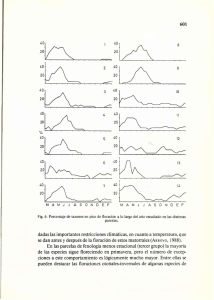
posteriores seria deseable cuantificar este esfuerzo, así como el de
Anuncio

posteriores seria deseable cuantificar este esfuerzo, así como el de los taxones nectaríferos (producción total de flores y cantidad de polen y/o néctar por flor). La familia Ericaceae es principalmente nectarífera, pero está bien representada sólo en parcelas de clima más moderado y húmedo, en las que no se observó diferenciación fenológica en ningún atributo. Las parcelas de mayor altitud (Grazalema b y c) son las que muestran mayor número de atributos con diferenciación fenológica. No hay ninguna razón para pensar que la altitud implique directa o indirectamente presiones selectivas que determinen esta organización de la floración. Más bien parece resultado de una representación muy particular de las familias, es decir, el componente filogenético de tal organización debe ser fuerte. De hecho KOCHMER & HANDEL (1986) demuestran en dos floras templadas no relacionadas de 2.298 y 1.575 taxones respectivamente, la existencia de diferencias fenológicas claras a nivel de familia, demostrando un papel muy importante de la filogenia en la floración. Esta idea ya había sido adelantada por CLARKE (1893). En cuanto a las características vegetativas, los taxones arbustivos están sobrerrepresentados en el presente estudio, haciendo estadísticamente difíciles las comparaciones fenológicas. No obstante, se ha encontrado en dos parcelas una floración significativamente precoz de los taxones arbóreos o de tamaño grande, debido probablemente a su mayor capacidad para almacenar reservas (GRAINGER, 1939; KOCHMER & HANDEL, 1986). Muchos de estos árboles son de floración histerántea y anemófilos (véase más arriba) o entomófilos. En este último caso la floración precoz e histerántea puede suponer una atracción visual mayor para los polinizadores. En este estudio se ha calculado el solapamiento de la floración de todos los pares de taxones de cada parcela, para hacer una aproximación a las posibles interacciones que los taxones que conviven pueden sufrir en su polinización. La idea de que las plantas zoófilas pueden competir por los polinizadores es antigua (ROBERTSON, 1895, 1924). Se ha propuesto que esta competencia puede ser responsable de la divergencia de hábitats, de la utilización de diferentes polinizadores o de la separación fenológica de los taxones que conviven (e.g. HEINRICH, 1975; PLEASANTS, 1983; WASER, 1983. Sólo el último fenómeno interesa aquí. Sin embargo, ha sido recientemente cuando se ha iniciado el estudio experimental de estos problemas, observándose casos tanto de competencia como de mutualismo (véanse las revisiones de KA Y, 1987; RATHCKE, 1983 y WASER, 1983). Este mutualismo ha sido indicado para la floración (WASER & REAL, 1979) y también para las semejanzas florales (mimetismo: DAFNI, 1984; LITTLE, 1983).