cromosomas eucariontes
Anuncio

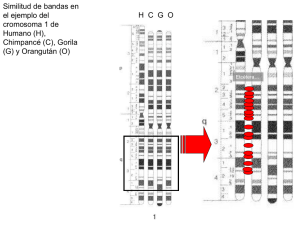
Estructura de los cromosomas eucariontes Se requieren grandes cantidades de DNA para codificar toda la información necesaria para producir solo una bacteria unicelular, y se necesita mucho más DNA para codificar las instrucciones que dirigen el desarrollo de los organismos multicelulares, como el del ser humano. Cada célula humana contiene cerca de 2 m de DNA, aunque el núcleo celular tiene solamente un diámetro de 5 a 8 µm. Intentar introducir todo este material en un espacio tan pequeño es equivalente a tratar de plegar 40 Km. de hebras extremadamente finas en una pelota de tenis. En las células eucariontes, las moléculas bicatenarias enormemente largas están contenidas en los cromosomas, que no solo ocupan cómodamente el interior del núcleo, sino que pueden ser distribuidos con facilidad entre las dos células hijas en cada división celular. Como se verá en esta sección, la tarea compleja de compactar el DNA es llevada a cabo por proteínas especializadas que se unen a éste y lo pliega, lo cual genera una serie de enrollamientos y asas que proporcionan niveles cada vez más altos de organización y que evitan que el DNA se transforme en una maraña difícil de controlar. Asombrosamente, el DNA está compactado de un modo tan ordenado que puede llegar a ser accesible a todas las enzimas y a otras proteínas que lo replican, lo reparan y utilizan sus genes para producir proteínas. Las bacterias típicamente transportan sus genes en una única molécula de DNA circular. Este DNA también se asocia con proteínas que lo condensan, pero que difieren de las proteínas que hacen lo propio en tas células eucariontes. Aunque es denominado con frecuencia un "cromosoma" bacteriano, este DNA procarionte no tiene la misma estructura que la de los cromosomas eucariontes, y se sabe menos acerca de cómo se condensan. Nuestra explicación de la estructura cromosómica estará centrada enteramente en los cromosomas eucariontes. El DNA eucarionte está condensado en los cromosomas En los eucariontes, como el ser humano, el DNA se distribuye en un grupo de diferentes cromosomas en el núcleo. El genoma humano, por ejemplo, contiene aproximadamente 3,2 x 109 nucleótidos distribuidos en 24 cromosomas. Cada cromosoma consiste de una única molécula de DNA lineal muy larga asociada a proteínas que pliegan la fina hebra de DNA en una estructura más compacta. El complejo de DNA y proteína se denomina cromatina (del griego chroma, "color," 1 a causa de sus propiedades de tinción). Además de las proteínas involucradas en la condensación del DNA, los cromosomas también se asocian con otras proteínas que intervienen en la expresión génica, la replicación del DNA y reparación de éste. Con la excepción de las células germinales (espermatozoides y óvulos) y de las células muy especializadas que carecen de la totalidad del DNA (como los glóbulos rojos sanguíneos), cada célula humana contiene dos copias de cada cromosoma, una heredada de la madre y otra del padre; los cromosomas maternos y paternos de un par se denominan cromosomas homólogos (homólogos). El único par de cromosomas no homólogo es el par sexual en los hombres, donde un cromosoma Y es heredado del padre y un cromosoma X de la madre. Los cromosomas humanos pueden individualizarse por hibridación del DNA (técnica que se describe en detalle en cap. 10); la técnica utiliza un grupo de moléculas de DNA unidas a moléculas fluorescentes para "pintar" cada cromosoma de un color distinto (fig. 5-12). Fig. 5-12. Cada cromosoma humano puede "pintarse" con un color diferente para permitir su identificación precisa con el microscopio óptico. Los cromosomas de un hombre fueron aislados de una célula en división nuclear (mitosis) y por ello se encontraban en un estado altamente compacto. El color de los cromosomas se obtiene mediante su exposición a un conjunto de moléculas de DNA humano que se acoplaron a una combinación de marcadores fluorescentes. Por ejemplo, las moléculas de DNA derivadas del cromosoma 1 están marcadas con una combinación específica, las del cromosoma 2 con otra y así sucesivamente. Como el DNA marcado puede formar apareamiento de bases, o hibridar solamente a su cromosoma de origen (véase el cap. 10), cada cromosoma está marcado de modo diferente. Para estos experimentos, los cromosomas se someten a tratamientos que separan la doble hélice de DNA en cadenas individuales, lo cual permite e) apareamiento de bases con el DNA marcado de cadena simple, al tiempo que se mantiene la estructura cromosómica relativamente intacta. (A) Los cromosomas se visualizan en su distribución original a partir del lisado celular- (B) Los mismos cromosomas alineados artificialmente en orden. En este cariotipo, los cromosomas homólogos están numerados y dispuestos en pares: la presencia del cromosoma Y indica que el DNA fue aislado de un hombre. (De E. Schróck y col., Soence 273:494-497, 1996.QAAAS.» 2 Pero el modo más tradicional para distinguir un cromosoma de otro es teñirlos con marcadores que se unen a determinados tipos de secuencias de DNA. Estos marcadores diferencian principalmente entre el DNA que es rico en los pares de nucleótidos A-T y el DNA que es rico en G-C, y producen un sorprendente y confiable patrón de bandas a lo largo de cada cromosoma (fig. 5-13). La disposición en bandas en cada tipo de cromosomas es única y permite identificar y numerar cada cromosoma. Fig. 5-13. Los patrones de bandeo único permiten la identificación de cada cromosoma humano. Los cromosomas 1 a 22 son enumerados en el orden aproximado en relación con su tamaño. Una célula somática (es decir, no germinal) humana típica contiene dos de cada uno de estos cromosomas más dos cromosomas sexuales: dos cromosomas X en la mujer y un cromosoma X y uno Y en el hombre. Los cromosomas utilizados para hacer estos mapas se tiñeron en un estadio temprano de la mitosis, cuando el DNA está compactado. La línea horizontal representa la posición del centrómero, que aparece como una constricción en los cromosomas mitóticos; las prominencias en los cromosomas 13,14,15.21 y 22 indican las posiciones de los genes que codifican para los RNA ribosómicos de mayor tamaño (véase cap. 7). Estos patrones son obtenidos mediante tinción de los cromosomas con el método de Giemsa, que produce bandas oscuras en regiones ricas en los pares de nucleótidos A-T. (Adaptado de U. Franke, Cytogenet. Ce// Genet-31:24-32, 1981.) Una muestra del grupo completo de 46 cromosomas se denomina cariotipo humano. Si se pierden partes de un cromosoma, o se intercambian entre los cromosomas, estos cambios pueden detectarse por las modificaciones en el patrón de las bandas. Los citogenetistas utilizan las alteraciones en el patrón de las bandas para detectar anomalías cromosómicas que se asocian con algunos defectos heredados (fig. 5-14) y con ciertos tipos de cáncer. 3 Fig. 5-14. Los cromosomas anormales se asocian con algunos defectos genéticos hereditarios. (A) Un par de cromosomas de un paciente con ataxia hereditaria, enfermedad caracterizada por el deterioro progresivo de las habilidades motoras. Los pacientes tienen un cromosoma 12 normal (izquierda) y un cromosoma 12 aberrante, como se aprecia por su mayor longitud. A partir de este patrón de bandas el material adicional contenido en el cromosoma 12 aberrante se dedujo que era una pieza del cromosoma 4 que se había adherido de forma inadecuada al cromosoma 12, (B) Los cromosomas se muestran aquí con el segmento que corresponde al DNA del cromosoma 4 de color azul y las partes que corresponden al DNA del cromosoma 12 de color rojo. (De E, Schróck y col., Science 273:294-497,1996. ©AAAS.) Los cromosomas contienen largas hileras de genes La función más importante de los cromosomas es transportar a los genes, que son las unidades funcionales de la herencia (fig. 5-15). Un gen se define en general como un segmento de DNA que contiene las instrucciones para producir una proteína particular (o, en algunos casos, una grupo de proteínas estrechamente relacionadas). Aunque esta definición se ajusta a la mayor parte de los genes, algunos dirigen la producción de una molécula de RNA, en lugar de una proteína, como su producto final. Al igual que las proteínas, las moléculas de RNA llevan a cabo diversas funciones catalíticas y estructurales en la célula, según se describirá en capítulos posteriores. Como puede preverse, existe una correlación entre la complejidad de un organismo y el número de genes en su genoma. Por ejemplo, el número total de genes es menor de 500 para una bacteria simple y cerca de 30 000 en el hombre. Las bacterias y algunos eucariontes unicelulares tienen genomas especialmente compactos: las moléculas de DNA que forman parte de sus cromosomas son poco más que filamentos o genes estrechamente agrupados. Sin embargo, los cromosomas de muchos eucariontes (incluidos los humanos) contienen, además de los genes, gran cantidad de DNA intercalado que no parece transportar información crítica. A veces se lo denomina DNA "basura", en la medida que su utilidad para la célula todavía no ha sido claramente demostrada. Aunque la secuencia de nucleótidos particulares de este DNA podría no ser importante, el DNA mismo -que actúa como material espaciador- podría ser significativo para la evolución a largo plazo de la especies y para la propia actividad de los genes. 4 Fig. 5-15. Se muestran los genes localizados en una porción del cromosoma 2 de la mosca de la fruta Drosophila melanogaster. Esta figura representa aproximadamente el 3% del total del genoma de la Drosophila, dispuesto como seis segmentos contiguos. Como se resume en tas claves, las representaciones simbólicas son barras horizontales con los colores del arco iris para el contenido de pares de bases de G-C; barras verticales coloreadas, los genes (conocidos y predecidos) codificados en una de las cadenas del DNA (por arriba de la línea media) y los genes codificados en la otra cadena (barras por debajo de la línea media). Como se indica en la clave, la altura de la barra de cada gen indica la certeza con la cual representa a un gen verdadero: la altura de confianza. El color de la barra de cada gen (véase código de color en la clave) señala si un gen íntimamente relacionado se halla en otros organismos. Por ejemplo, MWY significa que el gen tiene correspondencia estrecha en mamíferos, en el gusano nematodo Caenorhabditis elegans y en la levadura Saccharomyces cerevisiae. MW denota que se encuentra en mamíferos y el gusano, pero no en la levadura. (De Mark D. Adams y col., Science 287:2185-2195. 2000. © AAAS.) 5 En general, cuanto más complejo es un organismo, más grande es su genoma. Pero esta relación no siempre se cumple. El genoma humano, por ejemplo, es unas 200 veces más grande que el de la levadura S. cerevisiae, pero unas 30 veces más pequeño que el de algunos vegetales y 200 veces más pequeño que el de algunas especies de amebas. Además, también difiere de una especie a la otra en cómo se distribuye el DNA en los cromosomas. Los seres humanos tienen 46 cromosomas, pero una especie de ciervos pequeños tiene solamente 6, mientras que ciertas especies de carpas tienen más de 100. Incluso especies estrechamente relacionadas con tamaños de genomas similares pueden tener cromosomas con un número y tamaño distintos (fig. 5-16). Por lo tanto, aunque el número de genes está casi correlacionado con la complejidad de las especies, no mantiene una relación simple con el número de cromosomas y el tamaño total del genoma. Los genomas y los cromosomas de las especies actuales fueron modelados por una serie de acontecimientos genéticos aparentemente al azar en respuesta a una presión de selección. Fig. 5-16. Las especies estrechamente relacionadas pueden tener un número de cromosomas muy diferente. En la evolución del ciervo mutiaco de la India, los cromosomas que se separaron al inicio se fusionaron sin tener un efecto significativo sobre el animal. Las dos especies muestran tener aproximadamente el mismo número de genes. (Adaptado de M.W. Strickberger, Evolution, 3ª ed. Sudbury. MA; Jones & Bartlett Publishers, 2000.) Pregunta 5-2 En una doble hélice de DNA, los pares de nucleótidos adyacentes están a 0,34 nm de distancia. Utilice la figura 5-13 para estimar la longitud del DNA en el cromosoma humano 1 si éste estuviera desenrollado y estirado. Si la longitud real del cromosoma 1 en este estadio de la mitosis es aproximadamente de 10 µm, ¿cuál sería el grado de compactación del DNA? 6 Los cromosomas existen en diferentes estados durante la vida de la célula Para formar un cromosoma funcional, una molécula de DNA debe hacer algo más que transportar genes: necesita replicarse y las copias deben separarse y distribuirse con exactitud entre las células hijas en cada división celular. Estos procesos se producen a través de una serie ordenada de estadios, conocidos en conjunto como ciclo celular. Este ciclo de crecimiento celular se resume brevemente en la figura 5-17 y se examina en detalle en el capítulo 19. Solamente dos de estos estadios requieren nuestro interés en este capitulo: la interfase, cuando los cromosomas se duplican; y la mitosis, cuando se distribuyen en los núcleos de las dos células hijas. Fig. 5-17. La replicación y la segregación de los cromosomas tienen lugar a través de una serie de estadios, denominados ciclo celular. Durante la interfase, la célula expresa activamente sus genes. Todavía durante la interfase y antes de la división celular, el DNA es replicado y los cromosomas se duplican. Una vez que se completa la replicación del DNA, la célula puede entrar en la fase M, cuando se produce la mitosis. La mitosis es la división del núcleo. Durante este estadio, los cromosomas se condensan, la expresión génica cesa en su mayor parte, la envoltura nuclear se fragmenta y se forma el huso mitótico a partir de los microtúbulos y de otras proteínas. El huso mitótico captura los cromosomas condensados y lleva un grupo completo de cromosomas hasta cada extremo de la célula. Se forma una envoltura nuclear alrededor de cada grupo de cromosomas, y en la etapa final de la fase M, la célula se divide y forma dos células hijas. 7 Durante la interfase, los cromosomas están extendidos como hebras largas, finas y entrelazadas en el núcleo y no pueden distinguirse fácilmente con el microscopio óptico. Los cromosomas en este estado extendido se denominan como cromosomas en interfase. Las secuencias de DNA especializadas que se hallan en todos los eucariontes garantizan que los cromosomas en interfase se repliquen en forma enciente (Fig. 5-18). Un tipo secuencia de nucleótidos actúa como el origen de la replicación, la localización en la cual comienza la duplicación del DNA, como se verá en el capítulo 6. Los cromosomas eucariontes contienen muchos orígenes de replicación para asegurar que la totalidad del cromosoma pueda replicarse rápidamente. Otra secuencia de DNA forma los telómeros, que se hallan en cada uno de los dos extremos del cromosoma. Los telómeros contienen secuencias de nucleótidos repetidas que permiten que los extremos de los cromosomas se repliquen. También protegen al extremo del cromosoma de ser confundido por la célula con una molécula de DNA rota que necesita de reparación. La replicación y la función de los telómeros se analizan en capítulos posteriores. Fig. 5-18. Tres elementos de la secuencia de DNA son necesarios para producir un cromosoma eucarionte que pueda replicarse y luego segregarse en la mitosis. Cada cromosoma tiene múltiples orígenes de replicación, un centrómero y dos telómeros. Se muestra esquemáticamente la secuencia de procesos que experimenta un cromosoma típico durante el ciclo celular- En la interfase comienza la replicación del DNA en los orígenes de replicación y avanza bidireccionalmente a lo largo del cromosoma. En la fase M, el centrómero une a los cromosomas duplicados en el huso mitótico de modo que durante la mitosis será distribuida una copia a cada célula hija. Los centrómeros también contribuyen a mantener juntos a los cromosomas duplicados hasta que están listos para su separación. Los telómeros forman capuchones especiales en cada extremo del cromosoma. 8 A medida que avanza el ciclo celular, el DNA se enrolla y adopta una estructura cada vez más compacta, hasta que se forman cromosomas mitóticos muy condensados. Esta es la forma en la cual se visualizan con mayor facilidad; en efecto, todas las imágenes de los cromosomas mostradas hasta ahora en el capítulo son de cromosomas mitóticos. En este estado condensado, los cromosomas duplicados pueden separarse fácilmente cuando la célula se divide. Es la presencia de una tercera secuencia de DNA especializada, el centrómero, la que permite que una copia de cada cromosoma duplicado se distribuya en cada célula hija (véase fig. 5-17). Los cromosomas en interfase están organizados dentro del núcleo A pesar de que los cromosomas de las células en interfase son mucho más largos y finos que los cromosomas mitóticos, se piensa que están bien organizados dentro del núcleo. El núcleo se halla delimitado por una envoltura nuclear formada por dos membranas concéntricas. La envoltura nuclear está sostenida por dos redes de filamentos proteicos (véase cap. 17): una, denominada la lámina nuclear, forma una capa delgada subyacente que sostiene a la membrana nuclear interna; mientras que la otra, con una organización menos regular, rodea a la membrana nuclear externa (fig. 5-19). Las dos membranas están horadadas a intervalos por los poros nucleares, que transportan en forma activa ciertas moléculas hacia el citosol y desde éste. El interior del núcleo no es una mezcla al azar de muchos componentes de DNA, RNA y proteínas. Cada cromosoma en interfase probablemente ocupa una región particular del núcleo de modo que diferentes cromosomas no se llegan a enredar entre sí. Se piensa que esta organización se alcanza al menos en parte mediante la adherencia de partes de los cromosomas a sitios de la envoltura nuclear o de la lámina nuclear. El ejemplo más obvio de la organización de los cromosomas en el núcleo en interfase es el nucléolo, la estructura más destacada en ese estado con el microscopio óptico. Esta es una región donde las partes de los diferentes cromosomas que transportan genes para el RNA ribosómico están agrupados entre sí (véase figs. 5-13 y 5-19). Aquí, los RNA ribosómicos se sintetizan y combinan con proteínas para formar ribosomas, la maquinaria celular que sintetiza proteínas (véase cap. 7). 9 Fig. 5-19. Los cromosomas interfásicos están organizados dentro del núcleo. (A) Micrografía electrónica de una sección delgada a través del núcleo de un fibroblasto humano, El núcleo está rodeado por la envoltura nuclear, que consta de una membrana doble, perforada por poros nucleares. Dentro del núcleo, la cromatina aparece como una masa abigarrada difusa, con regiones cromosómicas especialmente densas, denominadas heterocromatina (tinción oscura), que se localizan sobre todo alrededor de la periferia, inmediatamente por debajo de la envoltura nuclear. La región oscura de gran tamaño es el nucléolo. (B) Representación esquemática de la sección transversal de un núcleo celular típico. La envoltura nuclear consiste en dos membranas, de las cuales la externa se continúa con el retículo endoplasmático. Dos redes de filamentos de citoesqueleto (verde) proporcionan sostén mecánico a la envoltura nuclear: los del interior del núcleo forman la lámina nuclear, que reviste la cara interna de la membrana nuclear Interna. El nucléolo (gris) es el sitio de la síntesis de RNA ribosómico. (A. cortesía de E. G. Jordán y J. Mc Govern.) El DNA en los cromosomas se halla muy condensado Como se ha visto, en todas las células eucariontes, estén en interfase o en mitosis, el DNA está condensado en los cromosomas. El cromosoma 22 humano, por ejemplo, presenta cerca de 48 millones de pares de nucleótidos; estirado de un extremo al otro, su DNA podría extenderse cerca de 1,5cm. Sin embargo, durante la mitosis, el cromosoma 22 mide solamente cerca de 2 µm de longitud, lo cual indica que es casi 10 000 veces más compacto que su forma extendida. Este fenómeno de condensación lo llevan a cabo proteínas que enrollan y pliegan el DNA en niveles cada vez más altos de organización. El DNA de los cromosomas en interfase, aunque menos condensado que el de los cromosomas mitóticos (fig. 5-20), todavía está muy condensado. en una proporción de cerca de 1000 veces. 10 Fig. 5-20. El DNA en los cromosomas en interfase es menos compacto que el de los cromosomas mitóticos. (A) Micrografía electrónica que muestra una enorme maraña de DNA cromosómico que se vuelca de un núcleo en interfase lisado. (B) Micrografía electrónica de barrido de un cromosoma mitótico; el cromosoma se ha duplicado, pero los dos cromosomas nuevos todavía están unidos entre si (véase fig. 5-18). La región de la constricción indica la posición del centrómero. Obsérvese la diferencia en las escalas. (A, cortesía de Victoria Foe; B, cortesía de Terry D. Alien.) En las siguientes secciones se considerarán las proteínas especializadas que hacen posible esta compresión. Téngase en mente, sin embargo, que el cromosoma es una estructura dinámica. Los cromosomas no se condensan y relajan solamente en relación con el ciclo celular, sino que diferentes regiones de los cromosomas en interfase deben desplegarse para permitirle a las células el acceso a secuencias de DNA especificas para la replicación, reparación o expresión génica. La condensación de los cromosomas, por lo tanto, debe ser lo bastante flexible como para hacer posible un acceso rápido y localizado al DNA según la necesidad. Los nucleosomas son las unidades básicas de la estructura de la cromatina Las proteínas que se unen al DNA para formar los cromosomas eucariontes tradicionalmente se dividen en dos clases generales: histonas y proteínas cromosómicas no histónicas. Las histonas están presentes en enormes cantidades (cerca de 60 millones de moléculas de varios tipos diferentes en cada célula), y su masa total en los cromosomas es casi igual a la del DNA mismo. El complejo que forman ambas clases de proteínas con el DNA nuclear se denomina cromatina. 11 Las histonas son responsables del primero y más esencial nivel de condensación de la cromatina, el nucleosoma, descubierto en 1974. Cuando los núcleos en interfase se fragmentan cuidadosamente y su contenido se examina con el microscopio electrónico, la mayor parte de la cromatina tiene la forma de una hebra de un diámetro cercano a 30 nm (fig. 5-21A). Si se somete esta cromatina a tratamientos que la descondensan parcialmente, puede observarse como una serie de "perlas o cuentas en un collar" (fig. 5-21B). El collar es el DNA, y cada cuenta es el núcleo de una partícula nucleosómica que consiste en DNA enrollado alrededor de un núcleo de proteínas formado por histonas. Fig. 5-21- Los nucleosomas pueden ser vistos con el microscopio electrónico. (A) La cromatina aislada directamente de un núcleo en Interfase aparece en el microscopio electrónico como una hebra de 30 nm de grosor. (B) Esta micrografía electrónica muestra una parte de la cromatina que se descondensó en forma experimental, después del aislamiento para mostrar los nucleosomas, (A, cortesía de Barbara Hamkalo; B, cortesía de Victoria Foe.) La estructura de los nucleosomas se determinó después de aislarlos por primera vez a partir de la cromatina descondensada mediante digestión con enzimas específicas (denominadas nucleasas) que fragmentan el DNA al cortarlo entre los nucleótidos. Tras un breve periodo de digestión, el DNA expuesto entre los núcleos de las partículas nucleosómicas, el DNA espaciador, se degrada. Cada partícula nucleosómica está constituida por un complejo de ocho histonas -dos moléculas de cada histona H2A, H2B, H3 y H4, y una cadena doble de DNA de cerca de 146 pares de nucleótidos que se enrollan alrededor de este octámero de histonas (fig. 5-22). La estructura de alta resolución del núcleo de una partícula nucleosómica fue resuelta en 1997 y reveló en el detalle atómico el complejo de histonas con forma de disco alrededor del cual el DNA esta muy enrollado y da 1,65 vueltas que giran hacia la izquierda (fig. 5-23). 12 Fig. 5-22. Los nucleosomas contienen DNA enrollado alrededor de un núcleo proteico de ocho moléculas de histonas. El núcleo de la partícula nucleosómica se libera de la cromatina mediante digestión del DNA espaciador con una nucleasa, enzima que fragmenta el DNA, (la nucleasa puede degradar al DNA expuesto, pero no puede atacar al DNA unido fuertemente alrededor del núcleo del nucleosoma). Después de la disociación de los nucleosomas aislados en su núcleo de proteínas y DNA, puede determinarse la longitud del DNA que estaba enrollado alrededor del núcleo proteico. Su longitud de 146 pares de nucleótidos es suficiente para enrollarse casi dos veces alrededor del núcleo de histonas. Fig. 5-23. La estructura del núcleo de la partícula nucleosómica, determinada por el análisis de difracción de rayos X, revela cómo el DNA está enrollado apretadamente alrededor de un núcleo de histona de forma discoidal. Cada histona se colorea de acuerdo con el esquema que se muestra la figura 5-22; la hélice de DNA es gris. Se puede ver una parte de la cola de la histona H3 extendiéndose desde el nucleosoma, pero las colas de las otras histonas no se muestran, (Reproducido con autorización de K. Luger y col. .Nature 389:251-260. 1997. © Macmillan Magazines Ltd.) 13 Cada núcleo de una partícula nucleosómica se separa del siguiente mediante una región de DNA espaciador, que puede variar en longitud desde unos pocos pares de nucleótidos hasta cerca de 80. (el término "nucleosoma" técnicamente se refiere a un núcleo de una partícula nucleosómica más uno de sus DNA espaciadores adyacentes, pero a menudo se lo utiliza como sinónimo de "núcleo de una partícula nucleosómica"). La formación de los nucleosomas convierte a la molécula de DNA en una hebra de cromatina aproximadamente de un tercio de su longitud inicial y proporciona el primer nivel de condensación del DNA. Las cuatro histonas que forman el núcleo del nucleosoma son proteínas relativamente pequeñas con una alta proporción de aminoácidos de carga positiva (lisina y arginina). Las cargas positivas ayudan a las histonas a unirse apretadamente al esqueleto azúcar-fosfato del DNA con carga negativa. Estas interacciones numerosas explican en parte por qué el DNA prácticamente de cualquier secuencia puede unirse a un núcleo de histonas. Cada uno de estos núcleos de histonas también tiene una larga "cola" del aminoácido N-terminal que se extiende hacia fuera desde su ADN. Las colas de histonas están sometidas a varios tipos de modificaciones covalentes que controlan muchos aspectos de la estructura de la cromatina, como se verá en breve. Las histonas que forman el núcleo del nucleosoma se hallan entre las más conservadas de todas las proteínas eucariontes conocidas; por ejemplo, hay solo dos diferencias entre las secuencias de aminoácidos de la histona H4 de los guisantes y de las vacas. Recientemente, se descubrieron histonas en Archaea, procariontes que forman un reino filogenético distinto del de los eucariontes y las eubacterias (véase cap. 1). Este extremo de conservación evolutiva refleja el papel vital de las histonas en la formación de la cromatina. 14 Los cromosomas tienen varios niveles de condensación del DNA Aunque se forman largas cadenas de nucleosomas sobre la mayor parte del DNA cromosómico, la cromatina en las células vivas adopta en raras ocasiones la forma extendida en collar de perlas ilustrada en la figura 5-21B. En lugar de ello, los nucleosomas se reúnen luego uno sobre el otro para generar una estructura más compacta, la fibra de 30 nm (véase fig. 5-21A). La compactación de los nucleosomas en la fibra de 30 nm depende de una quinta histona denominada H1, que agruparía a los nucleosomas en una disposición regularmente repetida. La estructura que resulta se muestra en la figura 5-24 como parte de un esquema más grande de varios niveles de condensación del cromosoma. Se han propuesto distintos modelos para explicar de qué manera los nucleosomas son contenidos en la fibra de cromatina de 30 nm; uno de los más concordantes con los datos disponibles es una serie de variaciones conocidas en conjunto como el modelo en zigzag (fig. 5-25). Es probable que la estructura de 30 nm hallada en los cromosomas sea un mosaico fluido de diferentes variaciones de zigzag. Fig. 5-24. La condensación de la cromatina se produce a varios niveles. Este dibujo esquemático muestra algunos de los órdenes de la cromatina que darían origen al cromosoma mitótico muy condensado. 15 Fig. 5-25. Las fibras de cromatina pueden ser condensadas de acuerdo con un modelo en zigzag. (A y B) Microscopía electrónica que sustenta las estructuras de los modelos que se muestran arriba y abajo a la izquierda en (C). La estructura de la fibra de cromatina de 30 nm puede ser una combinación de estas variaciones de zigzag. Puede producirse una interconversión entre las tres variantes mediante la expansión y la contracción en acordeón de la fibra. Obsérvese que los núcleos de histona no figuran en los esquemas en (C). (De J. Vendar y col., Proc. Nati. Acad. Sci. U.S.A. 95:1417314178,1998. @ National Academy of Sciences.) Se sabe que la fibra de cromatina de 30 nm puede condenarse todavía más. Se vio al comienzo de este capítulo que durante la mitosis la cromatina llega a estar tan condensada que los cromosomas pueden verse con el microscopio óptico. ¿Cómo se pliega la fibra de 30 nm para producir los cromosomas mitóticos? La respuesta a esta pregunta todavía no se conoce en detalle, pero se piensa que la fibra de 30 nm se organiza en bucles que proceden de un eje central (figs. 524 y 5-26). Finalmente, este filamento de bucles experimenta al menos un nivel más de condensación para formar el cromosoma mitótico (véase fig. 5-24). Fig. 5-26. Los cromosomas mitóticos se forman a partir de cromatina muy condensada. Esta micrografía electrónica de barrido muestra una región próxima al extremo de un cromosoma mitótico típico. Cada proyección observada como una prominencia se piensa que representa el extremo de un bucle de cromatina separado. El cromosoma en esta foto se duplicó, pero los dos nuevos cromosomas (también denominados cromátidas) todavía se mantienen unidos (véase fig. 5-20B). Los extremos de los dos cromosomas pueden distinguirse fácilmente en esta micrografía- (De M.P, Marsden y U.K. Laemmii. Ce// 17:849-858,1989. ©Elsevier.» 16 Los cromosomas en inferfase contienen tanto formas de cromatina condensada como de cromatina más extendida Cuando las células hijas completan su separación después de la mitosis, los cromosomas mitóticos se descondensan y se despliegan, son los cromosomas de interfase (véase fig. 5-18). Sin embargo, la cromatina en un cromosoma en interfase no está en el mismo estado de condensación en todas las partes del cromosoma. En general, las regiones del cromosoma que contienen genes que se expresan se hallan más desplegadas, mientras que las que contienen genes inactivos son más compactas. Por lo tanto, la estructura detallada de un cromosoma en interfase puede diferir de un tipo celular a otro, según qué genes se expresen. La forma de mayor grado de condensación de la cromatina en interfase se denomina heterocromatina (del griego heteros, que significa "diferente," más cromatina). Se observó primero con el microscopio óptico en la década de 1930 como regiones fuertemente teñidas dentro de la masa de cromatina. La heterocromatina típicamente forma cerca del 10 % del cromosoma en interfase. y en los cromosomas de mamíferos se concentra de manera característica alrededor de la región del centrómero y en los telómeros de los extremos de los cromosomas. La mayor parte del DNA que se condensa en heterocromatina no contiene genes. Sin embargo, los genes que llegan a ser contenidos en la heterocromatina son en general resistentes a la expresión porque la heterocromatina es extraordinariamente compacta (fig. 5-27). El resto de la cromatina en interfase, que está en una variedad de estados más extendidos, se denomina eucromatina (del griego eu, que significa "verdadero" o "normal," más cromatina). Todavía no se comprende cuál de estos niveles de condensación de los cromosomas mostrados en la figura 5-24 describen mejor a la eucromatina y a la heterocromatina; es probable que ambas presenten mezclas de varios estados de condensación. 17 Fig. 5-27. La expresión de un gen puede alterarse por el desplazamiento a otra localización en el genoma. Se muestran dos ejemplos de efectos de posición, en los que la actividad de un gen depende de su posición a lo largo del cromosoma. (A) El gen ADE2 de la levadura en su localización normal se expresa en todas la células. Cuando se lo sitúa cerca del extremo del cromosoma de una levadura, que se pliega en una forma particularmente compacta de cromatina, el gen no se expresa más en la mayor parte de las células de la población, ADE2 codifica una de las enzimas de biosíntesis de adenina, y la ausencia del producto del gen ADE2 lleva a la acumulación de un pigmento rojo. Por tal razón, una colonia de células que expresa ADE2 es blanca, y una constituida por células donde el gen ADE2 no se expresa es roja. Los sectores blancos alrededor de la colonia roja representan las células donde el gen ADE2 se ha vuelto espontáneamente activo. Esto es el resultado de un cambio heredable en el estado de condensación de la cromatina cerca del gen ADE2 en estas células, (B) Los efectos de posición también pueden observarse para el gen white en la mosca de la fruta Drosophila. El gen white controla la producción de pigmento ocular y su nombre surge después de la mutación que lo identificó por primera vez. Las moscas de tipo salvaje con un + gen white normal (white ) tienen una producción normal de pigmento, que les da a aquellas los ojos rojos, pero si el gen white es mutado e inactivado, en las moscas mutantes (white ) no se forma pigmento y tienen los ojos blancos. En las moscas + en las que se desplazó un gen white normal cerca de una reglón de heterocromatina. los ojos son moteados con parches rojos y blancos. Los parches blancos representan a las células + donde el gen white es silenciado por los efectos de la heterocromatina y los parches rojos representan a las células + que expresan al gen white . Las motas se producen porque el + silenciamiento del gen white por la heterocromatina no es completo. Como para las levaduras, la presencia de parches de gran tamaño de células rojas y blancas Indica que el estado del gen es heredado (activo o silenciado). 18 Aunque la mayor parte de los cromosomas eucariontes contienen regiones de eucromatina y de heterocromatina, existen algunas excepciones importantes. Tal vez el ejemplo más notable se halle en los cromosomas X en interfase de las hembras de mamíferos. Las células femeninas contienen dos cromosomas X, mientras que las células masculinas contienen un X y un Y. Como una dosis doble de cromosomas X sería letal, las hembras de los mamíferos desarrollaron un mecanismo para inactivar permanentemente a uno de los dos cromosomas X en cada célula: al azar, uno u otro de los dos cromosomas X en cada célula llega a estar muy condensado en heterocromatina en el desarrollo embrionario temprano. Después de esto, en todas de las numerosas progenies de las células se heredan los estados condensado e inactivo del cromosoma X (fig. 5-28). Fig. 5-28. Un cromosoma X puede inactivarse por completo por la formación de heterocromatina. Las células en un embrión de mamífero hembra contienen dos cromosomas X, uno de la madre (X) y el otro del padre (X). En un estadio temprano del desarrollo, uno de estos dos cromosomas en cada célula se condensa en heterocromatina, aparentemente al azar. En cada división celular después de este estadio, el mismo cromosoma se vuelve condensado en todos los descendientes de la célula original. En ratones, la Inactivación del cromosoma X se produce entre el tercero y el sexto día de desarrollo. En los seres humanos, también tiene lugar la inactivación del X muy tempranamente en el desarrollo, antes de que las células hayan sido asignadas a una vía particular de desarrollo, Por lo tanto, las hembras terminan con un mosaico de células que llevan al cromosoma X paterno o materno inactivado. En la mayor parte de los tejidos y órganos, cerca de la mitad de las células será de un tipo y la mitad restante del otro. Pregunta 5-3 19 Las mutaciones en un gen particular del cromosoma X causan ceguera para el color. Todos los hombres portadores de un gen mutante son ciegos para el color- La mayor parte de las mujeres portadoras de un gen mutan-te tienen una visión adecuada para el color, pero ven imágenes en color con una resolución reducida, por lo cual se piensa que las células funcionales de los conos (las células que contienen a los fotorreceptores del color) se encuentran mucho más espaciadas entre sí que en la retina normal. ¿Podría dar una explicación convincente de esta observación? Si una mujer es ciega para el color, ¿qué podría decir acerca de su padre? ¿Acerca de su madre? Explique sus respuestas. Los cambios en la estructura del nucleosoma permiten el acceso al DNA Hasta ahora se analizó cómo el DNA se condensa cuidadosa y estrechamente en la cromatina; ahora se estudiará cómo esta condensación puede ser dinámica y permitir el rápido acceso al DNA subyacente. Se expresó que el DNA en las células transporta una enorme cantidad de información codificada, y es especialmente importante que las células tengan acceso a ella según la necesidad. Como se verá, el acceso directo al DNA es típicamente localizado y solo se exponen aquellas regiones del genoma que las células necesitan en un momento particular. Las células eucariontes tienen varias maneras de ajustar rápidamente la estructura local de su cromatina. Una forma utiliza la ventaja de los complejos de remodelación de la cromatina, máquinas proteicas que emplean la energía de la hidrólisis del ATP para cambiar la estructura de los nucleosomas (fig. 5-29). Estos complejos permiten que el DNA subyacente sea más accesible a otras proteínas en la célula, sobre todo las que participan en la replicación, reparación y expresión génica del DNA. Durante la mitosis, al menos algunos de los complejos de remodelación de la cromatina se inactivan, lo cual podría ayudar a los cromosomas mitóticos a mantener su estructura intensamente condensada. 20 Fig. 5-29. El complejo de remodelación de la cromatina altera la estructura del nucleosoma. De acuerdo con este modelo, distintos complejos que remodelan la cromatina alteran y vuelven a formar a los nucleosomas, aunque, en principio, el mismo complejo podría catalizar ambas reacciones. La proteína de unión al DNA podría estar involucrada en la expresión génica, la replicación del DNA o la reparación del DNA. Otro modo de obtener un cambio en la estructura de la cromatina depende de la modificación reversible de las colas de histonas. Las colas N-terminal de cada una de las cuatro proteínas del núcleo de histonas llevan a cabo funciones decisivas en la regulación de la estructura de la cromatina. Cada cola esta sujeta a varios tipos de modificaciones covalentes. que se agregan y eliminan después que los nucleosomas se ensamblan por la actividad de enzimas que residen en el núcleo. Aunque las modificaciones de las colas de histonas tienen poco efecto directo sobre la estabilidad de un nucleosoma, algunas parecen afectar directamente la estabilidad de la fibra de cromatina de 30 nm y de algunas de las estructuras de orden superior mencionadas antes. Sin embargo, los efectos más profundos de las colas de histonas modificadas parecen ser la capacidad para unirse y de este modo atraer a proteínas específicas a regiones de la cromatina. Diferentes patrones de modificaciones de la cola de histona atraen a diferentes proteínas; algunas 21 de estas proteínas generan condensaciones posteriores de la cromatina y otras facilitan el acceso al DNA. En conjunto, las diferentes combinaciones de modificaciones de la cola y los distintos grupos de proteínas que se unen a la histona pueden indicarle a la célula cosas diferentes: un patrón podría indicar que un tramo determinado de la cromatina ha sido recién replicado; otro que la expresión génica debería tener lugar (fig. 5-30). Fig. 5-30. El patrón de modificación de las colas de las histonas puede establecer cómo la célula trata un tramo de cromatina. (A) Cada histona puede modificarse por la unión covalente de diferentes moléculas. La histona H3, por ejemplo, puede recibir un grupo acetilo (Ac), un grupo metilo (Me) o un fosfato (P). Obsérvese que algunas posiciones (p. ej, lisina 9 y 27) pueden ser modificadas de más de un modo- (B) Diferentes combinaciones de las colas de histonas pueden constituir un tipo de "código de histona." De acuerdo con la hipótesis del código de histona, cada marca transporta un significado específico al tramo de cromatina en el que esto se produce. Solamente se conocen unos pocos significados de las modificaciones. Como es válido para los complejos de remodelación de la cromatina, las enzimas que modifican a las colas de histonas están estrechamente reguladas. Son conducidas a una región particular de la cromatina por otras señales, en especial por sus interacciones con las proteínas que se unen a secuencias específicas del DNA, un aspecto que se examinará en el capítulo 8. Es probable que estas enzimas que modifican histonas funcionen en concierto con el complejo que remodela la cromatina para condensar y relajar tramos de ésta y permitir que la estructura de la cromatina local cambie rápidamente de acuerdo con las necesidades de la célula. Tomado de ALBERTS. B., et alt, 2006, Introducción a la Biología Celular, 2ª edición, Editorial Médica Panamericana. 22