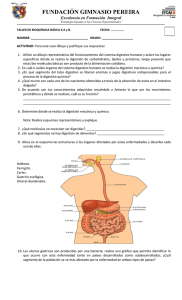
optimización de la digestión anaerobia de purines de
Anuncio

Universitat de Lleida Departament de Medi Ambient i Ciències del Sòl Laboratori d’Enginyeria Ambiental OPTIMIZACIÓN DE LA DIGESTIÓN ANAEROBIA DE PURINES DE CERDO MEDIANTE CODIGESTIÓN CON RESIDUOS ORGÁNICOS DE LA INDUSTRIA AGROALIMENTARIA ANTONIA ELENA CAMPOS POZUELO TESIS DOCTORAL UNIVERSITAT DE LLEIDA Escola tècnica superior d’Enginyeria Agrària Departament de Medi Ambient i Ciències del Sòl Laboratori d’Enginyeria Ambiental (LEA) OPTIMIZACIÓN DE LA DIGESTIÓN ANAEROBIA DE PURINES DE CERDO MEDIANTE CODIGESTIÓN CON RESIDUOS ORGÁNICOS DE LA INDUSTRIA AGROALIMENTARIA Memoria presentada por: Antonia Elena Campos Pozuelo Para optar al grado de Doctor Ingeniero Agrónomo por la Universitat de Lleida Director: Doctor Xavier Flotats i Ripoll Lleida, Julio de 2001 Tesis defendida el 27 de julio de 2001, en la Universidad de Lleida, ante el tribunal compuesto por: Dr. Iñaki Tejero Monzón Dr. Joan Mata Alvárez Dr. Jaime García de las Heras Dra. Núria Sala Martí Dra. Maria Rosa Teira Esmatges Obteniendo la calificación de Sobresaliente cum laudem por unanimidad Foto de portada: Imagen de la granja Mas El Cross (Santa Pau, Girona), que cuenta con un digestior anaerobio funcionando desde 1983. A mi madre, a la memoria de mi padre, a Chema, y a mi hija Elena. Agradecimientos Mi agradecimiento a todas aquellas personas e instituciones que de una u otra forma me han ayudado a llegar aquí, y de forma muy especial: Al profesor Dr. Xavier Flotats i Ripoll, director de este trabajo de tesis, por todas sus ideas, consejos, comentarios y ayuda tanto en la parte experimental como en la teórica, por las innumerables correcciones del documento de tesis, y en resumen por todo el apoyo que me ha dado, tanto en el ámbito científico como en el personal, durante todos estos años. A la CIRIT por la beca de formación de personal investigador que he disfrutado durante cuatro años, y a la Paeria de Lleida por la financiación del proyecto “Optimizació de la digestió anaeròbia termofílica de purins de porc”, básico para a realización del presente trabajo. A Jordi Palatsi y Antoni Casañé por su colaboración en la parte experimental, por todas las horas gastadas en el laboratorio, sus aportaciones y sugerencias, y por los buenos ratos que me han hecho pasar. A Mª Rosa Teira por sus ánimos y apoyo continuo, por sus consejos científicos y por las innumerables correcciones de inglés y catalán. A August, compañero de fatigas, por sus comentarios, sus ideas, su ayuda en el montaje experimental y su apoyo durante todo este tiempo. A los miembros del tribunal de tesis, Dr. Ignacio Tejero, Dr. Joan Mata, Dr. Jaime García de las Heras, Dra. Mª Rosa Teira y Dra. Nuria Sala, por todos sus comentarios, sugerencias y críticas constructivas, que han ayudado a la elaboración del presente documento. A todos los que durante estos años han pasado por el laboratorio, Lourdes, Patricia, Eduard, Silvia, Luci, Marta, Albert, Elisenda, David, Marina, Mercé, Vahida, Jordi,.... muchos de los cuales han colaborado en la parte experimental de este trabajo, y todos me han hecho pasar buenos momentos a lo largo de estos cinco años. A Rena Angelidaki por sus interesantes comentarios sobre la parte de modelización. A los propietarios y trabajadores de las granjas colaboradoras, Tribó, Ballester, Capdevila, Pinell y Cau y a Josep Millà, de la EDAR de Lleida, por su amabilidad. A todo el departamento, y de forma muy especial a Montse, por su ayuda en el laboratorio, a Marta por las lecturas de absorción y a Nuria, por su ayuda con electroforesis capilar; a los “precarios”, Asun, Francesc Domingo, Francesc Ferrer, Mikel, etc., por sus consejos, su apoyo y amistad; a Clara, por su ayuda en tareas administrativas; a Jaume Porta, por ponerme en contacto con Xavier; a Rosa Poch, Angela Bosch, Jordi Roca, y en general a todo el departamento, porque me han hecho sentirme bien durante todo este tiempo. A mis padres y a mis hermanos, por animarme a continuar, y hacerme sentir siempre que estaban ahí. Finalmente quiero agradecer a Chema todo su apoyo, y pedirle disculpas por todos esas horas, fines de semana y vacaciones que le he robado para dedicarlas a este trabajo, gracias por todo. ÍNDICE RESUMEN........................................................................................vii 1. INTRODUCCIÓN Y OBJETIVOS ............................................... 1 1.1. GENERACIÓN DE RESIDUOS ORGÁNICOS......................................................... 3 1.1.1. Residuos domésticos .......................................................................................... 3 1.1.2. Residuos industriales ........................................................................................ 6 1.1.3. Residuos ganaderos........................................................................................... 7 1.1.4. Gestión integrada de residuos .......................................................................... 10 1.2. BREVE REVISIÓN LEGISLATIVA ........................................................................ 11 1.3. ESTRATEGIAS DE TRATAMIENTO..................................................................... 13 1.4. CODIGESTIÓN DE RESIDUOS ORGÁNICOS ...................................................... 17 1.5. HERRAMIENTAS INFORMÁTICAS DE SIMULACIÓN DE SISTEMAS ANAEROBIOS .................................................................................................................. 18 1.6. OBJETIVOS GENERALES .................................................................................... 18 1.7. PLANIFICACIÓN DEL TRABAJO ......................................................................... 19 2. EL PROCESO DE LA DIGESTIÓN ANAEROBIA................... 21 2.1. HISTORIA DE LA TECNOLOGÍA ........................................................................ 23 2.2. PRODUCTOS FINALES DE LA DIGESTIÓN ANAEROBIA .................................. 24 2.3. CINÉTICA DE LAS REACCIONES BIOLÓGICAS ................................................. 26 2.3.1. Tasa de utilización de substrato (coeficientes de producción) .............................. 26 2.3.2. Tasa de crecimiento ........................................................................................ 26 2.3.3. Tasa específica de utilización de substrato........................................................ 29 2.3.4. Cinéticas de inhibición.................................................................................... 29 2.4. EL PROCESO MICROBIOLÓGICO Y BIOQUÍMICO DE LA DIGESTIÓN ANAEROBIA. ................................................................................................................... 31 2.4.1. Hidrólisis. ..................................................................................................... 32 2.4.2. Etapa fermentativa o acidogénica .................................................................... 36 2.4.3. Fase acetogénica. ............................................................................................ 42 2.4.4. Fase metanogénica.......................................................................................... 44 2.5. PARÁMETROS AMBIENTALES Y DE CONTROL ................................................. 47 2.5.1. Temperatura .................................................................................................. 47 2.5.2. Contenido de nutrientes .................................................................................. 52 2.5.3. Velocidad de carga orgánica (VCO) y tiempo de retención hidráulico (TRH)... 53 2.5.4. Agitación....................................................................................................... 53 2.5.5. pH y alcalinidad ............................................................................................ 54 2.5.6. Tóxicos e inhibidores ...................................................................................... 55 3. MÉTODOS ANALÍTICOS ..........................................................65 3.1. SÓLIDOS TOTALES Y VOLÁTILES ...................................................................... 67 ii Índice 3.2. DEMANDA QUÍMICA DE OXÍGENO (DQO) ................................................... 67 3.3. DEMANDA QUÍMICA DE OXÍGENO SOLUBLE (DQOSOL).............................. 68 3.4. NITRÓGENO KJELDHAL ................................................................................... 69 3.5. NITRÓGENO AMONIACAL ................................................................................. 70 3.6. NITRÓGENO ORGÁNICO ................................................................................... 70 3.7. AMONÍACO LIBRE .............................................................................................. 71 3.8. PH ........................................................................................................................ 71 3.9. ALCALINIDAD PARCIAL, TOTAL Y RELACIÓN DE ALCALINIDADES ............. 71 3.10. ÁCIDOS GRASOS VOLÁTILES .......................................................................... 72 3.11. COMPOSICIÓN DE BIOGÁS, CH4 Y CO2 ........................................................ 75 3.12. FÓSFORO TOTAL (PT) ...................................................................................... 77 3.13. CATIONES (NA, K, CA, MG, CU Y ZN)......................................................... 78 3.14. CONTENIDO EN GRASA. MÉTODO SOXLET................................................. 78 3.15. CONTENIDO EN FIBRA. MÉTODO FIBRA ÁCIDO DETERGENTE ................ 79 4. MEJORA DE UN MODELO ESTRUCTURADO DE DIGESTIÓN ANAEROBIA MEDIANTE LA INCORPORACIÓN DE LAS ECUACIONES PARA LA SIMULACIÓN DINÁMICA DEL PH DE LA TRANSFERENCIA LÍQUIDO-GAS ...............................79 4.1. RESUMEN ............................................................................................................ 81 4.2. INTRODUCCIÓN.................................................................................................. 81 4.2.1. La importancia del pH .................................................................................. 83 4.3. OBJETIVOS .......................................................................................................... 84 4.4. MODELIZACIÓN DEL PH EN SISTEMAS ANAEROBIOS. REVISIÓN BIBLIOGRÁFICA .............................................................................................................. 84 4.5. DESARROLLO DEL ALGORITMO DE SIMULACIÓN DINÁMICA DEL PH ........ 88 4.5.1. Notación........................................................................................................ 92 4.5.2. Establecimiento de las ecuaciones para la simulación dinámica del pH ............. 96 4.6. INCLUSIÓN DEL ALGORITMO DE SIMULACIÓN DEL PH EN UN MODELO DINÁMICO ESTRUCTURADO DE DIGESTIÓN ANAEROBIA PREVIO .......................... 98 4.6.1. Procesos y variables considerados ................................................................... 100 4.6.2. Cinética de los procesos biológicos considerados ............................................... 101 4.6.3. Establecimiento del sistema de ecuaciones diferenciales ordinarias.................... 106 4.6.4. Método numérico aplicado para la aproximación de las soluciones .................. 110 4.7. VALIDACIÓN DEL ALGORITMO DE SIMULACIÓN DINÁMICA DEL PH ....... 110 4.7.1. Establecimiento de las ecuaciones de simulación ............................................. 111 4.7.2. Dispositivo experimental para la validación del algoritmo .............................. 112 4.8. SIMULACIONES CON EL MODELO GENERAL DE DIGESTIÓN ANAEROBIA. ....................................................................................................................................... 115 4.9. CONCLUSIONES ................................................................................................ 120 Optimización de la digestión anaerobia de purines de cerdo iii 5. CODIGESTIÓN DE PURÍN CON RESIDUOS INDUSTRIALES. ENSAYOS DE VIABILIDAD EN DISCONTINUO. EFECTO DE LAS MEZCLAS. EFECTO DEL AMONIO..............................................................................................121 5.1. RESUMEN .......................................................................................................... 123 5.2. INTRODUCCIÓN................................................................................................ 123 5.2.1. Codigestión de residuos orgánicos................................................................... 123 5.2.2. Substratos posibles en el área de Lleida......................................................... 127 5.2.3. Metodología a utilizar para realizar estudios de viabilidad............................. 129 5.3. OBJETIVOS PARTICULARES DE LOS ENSAYOS EN DISCONTINUO. ............. 130 5.4. MATERIALES Y MÉTODOS .............................................................................. 131 5.4.1. Diseño del experimento................................................................................. 131 5.4.2. Métodos analíticos........................................................................................ 132 5.4.3. Desarrollo y seguimiento de los ensayos discontinuos ....................................... 132 5.4.4. Cálculo de la producción de gas acumulada.................................................... 133 5.4.5. Cálculo de índices de producción de metano .................................................... 134 5.4.6. Caracterización de los substratos utilizados ................................................... 138 5.4.7. Cálculo de las concentraciones finales de los parámetros físico-químicos............ 142 5.4.8. Análisis estadístico....................................................................................... 142 5.5. RESULTADOS Y DISCUSIÓN ............................................................................ 143 5.5.1. Experimento 1. Diluciones de purín a dos temperaturas: efecto del amonio ..... 143 5.5.2. Experimento 2. Codigestión de purín de cerdo con residuos de pera................. 167 5.5.3. Experimento 3. Codigestión de purín de cerdo con tierras decolorantes de aceite de oliva........................................................................................................................... 191 5.6. CONCLUSIONES ................................................................................................ 209 5.6.1. Sobre el método ............................................................................................ 209 5.6.2. Del experimento 1: Diluciones de purín - efecto del amonio. ........................... 210 5.6.3. Del experimento 2: Codigestión con residuo de pera. ...................................... 211 5.6.4. Del experimento 3. Adición de tierras decolorantes de aceite de oliva............... 212 5.6.5. Resumen de conclusiones ............................................................................... 212 6. ENSAYOS EN CONTINUO DE CODIGESTIÓN DE PURÍN Y TIERRAS DECOLORANTES DE ACEITE DE OLIVA. ............. 215 6.1. RESUMEN .......................................................................................................... 217 6.2. INTRODUCCIÓN................................................................................................ 218 6.2.1. Ensayos en régimen semicontinuo a escala laboratorio. ................................... 218 6.2.2. Importancia de la puesta en marcha. Riesgo de sobrecargas............................. 218 6.2.3. Digestión anaerobia de residuos ganaderos. .................................................... 220 6.2.4. Codigestión con tierras decolorantes de aceites vegetales.................................... 222 6.3. OBJETIVOS ........................................................................................................ 223 iv Índice 6.4. MATERIALES Y MÉTODOS. .............................................................................. 223 6.4.1. Dispositivo experimental .............................................................................. 223 6.4.2. Métodos analíticos........................................................................................ 229 6.4.3. Operación de los reactores ............................................................................. 229 6.4.4. Cálculo de parámetros .................................................................................. 229 6.4.5. Programación de los ensayos.......................................................................... 232 6.4.6. Caracterización de la alimentación ................................................................ 234 6.5. ESTRATEGIAS DE PUESTA EN MARCHA. RESULTADOS Y DISCUSIÓN. ....... 236 6.5.1. Reactores 1 y 2. Puesta en marcha directamente con purines de cerdo. ............. 237 6.5.2. Reactores 3 y 4. Puesta en marcha utilizando lodos de depuradora digeridos ... 241 6.5.3. Comparación de estrategias de puesta en marcha. ........................................... 246 6.5.4. Conclusiones parciales................................................................................... 248 6.6. RESULTADOS Y DISCUSIÓN DE LA CODIGESTIÓN DE PURÍN CON TIERRAS DECOLORANTES DE ACEITE DE OLIVA. ................................................................... 249 6.6.1. Digestión anaerobia de purín de cerdo. Rango termofílico (R1) ....................... 249 6.6.2. Digestión anaerobia de purín de cerdo. Rango mesofílico (R2) ........................ 257 6.6.3. Codigestión de purín con tierras decolorantes de aceite de oliva en rango mesofílico (R3). ......................................................................................................................... 264 6.6.4. Codigestión con tierras decolorantes de aceite de oliva - Rango termofílico (R4).273 6.6.5. Inhibición por amonio. Influencia de la temperatura....................................... 280 6.6.6. Producción de gas según los diferentes regímenes de alimentación...................... 283 6.6.7. Diferencias en los niveles de acidificación. Comparación de la eficiencia de la hidrólisis-acidogénesis.................................................................................................. 287 6.6.8. Influencia de la VCO sobre la producción de biogás. ..................................... 287 6.6.9. Comparación de la eficiencia de la hidrólisis entre los diferentes reactores, mediante el uso del parámetro NH4+/Nk................................................................................. 288 6.6.10. Comparación con los resultados obtenidos en discontinuo y con el modelo de simulación. ................................................................................................................. 290 6.7. CONCLUSIONES DE LOS ENSAYOS EN CONTINUO ....................................... 290 6.7.1. Sobre el método ............................................................................................ 290 6.7.2. Sobre el período de puesta en marcha............................................................. 291 6.7.3. Sobre la viabilidad de la digestión anaerobia de purines de cerdo..................... 291 6.7.4. Sobre el efecto de la codigestión con tierras decolorantes de aceite de oliva.......... 292 6.7.5. Resumen de conclusiones ............................................................................... 293 7. CONCLUSIONES GENERALES ............................................. 295 7.1. DEL MODELO DE SIMULACIÓN DINÁMICA DEL PH .................................... 297 7.2. EXPERIMENTOS EN DISCONTINUO ............................................................... 297 7.2.1. Sobre el método ............................................................................................ 297 7.2.2. Resultados del experimento 1: Diluciones de purín - efecto del amonio............. 298 Optimización de la digestión anaerobia de purines de cerdo v 7.2.3. Resultados del experimento 2: Codigestión con residuo de pera........................ 298 7.2.4. Resultados del experimento 3. Adición de tierras decolorantes de aceite de oliva. .................................................................................................................................. 299 7.3. EXPERIMENTOS EN CONTINUO ..................................................................... 300 7.3.1. Sobre el método ............................................................................................ 300 7.3.2. Sobre el método de puesta en marcha ............................................................. 300 7.3.3. Sobre la viabilidad de la digestión anaerobia de purines de cerdo..................... 300 7.3.4. Sobre el efecto de la codigestión con tierras decolorantes de aceite de oliva.......... 301 7.4. CONCLUSIONES GENERALES.......................................................................... 302 7.5. CONSIDERACIONES DE FUTURO. ................................................................... 303 8. REFERENCIAS BIBLIOGRÁFICAS........................................ 305 9. ANEJO 1. PROGRAMA DE SIMULACIÓN DE LA DIGESTIÓN ANAEROBIA DE SUBSTRATOS COMPLEJOS ...... 321 10. ANEJO 2. PROGRAMA DE SIMULACIÓN DEL PH EN SISTEMAS ACUOSOS SENCILLOS. ................................................ 346 11. ÍNDICES DE TABLAS Y FIGURAS ......................................... 362 vi Índice Optimización de la digestión anaerobia de purines de cerdo vii RESUMEN La digestión anaerobia de purines de cerdo puede ser una buena opción para la valorización económica de estos residuos. Una clara opción para mejorar la producción de metano, y por tanto la viabilidad económica de las plantas de digestión anaerobia, es la introducción como cosubstrato de residuos orgánicos de la industria agroalimentaria. La introducción de cosubstratos en un reactor puede comportar problemas de inhibición y sobrecarga orgánica, y ser capaz de preverlo puede ser vital para el funcionamiento de una planta. Por ello, los modelos capaces de predecir el comportamiento dinámico de reactores anaerobios son de gran interés. El pH es una variable fundamental en los sistemas anaerobios, cuyo comportamiento dinámico influye en multitud de procesos que ocurren en un reactor anaerobio, tanto biológicos como físico-químicos. Por tanto, un modelo dinámico estructurado debe ser capaz de simular de forma dinámica esta variable, siendo esta una asignatura pendiente en los modelos que se encuentran en la bibliografía. Se construyó un algoritmo de simulación dinámica del pH en sistemas anaerobios, basado en la ecuación del balance de cargas. Este algoritmo puede ser incorporado a un modelo dinámico de simulación, tratando el pH como una variable de estado del sistema y no como un parámetro a calcular en cada paso de integración. El algoritmo de cálculo de pH se validó para sistemas acuosos sencillos, habiendo obtenido buenos ajustes a datos experimentales. El algoritmo de cálculo de pH se introdujo en un modelo dinámico, previo, de digestión anaerobia de substratos complejos. Se incorporó, además, la simulación dinámica de las variables de la fase gaseosa. Este modelo general de digestión anaerobia, que utiliza parámetros obtenidos de la bibliografía, ha resultado una herramienta útil para prever la respuesta del sistema ante la introducción de cosubstratos, los estados de sobrecarga o la introducción de componentes inhibidores. Se realizaron ensayos de viabilidad en discontinuo de mezclas de purín con residuos de la elaboración de zumos de frutas (pulpa de pera) y con residuos del refinado de aceite de oliva (tierras decolorantes). El experimento fue desarrollado en los rangos termofílico y mesofílico (35º y 55º). En general, los resultados en mesofílico fueron mejores que en termofílico, ya que a 55ºC mostraron una importante inhibición por amonio. En ambos rangos de temperatura la producción de metano en términos absolutos mejoró por la adición de residuo de pera como cosubstrato. La producción máxima de metano respecto a sólidos volátiles se obtuvo para la mezcla con un 5% de viii Resumen TDO y 95% de purín, en el rango mesofílico, que alcanzó un valor 2,4 mayor que la producción de purín sólo, disminuyendo la producción de metano conforme aumentó la proporción de TDO. En el rango termofílico, en contraste, la codigestión con tierras decolorantes resultó en producciones de metano más bajas que el control. Con el objetivo de confirmar los resultados obtenidos en discontinuo para mezclas de purín y TDO, se pusieron en marcha cuatro reactores anaerobios en régimen semicontinuo, de 5 litros de capacidad, dos de ellos trabajando en el rango mesofílico (35ºC) y otros dos en termofílico (55ºC). En el rango mesofílico el reactor alimentado con mezcla de purín y TDO mejoró ampliamente los resultados obtenidos en el reactor con purín sólo. A pesar de la mejora en la producción de gas, se observó acumulación de AGV, indicando la inhibición parcial del sistema. En el rango termofílico, los resultados fueron en todos los casos peores que en mesofílico, con fuerte acumulación de AGV, especialmente acético, mostrando que el sistema estaba fuertemente inhibido por las altas concentraciones de amonio y de ácidos grasos de cadena larga. La codigestión se muestra como una opción interesante para incrementar las producciones de gas de purines, posibilitando el tratamiento de algunos residuos industriales, pero debe estudiarse previamente las condiciones óptimas de mezcla. Optimización de la digestión anaerobia de purines de cerdo ix RESUM La digestió anaeròbia de purins de porc pot ser una bona opció per a la valorització econòmica d’aquests residus. Una clara opció per a millorar la producció de metà i, per tant, la viabilitat econòmica de les plantes de digestió anaeròbia, consisteix en l’introducció, com cosubstrats, de residus orgànics de l’indústria agroalimentaria. La introducció de cosubstrats en un reactor pot comportar problemes de inhibició i sobrecàrrega orgànica i ser capaç de preveure-els pot ser vital per al funcionament d’una planta de tractament. Per aquest motiu els models capaços de predir el comportament dinàmic dels reactors anaerobis son de gran interès. El pH és una variable fonamental en els sistemes anaerobis, i el seu comportament dinàmic té influència en multitud dels processos que ocòrren en un reactor anaerobi, tant biològics com físico-químics. Per tant, un model dinàmic estructurat ha de ser capaç de simular de manera dinàmica aquesta variable, essent aquesta una assignatura pendent en els models que es troben a la bibliografia. Es va construir un algorisme de simulació dinàmica del pH en sistemes anaerobis, basat en l’equació del balanç de càrrega. Aquest algoritme pot ser incorporat a un model dinàmic de simulació, tractant el pH com una variable d’estat del sistema i no com un paràmetre a calcular en cada pas de la integració. L’algorisme de càlcul de pH es va validar per sistemes aquosos senzills, havent obtingut bons ajusts a dades experimentals. El algoritme de càlcul de pH es va introduir a un model dinàmic, previ, de digestió anaeròbia de substrats complexos. Es va incorporar, a més, la simulació dinàmica de les variables de la fase gasosa. Aquest model general de digestió anaeròbia, què utilitza paràmetres de la bibliografia, resulta una eina útil per preveure la resposta del sistema a la introducció de cosubstrats, a estats de sobrecàrrega o de introducció de components inhibidors. Es van realitzar assaigs en discontinu de viabilitat de mescles de purí amb residus de la elaboració de sucs de fruites (residu pera) i del refinat d'oli d’ oliva (terres decolorants d’oli d’oliva). L’experiment va ser desenvolupat en els rangs termofílic i mesofilic (35º i 55º). En general, els resultats en mesofilic van ser millors que en termofílic, donat que a 55ºC van patir una important inhibició per amoni. En ambdós rangs de temperatura la producció de metà en termes absolutes va millorar per l’addició de residu de pera com cosubstrat. La màxima producció de metà va ser obtinguda amb la codigestió de purí amb terres decolorants d'oli d'oliva (95% de purí i 5% terres), en el rang mesofílic, què va ser 2,4 vegades la producció de purí sol, disminuint en augmentar la proporció de terres decolorants. En contrast, en el rang termofílic, la codigestió amb terres decolorants va donar lloc a produccions de metà menors que el control. Amb l’objectiu de confirmar els resultats obtinguts en discontinu per a les mescles de purí i terres decolorants d’oli d’oliva, es van posar en marxa x Resumen quatre reactors anaerobis en règim semicontinu, de 5 litres de capacitat, dos dels quals treballaven en el rang mesofílic (35ºC) i els altres dos en termofílico (55ºC). En el rang mesofílic els reactors que es van alimentar amb terres decolorants d'oli d'oliva van millorar àmpliament els resultats obtinguts amb purí sol. A pesar de la millora en la producció de gas, es va observar acumulació d'àcids grassos volàtils, indicant la inhibició parcial del sistema. En el rang termofílic, els resultats van ser, en tots els casos, pitjors que en mesofílic, amb una forta acumulació d'àcids grassos volàtils, especialment d’àcid acétic, mostrant que el sistema estava fortament inhibit per les altes concentracions d'amoni i d’àcids grassos de cadena llarga. La codigestió apareix com una opció interessant per a incrementar les produccions de gas a partir de purins, possibilitant el tractament d’alguns residus industrials, però s’ha d’estudiar prèviament les condicions òptimes de mescla. Optimización de la digestión anaerobia de purines de cerdo xi ABSTRACT Anaerobic digestion of pig slurry can be a good option to valorise this waste. The use of organic wastes from food industry as cosubstrate is an interesting option to increase methane production and the economic feasibility of anaerobic plants. The addition of cosubstrates in an anaerobic reactor may involve inhibition and overloading problems. Being able to foresee these problems is very important for the plant running. Therefore, models capable of predicting the dynamic behaviour of anaerobic reactors are very interesting. pH is a very important parameter in anaerobic systems, and its dynamic behaviour largely influences physic-chemical and biological processes that take place in an anaerobic reactor. Therefore, a structured dynamic model must be able of simulating pH in a dynamic way, however no model in literature has considered it before. A mathematical algorithm was built for anaerobic dynamic simulation of pH in anaerobic systems, based on the charge balance. This algorithm can be added to a dynamic simulation model, treating pH as a system state variable, not as a parameter to be calculated in every step of integration. The algorithm of pH calculation was validated for a simple aqueous system, obtaining high regression coefficients for the experimental data. The pH calculation algorithm has been inserted in a previous anaerobic digestion model of complex substrates. The dynamic simulation of gas phase variables was also inserted. This general model of anaerobic digestion, which uses kinetic parameters from literature, has been a useful tool in order to foresee the system behaviour when adding cosubstrates, when overloading or when introducing inhibitors. Batch experiments of mixtures of pig slurry and wastes from juice factory (pear waste) and with wastes from refining oil factory (olive oil bleaching earth) were done. The experiment was developed at thermophilic and mesophilic temperatures (55ºC and 35ºC). In general, the mesophilic results were better than the thermophilic one. At 55ºC there was an important inhibition by ammonium. In both ranges of temperature, accumulated methane production was improved by the addition of pear waste as cosubstrate. The treatment with 5% of olive oil bleaching earth and 95% of pig slurry, in the mesophilic range, got the maxim methane yield (mL of CH4/g VS), which was 2,4 times the methane yield for pig slurry. The methane yield decreased with increasing olive oil bleaching earth proportions. In contrast, at the thermophilic range, the codigestion of pig slurry and olive oil bleaching earth resulted in methane yields lower than the control. xii Resumen With the aim of confirming the batch results, four anaerobic reactors of 5 litres of useful capacity were started up, in semicontinous regime. Two of them ran at mesophilic (35ºC) and other two in thermophilic (55ºC) temperatures. At mesophilic temperatures, the reactor fed with a mixture of slurry and olive oil bleaching earth showed better results than the control reactor, fed with pig slurry. In spite of improving methane production, volatile fatty acids accumulated, showing a partial inhibition of the system. In both thermophilic reactors, the results were worse than those at mesophilic temperature. A strong accumulation of volatile fatty acids, especially acetate, was observed, showing that the system was inhibited by the high concentration of ammonium and long chain fatty acids. Codigestion is an interesting option to increase the gas production from pig slurry, improving the feasibility of the treatment of some kinds of industrial wastes, but it is necessary to study the optimum conditions of mixture. 1. INTRODUCCIÓN Y OBJETIVOS 2 Introducción y objetivos Optimización de la digestión anaerobia de purines de cerdo 3 1.1. Generación de residuos orgánicos Los cambios socioeconómicos de las últimas décadas, altas concentraciones de población en núcleos urbanos, desarrollo de la industria agroalimentaria, intensificación de las explotaciones ganaderas, prácticas consumistas, etc., han propiciado la producción de grandes cantidades de residuos orgánicos que ocasionan graves problemas ambientales. Una primera clasificación de los residuos se puede realizar en función de su origen, distinguiendo los residuos procedentes del sector primario, residuos agrícolas, ganaderos y forestales, los procedentes del sector secundario, residuos industriales (agroalimentarios, textiles, etc.) y finalmente los procedentes del sector terciario o de servicios, constituidos por residuos sólidos urbanos (RSU) y lodos de estaciones depuradoras de aguas residuales (Pomares, 1998). 1.1.1. Residuos domésticos Uno de los grandes tipos de residuos son los residuos municipales o domésticos. Tanto la producción de este tipo de residuos, como las vías de tratamiento evolucionan muy rápidamente, debido, tanto al cambio en los hábitos de consumo, como al desarrollo de determinadas políticas estratégicas. Existe una clara tendencia hacia la recogida selectiva de materiales diversos, papel, vidrio, envases, residuos peligrosos, escombros y de materia orgánica. De esta forma la fracción orgánica de residuos sólidos urbanos resulta cada vez de “mayor calidad” y más fácil de estabilizar por métodos biológicos. La gestión tradicional de este tipo de residuos es el vertido en vertedero sanitario controlado, pero la tendencia, sobre todo a partir de la promulgación de la directiva 31/99, es a minimizar la fracción que va a vertedero. El nuevo Plan Nacional de Residuos Urbanos (Resolución 2110 de 13 de enero de 2000), establece la jerarquización de opciones para la gestión: 1.- Prevención-minimización 2.- Reutilización (especialmente para envases) 3.- Reciclado (papel, vidrio, plásticos, etc.) 4.- Valorización de la materia orgánica. 5.- Valorización energética (tratamientos térmicos: incineración y gasificación). 6.- Eliminación a vertedero, cumpliendo los requisitos técnicos 4 Introducción y objetivos establecidos en la directiva 31/99. La producción de residuos varía en función de la zona geográfica (Tabla 1.1), siendo mayor en las comunidades dónde los sectores industrial y servicios tienen mayor importancia (Baleares, Canarias, Cataluña, País Vasco y Comunidad Valenciana). Tabla 1.1. Generación de residuos en España en el año 1996 (Datos del Plan Nacional de Residuos Urbanos) Generación de residuos Tm/año Porcentaje Coeficiente de Comunidad autónoma generación kg/hab/día Andalucía 2.984.605 17,38 1,13 Aragón 416.419 2,42 0,96 Asturias 401.035 2,33 1,01 Baleares 559500 3,26 2,02 Canarias 966.516 5,63 1,65 Cantabria 194.875 1,13 1,01 Castilla la Mancha 673.581 3,92 1,08 Castilla y León 1.029.036 5,99 1,12 Cataluña 2.833.061 16,50 1,27 Extremadura 412.631 2,40 1,06 Galicia 810.275 4,72 0,81 Madrid 2.012.000 11,71 1,1 Murcia 394.494 2,30 0,99 Navarra 207.261 1,21 1,09 País Vasco 1.063.549 6,19 1,39 La Rioja 103.121 0,60 1,07 Comunidad Valenciana 2.048.377 11,93 1,4 Ceuta 32.000 0,19 1,27 Melilla 32.850 0,19 1,51 Total 17.175.186 100,00 1,21 Las principales vías de valorización de la fracción orgánica, previstas en el plan, son los tratamientos biológicos, especialmente el compostaje (se prevé compostar un 50% de la fracción orgánica de residuos sólidos urbanos FORSU) y la biometanización (5% de FORSU), seguidos de la incineración con aprovechamiento energético (17,7% de los residuos sólidos urbanos RSU). El uso final previsto para los materiales compostados es su aplicación al suelo como fertilizante o enmienda orgánica. La producción de residuos urbanos ha aumentado en los últimos años. La evolución es diferente en función del tipo de residuo, así como la incidencia de la recogida selectiva. En el año 98 la producción de RSU en Cataluña fue Optimización de la digestión anaerobia de purines de cerdo 5 de 2.996.391 toneladas, que corresponde a 1,35 kg/hab/día, mientras que en el año 96 la producción fue de 2.833.061 Tm, 1,27 kg/hab/día y en el año 91 la producción fue de 1,22 kg/hab*día. Es decir en 7 años se ha producido un incremento del 10,6%. La producción de RSU varía mucho en función de la zona de procedencia. En general se puede decir que la producción de residuos aumenta con el nivel de renta de la población. En Cataluña las máximas producciones por habitante se obtienen para las comarcas con alta incidencia del sector turístico, ya que la población censada es mucho menor que la real en épocas turísticas. En cuanto a la tipología de los residuos generados, la fracción más importante es la materia orgánica, aunque, según los datos del 98, es la que menor importancia de recogida selectiva presenta (Tabla 1.2). La fracción orgánica de RSU según estos datos fue del 56,5%, pero la tendencia actual es a disminuir, considerándose que lo normal para el período de vigencia del plan de residuos entre el 40 y 50 %. Tabla 1.2. Generación de residuos e incidencia de la recogida selectiva en el año 1998, en Cataluña (Generalitat de Catalunya, 1999). Generación (Tm) Recogida selectiva (Tm) % Recogida Selectiva Vidrio 239.711 69.941,3 29,18 Papel 674.188 95.942,6 14,23 Envases ligeros 119.886 10.206,8 8,51 Materia orgánica 1.348.376 9.245,4 0,67 Pilas 2.015 440,2 21,84 El destino principal de los residuos urbanos en Cataluña, en el año 99, fue la deposición controlada, que alcanzó un 71%, seguida de la incineración con un 22% y de la recogida selectiva para reciclaje (vidrio, papel, latas) con un 6,1%. La estabilización de la materia orgánica mediante tratamientos biológicos (compostaje) supuso, tan sólo, el 0,8% del volumen total. Otro residuo procedente del sector doméstico o municipal son los lodos de depuradora de aguas residuales urbanas. La mayoría de las depuradoras en servicio en Cataluña son depuradoras biológicas de tipo lodos activos, con 193 instalaciones. Existen también 9 depuradoras físico-químicas que sirven a un elevado porcentaje de población (42%). El principal residuo generado en las Estaciones Depuradoras de Aguas Residuales (EDAR) son los lodos, 6 Introducción y objetivos constituidos principalmente por la biomasa microbiana. Esta biomasa precisa de una estabilización de la materia orgánica y de un posterior postratamiento para su uso. Las principales instalaciones de postratamiento son compostaje, secado térmico, digestión anaerobia, cogeneración con gas natural y cogeneración con biogás, previa digestión anaerobia. El destino final de los lodos de depuradora es la aplicación a suelos agrícolas, vertido al mar o vertido en vertedero controlado (Tabla 1.3). Tabla 1.3. Destino final de lodos de EDAR en 1998 (Generalitat de Catalunya, 1999) Destino Lodo fresco Lodo seco Toneladas % Toneladas % Aplicación al suelo 237.968 9,14 45.544 18,59 Vertedero 125.343 4,81 34.456 14,07 Mar 2.239.903 86,04 164.939 67,34 Total 2.603.214 100,00 244.939 100,00 1.1.2. Residuos industriales El principal sector industrial en Europa es el alimentario (incluye alimentario, bebidas y fermentación), seguido de la industria papelera (Wheatley, 1990). En Cataluña, el sector de alimentación y bebidas es uno de los más importantes dentro del sector industrial (Tabla 1.4), con un 17,2% del VAB del sector. La tipología de residuos industriales es tan amplia como el número de industrias existentes. En los últimos años la declaración y la fracción de residuos que son correctamente gestionados ha aumentado mucho, pasando de 1 millón de toneladas en 1985 a cerca de 4,5 en 1997. El tipo de tratamiento de los residuos producidos dependerá, nuevamente, del tipo de residuos de que se trate, siendo las principales vías la valorización, el uso como subproducto, tratamiento físico-químico o biológico, vertido controlado e incineración. Recientemente ha sido aprobado en Cataluña el catálogo general de residuos (Decreto 34/1996 de 9 de enero, modificado por el Decreto 92/1999 de 6 de abril), dónde se catalogan todos los residuos procedentes tanto del sector industrial como urbano y agrícola, a la vez que se proponen alternativas de tratamiento y vías de valorización. El grupo 0201 del catálogo Optimización de la digestión anaerobia de purines de cerdo 7 incluye residuos de la producción primaria, la agricultura, mataderos y salas de despiece, y el grupo 0202 residuos de la industria agroalimentaria, excepto bebidas, y el 0203 la producción de bebidas. Se puede decir, por tanto, que la mayoría de los residuos industriales de naturaleza orgánica fermentables están comprendidos dentro del grupo 2 del catálogo. Menor importancia tienen los residuos del grupo 3, de la industria de la madera y del papel, y del grupo 4, de la industria de la piel y textil. La producción de residuos por parte del sector alimentario y bebidas supone, según la declaración de los años 97 y 98, el 17% del total de los residuos generados en Cataluña, siendo el grupo más importante en cuanto a la producción de residuos, seguido de la metalurgia, de la industria química y de la industria del papel, con una cantidad total de residuos declarados de 790.427 toneladas en 1997 (Generalitat de Catalunya, 1999). Tabla 1.4. Distribución de la contribución de los distintos sectores de actividad al Valor Añadido Bruto en Cataluña (1995) Sector de actividad Valor Añadido VAB Bruto (VAB) (%) Agricultura, ganadería, caza, silvicultura y pesca 152.344 1.21 Productos energéticos 655.978 5.19 Industria 3.295.385 26.09 Minerales metálicos férricos y no férricos 28.657 0.23 Minerales no metálicos 179.839 1.42 Productos químicos 537.405 4.25 789.673 6.25 Productos metálicos, maquinaria y material eléctrico Material de transporte 260.781 2.06 Productos alimentarios, bebidas y tabaco 566.871 4.49 Productos textiles, cuero, calzado y confección 396.568 3.14 Papel, edición y artes gráficas 253.917 2.01 Productos de industrias diversas 281.674 2.23 Construcción y obras de ingeniería civil 941.655 7.46 Servicios destinados a la venta 7.187.476 56.91 1.205.401 9.54 Servicios de administración general, enseñanza, investigación, sanidad, doméstico, etc. Producción imputada a servicios bancarios -807.688 -6.39 Total 12.630.551 100% 1.1.3. Residuos ganaderos Los residuos agrícolas son la mayor fuente de residuos y de potencial contaminante en Europa, y dentro de éstos, los residuos ganaderos constituyen el principal problema ambiental (Hobson, 1990). 8 Introducción y objetivos La problemática asociada a la gestión de los residuos orgánicos de origen ganadero se debe, básicamente, a la separación progresiva de la explotación ganadera y la agrícola, de forma que la mayoría de las explotaciones no poseen una base territorial suficiente para reutilizar los residuos ganaderos. Esto, junto con el aumento del censo ganadero, sobre todo el porcino, la disminución de la superficie agrícola útil, y el aumento de las dimensiones de las explotaciones ganaderas, hace equiparable el sector ganadero con la industria en cuanto a la problemática de gestión de residuos (Danés et al., 1996). Alemania y España son los dos países con mayor producción porcina de la Unión Europea. Así, en el año 1998, produjeron un 21% y un 17% respectivamente de la producción total de la Unión (Figura 1.1). Además, España muestra una tendencia ascendente, de forma que del año 97 al 98, aumentó en un 11,5%. Las principales áreas de producción dentro de España se localizan en la zona mediterránea, Cataluña, Aragón, Comunidad Valenciana y Murcia. (DARP, 1999), destacando Cataluña con el 25% de la producción porcina española, y con el 4,5% de la producción total de la Unión Europea. Dinamarca 9.5% Holanda 10.7% Resto España 12.6% Cataluña 4.5% Resto Europa 33.6% Francia 8.1% Alemania 21.0% Figura 1.1. Distribución de los cerdos de engorde en Europa (Datos de Estadístiques i Conjuntura agrària, 1999) El sector ganadero tiene un gran peso en la economía de Cataluña, especialmente en las comarcas de Lleida y sobre todo en los núcleos rurales. La ganadería contribuye de forma decisiva sobre el nivel de renta de la zona, teniendo un papel fundamental en el mantenimiento de la población rural. La producción porcina supuso, en el año 1996, un 33% de la renta agraria total de Catalunya, y sumando las producciones porcina, avícola y bovina superaban el 50% de la producción final agraria (Figura 1.2). El peso específico del sector ganadero sobre la renta agraria muestra, además, una Optimización de la digestión anaerobia de purines de cerdo 9 tendencia ascendente, comparando datos de 1992 a 1996. Dentro de Cataluña, la principal provincia productora de porcino es Lleida (Tabla 1.5), que produce el 46% del total de Cataluña. Dentro de la provincia, la mayor concentración se centra en las comarcas del Segrià y la Noguera, seguidas del Pla d’Urgell, Urgell y Garrigues (Tabla 1.6). Vino y subproductos 4% Flores y planta ornamental 0% Otros 13% Producción porcina 33% Huevos 3% Cereales 5% Leche 7% Hortalizas 5% Bovino 8% Fruta fresca 9% Avícola 11% Figura 1.2. Principales productos de la producción final agraria (adaptados de DARP, 1999) Tabla 1.5. Censo de ganado según la encuesta de diciembre de 1998 (DARP, 1999). Equivalentes en unidades de ganado mayor según el Código de buenas prácticas agrarias (DOGC 2761, 1998). Nº de animales Equivalentes de UGM Lleida 2.537.425 292 576 Porcino Cataluña 5.557.223 629 782 Lleida 255.355 50 158 Bovino Cataluña 674.332 120 114 Lleida 404.637 26 917 Ovino Cataluña 1.134.177 77 063 La mayoría de las granjas en Cataluña están integradas por grandes o pequeñas cooperativas y empresas que les proporcionan los lechones y el pienso y les facilitan la comercialización, estando en torno al 80-85% en 10 Introducción y objetivos Cataluña y al 90-93% en Lleida (DARP, 1999). No obstante en los últimos años ha aumentado la proporción de granjas de ciclo cerrado (con reproductores) en el área de Lleida, lo que proporciona un mayor valor añadido, pero aumenta la complejidad de la gestión de la granja. El aumento de la producción porcina conlleva, necesariamente, el aumento de la producción de residuos. El principal problema para la gestión de los residuos no es tanto la cantidad total, sino la excesiva concentración en determinadas áreas, que supera la capacidad de aceptación del medio. La cantidad y calidad de residuos producida varía mucho, dependiendo del tipo de animal, de la composición de la alimentación y del sistema de manejo de la granja (sistema de alimentación, bebederos, sistema de limpieza, tipo de estercolero o balsa, etc.). A pesar de ello, en la bibliografía hay valores medios de producción de purín por animal y día. A partir de los datos del censo de diciembre de 1998, y de las equivalencias propuestas por Danés et al. (1996), se puede estimar la producción de purín en las comarcas de Lleida (Tabla 1.6). Debido a que la cabaña ha presentado modificaciones en los últimos años, otras fuentes sitúan la producción de purines en 12 Mtoneladas/año. El contenido de nutrientes, N, P y K del residuo, depende directamente de la dieta. De hecho, mediante la formulación del pienso el contenido de N del purín se podría llegar a reducir hasta un 50-65% (Pomares et al., 1999). La aplicación excesiva de residuos ganaderos al suelo contribuye a la contaminación de las aguas, tanto superficiales como subterráneas, por nutrientes y por organismos patógenos. También son importante los efectos sobre la atmósfera, por la producción de olores y emisiones gaseosas de NH3, SH2, NOx, compuestos orgánicos volátiles, etc., procedentes las balsas de almacenamiento y de la aplicación al suelo. Finalmente, contribuyen a la contaminación del suelo, resultado de una aplicación excesiva de nutrientes, llevando a un desequilibrio y a la acumulación de determinados elementos, incluyendo algunos metales pesados. 1.1.4. Gestión integrada de residuos La coexistencia de diversas tipologías de residuos en una misma área geográfica posibilita la gestión integrada de residuos de diversos orígenes. Las ventajas de la gestión integrada van desde la adecuación de la composición del producto final a un suelo o cultivo concreto, la mejora del proceso de tratamiento, y el abaratamiento de los costes de transporte. Por otra parte, la gestión en sí misma puede resultar más complicada al intervenir diversos productores de residuos, que incluso pueden proceder de sectores Optimización de la digestión anaerobia de purines de cerdo 11 productivos diferentes. Tabla 1.6. Estimación de la producción anual de purín, basada en el censo de porcino del año 98 (DARP, 1999) y las equivalencias propuestas por Danés et al. (1996). Madre Cerdo de engorde Madre Madre Macho con gestante reprod. 20-50 kg 50-100 kg seca lechones L purín / 3.5 6.5 10 12 16 10 animal*día Nº de animales Garrigues 53032 95893 1914 10061 4207 247 Noguera 138457 255988 6602 33716 15262 925 Pla d'Urgell 80472 147449 1969 10674 4809 305 Segarra 48806 92281 1497 5598 3932 278 Segrià 212223 389075 8554 38406 19724 1042 Urgell 72842 136520 2250 11654 5572 347 Lleida 650868 1207631 26826 135087 63576 3832 Cataluña 1483722 2296314 49780 341736 136959 11857 Producción de purín (m3 purín/año) Garrigues 67.748 227.506 6.986 44.067 24.569 Noguera 176.879 607.332 24.097 147.676 89.130 Pla d'Urgell 102.803 349.823 7.187 46.752 28.085 Segarra 62.350 218.937 5.464 24.519 22.963 Segrià 271.115 923.080 31.222. 168.218 115.188 Urgell 93.056 323.894 8.213 51.045 32.540 Lleida 831.484 2.865.105 97.915 591.681 371.284 Cataluña 1.895.455 5.448.005 181.697 1.496.804 799.841 Total 902 371.778 3.376 1.048.490 1.113 535.763 1.015 335.247 3.803 1.512.627 1.267 510.013 13.987 4.771.455 43.278 9.865.079 1.2. Breve revisión legislativa La directiva 91/676/CEE relativa a la protección de las aguas por la contaminación producida por nitratos empleados en la agricultura, estableció la obligatoriedad para todos los estados miembros de definir las zonas vulnerables a la contaminación por nitratos, elaborar códigos de prácticas agrarias correctas, así como la elaboración de programas de acción respecto a las zonas vulnerables designadas. Con relación a la aplicación de residuos ganaderos al cultivo en zonas vulnerables, se establece como medida básica 12 Introducción y objetivos del programa de acción una cantidad máxima de equivalentes de nitrógeno aplicable, establecida en 170 kg N/Ha, aunque durante el primer programa de actuación cuatrienal se permitirá hasta 210 kg N/Ha, dejando abierta la posibilidad de que los estados miembros fijen otras cantidades máximas, justificadas con arreglo a criterios específicos: tipo de cultivo, duración del ciclo, precipitación, etc. Esta directiva, que tendría que haberse aplicado, en un plazo inferior a dos años desde su publicación, no fue transpuesta en el estado español hasta 1996, mediante el Real Decreto 261/1996 de 16 de febrero, sobre protección de las aguas contra la contaminación por nitratos procedentes de fuentes agrarias, en el que se transfieren todas las competencias a las Comunidades Autónomas. En Cataluña la definición de las zonas vulnerables se hizo mediante el Decreto 283/1998, de 21 de octubre, en el que se definen amplias zonas del territorio como vulnerables, coincidiendo en muchos casos con las zonas de mayor concentración ganadera. El área 6 comprende amplias zonas de las comarcas de La Noguera, Segarra, Urgell, Pla d’Urgell y Segriá. Casi simultáneamente se publicó el conjunto de recomendaciones conocidas como el código de buenas prácticas agrarias, orden de 22 de octubre de 1998, publicado en el DOGC Nº 2761 de 9/11/98. La administración está regulando el establecimiento de grandes plantas para el tratamiento de los excedentes, de acuerdo con los resultados de balances de nutrientes y planes de gestión de residuos globales por comarcas. El programa de medidas agronómicas aplicables a las zonas vulnerables se recoge en el Decreto 205/2000 de 13 de junio. En dicho programa se establecen las cantidades de nitrógeno por hectárea máximas aplicables, en función del tipo de cultivo y del área en que se encuentre. Por otro lado, la Unión Europea se ha planteado importantes objetivos en cuanto al aumento de la producción de energía eléctrica procedente de fuentes renovables, planteándose como objetivo para el año 2010, que el 12% de la energía total consumida en la Unión sea renovable. Las medidas para el fomento de las energías renovables a nivel español se han publicado en el Real Decreto 2818/1998 de 31 de diciembre, sobre la producción de energía eléctrica por instalaciones abastecidas por recursos o fuentes de energía renovables, residuos y cogeneración. Este RD dispone, entre otras medidas, de subvenciones sobre el precio de venta de energía eléctrica producida por energías renovables, utilizando como combustible principal biomasa primaria (b.6), biomasa secundaria (b.7), donde se encuadra la producción de biogás a partir de residuos, o mediante un proceso de cogeneración, siempre y cuando la energía térmica se utilice para Optimización de la digestión anaerobia de purines de cerdo 13 el tratamiento de purines (grupo d.1), lodos (grupo d.2) u otros residuos (grupo d.3), independientemente del tipo de combustible utilizado. La coyuntura de precios y primas favorece en la actualidad el uso de combustibles fósiles (gas natural) para la aplicación de la tarifa d.1, lo cual contribuye al aumento de las emisiones de gases de efecto invernadero. Teniendo en cuenta la Directiva 96/61 relativa a la prevención y a la reducción integrada de la contaminación, es necesario emplear las mejores técnicas disponibles, entendiendo como tales la fase más eficaz y avanzada de desarrollo de actividades y de sus modalidades de explotación que demuestren la capacidad práctica para reducir en general las emisiones y el impacto en el conjunto del medio ambiente (considerando emisiones al aire, agua y suelo). Para ello es necesario desarrollar procesos de tratamiento complejos capaces, no sólo de solucionar el problema de contaminación de las aguas en una determinada zona, sino que controlen de forma integral los riesgos de contaminación sobre todos los ámbitos. 1.3. Estrategias de tratamiento Un tratamiento es una combinación de procesos unitarios cuyo objetivo es la modificación de las características del residuo para su adecuación a la demanda como producto de calidad (Teira et al., 1999). Esta adecuación puede ser para equilibrar oferta y demanda en el tiempo, para mejorar el transporte y aplicación o para mejorar la composición. La idoneidad de un proceso de tratamiento dependerá de cada zona geográfica, de las necesidades que hayan puesto de manifiesto los estudios preliminares del plan de gestión, de la calidad del producto final obtenido y de los costes económicos asociados. En todo caso, el objetivo básico que se debe perseguir es el de aumentar la capacidad de gestión sobre el residuo. Los objetivos particulares pueden ser (Flotats et al., 2000): 1.- Adecuar la producción de residuos a las necesidades estacionales de los cultivos. 2.- Transportarlo fuera de la zona de aplicación del plan de gestión 3.- Valorar económicamente el residuo 4.- Adecuar la composición a los requerimientos del entorno (de suelos, de cultivos, de mínimo impacto ambiental - malos olores) 5.- Extraer y recuperar nutrientes valorizables (nitrógeno, fósforo,...) 6.- Higienizar –reducir o eliminar patógenos En la Tabla 1.7, se sintetizan las características básicas de algunos procesos 14 Introducción y objetivos susceptibles de ser aplicados en el tratamiento, indicando la fracción a la que se puede aplicar y los objetivos que cumple el tratamiento. Las combinaciones posibles de dichos procesos es muy elevada. Cuando existe una excedencia estructural, sólo caben estrategias que tiendan a la generación de productos con demanda en el mercado de los fertilizantes orgánicos y minerales, o de enmiendas orgánicas, que justifiquen económicamente el transporte y el control de calidad sobre el producto ofertado. En el planteamiento del proceso de tratamiento, y de los objetivos a cumplir, es muy importante la calidad y variabilidad del producto a tratar. Para purines, su composición varia según la dieta alimentaria, el estado fisiológico de los animales, la edad del purín, y las prácticas de manejo y limpieza de cada granja. La práctica usual, en granjas de engorde por ejemplo, es vaciar los fosos una vez acabado el ciclo, con lo cual se obtienen purines envejecidos, con elevada relación de alcalinidad, materia orgánica hidrolizada y elevada concentración de ácidos grasos volátiles. Para evitar problemas de emisiones atmosféricas de compuestos orgánicos, que constituyen junto al amoníaco, los principales causantes de males olores, caben dos estrategias: 1. Transformar parte de los materiales disueltos (orgánicos y minerales) a formas en suspensión (biomasa), mediante el proceso aeróbico heterótrofo, con el consecuente consumo de energía, para la obtención de un compuesto final de tipo orgánico. 2. Transformar parte de los materiales orgánicos a formas gaseosas combustibles (biogás), mediante el proceso anaerobio heterótrofo, para la obtención de un compuesto final de tipo mineral. Necesariamente contendrá una parte de materia orgánica, aunque sea mínima, y su calidad dependerá de los parámetros de control del proceso anaerobio. El producto final, para que tenga valor como producto fertilizante, deberá cumplir los siguientes requisitos: producto estable, con mínima concentración de materia orgánica fácilmente degradable; mínimo volumen con máxima concentración de nutrientes; relación N:P:K adecuada; mínima concentración de metales pesados y tóxicos; higienizado, con nula concentración de patógenos, semillas de malas hierbas, larvas o huevos de insectos, etc.; olor agradable, o en todo caso que no recuerde su origen; composición estable, con mínimas variaciones temporales. La digestión anaerobia, como proceso previo al secado, cumple con todos los condicionantes expuestos. Las ventajas de la inclusión del proceso de digestión anaerobia, en la estrategia de tratamiento, se sintetizan en la Tabla 1.8. Optimización de la digestión anaerobia de purines de cerdo 15 Tabla 1.7. Síntesis de operaciones aplicables al tratamiento de residuos ganaderos, en especial a purines de cerdo (T: residuo íntegro; S: fracción sólida; L: fracción líquida). (Flotats et al., 2000) Proceso 1. Balsas homogeneización, estercoleros 2. Separación de fases 3. Aplicación de encimas y bacterias a balsas 4. Nitrificación 5. Desnitrificación 6. Descomposición aeróbica heterótrofa 7. Digestión anaerobia 8. Compostaje 9. Reducción biológica de fósforo 10. Precipitación química 11. Secado/peletización 12. Evaporación/ concentración 13. Stripping/ absorción 14. Higienización térmica 15. Dosificación de aditivos 16. Ozonización 17. Filtración en membrana/osmosis inversa Aplicado a fracción S, L, o T T, S, L T T L L L, T Objetivo Regular la producción continua al consumo estacional de cultivos. Regular entradas discontinuas a plantas de tratamiento. Reducir patógenos Separar para propiciar líneas específicas de tratamiento, transporte o aplicación a fracción S o L resultante Aumentar concentración de sólidos. Transformar N amoniacal a orgánico Transformar N amoniacal a nítrico Transformar N nítrico a N2. Eliminar materia orgánica fácilmente degradable Eliminar materia orgánica S Producir CH4 (energía). Eliminar materia orgánica. Higienizar Eliminar/estabilizar materia orgánica. Higienizar. Obtener abono orgánico de calidad Transferir P soluble a fase biológica sedimentable. Eliminar materia orgánica fácilmente degradable. Transferir algunos componentes a fase sedimentable. Separar P (apatitas, estruvita) Separar agua. Reducir volumen L Separar agua. Reducir volumen L Recuperar N amoniacal T Eliminar/inactivar patógenos. Hidrólisis térmica T, L, S S L L L Modificar composición para adecuarla a cultivos o posibilitar otros procesos Oxidación compuestos orgánicos recalcitrantes L Separar sales. Reducir conductividad T, S, L 16 Introducción y objetivos Tabla 1.8. Ventajas del proceso de digestión anaerobia (Flotats et al., 2000) FACTOR Variabilidad en la composición Malos olores y compuestos orgánicos volátiles VENTAJAS DE LA DIGESTIÓN ANAEROBIA Homogeneización de la composición, más intensa cuanto mayor es el tiempo de retención. Eliminación de ácidos grasos volátiles (AGV) y otros compuestos fácilmente degradables. La materia orgánica resultante es lentamente o difícilmente degradable; los purines digeridos no presentan olor desagradable y son un producto más estable. En procesos térmicos posteriores se evitan problemas por volatilización de compuestos orgánicos. La reducción o eliminación de AGV disminuye la fitotoxicidad a los cultivos por estos compuestos. Reducción de Reducción de sólidos totales y volátiles. Reducción de materia materia orgánica orgánica degradable y mantenimiento de las concentraciones de y total. nutrientes. Transformación de nitrógeno orgánico a amoniacal. En Mineralización caso de separar la fase acuosa, el producto resultante presentará menor volumen, manteniendo la misma riqueza fertilizante Distribución de Homogeneización en la distribución de partículas, lo cual favorece partículas y de el diseño y aplicación de procesos posteriores de secado. Hidrólisis fracción soluble de partículas de pequeño tamaño y coloidales, y reducción de orgánicos solubles, con lo cual se facilita la separación entre fases solubles y en suspensión. Consistencia Consistencia pastosa de la fracción sólida de los purines digeridos, lo cual favorece su manipulación y peletización Alcalinidad Disminución muy significativa de la relación de alcalinidad. Aporte de alcalinidad para favorecer un proceso posterior de nitrificación, total o parcial. A su vez, y debido a la reducción de materia orgánica, el consumo energético en este proceso será inferior al de la nitrificación de la fracción líquida de purines frescos. Balance Balance energético positivo y proceso productor neto de energía energético renovable. Contribuye a disminuir las necesidades externas de energía para procesos térmicos posteriores. Permite el tratamiento de mezclas con otros residuos para optimizar la producción energética (codigestión), y facilitar la gestión integral de residuos orgánicos en la zona de aplicación del plan (cogestión). Emisiones de El proceso contribuye a la disminución en la generación de gases gases de efecto de efecto invernadero, si el metano producido sustituye una fuente invernadero no renovable de energía. Optimización de la digestión anaerobia de purines de cerdo 17 1.4. Codigestión de residuos orgánicos Los potenciales de producción de biogás a partir de purines de cerdo son relativamente bajos, debido al bajo contenido en materia orgánica de los mismos, comparados con otros tipos de residuos, y la baja biodegradabilidad de la misma. La codigestión de residuos ganaderos y residuos orgánicos en sistemas de mezcla completa es una metodología exitosa tanto en régimen termofílico como en el mesofílico (Brinkman, J., 1999). La principal ventaja de la codigestión está en aprovechar la sinergia de las mezclas, y compensar carencias de cada uno de los substratos por separado. En Dinamarca funcionan alrededor de 20 plantas centralizadas de producción de biogás desde los años ochenta, lo que ha posibilitado el tratamiento combinado de residuos ganaderos y residuos orgánicos procedentes de la industria alimentaria, de plantas depuradoras de aguas residuales urbanos, residuos de mataderos y la fracción orgánica de residuos sólidos urbanos (Angelidaki y Ahring, 1997a). Los residuos urbanos e industriales suelen contener altas concentraciones de materia orgánica fácilmente degradable (lípidos, carbohidratos y proteínas), por lo que presentan un mayor potencial de producción de biogás que los residuos ganaderos, de 30 a 500 m3/ton (Ahring et al., 1992; Angelidaki y Ahring, 1997a; Bardiya et al., 1996), mejorando la viabilidad económica de las plantas (Ahring et al., 1992). Sin embargo, estos residuos pueden presentar problemas para su digestión, como deficiencia en nutrientes necesarios para el desarrollo de los microorganismos anaerobios, baja alcalinidad, o excesivo contenido en sólidos que provoque problemas mecánicos (Banks y Humphreys, 1998). Los residuos ganaderos, y en concreto el purín de cerdo, pueden ser una buena base para la codigestión, porque, generalmente, presentan un contenido de agua más alto que la mayoría de residuos industriales, una mayor capacidad tampón y aportan una amplia variedad de nutrientes necesarios para el crecimiento de los microorganismos anaerobios (Angelidaki y Ahring, 1997a). Muchas experiencias de codigestión han sido llevadas a cabo, mezclando diferentes tipos de residuos, desde las experiencias a escala industrial de Dinamarca, hasta muchas otras a escala laboratorio, corroborando casi siempre las expectativas de un mayor potencial de biogás. Los efectos beneficiosos de la introducción de mezclas de residuos ganaderos con residuos industriales se han puesto de manifiesto en las plantas a escala 18 Introducción y objetivos industrial en Dinamarca. La producción media de las plantas que utilizan mezclas fue en el mes de septiembre del año 1999 de 38,5 m3 de gas/m3 de biomasa, con un máximo para la planta de Vegger de 90 m3 gas/m3 de biomasa introducida en el reactor, mientras que la media de producción para las plantas que trabajan sólo con estiércol fueron siempre menores a los 26,6 m3 de gas/m3 de biomasa, con un valor medio en el mismo período de 14,5 m3 de gas/m3 de biomasa (Danish Energy Agency, 1999). En el capítulo 5 se hace una revisión bibliográfica de detalle de experiencias previas en codigestión de diferentes tipos de residuos. 1.5. Herramientas informáticas sistemas anaerobios de simulación de La introducción de cosubstratos en un reactor puede comportar problemas de inhibición y sobrecarga orgánica y ser capaz de preverlo puede ser vital para el funcionamiento de una planta de tratamiento. Por ello los modelos capaces de simular el comportamiento dinámico de reactores anaerobios son de gran interés. El pH es una variable fundamental en los sistemas anaerobios, cuyo comportamiento dinámico influye en multitud de los procesos que ocurren en un reactor anaerobio, tanto biológicos como físico-químicos. El principal modulador de la digestión anaerobia de residuos ganaderos, especialmente en purines de cerdo, es la inhibición por amoníaco libre (Angelidaki y Ahring, 1993b), cuya concentración es una función del pH y la temperatura, aumentando con dichas variables. Pero si el efecto tóxico aumenta, el pH puede bajar, debido a la acumulación de ácidos grasos volátiles (principales intermediarios del proceso), con lo que disminuirá la concentración de amoníaco libre y, por tanto el efecto inhibidor. Además el pH afecta a las velocidades de crecimiento de los microorganismos. Por otro lado, muchos procesos de conversión biológicos en la industria son dependientes del pH, por lo que un algoritmo capaz de predecir la evolución del pH puede ser útil, no solo para una correcta simulación del proceso anaerobio, sino, en general de cualquier proceso en el que intervenga. 1.6. Objetivos generales Los objetivos generales del trabajo se resumen en los siguientes puntos: Optimización de la digestión anaerobia de purines de cerdo 19 1. Estudiar la mejora de la producción de biogás a partir de purines de cerdo mediante la codigestión de purines con otros residuos. Para la consecución de este objetivo general se plantearon los siguientes objetivos específicos: 1.1. Estudio del proceso anaerobio de mezclas de purín con residuos orgánicos procedentes de la industria agroalimentaria, tanto en discontinuo como en continuo, con diferentes proporciones de mezcla. 1.2. Comparación de la eficiencia del proceso anaerobio en los rangos mesofílico y termofílico. 1.3. Estudio de la inhibición por amonio/amoniaco en purín. 1.4. Elaboración de una metodología de trabajo sencilla y rápida en discontínuo, que proporcione suficiente información sobre la viabilidad técnica del proceso anaerobio con un nuevo substrato o mezclas de substratos. 1.5. Puesta en marcha de un banco de ensayos con reactores en régimen semicontinuo, de tipo mezcla completa, a escala piloto-laboratorio. Comparación de estrategias con el objetivo de reducir la duración de la fase de puesta en marcha, para los rangos mesofílico y termofílico. 2. Contribuir a la mejora de uno de los modelos de simulación de sistemas anaerobios más completos que se recogen en la bibliografía. Este modelo permite trabajar con diferentes tipos de substratos complejos, y es por ello válido para simular los procesos de codigestión. La mejora propuesta se basa en la introducción del pH como variable de estado. Este objetivo se divide en dos objetivos parciales: 2.1. Construcción y validación de un algoritmo que permita la predicción dinámica del pH, considerando éste una variable de estado. 2.2. Desarrollar el sistema de ecuaciones que permitan la modelización en la interfase líquido-gas. 2.3. Introducción de las ecuaciones desarrolladas en el modelo general de simulación del proceso anaerobio de substratos complejos y conseguir una herramienta útil para la simulación y optimización del proceso de codigestión. 1.7. Planificación del trabajo En el capítulo 2, se hace una revisión bibliográfica general del proceso de digestión anaerobia, válida para todos los capítulos siguientes. En el capítulo 3, se exponen los métodos analíticos de los diferentes 20 Introducción y objetivos parámetros analizados, utilizados en la parte experimental del trabajo. En el capítulo 4 se desarrolla un algoritmo de simulación dinámica del pH y su inclusión en un modelo dinámico de simulación de la digestión anaerobia. En los capítulos 5 y 6, se exponen los experimentos realizados de codigestión de purines y diferentes cosubstratos, tanto en discontinuo (capítulo 5), como en continuo (capítulo 6). En el capítulo 7 se resumen las conclusiones generales del trabajo. En el capítulo 8 se detallan las referencias bibliográficas correspondientes a todos los capítulos. Los programas de simulación, escritos en FORTRAN 77, se recogen en los Anejos 1 y 2. 2. EL PROCESO DE LA DIGESTIÓN ANAEROBIA 22 El proceso de la digestión anaerobia Optimización de la digestión anaerobia de purines de cerdo 23 La digestión anaerobia es un proceso biológico degradativo en el cual parte de los materiales orgánicos de un substrato son convertidos en biogás, mezcla de dióxido de carbono y metano con trazas de otros elementos, por un consorcio de bacterias que son sensibles o completamente inhibidas por el oxígeno. Utilizando el proceso de digestión anaerobia es posible convertir gran cantidad de residuos, residuos vegetales, estiércoles, efluentes de la industria alimentaria y fermentativa, de la industria papelera y de algunas industria químicas, en subproductos útiles. En la digestión anaerobia más del 90% de la energía disponible por oxidación directa se transforma en metano, consumiéndose sólo un 10% de la energía en crecimiento bacteriano frente al 50% consumido en un sistema aerobio (Muñoz Valero et al., 1987). 2.1. Historia de la tecnología El proceso anaerobio ocurre de forma espontánea en la naturaleza para degradar la materia orgánica, produciendo, por ejemplo, el gas de los pantanos, el gas natural de yacimientos subterráneos o incluso el gas metabólico producido en el estómago de los rumiantes. En el siglo XVIII Volta investigó e identificó el gas de los pantanos. Dalton, en 1804, estableció la composición química del metano (CH4). Hasta mediados del siglo XIX no se tuvo certeza de la participación de organismos vivos unicelulares en el proceso, siendo Beauchamp, en 1868, quién estableció la presencia de microorganismos en los procesos de producción de metano. Pasteur descubrió que mediante la temperatura se podía favorecer el desarrollo de los microorganismos más interesantes. Propoff, en 1875, descubrió que la formación de biogás sólo se producía en condiciones anaerobias. En 1884, Pasteur investigó sobre la producción de biogás a partir de residuos animales, proponiendo la utilización del biogás para la iluminación de las calles (Muñoz Valero et al., 1987). En la primera mitad del siglo XX se realizaron numerosas experiencias a escala laboratorio y piloto, alcanzando una especial importancia durante la segunda guerra mundial debido a la escasez de combustibles. Con el fin de la guerra y la fácil disponibilidad de combustibles fósiles la mayoría de las instalaciones fueron cesando en su funcionamiento. En la India, a partir de la década de los 60, se impulsó notablemente la tecnología de producción de biogás a partir de estiércol bovino con el doble objetivo del aprovechamiento energético y mantenimiento de las 24 El proceso de la digestión anaerobia propiedades fertilizantes. En China se ha fomentado, también, desde la década de los 70, la construcción de digestores, mediante programas de ámbito nacional. En los países industrializados la historia de la tecnología de biometanización ha sido diferente y el desarrollo ha estado motivado más por motivaciones medioambientales que puramente energéticas, siendo un método clásico de estabilización de lodos activos residuales de estaciones depuradoras de aguas residuales urbanas. A partir de la crisis energética de 1973, y durante la década de los ochenta, volvió a adquirir cierta importancia como forma de recuperación energética en explotaciones agropecuarias y agroindustriales. Con la bajada de los precios del petróleo, a finales de los años ochenta, el interés por la tecnología de digestión anaerobia volvió a decaer, aunque en algunos países industrializados se han desarrollado importantes programas de desarrollo de plantas anaerobias a escala industrial, teniendo como objetivos principales la gestión de residuos, principalmente ganaderos, la estabilización e higienización de los mismos, y el fomento de las energías renovables, para disminuir la emisión neta de gases de efecto invernadero. El principal exponente es Dinamarca, donde, en 1985, comenzó un programa demostración, desarrollado conjuntamente por los ministerios de agricultura, energía y medio ambiente, en un esfuerzo por demostrar el potencial de grandes plantas de digestión anaerobia como productores de energía eléctrica. Así, en 1997 se contabilizaban 19 grandes plantas que tratan conjuntamente residuos de origen industrial, residuos urbanos, lodos de depuradora y residuos ganaderos (Angelidaki y Ahring, 1997a), aunque en el año 2000 los objetivos eran duplicar la producción, y continuar aumentando hasta el año 2030. 2.2. Productos finales de la digestión anaerobia Los principales productos del proceso de digestión anaerobia, trabajando en sistemas de alta carga orgánica y en mezcla completa, son el biogás y un efluente estabilizado. Biogás Es una mezcla gaseosa formada, principalmente, por metano y dióxido de carbono y pequeñas proporciones de otros gases, como H2S, H2, NH3, etc. La composición o riqueza del biogás depende del material digerido y del Optimización de la digestión anaerobia de purines de cerdo 25 funcionamiento del proceso. En la Tabla 2.1 se muestran valores medios de composición del biogás en función del substrato utilizado. La potencial calorífica inferior del biogás es aproximadamente de 5.250 kcal/m3, para una riqueza en metano de 60%. Tabla 2.1. Componentes del biogás en función del substrato utilizado (Coombs, 1990) Componente Residuos agrícolas Lodos de Residuos depuradora industriales Gas de vertedero Metano 50-80% 50-80% 50-70% 45-65% Dióxido de carbono 30-50% 20-50% 30-50% 34-55% Agua Saturado Saturado Saturado Saturado 0-2% 0-5% 0-2% 0-1% 100-700 ppm 0-1% 0-8% 0.5-100 ppm Amoníaco Trazas Trazas Trazas Trazas Monóxido de carbono 0-1% 0-1% 0-1% Trazas Nitrógeno 0-1% 0-3% 0-1% 0-20% Oxígeno 0-1% 0-1% 0-1% 0-5% Compuestos orgánicos Trazas Trazas Trazas 5 ppm (terpernos, esteres,...) Hidrógeno Sulfuro de hidrógeno Efluente Las características del efluente, dependen mucho del tipo de sistema, pero tratando con sistemas de mezcla completa y con residuos orgánicos, se puede decir que el efluente es la mezcla del influente estabilizado y la biomasa microbiana producida. Durante el proceso anaerobio parte de la materia orgánica se transforma en metano, por lo que el contenido en materia orgánica es menor que en el influente. Se trata, además, de un producto más mineralizado que el influente, con lo que normalmente aumenta el contenido de nitrógeno amoniacal y disminuye el nitrógeno orgánico. 26 El proceso de la digestión anaerobia 2.3. Cinética de las reacciones biológicas 2.3.1. Tasa de utilización de substrato (coeficientes de producción) El crecimiento celular engloba la conversión metabólica de un substrato en sus productos, lo que hace que se libere energía en forma de ATP (ruta catabólica), que será utilizada para la síntesis celular (ruta anabólica). Catabolismo : Substrato o Pr oductos Energía Anabolismo : Substrato Energía Nutrientes o Masa Celular Resultado Global : Substrato Nutrientes o Masa Celular Productos La cantidad de masa celular, o biomasa formada, es proporcional a la cantidad de substrato y de producto. Se puede definir un coeficiente para cada tipo de bacterias, llamado coeficiente de producción, Y, que puede ser determinado experimentalmente: Producción de biomasa : Y X ,S Producción de producto : Y P ,S 'X 'S , 'P 'S (2.1) donde S: Substrato, X: Biomasa y P: Producto. Como ejemplo, el proceso metanogénico a partir de acético se puede expresar como el resultado general de la reacción de respiración y la síntesis celular, expresado por las siguientes reacciones (Mosey, 1983): Re spiración : CH 3 COOH o CH 4 CO 2 Síntesis Celular : CH 3 COOH 0,4 NH 3 o 0,4C 5 H 9 O3 N 0 ,8H 2 O General : CH 3 COOH 0,0183 NH 3 o 0,4C 5 H 9 O3 N 0.954CH 4 0.954CO 2 0 ,0366 H 2 O (2.2) El crecimiento de la población de microorganismos se puede asociar al consumo de substrato mediante el uso del coeficiente de producción: dX dt Y X ,S dS bX dt (2.3) donde bX es un término que refleja la lisis bacteriana, habiéndose asociado, también, con el concepto de energía de mantenimiento, o energía utilizada no para el crecimiento sino para el mantenimiento de los microorganismos (respiración). 2.3.2. Tasa de crecimiento En condiciones ideales, el crecimiento de las poblaciones bacterianas sigue un crecimiento exponencial en el tiempo. Puesto que el crecimiento de la Optimización de la digestión anaerobia de purines de cerdo 27 población bacteriana ocurre por división de las células individuales, la tasa de crecimiento bacteriano es proporcional al tamaño de la población. Esto lleva a la reacción autocatalítica descrita mediante la cinética de primer orden (Monod, 1950). dX dt PX , (2.4) donde P: la tasa de crecimiento específica. Considerando la respiración endógena, la tasa de crecimiento se expresará mediante la siguiente expresión: dX dt PX bX , (2.5) dónde b es la tasa de lisis o coeficiente de respiración. Al integrar la ecuación 2.5, considerando P y b constantes, se obtiene la función de la concentración de la población bacteriana en el tiempo, X X ( t 0 )e P b ·t . (2.6) En la práctica existen limitaciones al crecimiento, dadas por ejemplo, por la limitación del substrato disponible o por la presencia de tóxicos. La concentración de substrato disponible limita la velocidad de crecimiento de las poblaciones bacterianas. La forma de simular esta influencia ha sido objeto de controversia, siendo diferente en función de las condiciones y del grupo de microorganismos. En general se acepta que se cumple la cinética de Monod (1950), quién propuso una expresión similar a la ecuación de Michaelis-Menten de velocidad de reacción enzimática: P Pm S , KS S (2.7) donde P: tasa de crecimiento específica; Pm: tasa máxima de crecimiento específica; S: concentración de substrato; KS: constante de saturación. La dependencia del substrato de la velocidad de crecimiento específica (P) es de forma que si la cantidad de substrato es muy grande la tasa específica se aproxima al valor máximo y si la concentración del substrato tiende a cero, se aproxima a cero (Figura 2.1). Otras funciones del substrato se han considerado para simular la cinética de crecimiento de microorganismos anaerobios, tal y como se muestra en la Tabla 2.2. 28 El proceso de la digestión anaerobia P 100% P max 50% Ks 0% S Figura 2.1. Ilustración de la cinética de crecimiento de Monod. Tabla 2.2. Diferentes cinéticas de crecimiento de microorganismos usadas en la bibliografía (Pavlostathis y Giraldo-Gómez, 1991) Tipo de cinética dS Primer orden K ·S Grau et al. (1975) P· S X Y S0 P S · X Y KS S Monod (1950) Contois (1959) Chen y Hashimoto (1978) P dt Pm S · X Y BX S S P · X K · X Y ·S kS S0 S S P· S0 P· S KS S S BX S S P· K ·S 0 ( 1 K )S Pm· (2.8) (2.9) (2.10) (2.11) (2.12) Sobre la base de la cinética de Monod se pueden introducir nuevas modificaciones, por ejemplo para el caso de la existencia de varios substratos limitantes, mediante términos tipo Monod multiplicativos (Angelidaki et al., 1999). Así si se consideran dos substratos limitantes, S1 y S2, P P max S1 S2 . S1 K S1 S 2 K S 2 (2.13) Optimización de la digestión anaerobia de purines de cerdo 29 2.3.3. Tasa específica de utilización de substrato A partir de las expresiones 2.3 y 2.5 se puede definir la tasa específica de utilización de substrato (U), es decir la variación de substrato en el tiempo por unidad de biomasa: dX dt PX bX Y X ,S dS bX PX dt Y X ,S dS U dt 1 dS X dt P . (2.14) Y 2.3.4. Cinéticas de inhibición La presencia de un compuesto tóxico para los microorganismos se refleja en una menor tasa de crecimiento de los mismos. No todos los microorganismos se ven afectados de la misma forma por los mismos compuestos. Se dice que hay tres tipos básicos de inhibición, en función de la reversibilidad y del parámetro cinético al que afecta. A través de las constantes “biocinéticas” de la ecuación de Monod (2.7) para la tasa de crecimiento específico y de utilización de substrato, se puede ajustar el modelo para tener en cuenta los factores inhibidores. Lo más común en los modelos consultados en la bibliografía es que sea la velocidad de crecimiento específica μ la variable afectada, aunque algunas substancias pueden afectar al coeficiente de producción o a la tasa de lisis (Pavlostathis y Giraldo-Gómez, 1991). En la Tabla 2.3 se muestran las ecuaciones de los diferentes tipos de inhibición que afectan a la velocidad de crecimiento específica. Tabla 2.3. Tipos de inhibición y expresión matemática de la cinética Parámetro afectado Expresión de la cinética Inhibición no Tasa máxima de KI S * (2.15) P P max competitiva crecimiento Ks S KI I Inhibición Constante de P max S P (2.16) Competitiva saturación § I · ¸ ¨ K s ¨1 S K I ¸¹ © Inhibición Tasa máxima y P max P (2.17) Acompetitiva constante de Ks I 1 saturación. S KI En todos los casos, KI es la constante de inhibición y I la concentración de compuesto inhibidor. La expresión de la inhibición acompetitva se denomina, también, cinética de Haldane, y ha sido utilizada para expresar la inhibición por el propio substrato (S) o por el producto (P) (Andrews y Graef, 1971): 30 El proceso de la digestión anaerobia P P max 1 ; KS S 1 S Ki P P max 1 . KS P 1 S KI (2.18) También se utiliza la llamada “ecuación de Haldane generalizada” , P P max § S K 1 s ¨¨ S © Ki · ¸ ¸ ¹ n , (2.19) dónde n es el orden de inhibición. Con estos tipos de expresiones, hay un valor de concentración de substrato para el que la tasa de crecimiento es máxima (Figura 2.2). Este valor se puede determinar analíticamente en función de los valores de las constantes de saturación e inhibición, S P max n n 1 K S • K I n >P @max ; P max n 1 · § K S · n 1 § ¸¸ 1 ¨1 n n ¸ • ¨¨ n © ¹ © KI ¹ . (2.20) Relación tasa específica de crecimiento/tasa máxima (P/Pmax) 1 Sin inhibición 0,8 0,6 n=0 0,4 n=1 0,2 n=2 n=10 n=3 0 0 2 4 6 8 10 S/Ks Figura 2.2. Efecto de la concentración de substrato sobre la relación tasa específica de crecimiento/tasa máxima, para diferentes valores de n, considerando que la constante de inhibición es KI=3 KS Los procesos biológicos están afectados por la temperatura. De forma general, al aumentar la temperatura aumenta la tasa específica de crecimiento, hasta llegar al óptimo. A partir del óptimo, diferente para cada grupo de Optimización de la digestión anaerobia de purines de cerdo 31 microorganismos, la velocidad disminuye. La expresión más ampliamente utilizada en la modelización de los procesos anaerobios es la ecuación de Arrhenius (Pavlostathis y Giraldo-Gómez, 1991), (2.21) P k1 exp a1 T Tref k 2 exp a 2 T Tref . > @ > @ El pH influye en la velocidad de crecimiento de los microorganismos anaerobios. Generalmente se considera que afecta a la tasa máxima de crecimiento, mediante expresiones descritas por la función de Michaelis normalizada (Angelidaki et al., 1993), (2.22) P pH P F pH . 2.4. El proceso microbiológico y bioquímico de la digestión anaerobia. Los modelos tradicionales de digestión anaerobia dividen las reacciones que ocurren durante el proceso de mineralización de la materia orgánica en varias fases, llevadas a cabo por diferentes grupos de bacterias, relacionados entre ellos. De hecho muchas de estas reacciones ocurren simultáneamente sin una separación clara de fases. En la Figura 2.3 se muestra el esquema de las diferentes fases de la digestión anaerobia, con los principales microorganismos de los diferentes procesos y los compuestos intermediarios. La primera fase es la hidrólisis de partículas y moléculas complejas que son hidrolizadas por enzimas extracelulares producidas por los microorganismos fermentativos. Como resultado se producen compuestos solubles, que serán metabolizados por las bacterias anaerobias en el interior de las células. Los compuestos solubles, básicamente diferentes tipos de oligosacáridos y azúcares, alcoholes, aminoácidos y ácidos grasos, son fermentados por los microorganismos acidogénicos que producen, principalmente, ácidos grasos de cadena corta, alcoholes, dióxido de carbono e hidrógeno. Los ácidos grasos de cadena corta son transformados en acético, hidrógeno y CO2, mediante la acción de los microorganismos acetogénicos. Finalmente ocurre la metanogénesis, que produce metano principalmente a partir de acético y a partir de H2 y CO2. 32 El proceso de la digestión anaerobia MATERIA ORGÁNICA COMPLEJA PROTEÍNAS CARBOHIDRATOS 1 HIDRÓLISIS AMINOÁCIDOS, LÍPIDOS 1 1 AZÚCARES ÁCIDOS GRASOS, ALCOHOLES PRODUCTOS INTERMEDIOS OXIDACIÓN ANAEROBIA PROPIONICO, BUTÍRICO, VALÉRICO, ETC. FERMENTACIÓN 1 2 1 ACETOGÉNESIS 3 HIDRÓGENO, CO2 ACETICO HOMOACETOGÉNESIS METANOGÉNESIS ACETOCLÁSTICA 5 4 METANOGÉNESIS HIDROGENOTRÓFICA METANO, DIOXIDO DE CARBONO Figura 2.3. Esquema de reacciones de la digestión anaerobia de materiales poliméricos. (Pavlostathis y Giraldo-Gómez, 1991). Los números indican la población bacteriana responsable del proceso: 1: bacterias fermentativas; 2: bacterias acetogénicas que producen hidrógeno; 3: bacterias homoacetogénicas; 4: bacterias metanogénicas hidrogenotróficas; 5: bacterias metanogénicas acetoclásticas. 2.4.1. Hidrólisis. La materia orgánica polimérica no puede ser utilizada directamente por los microorganismos a menos que se hidrolicen en compuestos solubles, que puedan atravesar la membrana celular. La hidrólisis es, por tanto, el primer paso necesario para la degradación anaerobia de substratos orgánicos complejos. La hidrólisis de estas partículas orgánicas es llevada a cabo por enzimas extracelulares excretadas por las bacterias fermentativas. La etapa hidrolítica puede ser la etapa limitante de la velocidad del proceso global, sobre todo tratando residuos con alto contenido en sólidos. Incluso en casos donde las fases acidogénicas o metanogénicas son consideradas como pasos limitantes, la hidrólisis puede afectar el conjunto del proceso (Pavlostathis y Giraldo-Gómez, 1991). Cualquier substrato se compone de los tres tipos básicos de macromoléculas: Optimización de la digestión anaerobia de purines de cerdo 33 hidratos de carbono, proteínas y lípidos. La hidrólisis de cada tipo de compuesto se realiza por diferentes grupos enzimáticos. El grado de hidrólisis y la velocidad del proceso depende de muchos factores, entre otros del pH, de la temperatura, de la concentración de biomasa hidrolítica, del tipo de materia orgánica particulada (Pavlostathis y Giraldo-Gómez, 1991), y del tamaño de partícula (Hills y Nakano, 1984). Uno de los principales componentes de la materia orgánica, sobre todo en residuos ganaderos, son los materiales lignocelulósicos, compuestos principalmente por lignina, celulosa y hemicelulosa. La lignina es un material altamente refractario a la degradación anaerobia, afectando también a la biodegradabilidad de la celulosa, de la hemicelulosa y de otros polímeros, convirtiéndose su degradación en el proceso limitante de la velocidad de la hidrólisis y por tanto, de la degradación anaerobia de determinados substratos (Sleat y Mah, 1987; Pavlostathis y Giraldo-Gómez, 1991; Veeken y Hamelers, 1999). Los principales productos de la hidrólisis de la celulosa son celobiasa y glucosa, mientras que la hemicelulosa produce pentosas, hexosas y ácidos urónicos. Las proteínas son hidrolizadas por proteasas en proteosas, peptonas, péptidos y aminoácidos. Hay proteasas extracelulares, conocidas como proteinasas que atacan la proteína entera, y las peptidasas, intracelulares, que cortan aminoácidos del extremo de proteínas y péptidos. Los aminoácidos producidos son degradados a ácidos grasos volátiles, dióxido de carbono, hidrógeno, amonio y sulfuro reducido. Generalmente la tasa de hidrólisis de proteínas es menor que la de los carbohidratos (Pavlostathis y GiraldoGómez, 1991). La degradación de lípidos en ambientes anaerobios consiste en una ruptura inicial de las grasas por un grupo de enzimas hidrolíticas (lipasas) en los correspondientes ácidos grasos de cadena larga y moléculas de glicerol o galactasa. Una molécula de fosfolípidos produce un equivalente de ácido fosfórico, uno de glicerol y dos de ácidos grasos (Pavlostathis y GiraldoGómez, 1991). La tasa de hidrólisis, en general, aumenta con la temperatura (Pavlostathis y Giraldo-Gómez, 1991; Siegrist et al., 1993; Veeken y Hamelers, 1999), independientemente del compuesto de que se trate. Hills y Nakano (1984) demostraron que la tasa de hidrólisis depende, también, del tamaño de las partículas, debido fundamentalmente a la disponibilidad de superficie para la adsorción de las enzimas hidrolíticas. Los pretratamientos físico-químicos, cuyo principal efecto es la reducción del tamaño de las partículas, producen un aumento en la tasa de hidrólisis, y si 34 El proceso de la digestión anaerobia esta fase es la limitante del proceso anaerobio, supone un beneficio para el proceso general, produciendo menores tiempos de retención y tamaños de reactor menores. En la bibliografía se relatan numerosas experiencias positivas en este sentido: pretratamiento con ultrasonidos de lodos de depuradora (Tiehm et al., 1997); pretratamiento mecánico de diferentes tipos de substratos (Baier y Schmidheiny, 1997; Hartmand et al., 1999; Palmowski y Müller, 1999); pretratamientos que combinan ultrasonidos y ataque alcalino (Chiu et al., 1997); pretratamientos térmicos (Bonmatí et al., 2000); o termoquímicos (Delgenès et al.,. 1999). La dependencia del tamaño de partícula ha motivado el desarrollo de diversos modelos que se basan en este parámetro para simular la velocidad del proceso hidrolítico (Hills y Nakano, 1984; Vavilin et al., 1995; Valentini et al., 1997; Sanders et al., 1999; Palmowski y Müller, 1999). Cinética del proceso hidrolítico. Sólo unos pocos autores consideran el paso de hidrólisis y acidificación en sus modelos, entre los que destacan Hill (1982), Jain et al. (1992), Siegrist et al. (1993), Angelidaki et al., (1993, 1996, 1999), Vavilin et al. (1994), Shin et al. (1995), Bagley et al. (1999). La mayoría de estos modelos utilizan la cinética de primer orden (Tabla 2.4) para simular la fase de hidrólisis, algunos considerando un único proceso general de hidrólisis de materia orgánica insoluble (Hill, 1982; Siegrist et al., 1993). La utilización este tipo de cinética proporciona diferentes valores de la constante de hidrólisis en función del tipo de substrato (Veeken y Hamelers, 1999), o del pretratamiento aplicado (Shimizu et al., 1993), lo que hace que haya grandes discrepancias en los valores de dicho parámetro en la bibliografía (Pavlostaths et al., 1991). Otros modelos, como el de Angelidaki et al. (1999), consideran la cinética de primer orden pero para cada uno de los tres grandes grupos de macromoléculas, siendo por ello, más extrapolable a diferentes tipos de substratos. El contenido de lignina en la fracción de carbohidratos de un substrato puede hacer variar sustancialmente la biodegradabilidad de la fracción lignocelulósica, hecho que Angelidaki et al.(1999) simulan mediante la utilización de diferentes coeficientes estequiométricos en función del tipo de substrato. Algunos modelos consideran la tasa de hidrólisis dependiente, no sólo de la concentración de substrato a hidrolizar sino también de la concentración de biomasa responsable de la producción de las enzimas hidrolíticas (Tabla 2.4). Los modelos ASM nº 2 y nº 3 y aquellos que se basan en estos (Henze et al.,1995; Henze et al., 1999; Bagley et al., 1999), simulan la fase hidrolítica del Optimización de la digestión anaerobia de purines de cerdo 35 proceso anaerobio mediante la cinética de Contois, modificación de la de Monod, en la que la constante de saturación es proporcional a la concentración de dicha población bacteriana. De hecho la cinética de primer orden se puede considerar como un caso límite de la cinética de Contois, ya que si la población bacteriana es mucho mayor que la concentración de substrato a degradar la degradación de éste es sólo dependiente de su concentración. Tabla 2.4. Comparación de las diferentes cinéticas utilizadas en la simulación de la fase hidrolítica en modelos de digestión anaerobia de substratos complejos (XS Substrato a hidrolizar; XH: Biomasa hidrolítica; Kh, kh, A y KHA, parámetros cinéticos) Cinética Contois Referencias Henze et al., 1995; Bagley et al., 1999. Primer orden Siegrist et al., 1993; Angelidaki et al, 1999. Tamaño de partícula Valentini et al., 1997. Expresión dX s dt k h · dX s dt dX S dt Xs XH KX Xs XH XH (2.23) -K h ·X s (2.24) K HA · X s · X H A (2.25) Una nueva generación de modelos están siendo desarrollados en los últimos años, basados en la disminución del tamaño de las partículas (Tabla 2.4), dado que es uno de los parámetros más influyentes (Hills y Nakano, 1984; Vavilin et al., 1995; Valentini et al., 1997; Sanders et al., 1999; Palmowski y Müller, 1999). Valentini et al.(1997) propusieron una nueva cinética general (ecuación 2.25), que en función de las condiciones particulares se aproximaría a una cinética de primer orden y que depende, no sólo de la concentración de substrato, sino también de la concentración de biomasa hidrolítica. Este autor considera que la constante de hidrólisis (KHA) depende, además, del tamaño de partícula. En función del valor de A se aproximaría a la cinética de primer orden respecto al substrato o a la cinética de primer orden respecto al substrato y la biomasa (A=1), o de orden 0,5. El valor de A que mejor se ajustó a sus datos experimentales estuvo alrededor de 0,5. El significado físico de A se asocia con la superficie de substrato disponible. Un modelo muy importante es el propuesto por Vavilin et al. (1996). Se trata 36 El proceso de la digestión anaerobia de un modelo determinístico basado en que la fase hidrólitica se divide en dos fases principales: la primera de colonización de las bacterias hidrolíticas de la superficie de las partículas de substrato; y la segunda fase de reacción de hidrólisis propiamente dicha, es decir las enzimas liberadas por las bacterias catalizan la reacción de hidrólisis, liberándose materia orgánica soluble que puede ser usada por las mismas bacterias hidrolíticas o ser liberadas al medio para ser utilizadas por otros grupos bacterianos (bacterias fermentativas). La simulación de la segunda fase la realizan mediante cinética de primer orden, donde la constante de hidrólisis es función del tamaño de las partículas a degradar y de la densidad y la forma de las mismas, aumentando al disminuir la densidad y el tamaño de las partículas. El modelo considera que la concentración de biomasa está en exceso, por lo que la tasa de hidrólisis es constante por unidad de superficie, y el factor limitante de la velocidad es la superficie disponible para la colonización bacteriana. Péringer (1999) presentó un modelo similar al anterior, simulando la fase de colonización enzimática de la superficie de las partículas y la reacción hidrolítica en sí misma, simulada mediante la cinética tipo Michaelis-Menten. Sin embargo no considera la dependencia del tamaño de las partículas, considerando la tasa máxima de hidrólisis como función del pH. Sanders et al. (1999) han desarrollado un modelo similar que considera tanto el tamaño como la densidad de las partículas a hidrolizar. Inhibición de la hidrólisis de macromoléculas La hidrólisis puede verse afectada por la presencia de algún compuesto que sea tóxico o inhibidor de la población bacteriana responsable de la producción de enzimas extracelulares. Gallert et al. (1997) encontraron que la concentración de amonio influye negativamente en la desaminación de peptonas. Angelidaki et al. (1999) consideran que la tasa de hidrólisis de carbohidratos y proteínas está limitada por la concentración total de ácidos grasos volátiles (AGV). Henze et al. (1995) considera que la tasa de hidrólisis está inhibida por la concentración de oxígeno y nitrato. 2.4.2. Etapa fermentativa o acidogénica Las moléculas orgánicas solubles son fermentadas por varios organismos fermentativos formando compuestos que pueden ser utilizados directamente por las bacterias metanogénicas (acético, fórmico, H2) y compuestos orgánicos más reducidos (láctico, etanol, propiónico, butírico, principalmente) que tienen que ser oxidados por bacterias acetogénicas a substratos que puedan utilizar las metanogénicas (Stams, 1994). Las Optimización de la digestión anaerobia de purines de cerdo 37 proporciones entre los productos de la fermentación varían en función del consumo de H2 por parte de las bacterias que utilizan hidrógeno. Cuando el H2 es eliminado de forma eficiente las bacterias fermentativas no producen compuestos reducidos como el etanol, favoreciendo la producción de H2 y la liberación de energía en forma de ATP (Pavlostathis y Giraldo-Gómez, 1991). La actividad de algunas bacterias fermentativas y acetogénicas depende de la concentración de H2, siendo posible sólo a valores muy bajos de presión parcial de H2. La eliminación continua de H2 mediante oxidación por CO2 (bacterias metanogénicas hidrogenotróficas) estimula la acción de las bacterias fermentativas, al eliminar un producto de la reacción (Boone y Xun, 1987). Fermentación de carbohidratos solubles. La ruta de degradación de la glucosa en los sistemas anaerobios proporciona como principales productos ácidos grasos volátiles, H2 y CO2. La estequiometría que se propone para este proceso varía en función de la fuente consultada. Así, McCarty (1971), como posteriormente recogió Pavlostathis y Giraldo-Gómez (1991), propone una estequiometría basada en principios bioenergéticos y termodinámicos, considerando la energía producida en la respiración y la consumida para la síntesis celular de los microorganismos responsables. Siguiendo a Pavlostathis y Giraldo-Gómez (1991), las fracciones de glucosa consumida para la respiración y para la síntesis son, respectivamente, 0.504 y 0.496. 'Gº’=-50.39 kcal/mol de glucosa, C 6 H 12 O6 0.595 NH 4 1.458HCO3 o 0.595C 5 H 7 NO 2 0.535CH 3 COO 0.190CH 3 CH 2 COO 0.140CH 3 CH 2 2 COO 2.272CO 2 2.898H 2 O º ' 'G 50.39kcal / mol glucosa. (2.26) La principal ruta metabólica de degradación de glucosa para formar ácidos orgánicos es la de Embden-Meyerhof (Figura 2.4), que tiene como principal intermediario el piruvato (Mosey, 1983). La fermentación de azúcares se realiza por diversos tipos de microorganismos, siguiendo diferentes rutas metabólicas, en función del organismo responsable, y obteniendo productos finales diferentes. Los principales microorganismos son los que producen butírico o butanol, básicamente del género Clostridium, que convierten la glucosa y algunos aminoácidos en ácido butírico, acético, CO2 y H2. La glucosa se convierte en piruvato mediante la ruta Embden-Meyerhof, y el piruvato se desdobla a 38 El proceso de la digestión anaerobia Acetil-CoA y CO2 (Madigan et al., 1998). El Acetil-CoA se reduce en los productos de fermentación empleando como transportador de electrones el NADH derivado de las reacciones glucolíticas de la ruta Embden-Meyerhof (Metzler, 1981). Las proporciones de los diversos productos se modifican por la duración y las condiciones de la fermentación, siendo el butírico y el acético los productos mayoritarios si el pH se mantiene alcalino (Madigan et al., 1998). Glucosa ATP ADP Glucosa Glucosa 6 fosfato ATP ADP Fructosa 1:6 difosfato 2 [gliceraldehido-fosfato] 2[fosfato] 2NAD+ 2NADH +2H+ 2 [1:3 difosfoglicerato] 4ADP 2CO2 2NAD+ 2HSCoA 2 [piruvato] 2 [acetil-CoA] 2NADH+2H+ 4ATP 2H2 2NADH+2H+ 4NADH+4H+ 2NAD+ 4NAD+ 2HSCoA 2H2 2 [ácido acético] [ácido butírico] 2 [ácido propiónico] Figura 2.4. Simplificación de las rutas metabólicas de degradación de la glucosa por las bacterias acidogénicas (Mosey, 1993). Las bacterias ácido-propiónicas, del género Propionibacterium, llevan a cabo un proceso distinto, conocido como fermentación acido-propiónica, en el que se produce la fermentación del ácido láctico, carbohidratos y polihidroxialcoholes, produciendo, principalmente, ácido propiónico, succínico, acético y CO2. Sus requerimientos nutricionales son complejos y crecen con lentitud. Las diferencias en el metabolismo respecto a los Clostridium se producen a partir de la formación del piruvato por la ruta Optimización de la digestión anaerobia de purines de cerdo 39 Embden.-Meyerhof. La base de la fermentación ácido-propiónica es la conversión del piruvato a oxalacetato por carboxilación y la conversión ulterior, a través de succinato y succinil-CoA a metilmalonil-CoA y propionil-CoA. Con objeto de que la oxidación-reducción resulte equilibrada dos tercios de la glucosa se transforman en propionato y un tercio en acetato (Metzler, 1981). C 6 H 12 O6 o 1,33CH 3 CH 2 COO 0 ,66CH 3 COO 2 H 0,66CO 2 0,66 H 2 O 'G ' ( pH 7 ) 310kJ 74.16 (2.27) Fermentación de aminoácidos Los principales productos de la fermentación de aminoácidos y de otras moléculas nitrogenadas son ácidos grasos de cadena corta, succínico, aminovalérico y H2 (Tabla 2.6). La fermentación de aminoácidos se considera un proceso rápido y que en general, no limita la velocidad de la degradación de compuestos proteicos (Pavlostathis y Giraldo-Gómez, 1991). La estequiometría varía mucho en función de la fuente consultada, siendo además diferente para cada aminoácido. La estequiometría propuesta por McCarty, 1974, y adaptada por Pavalostathis et al. (1991), considerando las diferentes fracciones para respiración y síntesis de biomasa, se expresa como sigue, C 4 H 6 ON 0,114 HCO3 1,478H 2 O o 0 ,247C 5 H 7 O 2 N 0 ,30CH 3 COO 0,28CH 3 CH 2 COO 0 ,286CH 3 CH 2 2 COO 0 ,753NH 4 0 ,298CO 2 'G 0 8.87kcal / mol . (2.28) Algunos organismos del género Clostridium pueden fermentar aminoácidos. Los productos finales de la oxidación son NH3, CO2 y un ácido carboxílico con un átomo de carbono menos que el aminoácido oxidado (Madigan et al., 1998). Producen n-butírico y ácido isobutírico, isovalérico, caproico, sulfuro de hidrógeno, metilmercaptano, cadaverina, putrescina (en función del tipo de aminoácido de que proceda), etc. Cinética de la fermentación de hidratos de carbono y aminoácidos La mayoría de los modelos publicados simulan la velocidad de la fermentación mediante la cinética de Monod (2.9), modificada por funciones de inhibición, aunque muchos autores no consideran inhibición, debido a la 40 El proceso de la digestión anaerobia versatilidad de los microorganismos fermentativos. La tasa específica máxima de crecimiento de los microorganismos que degradan carbohidratos varía de 0,50 a 0,20 h-1, y la constante de saturación (Ks) de 0,004 a 11,76 mM (Pavlostathis y Giraldo-Gómez, 1991). En la Tabla 2.5 se muestran una colección de parámetros cinéticos adaptada de Pavlostathis y Giraldo-Gómez (1991). Inhibidores de la fermentación de hidratos de carbono y aminoácidos. No se han descrito muchos inhibidores de esta etapa, destacándose tan sólo los ácidos grasos de cadena larga (AGCL), señalados por Angelidaki et al. (1999). La concentración de hidrógeno juega un papel regulador importante del funcionamiento de la fermentación, tal y como describen Boone y Xun (1987). Oxidación anaerobia de ácidos grasos de cadena larga (Acetogénesis de AGCL). La ruta principal de degradación de AGCL es la E-oxidación. Los ácidos grasos libres son introducidos en la célula a través de la pared celular. Este proceso puede ser desarrollado por un gran número de microorganismos, incluso un número mayor que los organismos capaces de hidrolizar las grasas. Una vez dentro de la célula, el ácido graso es convertido en el correspondiente tio-ester-CoA, lo que sirve tanto para activar su degradación, como para disminuir el efecto tóxico de los ácidos grasos libres. La E-oxidación es un ciclo en espiral que va liberando un acetil-CoA en cada bucle, produciendo, principalmente, ácido acético. Si se trata de un ácido con un número, n, impar de átomos de carbono, al final se obtendrían n-1 acetilCoA y un propionil-CoA (Ratledge, 1992). Durante el proceso se produce la deshidrogenación del ácido graso, liberándose hidrógeno molecular a través del intermediario NADH. El H2 es el principal aceptor de electrones. La E-oxidación de AGCL es una reacción endotérmica, lo que unido a la linealidad de la ruta metabólica, hace que el proceso sea muy dependiente de la acción simbiótica de los microorganismos consumidores de hidrógeno para que se pueda producir. A modo de ejemplo la degradación de un ácido graso de cadena larga (palmítico, de 16 átomos de carbono), sería de la siguiente forma (Hanaki et al., 1981): CH 3 CH 2 14 COO 14 H 2 O o 8CH 3COO 7 H 14 H 2 'G 0' 345.6kJ / mol (2.29) Optimización de la digestión anaerobia de purines de cerdo 41 Tabla 2.5. Resumen de parámetros cinéticos para las diferentes fases y diferentes grupos en el proceso de digestión anaerobia (adaptado de Pavlostathis y Giraldo-Gómez, 1991). b Ks μmax Y Substrato Tª k d-1 d-1 g DQO mg DQO/L gSSV gSSV x d g DQO Fermentación de carbohidratos Glucosa 35 427 0.0125 0.15 Glucosa 36.5 22.5 0.052 0.17 Glucosa 37 527 0.013 Glucosa 37 370 0.125 0.14 Oxidación anaerobia de ácidos grasos de cadena larga AGCL 20 3.85 4620 0.139 0.04 25 4.65 3720 0.171 0.04 35 6.67 2000 0.252 0.04 AGCL saturados Místirico 37 0.95 105 0.105 0.11 Palmítico 1.00 143 0.110 0.11 Estereato 0.77 417 0.085 0.11 AGCL insaturados Oleico 37 4 3180 0.44 0.11 Linoleico 5 1816 0.55 0.11 Acetogénesis a partir de AGV Prop. 25 7.8 1145 0.358 0.051 Prop. 33 6.2 246 0.155 0.025 Prop. 35 7.7 60 0.313 0.042 Butírico 35 8.1 13 0.354 0.047 Butírico 60 12 0.77 Mezcla 35 17.1 166 0.414 0.030 6.1 0.015 0.015 0.015 0.01 0.01 0.01 0.01 0.01 0.04 0.010 0.027 0.099 Según Pavlostathis y Giraldo-Gómez (1991), considerando la síntesis celular, y considerando de forma inseparable la metanogénesis a partir de hidrógeno, la ecuación estequiométrica, queda (Pavlostathis y Giraldo-Gómez, 1991): CH 3 CH 2 14 COO 0 ,235 NH 4 6,822 HCO3 o 0 ,235C 5 H 7 NO 2 7.587CH 3 COO 3,319CH 4 3,850CO 2 0,530 H 2 O 'G 0 -17.50 kcal/mol (2.30) Cinética de la oxidación anaerobia de ácidos grasos de cadena larga. Pocos autores han introducido la degradación de ácidos grasos de cadena larga en sus modelos, y los que lo han hecho han utilizado la cinética tipo Monod modificada por factores de inhibición (Siegrist et al., 1993). 42 El proceso de la digestión anaerobia Angelidaki et al. (1999) consideran dos términos multiplicativos (uno referente a la limitación de AGCL y otro referente a la concentración de amonio, nutriente necesario para la síntesis celular), teniendo en cuenta también un factor multiplicativo en función del pH. La inhibición por el substrato la simulan mediante cinética de Haldane. Inhibición de la oxidación anaerobia de ácidos grasos de cadena larga. El principal inhibidor de este proceso es el H2 (Pavlostathis y GiraldoGómez, 1991; Stam, 1994). Siegrist et al. (1993) señalan también como inhibidor de este proceso la concentración de ácido acético. Angelidaki et al. (1999) consideran como principal inhibidor la propia concentración de AGCL, junto con el pH. 2.4.3. Fase acetogénica. Mientras que algunos productos de la fermentación pueden ser metabolizados directamente por los organismos metanogénicos (H2 y acetato), otros (valeriato, butirato, propionato, algunos aminoácidos, etc.) necesitan ser transformados en productos más sencillos, acetato e hidrógeno, a través de las bacterias acetogénicas (Tabla 2.6). Representantes de los microorganismos acetogénicos son Syntrophomonas wolfei y Syntrophobacter wolini (Boone y Bryant, 1980). Los procesos acetogénicos son energéticamente difíciles, por lo que necesitan ser ”ayudados” por los organismos metanogénicos u otros organismos consumidores de hidrógeno (Stams, 1994) y la energía libre de la reacción depende de la presión parcial de hidrógeno del medio (Figura 2.5). Un tipo especial de microorganismos acetogénicos, son los llamados homoacetogénicos, que consumen H2 y CO2, y producen acetato. Los principales exponentes son Acetobacterium woodii o Clostridium aceticum. Este tipo de bacterias son capaces de crecer heterotróficamente en azúcares, al contrario que los metanogénicos, siendo más parecidos a los fermentativos que a los metanogénicos, a pesar de utilizar los mismos substratos (Madigan et al., 1998). Algunos autores han considerado este proceso en sus modelos, como Hill (1982) que considera un 10% del acetato formado por esta vía. Cinética de la acetogénesis. La mayoría de los modelos que consideran esta fase aplican la cinética de Monod para su simulación matemática, aunque modificada por coeficientes de inhibición (Hill, 1982; Mosey, 1983; Ahring y Westermann, 1987b; Optimización de la digestión anaerobia de purines de cerdo 43 Costello et al., 1991; Siegrist et al., 1993; Angelidaki et al., 1999). La mayoría de las funciones de inhibición utilizadas son del tipo de inhibición no competitiva reversible (2.15). Un resumen de las constantes cinéticas del modelo de Monod se muestran en la Tabla 2.5, recogidas de Pavlostathis y Giraldo-Gómez (1991). Tabla 2.6. Reacciones acetogénicas que ocurren en los sistemas anaerobios (Stams, 1994) 'Gº ' ( KJ ) Reacciones acetogénicas Etanol y láctico Etanol H O o Acetato H 2H 2 2 +9,6 -4,2 Lactato 1 2 H O o Acetato1 H 2 H HCO 2 2 3 Ácidos Grasos Acetato1 4 H O o H 4 H 2 HCO 2 2 3 +104.6 Pr opionato 1 3H O o Acetato 1 HCO H 3H 2 3 2 +76.1 Butirato 1 2 H O o 2 Acetato1 H 2 H +48.1 2 Valerato 1 3H O o 3 Acetato1 2 H 4 H 2 2 +96.2 2 Aminoácidos Alanina 3H O o Acetato 1 HCO NH H 2 H 2 3 4 2 +7.5 Aspartato 1 4 H 2 O o Acetato 1 2 HCO3 NH 4 H 2 H 2 -14.0 Leucina 3H O o isovalerato 1 HCO NH H 2 H 2 3 4 2 +4.2 Glutamato 1 4 H 2O o propionato 1 2 HCO3 NH 4 H 2 H 2 -5.8 Glutamato1 7 H O o acetato1 3HCO NH 3H 5H +70.3 2 3 4 2 Inhibidores de la acetogénesis. El principal inhibidor de la acetogénesis, cuya acumulación provoca la rápida acumulación de los substratos, es el hidrógeno molecular (Ahring y Westermann, 1987a y Fukuzaki et al., 1990), pudiéndose decir que la oxidación del propiónico sólo es posible si la presión parcial de H2 está por 44 El proceso de la digestión anaerobia debajo de 5,8*10-5 atmósferas (Boone y Mah, 1987). Otros compuestos pueden inhibir también el correcto desarrollo de las poblaciones acetogénicas, como el propio ácido acético (producto de la acetogénesis) (Ahring y Westermann, 1988; Angelidaki et al., 1993; Siegrist et al., 1993; Hyun et al., 1998), o los ácidos grasos de cadena larga (Galbraith et al., 1971; Hanaki et al., 1981; Angelidaki et al., 1999), además de estar muy afectado por el valor de pH (Siegrist et al., 1993; Angelidaki et al., 1993). 'G (kJ/e-) Oxidación de propiónico a hidrógeno, CO2 y acético Oxidación de butírico a acético Metanogénesis hidrogenotrópica Presión de Hidrógeno (atm) Figura 2.5. Relación entre la energía libre de la reacción y la presión parcial de hidrógeno (Boone y Mah, 1987). 2.4.4. Fase metanogénica. Los microorganismos metanogénicos pueden ser considerados como los más importantes dentro del consorcio de microorganismos anaerobios, ya que son los responsables de la formación de metano y de la eliminación del medio de los productos de los grupos anteriores, siendo, además, los que dan nombre al proceso general de biometanización. Las bacterias metanogénicas son las responsables de la formación de metano a partir de substratos monocarbonados o con dos átomos de carbono unidos por un enlace covalente: acetato, H2, CO2, formato, metanol y algunas metilaminas. Los organismos metanogénicos se clasifican dentro Optimización de la digestión anaerobia de purines de cerdo 45 del dominio Archaea, y, morfológicamente, pueden ser bacilos cortos y largos, cocos de varas ordenaciones celulares, células en forma de placas y metanógenos filamentosos, existiendo tanto Gram positivos como Gram negativos (Madigan et al., 1998). Todas las bacterias metanogénicas que se han estudiado poseen varias coenzimas especiales, siendo la coenzima M, la que participa en el paso final de la formación de metano (Madigan et al., 1998). Se pueden establecer dos grandes grupos de microorganismos, en función del substrato principal, dividiéndose en los hidrogenotróficos, que consumen hidrógeno y fórmico, y los metilotrópicos o acetoclásticos, que consumen grupos metilos del acetato, metanol y algunas aminas (Cairó y París, 1988). Las principales reaccciones metanogénicas se recogen en la Tabla 2.7. La mayoría de los organismos metanogénicos son capaces de utilizar el H2 como aceptor de electrones, mientras que sólo dos géneros son capaces de utilizar el acetato (Ferguson y Mah, 1987). A pesar de ello, en ciertos ambientes anaerobios, éste es el principal precursor del metano, considerándose que alrededor del 70% del metano producido en los reactores anaerobios se forma a partir de acetato (Jeris et al.,1965, citado en Ferguson y Mah, 1987). Los dos géneros que tienen especies acetotróficas son Methanosarcina y Methanothrix, siendo el principal exponente Methanosarcina barkeri, que es capaz de crecer en diversos substratos, entre los que están H2 y CO2, acetato, metanol, metilaminas y CO (Cairó y París, 1988). Cinética de la metanogénesis La mayoría de los modelos utilizan la cinética de Monod para simular el crecimiento de los microorganismos metanogénicos, considerando como substrato principal el acetato. Una colección de parámetros cinéticos, tomada de Pavlosthatis y Giraldo-Gómez (1991), se muestra en la Tabla 2.8. Algunos autores consideran separadamente la simulación de los organismos hidrogenotróficos, aunque muchos otros lo consideran inseparable de la fase acetogénica (Angelidaki et al., 1993; 1999). Puesto que la acetogénesis no puede desarrollarse a no ser que el consumo de hidrógeno sea muy eficiente (Figura 2.5), puede, que en los ambientes donde no haya problemas de acumulación de hidrógeno, sea suficiente con este tipo de modelo. No obstante, hay toda una generación de modelos que se basan, precisamente, en el papel regulador del hidrógeno. 46 El proceso de la digestión anaerobia Tabla 2.7. Principales reacciones metanogénicas y otras consumidoras de hidrógeno (adaptada de Stams, 1994 y Fergusson et al., 1987). Reacciones hidrogenotróficas 4 H 2 H 2 HCO3 o Acetato 4 H 2 O 'G0(KJ) -104.6 4 H 2 4 Sº o 4 HS 4 H -112 4 H 2 2 HCO3 H o CH 4 3H 2 O -135.6 4 H 2 4 SO42 H o HS 4 H 2 O -151.9 4 H 2 4 fumarato o 4 succinato -344.6 -599.6 4 H 2 NO3 2 H o NH 4 3H 2O Interconversión formato-hidrógeno H 2 HCO3 o formato H 2 O -1.3 Metanogénesis acetoclástica Acetato H 2 O o HCO3 CH 4 -31.0 Metanogénesis a partir de otros substratos Fórmico 4 HCOOH o CH 4 3CO2 2 H 2 O Metanol 4CH 3 OH o 3CH 4 CO 2 2 H 2 O Trimetil-amina 4CH 3 3 N 6 H 2 O o 9CH 4 3CO 2 4 NH 3 Dimetil-amina 2 CH 3 NH 2 H 2 O o 3CH 4 CO 2 2 NH 3 2 Monometil-amina 4CH 3 NH 2 2 H 2 O o 3CH 4 CO 2 4 NH 3 Inhibición de la metanogénesis. Diversos compuestos se han descrito como inhibidores del crecimiento de los microorganismos metanogénicos. Entre los más conocidos están el nitrógeno amoniacal, los ácidos grasos de cadena larga, ácidos grasos volátiles, algunos cationes, etc. No todos los grupos de metanogénicos resultan igualmente inhibidos por los mismos compuestos. La inhibición por amoníaco libre es más fuerte para los metanogénicos acetoclásticos que para los hidrogenotróficos (Hansen et al., 1998). Optimización de la digestión anaerobia de purines de cerdo 47 Tabla 2.8. Resumen de parámetros cinéticos de la fase metanogénica (adaptado de Pavlostathis y Giraldo-Gómez, 1991) Tipo cultivo Tª k Ks μmax b Y -1 -1 d d gDQO mgDQO gSSV gSSV ·d L gDQO Metanogénesis acetoclástica (substrato acetato en todos los casos) Cultivo mezcla 25 5.0 930 0.25 0.050 0.011 Cultivo mezcla 30 5.1 356 0.275 0.054 0.037 Cultivo mezcla 35 8.7 165 0.357 0.041 0.015 Cultivo mezcla 60 26 0.28 Methanosarcina barkeri 37 8.6 257 0.206 0.024 0.004 Methanobacterium sp. 30 26 11 0.26 0.01 Methanothrix soehngenii 37 30 0.11 0.023 Metanogénesis hidrogenotrófica a partir de H2 y CO2 Methanobrevibacter 33 0.6 1.4 0.04 arboriphilicus Methanobrevibacter smithii 37 90 0.018 4.02 0.045 0.088 Methanobrevibacter smithii 37 Bacterias del rumen 37 2-8 0.016 Lodo de digestor 30 11-69 0.07-0.11 60 50-54 0.09-0.14 0.13 Methanobacterium thermoautotrophicum Methanospirillum hungatei 37 1.92 0.09-0.12 0.05 0.017-0.03 Cultivo mezcla 35 16.5 4.8*10-5 A modo de resumen, se puede decir que, a excepción del paso hidrolítico, los demás procesos pueden ser simulados suficientemente bien utilizando la cinética de Monod. Un resumen de los parámetros cinéticos, adaptado de Pavlosthatis et al. (1991), se muestra en la Tabla 2.9. Es de destacar la gran variación de los parámetros, debido a la gran variabilidad en el modo de operación, condiciones ambientales y operacionales. 2.5. Parámetros ambientales y de control 2.5.1. Temperatura De forma general, a altas temperaturas las tasas de reacción químicas y biológicas son más rápidas que a bajas temperaturas. La velocidad de reacción de los procesos biológicos dependen de la velocidad de crecimiento de los microorganismos responsables, que a su vez es dependiente de la temperatura (van Lier, 1995). 48 El proceso de la digestión anaerobia Tabla 2.9. Resumen de constantes cinéticas propuestas para los diferentes procesos (Pavlosthatis et al., 1991) k Ks b μmax Y -1 -1 d Substrato Proceso d mgDQO gDQO gSSV L gSSV ·d gDQO Carbohidratos Acidogénesis 1.33-0.76 22.5-630 7.2-30 0.14-0.17 6.1 AGCL Oxidación anaerobia 0.77-6.76 105-3180 0.085-0.55 0.04-0.11 0.01-0.015 AGV Acetogénesis 6.2-17.1 12-500 0.13-1.20 0.025-0.047 0.01-0.027 Acetato Metanogénesis acetoclástica 2.6-11.6 11-421 0.08-0.7 0.01-0.054 0.004-0.037 H2/CO2 Metanogénesis hidrogenotrófica 1.92-90 4.8*10-5-0.60 0.05-4.07 0.017-0.045 0.088 Influencia de la temperatura sobre aspectos físico-químicos La solubilidad de los gases NH3, H2S y H2 desciende al aumentar la temperatura, favoreciéndose la transferencia líquido-gas, y por tanto desapareciendo más rápidamente del medio acuoso. Esto supone un efecto positivo, dada la toxicidad sobre el crecimiento de los microorganismos anaerobios de los citados compuestos. Una posible desventaja de este fenómeno es que el descenso de la solubilidad del CO2, que implicará un aumento del pH en los reactores termofílicos, lo que en condiciones de alta concentración de amonio puede ser negativo (van Lier, 1995). La solubilidad de la mayoría de las sales aumenta con la temperatura. Las sales orgánicas son más solubles a altas temperaturas, por lo que la materia orgánica es más accesible para los microorganismos, y aumenta la velocidad del proceso. Sin embargo, si se trata de compuestos tóxicos, al aumentar su solubilidad con la temperatura serán potencialmente más tóxicos, lo que puede explicar parcialmente la mayor inhibición de determinados compuestos orgánicos en el rango termofílico, como los ácidos grasos de cadena larga (Hwu et al., 1997). Además, la temperatura influye directamente en determinados equilibrios químicos, con gran influencia sobre el proceso anaerobio, como los del amonio-amoníaco libre o ácidos grasos volátiles ionizados-no ionizados. En general, con la temperatura se favorecen las formas no ionizadas, que resultan más tóxicas para los microorganismos (NH3 y AH). Optimización de la digestión anaerobia de purines de cerdo 49 La viscosidad de los líquidos y semisólidos disminuye al aumentar la temperatura, lo que implica menores requerimientos energéticos para la mezcla (agitación). A altas temperaturas se produce también una mejor sedimentación de los sólidos. Influencia de la temperatura sobre aspectos bioquímicos Tasa de crecimiento de metanogénicos (%) El proceso anaerobio se produce en la naturaleza en un amplio rango de temperaturas, que van desde 0º a 97ºC (Muñoz-Valero et al., 1987). La eficiencia del proceso, no obstante, es muy diferente en función de la temperatura del medio. Se habla de tres rangos principales de temperatura, psicrofílico (por debajo de 25ºC), mesofílico (entre 25 y 45ºC) y termofílico (entre 45ºC y 65ºC), siendo la tasa máxima específica de crecimiento (Pmax) mayor conforme aumenta la temperatura (van Lier et al., 1993). Dentro de cada rango de temperatura, existe un intervalo en que dicho parámetro se hace máximo. Será interesante, por tanto, trabajar en torno a este punto (Figura 2.6). La velocidad del proceso aumenta con la temperatura, aunque también aumentan los requerimientos energéticos, y puede disminuir la estabilidad del proceso (Fannin, 1987), al menos en presencia de determinados tóxicos. Por otro lado, es preciso desarrollar un completo balance energético para establecer el interés de mantener una determinada temperatura. El rango psicrofílico se plantea como poco viable debido al gran tamaño de reactor necesario. Sin embargo, simplifica mucho el diseño y hay menos problemas de estabilidad. Cuanto mayor es la duración del tiempo de retención menor es la diferencia entre las velocidades de degradación a diferentes temperaturas (Fannin, 1987). 100 Termofílico 80 60 40 Mesofílicos Psicrofílicos 20 0 20 40 60 80 Temperatura (ºC) Figura 2.6. Dependencia de la constante de crecimiento de la temperatura (van Lier et al. , 1993). 50 El proceso de la digestión anaerobia La temperatura más utilizada en la digestión anaerobia de residuos es dentro del rango mesofílico, alrededor de 35-37ºC, aunque hay cierta tendencia en los últimos años a pasar al rango termofílico tanto para conseguir una mayor velocidad del proceso, como para mejorar la destrucción de organismos patógenos. La producción de biogás, en ausencia de inhibidores, aumenta con la temperatura, puesto que aumenta la tasa de crecimiento de los microorganismos; temperaturas más bajas implican tiempos de retención más largos, y por tanto mayores volúmenes de reactor. La tasa de hidrólisis también aumenta con la temperatura (Veeken y Hamelers, 1999), por lo que el régimen termofílico puede tener gran interés al tratar residuos en los que la hidrólisis sea la etapa limitante, como los residuos con alto contenido en componentes lignocelulósicos. El régimen termofílico se ha relacionado tradicionalmente con mayores problemas de estabilidad (Hobson,1990). Sin embargo otros autores consideran que las plantas termofílicas son tan estables y tan operables como las mesofílicas, presentando, además de las ventajas antes mencionadas, una mayor producción de gas por unidad de sólidos volátiles y una mejora en el postratamiento, ya que el efluente de la digestión termofílica es más fácilmente deshidratable, junto con una menor producción de malos olores (Ahring, 1995; Krugel et al., 1998). La temperatura óptima para el crecimiento bacteriano depende de cada especie, tal y como se muestra en la Tabla 2.10. La mayoría de las bacterias termofílicas presentan tasas específicas de crecimiento máximas mayores que los organismos mesofílicos. La temperatura normal de operación dentro del rango termofílico está sobre los 55ºC (52-56ºC). Por encima de este nivel los microorganismos acetogénicos disminuyen drásticamente su velocidad de crecimiento. Sin embargo, la tasa específica de crecimiento de los microorganismos metanogénicos continua aumentando hasta los 70ºC (Ahring, 1995). La sensibilidad a los cambios de temperatura ambiental depende de diversos factores, principalmente del grado de adaptación del cultivo, del modo de operación y del tipo de bioreactor. En el rango termofílico un aumento brusco de la temperatura puede provocar un importante descenso en la producción de gas (van Lier et al., 1993), mientras que una bajada puede suponer un descenso en la producción de gas, pero completamente reversible (van Lier et al., 1993), o puede no mostrar diferencias (Ahring et al., 1995), debido a la disminución del efecto de inhibición por amoníaco. Para pasar un reactor del rango mesofílico al termofílico sin que se produzca una Optimización de la digestión anaerobia de purines de cerdo 51 parada en la producción de gas, se deberá realizar muy lentamente (van Lier et al., 1993). Tabla 2.10. Temperatura óptima y máxima, parámetros cinéticos de crecimiento para diferentes cultivos metanogénicos acetoclásticos (van Lier et al., 1993) μmax Ks(Ac) Metanogénicos acetoclásticos Topt (ºC) Tmax (ºC) (d-1) mg DQO/L Methanosarcina barkeri 35-40 0.023 320 Methanosarcina thermophila 50 55-60 0.058 288 Methanosarcina CALS-1 55-58 60 0.058 Methanosarcina MP 55 60 Methanosarcina MSTA-1 55 65 0.053 685 Methanosarcina CHTI55 57 63 0.085 614 Methanothirx soehngenii 37 45-50 0.0085 45 Methanothirx conciliii 35-40 40-45 0.029 77 55 65-70 0.020 Methanosaeta sp. PT TAM 60 70 0.012 51 Methanothirx sp. CALS-1 60 65-70 0.028 <64 Methanothirx thermoacetophila 65 70 Co-cultivo oxidante de acetato 60 0.019 - El efecto inhibidor del amonio es mayor en el rango termofílico que en el mesofílico por el aumento de la concentración de la forma tóxica, NH3, al aumentar la temperatura (Angelidaki y Ahring, 1994 y Hansen et al.,1998), a pesar de la mayor sensibilidad de los microorganismos mesofílicos al amoníaco libre (Gallert et al., 1998). Un problema adicional al de inhibición por amonio es la mayor tasa de hidrólisis de proteínas en el rango termofílico frente al mesofílico (Gallert et al., 1998). El tratamiento termofílico presenta la importante ventaja de la mayor eliminación de organismos patógenos, que puede ser un factor clave en función del destino final del efluente, sobre todo para su uso como fertilizante orgánico. Un tratamiento termofílico por encima de 50ºC reúne en un sólo paso el tratamiento de higienización y el de digestión anaerobia (Angelidaki y Ahring, 1997). El proceso de digestión anaerobia produce la inactivación de algunos virus patógenos (enterovirus y parvovirus), pero la tasa de inactivación depende del tipo de virus, de la duración del proceso y de la temperatura de operación (Turner y Burton, 1997). Sobre lodos de depuradora se han realizado numerosos estudios de comparación de la eficiencia de eliminación de patógenos en función de la temperatura de tratamiento, resultando generalmente muy favorable el rango termofílico 52 El proceso de la digestión anaerobia (Watanabe et al., 1997, Krugel et al., 1998). Una técnica interesante, especialmente en instalaciones ya existentes de digestión de lodos de depuradora, es la combinación de dos fases de digestión, una primera de alta carga en termofílico y una segunda con menor carga en mesofílico. Con este sistema aprovechan las ventajas del sistema termofílico (reducción de patógenos, menos problemas de formación de espumas, foaming) y evitan posibles problemas de inestabilidad (Han et al., 1997; Oles et al., 1997). 2.5.2. Contenido de nutrientes El proceso anaerobio se caracteriza, frente a procesos aerobios, por los bajos requerimientos de nutrientes, debido fundamentalmente a los bajos índices de producción de biomasa. A pesar de ello, la biomasa necesita para su desarrollo el suministro de una serie de nutrientes minerales, además de una fuente de carbono y de energía. Para determinados procesos necesita, además, compuestos orgánicos especiales, como vitaminas. Los principales nutrientes del sistema anaerobio son nitrógeno, sulfuro, fósforo, hierro, cobalto, níquel, molibdeno, selenio, riboflavina y vitamina B12 (Speece, 1987a). Los valores mínimos necesarios para el correcto crecimiento de los microorganismos se muestra en la Tabla 2.11. Otros autores han expresado las necesidades de nitrógeno y fósforo en función de la concentración de carbono de la alimentación, considerándose que la relación C/N debe oscilar entre 15-30:1, y la C/P de 75-113/1 (Speece, 1987a). Tabla 2.11. Rangos de concentración de nutrientes, necesarios para el correcto crecimiento de las bacterias anaerobias (Henze, 1995) g/kg SSV g/kg DQO (B) Nitrógeno 80-120 55-85 Fósforo 10-25 7-18 Azufre 10-25 7-18 Hierro 5-15 4-11 En general, los residuos ganaderos suministran una suficiente concentración de todos los nutrientes, siendo más común la presencia de problemas por exceso que por defecto. Optimización de la digestión anaerobia de purines de cerdo 53 2.5.3. Velocidad de carga orgánica (VCO) y tiempo de retención hidráulico (TRH) El tiempo de retención es un parámetro muy importante, que dependerá típicamente del tipo de reactor utilizado. En los sistemas de mezcla completa el tiempo de retención hidráulico coincide con el tiempo de retención celular, es decir de la biomasa, por lo que el tiempo de retención deberá ser suficientemente largo para permitir el desarrollo de la población bacteriana. El tiempo de retención, junto con la velocidad de carga, determinada por el tipo de substrato, son los principales parámetros de diseño, definiendo el volumen del digestor. La fracción de materia orgánica degradada aumenta al aumentar el TRH, sin embargo la producción volumétrica de metano (producción por unidad de reactor) disminuye, una vez superado el óptimo. Es por tanto necesario determinar para cada tipo de residuo y de digestor el tiempo de retención que optimiza el proceso. Los tiempos de retención usuales tratando residuos ganaderos varían mucho según la fuente consultada y van de 10 a 30 días (Hobson, 1990). La carga orgánica es la relación es la cantidad de materia orgánica, expresada normalmente en unidades de DQO o de sólidos volátiles, por unidad de reactor y unidad de tiempo, siendo directamente dependiente de la concentración del substrato y del tiempo de retención. Altas cargas orgánicas, en ausencia de inhibidores, proporcionan altas producciones volumétricas de biogás. Parece que la resistencia a ciertos inhibidores puede aumentar con la carga orgánica (Angelidaki et al., 1993). Sin embargo la inestabilidad aumenta también con el aumento de carga, especialmente en el caso de “sobrecargas” puntuales, que conllevan la acumulación de ácidos grasos volátiles (Ahring et al., 1995). 2.5.4. Agitación La agitación de los reactores anaerobios tiene diversos objetivos, que se resumen en los siguientes puntos (Noone, 1990): poner en contacto el substrato fresco o influente con la población bacteriana, y eliminar los metabolitos producidos por los metanogénicos, al favorecer la salida de los gases; proporcionar una densidad uniforme de población bacteriana; prevenir la formación de capa superficial y de espumas, así como la sedimentación en el reactor; prevenir la formación de espacios muertos que reducirían el volumen efectivo del reactor, y la formación de caminos preferenciales en función de la hidráulica del sistema; eliminar la estratificación térmica, manteniendo una temperatura uniforme en todo el reactor. 54 El proceso de la digestión anaerobia Algunos tipos de reactores pueden funcionar bien sin ningún sistema de agitación. Se suelen utilizar para substratos con muy alto contenido en sólidos o sobre substratos básicamente solubles, con regímenes de flujo tipo pistón. La agitación puede ser de varios tipos, mecánica, hidráulica o neumática. Para grandes tamaños parece que la agitación por gas es la que mayores ventajas presenta, tanto por el efecto de agitación, como por su sencillez de diseño y operación (Muñoz Valero et al., 1987). La velocidad de agitación es un parámetro que puede influir en el desarrollo del proceso, siendo necesario un equilibrio entre la buena homogeneización y la correcta formación de agregados bacterianos (Fannin, 1987). Una velocidad de agitación alta, por encima de 700 rpm, puede disminuir ligeramente la producción de biogás (Stafford, 1982), por ruptura de agregados bacterianos. 2.5.5. pH y alcalinidad Los microorganismos anaerobios necesitan un pH en torno a la neutralidad para su correcto desarrollo, aunque permiten cierta oscilación (Clark y Speece, 1989). Parece ser que el pH afecta fundamentalmente a la actividad enzimática de los microorganismos (Webb, J.L., 1963, citado en Clark y Speece, 1989), mediante: cambios de estado de los grupos ionizables de las enzimas como el carboxil y amino; alteración de los componentes no ionizables del sistema, como por ejemplo el substrato; y desnaturalización de la estructura proteica de las enzimas. Para que el proceso se desarrolle de forma satisfactoria, el pH debe estar en torno a la neutralidad, presentando problemas graves si el pH baja por debajo de 6 o sube por encima de 8,3 (Lay et al., 1997). Sin embargo, el proceso de inhibición parece ser completamente reversible, aunque el tiempo de recuperación depende de la duración de la alteración. El pH es también una importante variable de diagnóstico de los sistemas anaerobios, pues muchos fenómenos tienen influencia sobre el mismo. Ejemplos clásicos son las sobrecargas orgánicas, o la presencia de un inhibidor de la etapa metanogénica, que pueden provocar desequilibrios entre la producción y el consumo de ácidos grasos volátiles, produciendo la acumulación de éstos y el consiguiente descenso del pH, produciéndose la acidificación del reactor. En función de la alcalinidad del medio, la bajada de pH será más o menos rápida. En residuos ganaderos, que presentan altas alcalinidades, la bajada de pH será poco importante incluso aunque se produzcan importantes acumulaciones de ácidos. Optimización de la digestión anaerobia de purines de cerdo 55 En cualquier caso, el pH no se considera una buena variable de control por resultar demasiado lenta: una vez detectada una variación importante del pH, el fracaso del sistema puede ser ya irreversible (Iza, 1995). Por ello se consideran otras variables como mejores indicadores del estado del proceso anaerobio, como la producción de biogás y su contenido en metano (Hill y Holmberg, 1988), el contenido de ácidos grasos volátiles o la relación entre ellos (Hill et al., 1987; Ahring et al., 1995), la presión parcial de hidrógeno, o indicadores basados en el número de bacterias o actividad bacteriana (Angelidaki et al., 1997b). Por otro lado, el pH es un importante modulador del sistema puesto que influye en varios equilibrios químicos, pudiendo desplazarlos hacia la formación de una determinada componente que tenga influencia en el proceso. Su papel es fundamental en el equilibrio amonio - amoníaco, teniendo, por tanto, una gran importancia en el proceso general, por ser el amoniaco libre un importante inhibidor de la fase metanogénica (Zeeman et al., 1985). El pH influye también en el mecanismo de inhibición de degradación de propionato por acético, habiéndose descrito una mayor inhibición a pH bajos (Fukuzaki et al., 1990), debido a que, en este caso, el componente tóxico es la forma no ionizada del ácido acético, que aumenta con la acidez del medio. La alcalinidad es una medida de la capacidad tampón del medio. Esta capacidad tampón puede ser proporcionada por un amplio rango de sustancias, siendo por tanto una medida inespecífica. En el rango de pH de 6 a 8, el principal equilibrio químico que controla la alcalinidad es el dióxido de carbono-bicarbonato. La relación de alcalinidad RA, se define como la relación entre la alcalinidad debida a los AGV (AI) y la debida al bicarbonato (AT), recomendándose no sobrepasar un valor de 0,3-0,4 para evitar la acidificación del reactor (Iza, 1995). La alcalinidad al bicarbonato debe mantenerse por encima de 2500 mg/L para asegurar la estabilidad del digestor (Fannin, 1987). 2.5.6. Tóxicos e inhibidores La magnitud de toxicidad observada o recogida en la bibliografía es una función de diversos factores, incluyendo concentración, antagonismos, sinergismos, formación de complejos y aclimatación (Kugelman y Chin, 1971). La concentración es el único factor que usualmente se considera, lo que lleva a afirmaciones absolutistas, muchas veces erróneas. En general la velocidad de crecimiento bacteriano aumenta con la 56 El proceso de la digestión anaerobia concentración de substrato, llegando a un punto en que se estabiliza y, dependiendo de cada caso concreto, puede llegar a descender (inhibición por el substrato). Así, en términos absolutos, una substancia es un tóxico o un substrato dependiendo de su concentración. Los fenómenos de antagonismo y sinergismo son muy importantes al hablar de toxicidad. Antagonismo es una reducción de la toxicidad de un substrato en presencia de otro y sinergismo es el aumento del efecto tóxico de una substancia causada por la presencia de otra. La formación de complejos resulta, también, fundamental. Si una sustancia no está en solución, no puede penetrar dentro de la célula, y por tanto no podrá afectar el metabolismo del organismo. La magnitud del efecto tóxico de una substancia puede ser reducido significativamente por aclimatación de la población de microorganismos al tóxico. La aclimatación implica una reorganización de los recursos metabólicos para vencer los obstáculos metabólicos producidos por el substrato tóxico, más que mutación o selección de las poblaciones (Kugelman y Chin, 1971). Otros factores pueden afectar también la toxicidad de un determinado compuesto, por ejemplo, el tipo de agregados bacterianos, siendo más resistentes, en general, los lodos granulares que los floculentos (Hwu et al., 1997). También la temperatura juega un importante papel en el efecto tóxico de determinados compuestos (amonio, sulfuro, ácidos grasos volátiles, etc.). Son muchas las substancias que pueden resultar inhibidoras del crecimiento de los microorganismos anaerobios. A continuación se describe brevemente los compuestos que más comúnmente presentan problemas de toxicidad, en los substratos utilizados en el presente trabajo. Nitrógeno amoniacal Los residuos ganaderos contienen altas concentraciones de compuestos nitrogenadas, función del sistema de alimentación, de la composición de los piensos, del tipo de animales de los que procede, así como del tipo de granja. El nitrógeno orgánico durante el proceso anaerobio se hidroliza produciendo formas amoniacales. Aunque el nitrógeno amoniacal es un importante nutriente para el crecimiento de los microorganismos (Bryant et al., 1971), cuya carencia puede provocar el fracaso en la producción de gas, una concentración excesivamente alta del mismo puede limitar su crecimiento. Hay una gran dispersión en la bibliografía sobre la concentración de amonio inhibidora del proceso anaerobio. Así, a 50ºC una concentración de 1,7 g N-NH4+/L resultó inhibitoria para la digestión anaerobia de estiércol bovino (Zeeman et al., 1985). Hashimoto (1986), en reactores sin aclimatar, encontró signos de inhibición a una concentración de nitrógeno amoniacal Optimización de la digestión anaerobia de purines de cerdo 57 de 2,5 g N-NH4+/L, tanto en mesofílico como termofílico, aunque al trabajar con reactores adaptados previamente la concentración inicial de inhibición fue de 4 g N-NH4+/L en el rango termofílico. Koster y Lettinga (1988) consiguieron que el proceso se desarrollara a concentraciones de amonio extremadamente altas, hasta 12 g N-NH4+/L con un lodo granular, aunque a partir de 2,5 g N-NH4+/L se observó una disminución en la tasa específica máxima de crecimiento de los microorganismos metanogénicos, destacando la reversibilidad de la toxicidad por amonio, así como que la alta concentración de amonio apenas afecta la acidogénesis. Robbins et al. (1989) a niveles de 2,8 g N-NH4+/L no observaron inhibición, al menos de manera permanente, siendo el cultivo capaz de recuperarse al poco tiempo. Angelidaki y Ahring (1993) concluyeron que aunque es posible mantener un proceso estable con concentraciones de amonio por encima de 6 g NNH4+/L en el rango termofílico, a partir de esta concentración el volumen de gas producido es afectado seriamente. Krylova et al. (1997) trabajando con gallinaza encontraron que una concentración de amonio superior a 2,8 g NNH4+/L suponía un descenso en el biogás producido de entre el 50 y 90% y del metano entre el 80 y el 90%. Hansen et al. (1998) obtuvieron un proceso de producción de biogás estable con purines de cerdo a una concentración de amonio de 6 g N-NH4+/L, pero los índices de producción de biogás fueron menores que el potencial y además disminuyeron de forma importante al aumentar la temperatura y por tanto al aumentar el amonio libre. Flotats et al. (1999) observaron el efecto inhibidor del amonio sobre la producción de gas, a partir de una concentración de 2,1 g N-NH4+/L en el rango termofílico. La carga orgánica del reactor, y por tanto el número de microorganismos activos, parece afectar la inhibición por amonio (Ahring, 1995), lo que puede explicar en parte la variabilidad encontrada. Van Velsen (1979) demostró que la adaptación del lodo metanogénico hace viable la digestión anaerobia en régimen mesofílico a concentraciones de 3 g N-NH4+/L para purines de cerdo y hasta 5 g N-NH4+/L para lodos de depuradora. A pesar de la adaptación observada, la velocidad específica máxima de crecimiento desciende al aumentar la concentración de nitrógeno amoniacal. El efecto de adaptación de los microorganismos ha sido constatado por diversos autores (Hashimoto, 1986; Koster, 1986; Angelidaki y Ahring, 1993b; 1994; Hansen et al., 1998; Gallert et al., 1998). La forma que parece causar la inhibición por amonio es el amoníaco libre (NH3) ya que el efecto inhibitorio del amonio parece aumentar a pH alcalinos y a altas temperaturas (Zeeman et al., 1985). Los límites de inhibición de nuevo varían mucho según el autor. Hashimoto (1986) 58 El proceso de la digestión anaerobia encontró inhibición a una concentración de 0,02 g N-NH3/L en el rango mesofílico, 0,2 g N-NH3/L en el rango termofílico, sin aclimatación y 0,39 g N-NH3/L en el rango termofílico con aclimatación. Angelidaki y Ahring (1993b) observaron que con una concentración de 0,650 g N-NH3/L la velocidad de crecimiento de los metanogénicos a partir de acético disminuye un 20%. Hansen et al. (1998), trabajando con purines de cerdo, observaron inhibición sólo a partir de la concentración 1,1 g N-NH3/L, disminuyendo acusadamente la tasa de crecimiento específico. Este valor de la concentración de inhibición tan alto se debe a la larga adaptación previa del inóculo utilizado. Los principales microorganismos afectados por altas concentraciones de amonio son los metanogénicos. Un cambio brusco en la concentración de amonio produce un descenso en la velocidad de crecimiento de los organismos metanogénicos, pero no en la tasa de crecimiento de los acidogénicos o acetogénicos (Koster y Lettinga, 1988; Robbins et al., 1989). Los microorganismos metanogénicos que consumen acético son más sensibles a la inhibición por amonio que los consumidores de H2 (Angelidaki y Ahring, 1993b, Hansen et al., 1998). La función de inhibición que relaciona la tasa específica de crecimiento con la concentración de amoníaco libre es función del tipo de microorganismos, siguiendo para los metanogénicos acetoclásticos un modelo sigmoidal, mientras que los que consumen H2 siguen un modelo lineal (Angelidaki et al., 1993). La consecuencia del modelo sigmoidal es que en un rango relativamente estrecho de concentraciones de amoníaco libre la velocidad de crecimiento desciende bruscamente, manteniéndose, sin embargo, más estable a concentraciones más altas. Una concentración de amonio superior a los 7-8 g N-NH4+/L puede inhibir la hidrólisis de proteínas (Krylova et al., 1997). Gallert et al. (1998) observaron que al aumentar la concentración de 0,5 a 6,5 g N-NH4+/L en el régimen mesofílico aumentó la inhibición sobre la desaminación de peptonas (proteolisis) y la metanogénesis, y sin embargo no observaron acumulación de hidrógeno, indicando que la metanogénesis hidrogenotrófica no resulta inhibida a este nivel de concentración. Algunos autores han apuntado diferencias de comportamiento frente a la inhibición por amonio de microorganismos mesofílicos o termofílicos. Aunque normalmente se considera más problemático el proceso termofílico, puesto que el agente inhibidor es el amoníaco libre, Gallert et al. (1998) encontraron que los microorganismos mesofílicos son más sensibles a la inhibición por NH3, con un valor de la constante de inhibición (considerando inhibición no competitiva reversible) de los metanogénicos, Optimización de la digestión anaerobia de purines de cerdo 59 de 92 mg NH3/L, mientras que en termofílico el valor de esta constante se cifró 251 mg NH3/L; también encontraron que la degradabilidad de las proteínas (peptona) es mayor en termofílico que en mesofílico. Ácidos grasos volátiles Los ácidos grasos volátiles son los más importantes intermediarios del proceso anaerobio, siendo, por ello, fundamental conocer su evolución. Juegan un papel muy importante en la monitorización y control de reactores anaerobios, mostrando una rápida respuesta a las variaciones en el sistema, por ejemplo en el caso de sobrecargas orgánicas (Ahring et al., 1995), o en el caso de la introducción de tóxicos. El aumento de su concentración está relacionado con la disminución en la producción de biogás (Hill et al., 1987). Además, los ácidos grasos volátiles pueden inhibir algunos de los procesos que tienen lugar en un reactor anaerobio, aunque hay cierta dispersión en la bibliografía. La acumulación de propiónico en el reactor, especialmente de la forma no ionizada, puede inhibir la acetogénesis a partir de propiónico (Fukuzaki et al., 1990), y la metanogénesis acetoclástica (Barredo y Evison, 1991). La acumulación de acético, puede inhibir la acetogénesis a partir de propiónico (Fukuzaki et al., 1990), y la acetogénesis a partir de butírico (Ahring y Westermann, 1988). Sin embargo, son necesarias concentraciones de acético muy altas para que llegue a afectar a la producción de metano, por encima de 4000 mg/L o más (Stafford, 1982; Ahring et al., 1995). Ahring et al. (1995) concluyeron que concentraciones de ácidos grasos volátiles por debajo de 50 mM, equivalente a 3000 mg acético/L, no producen ninguna disminución de la producción de metano. Son los ácidos propiónico y valérico los primeros que afectan al proceso, mientras que el butírico y el acético han de acumularse por encima de 100 mM para afectar a la tasa de producción de metano. Hidrógeno El hidrógeno es un importante intermediario del proceso anaerobio, tal y como se vio en el apartado 1.5.4, y su acumulación puede provocar la inhibición de la acetogénesis, con la consiguiente acumulación de ácidos grasos volátiles, estando especialmente descrita la acumulación de propiónico (Harper y Pohland, 1986; Boone y Xun, 1987; Fukuzaki et al, 1990). Hill y Cobb (1993) relacionaron altos valores de la presión parcial de H2 con el aumento de la fracción iso sobre la fracción n de los ácidos butírico y valérico. 60 El proceso de la digestión anaerobia Compuestos azufrados en los sistemas anaerobios En presencia de sulfatos las bacterias metanogénicas compiten con las bacterias sulfato-reductoras por los substratos útiles, mostrando las últimas, ventajas termodinámicas y cinéticas sobre las primeras, tanto sobre las que consumen hidrógeno como sobre las acetoclásticas (Hulshoff Pol et al., 1998). El resultado de esta competición determinará la proporción de sulfhídrico y metano en el biogás producido. El sulfato es, además, un importante inhibidor, aumentando el efecto inhibidor en función de la relación DQO/sulfato, de forma que los substratos adaptados no muestran inhibición para valores de la relación por encima de 10, y sí hay signos de inhibición por debajo de un valor de 7-8, aunque es posible mantener un proceso estable (Omil et al., 1995). Además de la competición, el sulfíhidrico es tóxico a altas concentraciones para muchos grupos bacterianos. Parece que la forma tóxica es la no ionizada, ya que es la que puede atravesar la membrana celular, por lo que la inhibición se ve favorecida a pH bajos y a bajas temperaturas (predominio de la forma no ionizada y mayor solubilidad en la fase líquida). En general el lodo granular es menos sensible que la biomasa en suspensión a la inhibición por H2S a pH bajos y neutros, aunque parece ser similar a altos pH. A pH altos la inhibición de las metanogénicas es mayor que la de las sulfatoreductoras, mientras que a bajos pH no hay diferencia entre ambos grupos. En general, los metanogénicos son más sensibles que los acetogénicos y los acidogénicos. A pH de 7,5 a 9, la inhibición de las sulfato-reductoras acetoclásticas se determina por la concentración de sulfuros totales, más que por la concentración de H2S. Además del pH, la relación DQO/sulfato influye en la sensibilidad de un lodo, debido a las diferentes asociaciones bacterianas, así como la temperatura (Hulshoff Pol et al., 1998). A altas temperaturas se favorece el paso de H2S del líquido al gas al disminuir la solubilidad de éste, por lo que, en principio, habrá menos problemas en el rango termofílico. Los niveles de concentración de H2S a los cuales se produce la inhibición al 50% varían en función de los citados parámetros, pero están entre 50 y 250 mg/L. Sin embargo, concentraciones más bajas, 23 mg S/L en el rango termofílico, pueden producir inhibición del proceso metanogénico si se digiere un material con alto contenido en nitrógeno amoniacal, por ejemplo purín de cerdo (Hansen et al., 1999). Ácidos grasos de cadena larga Altas concentraciones de ácidos grasos de cadena larga pueden inhibir el proceso de digestión anaerobia (Galbraith et al., 1971; Hanaki et al., 1981; Optimización de la digestión anaerobia de purines de cerdo 61 Angelidaki et al., 1990; Angelidaki y Ahring, 1992; Rinzema et al., 1994; Hwu et al., 1997). Las grasas neutras (triglicéridos) son hidrolizadas rápidamente a ácidos grasos de cadena larga (AGCL). Las concentraciones límite de inhibición no están muy claras en la bibliografía y depende mucho del tipo de ácido graso, así como de la forma en que se encuentra. El efecto inhibidor de los ácidos grasos de cadena larga provoca un aumento de la duración de la fase lag en ensayos en discontinuo (Hanaki et al., 1981). Aunque la adsorción de los AGCL sobre la pared celular puede jugar un papel importante sobre la inhibición, ésta se ha relacionado con la concentración de AGCL, más que con la relación de ácidos grasos/biomasa (Koster y Cramer, 1987; Angeldaki y Ahring, 1992). El efecto tóxico se ha descrito como no reversible, y la forma tóxica son los ácidos grasos libres. El efecto inhibidor de los lípidos está muy relacionado con la adaptación de los microorganismos, y prácticamente condicionados a la existencia de microorganismos acetogénicos que degraden AGCL a medida que se van produciendo por hidrólisis de las grasas (triglicéridos u otras formas), evitando así, alcanzar concentraciones tóxicas (Angeldaki y Ahring,1992, Rinzema et al., 1994). Según la bibliografía, la toxicidad de los ácidos grasos de cadena larga, en especial del oleico, es mayor en el rango termofílico que en el mesofílico, estando también afectada por el tipo de lodo (granular o floculento) (Hwu et al., 1997). Angeldaki y Ahring concluyeron que los ácidos grasos libres de cadena larga, oleico y estérico, inhiben todos los pasos de la digestión anaerobia termofílica, provocando, a una concentración de 0,2 g/L de oleico, el aumento en la duración del desfase inicia en la producción de metano, fase lag, mientras que el crecimiento bacteriano es completamente inhibido a una concentración de 0,5 g/L de oleico y 1,0 de esteárico (Angeldaki y Ahring, 1992). En presencia de calcio el efecto tóxico de los ácidos grasos de cadena larga disminuye debido a la precipitación de las sales cálcicas (Galbraith et al., 1971; Koster, 1987; Angelidaki et al., 1990). Cationes y metales pesados Todos los cationes pueden proporcionar toxicidad a algún nivel de concentración, aumentando la toxicidad con el peso molecular, por lo que los metales pesados son los que provocan toxicidad a menor concentración, (Hayes y Theis, 1978). El orden de toxicidad de los metales pesados es Ni > Cu >> Cr(IV) # Cr(III) > Pb > Zn (Hayes y Theis, 1978). Los niveles de inhibición varían 62 El proceso de la digestión anaerobia mucho en función de la fuente, debido a varios factores. En primer lugar la toxicidad es menor si la introducción en el reactor es gradual (Tabla 2.12). Los metales pesados precipitan en presencia de sulfuros, desapareciendo de la solución, por lo que resultan menos tóxicos para los microorganismos, pudiendo llegar a tolerarse elevadas concentraciones de metales pesados (Kugelman y Chin, 1971). Tabla 2.12. Concentración de inhibición y toxicidad de metales pesados (Hayes y Theis, 1978) Alimentación gradual Alimentación brusca Límite de toxicidad Metal Concentración de Límite de toxicidad (mg/L) Inhibición* (mg/L) (mg/L) Cr(III) 130 260 <200 Cr(VI) 110 420 <180 Cu 40 70 <50 Ni 10 30 <30 Cd >20 >10 Pb 340 >340 >250 Zn 400 600 <1700 * Inicio de la disminución de la producción de gas Otros cationes como calcio, sodio, potasio, etc., pueden resultar inhibidores para el proceso anaerobio, a concentraciones altas (Kugelman y Chin, 1971; Omil et al., 1995; Kim et al., 1999). La concentración de inhibición por cationes depende mucho de la presencia de posibles antagonistas, tal y como se muestra en laTabla 2.13. El potasio es antagonista del sodio, del magnesio y del calcio; el sodio lo es del amonio, potasio, magnesio y calcio; el calcio del potasio; el magnesio del potasio y el amonio del potasio (Kugelman y Chin, 1971). Tabla 2.13. Concentración límite de cationes en sistemas anaerobios (Kugelman y Chin, 1971). Alimentación sencilla Alimentación continua Cation simple En presencia de Cation simple En presencia de Cation (M) antagónicos (M) (M) antagónicos (M) Sodio 0.2 0.3-0.35 0.3 70.35 Potasio 0.09 0.15-0.2 0.13 0.35 Calcio 0.07 0.125-0.15 0.15 0.2 Magnesio 0.05 0.125 0.065 0.14 Optimización de la digestión anaerobia de purines de cerdo 63 Desinfectantes y antibióticos Los antibióticos son comúnmente añadidos a las dietas de los animales, para mejorar la producción o para control de enfermedades. La presencia de penicilina y tetraciclina, procedentes de la dieta de los animales, en el purín de cerdo, tiene un efecto inhibitorio sobre el proceso, aunque parece haber una buena aclimatación a la presencia de dichas substancias (Massé et al., 2000). Hilpert et al. (1987) encontraron que dos antibióticos, monensin y lasalocid, usados normalmente en las dietas inhiben el proceso anaerobio, aunque el segundo precisa una concentración muy alta. Sin embargo otros muchos antibióticos no han mostrado ningún efecto sobre el proceso, como flavomicin y bacitracin (Hilpert et al., 1987) y tylosin, lyncomycin, sulphamethazine y carbadox (Massé et al., 2000). Los restos de desinfectantes, procedentes de la limpieza y desinfección de las granjas en los residuos animales pueden resultar tóxicos para el sistema anaerobio (Hilpert et al., 1987). La toxicidad dependerá, básicamente de la concentración, de la biodegradabilidad de los mismos, y del tiempo transcurrido desde su utilización, hasta la entrada del residuo en el sistema anaerobio. Los residuos de cultivos agrícolas, susceptibles de ser utilizados para producción de metano, pueden contener algunos compuestos tóxicos para el crecimiento de los microorganismos anaerobios, por ejemplo, residuos de pesticidas, compuestos fenólicos, terpenos, resinas, etc. Al realizar un pretratamiento químico con ataque ácido o básico, se pueden formar, también, compuestos tóxicos (Speece, 1987b). 64 El proceso de la digestión anaerobia 3. MÉTODOS ANALÍTICOS 66 Métodos analíticos Optimización de la digestión anaerobia de purines de cerdo 67 A continuación se exponen las técnicas analíticas empleadas para la cuantificación de los parámetros físico-químicos analizados en todos los experimentos llevados a cabo, correspondientes a los capítulos 4, 5 y 6. 3.1. Sólidos totales y volátiles La determinación del contenido de sólidos totales (ST) y de sólidos volátiles (SV) se realizó de acuerdo con el método 2540 E de Standard methods for examination of water and wastewater (APHA, 1995). Los ST se determinaron mediante el peso del residuo seco, secado a 105ºC en estufa, Selecta 202, durante 48 horas, referido al peso de materia fresca inicial. Para el cálculo de se utiliza la siguiente expresión: Sólidos Totales(%) ST(%) Peso105º C Tara * 100 Peso muestra (3.1) La determinación de los sólidos volátiles (SV) se realizó sobre la misma muestra, mediante calcinación, en un mufla Heron 12-PR/300, a 550ºC durante 6 horas. El contenido en sólidos volátiles se determina por diferencia entre el residuo seco y las cenizas, siguiendo la siguiente expresión: Sólidos Volátiles(%) SV(%) Cenizas(%) Peso 105º C - Peso550º C * 100 Peso muestra Sólidos Totales (%) - Sólidos Volátiles (%) (3.2) (3.3) 3.2. Demanda química de Oxígeno (DQO) La DQO es una medida indirecta del contenido de materia orgánica y compuestos oxidables en una muestra. Se define como la cantidad de oxígeno necesaria para oxidar completamente la materia orgánica y los compuestos oxidables de una determinada muestra. Se ha utilizado el método 5220B propuesto en Standard methods for examination of water and wastewater (APHA; 1995), conocido también como método de reflujo abierto. Los resultados se expresan en mg O2/kg o g O2/kg. Se realiza la digestión de la muestra con exceso de dicromato potásico en un medio fuertemente ácido (H2SO4), durante dos horas a 150ºC. Se utiliza un bloque de digestión con capacidad para 20 tubos con reflujo abierto, marca Selecta. La reacción es catalizada por sulfato de plata (Ag2SO4) y se 68 Métodos analíticos utiliza HgSO4 para eliminar problemas de interferencias de los haluros presentes. El exceso de dicromato se valora con sal de Mohr, sulfato ferroso amónico (Fe(NH4)2(SO4)2·6H2O), usando ortofenantrolina como indicador. En cada grupo de muestras se realiza un blanco, siguiendo el mismo procedimiento que con el resto de las muestras, sustituyendo ésta por agua desionizada. Diariamente se comprueba la normalidad de la sal de Mohr utilizada. La determinación de la DQO se determina según la siguiente expresión: DQO( mgO2 / Kg ) ( V Bl Vm )* 8000 * N Peso muestra ( g ) (3.4) VBl= Volumen de Ferro-amonio sulfato consumido en la valoración del blanco. Vm= Volumen de Ferro-amonio sulfato consumido en la valoración de la muestra. N= Normalidad de la Sal de Mohr: N VolCr2 O7 * N Cr2 O7 Vol sal de mohr , donde: VolCr2O7= Volumen de Cr2O7 utilizado para la determinación de la normalidad NCr2O7= Normalidad del Cr2O7 utilizado Volsal de Mohr= Volumen de sal de Mohr utilizada para la valoración Se utiliza dicromato potásico 1 N, manteniéndose la relación volumétrica entre el dicromato y el ácido sulfúrico, de 1:3. Para la mayoría de las determinaciones de purín el volumen de dicromato fue de 10 mL, y de sulfúrico 30 mL, para una muestra de aproximadamente 0,5 g. Para algunas determinaciones de susbtratos más concentrados se tuvieron que utilizar 15 mL. 3.3. Demanda Química de Oxígeno soluble (DQOsol). El fundamento del método es el mismo que el utilizado para la determinación de la DQO total, pero se ha utilizado el método de reflujo cerrado (número 5220 C). El análisis se realiza sobre la fracción soluble. La obtención de la fracción soluble se realizó mediante la centrifugación y separación sucesiva del sobrenadante, a una velocidad de 3500 rpm, y filtrado mediante membrana de acetato de celulosa de 0,45 μm. Se utilizó el método de reflujo cerrado, utilizando un tubo de 10 mL de capacidad, con tapón de rosca, al que se añaden 2 mL de la muestra filtrada y Optimización de la digestión anaerobia de purines de cerdo 69 diluida, 1 mL de dicromato 0,25 N y 3 mL de H2SO4 con sulfato de plata como catalizador. La digestión se realiza con los tubos cerrados, en una estufa ”Selecta 202”, a 150ºC, durante 2 horas. Una vez enfriados, el contenido se trasvasa a matraces Erlenmeyer, arrastrando los posibles restos con agua desionizada. Se añaden 5 mL de H2SO4, 5 gotas de ortofenantrolina como indicador, y se valora con Sal de Mohr 0,0125 N. Para el cálculo se ha utilizado la ecuación 3.4. 3.4. Nitrógeno Kjeldhal El método Kjeldhal determina el nitrógeno en estado trinegativo. No tiene en cuenta el nitrógeno en forma de azida, azina, azo, hidrazona, nitrato, nitrito, nitrilo, nitroso, oxamina y semicarbazona. A pesar de ello normalmente se asocia a nitrógeno total, por considerar las fracciones más importantes de formas nitrogenadas en los residuos animales, nitrógeno orgánico y amoniacal. El método ha sido adpatado del método 4500 de Standard methods for examination of water and wastewater (APHA, 1995). Se basa en digerir la muestra con temperatura, en medio ácido con un catalizador de Selenio, de forma que los compuestos orgánicos nitrogenados formen formas amoniacales, sulfato amónico. La reacción se podría resumir en: C a H b N c O d H 2 SO 4 Catl. Selenio o SO 2 n CO 2 n H 2 O (NH 4 )2 SO 2 (3.5) Posteriormente se analiza el contenido total de sales amoniacales mediante la destilación de la muestra digerida, con un destilador Tecator (Kjeltec System 1026 Distilling Unit). Se añade una base fuerte (NaOH 40%) para subir el pH y desplazar el equilibrio hacia la formación de amoníaco libre, siguiendo la siguiente expresión: (3.6) (NH 4 )2 SO4 2NaOH o 2NH 3 Na 2 SO4 2H 2 O El destilado se recoge en ácido bórico con indicador (rojo de metil). Al recogerse el vapor en un medio ácido el amoníaco pasará a la forma iónica no volátil. NH 3 H 3 BO3 (en exceso) o NH 4 H 2 BO3- (3.7) Finalmente se realiza la valoración del borato que ha reaccionado con el amoníaco, mediante titulación con un ácido fuerte, HCl, de normalidad conocida. 70 Métodos analíticos H 2 BO3- H o H 3 BO3 (3.8) El nitrógeno total se estima utilizando la siguiente expresión: N k ( mg / kg ) 14000 * Vm Vbl * N HCl , M muestra (3.9) Mmuestra : cantidad de muestra (g), Vm es el volumen de HCl consumido en la valoración de la muestra (mL), Vbl es el volumen de HCl consumido en la valoración del blanco y NHCl es la normalidad del HCl. 3.5. Nitrógeno amoniacal El nitrógeno amoniacal se ha analizado por el método de destilación, con un destilador marca Tecator (Kjeltec System 1026 Distilling Unit), siguiendo el método 4500-NH3 B de Standard methods for examination of water and wastewater (APHA, 1995). El método es exactamente el mismo que el utilizado para destilar y valorar el nitrógeno Kjeldhal, sin la digestión previa y utilizando como base MgO. La determinación se realiza sobre la fracción el sobrenadante de centrifugar a 3500 rpm, tomando 1 mL de muestra. El cálculo de la concentración de nitrógeno amoniacal expresado en mg de N-NH4+/L sería: N - NH 4 (mg/L) = 14000 * Vm Vbl * N HCl , Vmuestra (3.10) donde Vmuestra es el volumen de muestra utilizado. 3.6. Nitrógeno orgánico El contenido de nitrógeno orgánico de una muestra se determina por diferencia entre el nitrógeno Kjeldahl y el nitrógeno amoniacal: >N org @ >N k @ >N NH 4 @ (3.11) El contenido total de proteínas se puede obtener de forma aproximada, multiplicando la concentración de nitrógeno orgánico por el factor 6,25 (MAPA, 1994). (3.12) >Pr oteínas @ 6,25 x N org > @ Optimización de la digestión anaerobia de purines de cerdo 71 3.7. Amoníaco libre El amoníaco libre se calculó a partir de la concentración de amonio total, del pH del medio y de la temperatura, a partir del siguiente equilibrio: Ka NH 4 NH 3 H , (3.13) donde Ka es función de la temperatura. La función se obtuvo mediante regresión polinómica a partir de los datos de Lide (1993). La expresión obtenida fue: pKa 10.072 0.0356 * temp 9 * 10 5 * temp 2 4 * 10 8 * temp 3 . (3.14) A partir del equilibrio, la concentración de amoníaco libre se calcula como función del pH y de la concentración de amonio total, quedando la siguiente expresión: >NH 3 @ >NH 4 @T 1 10 pKa pH . (3.15) 3.8. pH El pH es la forma común de expresar la concentración del ion hidrógeno en las soluciones acuosas: pH > @ log10 H . (3.16) Se midió directamente sobre la muestra, con un electrodo Crison 52-11, especial para líquidos viscosos y con alto contenido en proteína, conectado a un medidor de pH/mV Crison GLP 22. Se realizó la calibración con disoluciones tampón estándar CRISON de pH 7,02 y 4,00 a 20ºC. La resolución de la lectura es de 0,01 unidades de pH y la precisión de r0,01. 3.9. Alcalinidad parcial, total y relación de alcalinidades La alcalinidad de un agua es su capacidad para neutralizar ácidos y constituye la suma de todas las bases valorables. La alcalinidad depende del pH de punto final utilizado. El método estándar (2320) de Standard methods for examination of water and wastewater (APHA, 1995), consiste en la valoración con 72 Métodos analíticos un ácido fuerte hasta pH 4,3. A pH 4,3 más del 99% del bicarbonato del sistema es convertido a CO2. Sin embargo al hacer esta valoración se considera más del 80% de los de ácidos grasos volátiles (Hill y Jenkins, 1989), compuestos presumiblemente abundantes en los sistemas anaerobios. Por ello Hill y Jenkins (1989) propusieron la utilización de la valoración hasta pH 5,75, que se ajusta mucho mejor al valor real de alcalinidad debida al bicarbonato. En el presente trabajo se propone hacer una valoración de la alcalinidad en dos pasos, primero a 5,75 y posteriormente a 4,3. Tomando estos dos puntos finales de pH se definen tres parámetros de medida de la alcalinidad: alcalinidad total (AT) medida al punto de pH 4,3; alcalinidad parcial (AP), asociada a la alcalinidad al bicarbonato, medida al punto de pH 5,75 y alcalinidad intermedia (AI), asociada a la concentración de AGV, y estimada como diferencia de ambas. La valoración se realizó con ácido clorhídrico de normalidad exacta conocida con medida continua del pH, hasta los dos puntos citados. El instrumental utilizado fue el mismo que el utilizado para medir el pH. El cálculo de la alcalinidad se realizó utilizando las siguientes expresiones: V4.3 N HCl * 50 Vol muestra (mL) AT Al cal. Total(mg CaCO3 /L ) = AP Al cal. Bicarbonato(mg CaCO3 /L ) = V5.75 N HCl * 50 Vol muestra (mL) V V5.75 RA (Relación de Alcalinidades) = 4.3 V4.3 (3.17) (3.18) (3.19) 3.10. Ácidos Grasos Volátiles Los ácidos grasos de cadena corta, o volátiles, AGV, analizados han sido acético, propiónico, butírico y valérico, siendo posible separar los dos isómeros del butírico y del valérico (formas iso y n). Se analizan por cromatografía de gases, con un cromatógrafo para columna capilar Trace 2000 (ThermoInstruments), equipado con un inyector split-splitless, un detector de ionización de llama (FID) y un inyector automático (Autosampler AS2000), con un carro con capacidad para 90 muestras. Como gas portador se utiliza helio. La columna es una columna capilar FFAP de 0,25 mm de diámetro interior y 30 m de longitud. Para el detector se utiliza hidrógeno y aire sintético y helio como make-up con un caudal de 30 Optimización de la digestión anaerobia de purines de cerdo 73 mL/min. Se inyecta un volumen de muestra de 1 μL, en modo split, con un flujo de gas portador constante a 1 mL/min, una relación de split de 20/1 y una relación de purga de septum de 5/1. La temperatura inicial del horno es de 100ºC, durante 4 minutos, momento a partir del cual sueb hasta los 155ºC con una velocidad de 5ºC/minuto. Finalmente sube hasta 240ºC. Todos los ácidos grasos analizados salen en un tiempo de 12,5 minutos, pero la duración total del análisis es de 27 minutos para purgar los compuestos retenidos en la columna. La temperatura del inyector y del detector es 230ºC. En la Figura 3.1 se muestra un cromatograma como ejemplo. Figura 3.1. Ejemplo de cromatograma del análisis de AGV. La muestra se centrifuga a 5000 rpm, separando el sobrenadante. Si es necesario se procede a la correspondiente dilución; casi todas las muestras se diluuen al 10%. Se vuelve a centrifugar a 5000 rpm y el sobrenadante se filtra con una membrana de acetato de celulosa de 0,45 μm. Una vez filtrada la muestra, se realiza la extracción con dietil-éter, 1/1, previa acidificación con HCl concentrado. La calibración se realiza con mezcla de 6 patrones a diferentes concentraciones, desde 1 mg/L hasta 2000 mg/L. Se utiliza un método lineal de calibración, obteniendo altos coeficientes de regresión (Figura 3.2). Cada día se inyecta al menos un patrón para detectar posibles variaciones en los tiempos de retención. 74 Métodos analíticos Optimización de la digestión anaerobia de purines de cerdo Área Área Acético Propiónico y = 286.21x 75 y = 1039.2x R2 = 0.9961 2 R = 0.9972 700000 2500000 600000 2000000 500000 400000 1500000 300000 1000000 200000 500000 100000 0 0 0 500 Área 1000 1500 500 1000 1500 2000 2500 Conc (mg/L) Área y = 1791.6x Iso-butírico 0 2000 2500 Conc (mg/L) N-butírico R2 = 0.997 y = 1771.6x R2 = 0.9968 4000000 4000000 3500000 3500000 3000000 3000000 2500000 2500000 2000000 2000000 1500000 1500000 1000000 1000000 500000 500000 0 0 0 500 1000 1500 2000 0 2500 500 1000 1500 Área Iso-valérico Área y = 2224.9x 2000 2500 Conc (mg/L) Conc (mg/L) N-valérico y = 2233.3x R2 = 0.9972 R2 = 0.9972 5000000 4500000 4000000 3500000 3000000 2500000 2000000 1500000 1000000 500000 0 5000000 4500000 4000000 3500000 3000000 2500000 2000000 1500000 1000000 500000 0 0 500 1000 1500 2000 2500 0 500 1000 Conc (mg/L) 1500 2000 2500 Conc (mg/L) Figura 3.2. Ejemplo de rectas de calibración para los diferentes AGV. 3.11. Composición de biogás, CH4 y CO2 El biogás se analiza mediante cromatografía de gases con el equipo GC8000 Top de ThermoInstruments, equipado con un inyector para columnas empacadas J70 y un detector de conductividad térmica. Se utiliza una columna empacada Porapak N de 2 mm de diámetro y 2 m de longitud. La temperatura del detector es de 120ºC y la del inyector de 130ºC. La 76 Métodos analíticos temperatura del horno se mantiene constante a 30ºC. El tiempo total de análisis es de unos 4 minutos. Se utiliza Helio como gas portador con un flujo de 20 mL/min. La calibración se realiza utilizando dos patrones, uno mezcla de CO2 y CH4 y, el otro mezcla de N2 y CO2, de concentración conocida. Se utilizaron diferentes volúmenes de muestra, para obtener los diferentes puntos de la recta de calibración. Las rectas obtenidas en una de las calibraciones realizadas se muestra en la Figura 3.3. Optimización de la digestión anaerobia de purines de cerdo N2 Area y = 359101x Area CH4 77 y = 319482x 2 R = 0.9972 2 R = 0.996 4000000 8000000 3000000 6000000 2000000 4000000 1000000 2000000 0 0 0 2 4 6 8 10 12 Area CO2 0 5 10 15 20 25 Conc (micromoles) Conc (micromoles) y = 440732x 2 R = 0.9994 8000000 6000000 4000000 2000000 0 0 5 10 15 20 Conc (micromoles) Figura 3.3. Rectas de calibración para los tres componentes del biogás considerados 3.12. Fósforo total (PT) A partir de las cenizas obtenidas por calcinación a 550ºC, se realiza una extracción con ácido clorhídrico concentrado. El fósforo inicialmente contenido en las cenizas se disuelve y pasa a la solución. Utilizando un método colorimétrico, adaptado del método 4500-P C de Standard methods for examination of water and wastewater (APHA, 1995), se determina el contenido de PT. La lectura de absorción se realiza a 430 nm mediante el espectrofotómetro “ATI-UNICAM UV2”. Se utiliza el reactivo nitrovanadomolíbdato, que al reaccionar con el ácido fosfórico, forma fosfomolibdovanadato, que da una coloración amarilla. La cantidad de reactivo es un 20% del volumen del matraz. Una vez preparada la solución se debe dejar reposar 1 hora antes de realizar la lectura. El cálculo 78 Métodos analíticos de la concentración se obtiene mediante la siguiente expresión: V (L) VA( L ) Cenizas(%) , x x PT ( mg / g ) C( mg / L ) x B V A ( L ) Cenizas( g ) 100 (3.20) donde C, es el valor obtenido a partir de la lectura en el espectrofotómetro, VA es el volumen de la dilución ácida de las cenizas y VB es el volumen de la dilución realizada para hacer la determinación. 3.13. Cationes (Na, K, Ca, Mg, Cu y Zn). Los cationes se analizan por fotometría de llama, utilizando el equipo “PERKIN-ELMER 5000”, sobre la base de un extracto ácido sobre las cenizas, realizado con el mismo procedimiento que en el método de determinación del fósforo. Los cationes Na y K se determinan mediante la técnica de fotometría de llama en emisión, mientras que para la determinación del Ca, Mg, Cu y Zn, se trabaja en absorción. En la determinación de Na y K, es necesario añadir 1 mL de ClLi 2,5N por cada 100 mL antes de aforar, para evitar su ionización. En la determinación del Ca y Mg, es necesario añadir 1 mL de SrCl2 al 3% por cada 10 mL de solución. Para la determinación de Cu y Zn es necesario añadir a los patrones HCL, en igual cantidad que la presente en las muestras. Se deberá aplicar la ecuación 3.20 al valor obtenido por la lectura en el instrumento, para referirlo a la muestra fresca. 3.14. Contenido en grasa. Método Soxlet. El contenido en grasa se ha determinado en el laboratorio de Producción Animal de la ETSEA, siguiendo el método oficial (MAPA, 1994). El contenido en grasa bruta de un producto se define convencionalmente como la parte del mismo, extraíble por éter etílico en determinadas condiciones. Incluye además de la grasa, otras sustancias solubles en éter como ceras, pigmentos, vitaminas, etc. Se pesa una cantidad conocida de muestra seca y molida (500 μm) y se introduce en un cartucho taponado con algodón. Se tara un matraz (desecado en estufa y enfriado en desecador) y se introduce el cartucho en el extractor, añadiendo éter etílico una vez conectado el matraz. Se procede a la extracción, continuando hasta que el éter es incoloro, con una duración aproximada de 6 horas. Se saca el cartucho del extractor y se recupera el Optimización de la digestión anaerobia de purines de cerdo 79 éter. El matraz con el resto del extracto y del disolvente se seca durante media hora en una estufa de desecación a 100ºC. Se deja enfriar en desecador y, finalmente, se pesa cuando esté a temperatura ambiente. El contenido en grasa se calcula mediante la siguiente expresión: Grasa bruta (% M.S.) P1 P2 * 100 , P (3.21) donde P1: peso del matraz con el extracto etéreo (g), P2: peso del matraz vacío (g) y P: peso de muestra seca empleada (g). 3.15. Contenido en fibra. Método fibra ácido detergente El contenido en celulosa, hemicelulosa y lignina, se determinó por diferencia de los resultados obtenidos al aplicar la técnica de fibra ácido detergente, FAD (celulosa +lignina), lignina y fibra neutro detergente (celulosa, hemicelulosa y lignina), sobre matera seca. Se ha determinado en el laboratorio agroalimentario de Cabrils, de la Generalitat de Catalunya, siguiendo el método oficial (Métodos oficiales de análisis de los alimentos, MAPA, 1994). 80 Métodos analíticos 4. MEJORA DE UN MODELO ESTRUCTURADO DE DIGESTIÓN ANAEROBIA MEDIANTE LA INCORPORACIÓN DE LAS ECUACIONES PARA LA SIMULACIÓN DINÁMICA DEL PH DE LA TRANSFERENCIA LÍQUIDO-GAS 80 Simulación dinámica del pH Optimización de la digestión anaerobia de purines de cerdo 81 4.1. Resumen Con el objetivo de contribuir a la construcción de herramientas informáticas de simulación de sistemas anaerobios, se ha construido un algoritmo de modelización dinámica del pH. Se llegó a una ecuación diferencial que simula la variación del pH en sistemas dinámicos, a partir de la simulación de la variación de las componentes del balance de cargas. Se considera también la simulación dinámica de las variables de la fase gaseosa, que resulta fundamental para la correcta simulación del pH. Este algoritmo puede ser introducido en un modelo de simulación dinámica, tratando el pH como una variable de estado y no como un parámetro a calcular en cada paso de integración, lo que proporciona una mayor rapidez de cálculo. Integrando el sistema de ecuaciones diferenciales ordinarias mediante la rutina RungeKutta-Felhberg, se consiguió un muy buen ajuste del modelo y una gran rapidez de cálculo. El algoritmo de pH se ha validado para sistemas acuosos sencillos, habiendo obtenido muy buenos ajustes a datos experimentales. El algoritmo de simulación de pH se insertó en un modelo dinámico de digestión anaerobia, basado en el modelo propuesto por Angelidaki et al. (1999). Además de la simulación dinámica del pH, se realizó también la simulación dinámica de la transferencia líquido-gas. Este modelo de digestión anaerobia, pese a la dificultad para su calibración completa, resulta una herramienta útil para prever la respuesta del sistema a posibles estados de sobrecarga o de introducción de componentes inhibidores, y en general para predecir la tendencia de un sistema anaerobio. 4.2. Introducción Un modelo validado del proceso de digestión anaerobia constituye una herramienta muy útil, tanto para la optimización del diseño, como para la operación de plantas de tratamiento, permitiendo determinar el efecto de las características del substrato y la carga sobre el desarrollo del proceso, desarrollar sistemas de control en la operación, predecir el desarrollo de procesos, mejorar el conocimiento de los mismos mediante comparación con datos experimentales y testar hipótesis. Un modelo puede ser de estado estacionario o dinámico. Los primeros pueden ser útiles en la fase de diseño y dimensionamiento de digestores, 82 Simulación dinámica del pH mientras que los modelos dinámicos pueden predecir la evolución de los procesos de forma continua, permitiendo estudiar el efecto de cambios en el sistema, siendo muy útiles para el mejor conocimiento de los procesos que ocurren en un reactor anaerobio. La digestión anaerobia consiste en la degradación de la materia orgánica mediante la acción de un amplio grupo de microorganismos en ausencia de oxígeno. Los modelos tradicionales de digestión anaerobia dividen el proceso en varias fases o estadios, llevados a cabo por diferentes grupos de bacterias, que mantienen relaciones entre ellas. En la literatura se recogen gran variedad de modelos dinámicos estructurados, que, aunque se basan en los mismos principios, dan un peso especial a un determinado parámetro, considerándolo como el principal modulador del proceso. Hay modelos que utilizan el hidrógeno como parámetro fundamental, siendo el más importante de entre estos, el propuesto por Mosey (1983). Algunos consideran como principal modulador la concentración de ácidos grasos no ionizados, cuyo principal exponente es el modelo postulado por Andrews y Graef (1971). A partir de éste se han desarrollado otros, incluyendo nuevos variables, como el amoníaco, el pH o la temperatura (Kleinstreuer y Poweigha, 1982). Otros modelos han considerado también los ácidos grasos volátiles como el principal modulador, aunque considerando la concentración total, como el que propuso Hill (1982). Este modelo fue desarrollado para simular reactores trabajando con residuos ganaderos como substrato, por lo que presenta un gran interés. En modificaciones posteriores incluyeron la acetogénesis a partir del valérico (Hill y Cobb, 1993). Los coeficientes de producción fueron determinados experimentalmente a partir de plantas piloto trabajando con residuos ganaderos de diferente tipo. Otros modelos, como el de Angelidaki et al. (1993), se centran en el efecto de inhibición por amoníaco libre y la interdependencia del pH. Varias modificaciones de este modelo han sido publicadas posteriormente, incluyendo la degradación de otros tipos de macromoléculas, como los lípidos (Angelidaki et al., 1996) y proteínas (Angelidaki et al., 1999). Otros modelos han considerado el importante papel del amonio como regulador, incluyendo también otros parámetros que pueden jugar un papel muy importante en la digestión de residuos ganaderos, como es la actividad de las bacterias sulfato-reductoras y la inhibición por sulfuros (Vavilin et al., 1994, 1995 y 1996; Kalyuzhnyi et al., 1997). Optimización de la digestión anaerobia de purines de cerdo 83 4.2.1. La importancia del pH El pH puede influir sobre el crecimiento de los microorganismos, al favorecer la formación de formas inhibidoras, caso de la inhibición por amoníaco (favorecida a pH alcalinos), la inhibición por ácidos grasos y la inhibición por H2S (favorecida a pH ácidos). En la digestión anaerobia termofílica de residuos ganaderos, y especialmente purines de cerdo, el principal modulador del sistema es el amoníaco libre (Angelidaki y Ahring, 1993b), cuya concentración es función del pH y temperatura, aumentando con estas variables. El principal efecto del amoníaco es sobre la población metanogénica, especialmente la acetoclástica, por lo que al aumentar la concentración de amoníaco libre aumenta la concentración de acético (Koster y Lettinga, 1988; Angelidaki y Ahring, 1993b; Hansen et al., 1998), que a su vez puede provocar la inhibición de la acetogénesis y la consiguiente acumulación de otros AGV, como propiónico, butírico y valérico (Ahring y Westermann, 1988; Fukuzaki et al., 1990), lo que puede provocar una bajada del pH del sistema. La bajada del pH será más o menos acentuada en función de capacidad tampón del medio. La bajada del pH, sin embargo, provoca una bajada en la concentración de amoníaco libre, y por tanto, la reactivación del proceso, aunque con una tasa de producción de metano menor (Angelidaki y Ahring, 1993b). El pH tiene un importante efecto sobre las tasas de crecimiento de los microorganismos anaerobios, hidrolíticos, acidogénicos, acetogénicos y metanogénicos (Clark y Speece, 1989), que alcanzan un óptimo para un pH en torno a la neutralidad. Por otro lado, la acumulación de ácidos grasos volátiles puede provocar la inhibición del proceso enzimático de hidrólisis (Angelidaki et al. 1993), lo que resulta en una menor eliminación de materia orgánica. En general, se considera que el pH del medio afecta a la velocidad máxima de crecimiento de los microorganismos, siguiendo una función de Michaelis normalizada (Angelidaki et al., 1993; Kalyuzhnyi, 1997) o similar (Siegrist et al., 1993; Vavilin et al., 1995). La simulación dinámica del pH en los sistemas anaerobios es interesante para el desarrollo de un modelo de simulación matemática del proceso global de la digestión anaerobia de residuos ganaderos. La mayoría de los modelos dinámicos de digestión anaerobia tienen en cuenta el pH, aunque lo consideran como un parámetro a calcular en cada paso, sin establecer las ecuaciones de su variación temporal. Estos modelos han obtenido buenas aproximaciones a valores reales, pero podrían ser mejorados introduciendo un algoritmo de simulación dinámica del pH como el que se propone, dado el carácter dinámico de esta variable. 84 Simulación dinámica del pH 4.3. Objetivos El objetivo del presente trabajo es la mejora del modelo de simulación de Angelidaki et al. (1999), mediante la introducción de la simulación dinámica del pH y de las variables de la fase gaseosa, de forma que mejore la precisión del mismo. Para ello se propusieron los siguientes objetivos parciales: Construcción y validación de un algoritmo que permita la simulación x dinámica del pH, considerando éste como una variable de estado. x Desarrollar el sistema de ecuaciones que permitan la modelización en la interfase líquido-gas. Introducción de las ecuaciones desarrolladas en el modelo general de x simulación del proceso anaerobio de substratos complejos y conseguir una herramienta útil para la simulación y optimización del proceso de codigestión. 4.4. Modelización del pH en sistemas anaerobios. Revisión bibliográfica La mayoría de los autores que han simulado el pH en los sistemas anaerobios se han basado en la resolución de la ecuación del balance de cargas en un momento determinado, o bien, en aproximaciones a las soluciones analíticas. El número de compuestos utilizados como variables varía en función del autor considerado, yendo desde los que tan sólo consideran el sistema carbonato/ácidos grasos, hasta los que utilizan más parámetros (Tabla 4.1). Andrews y Graef (1971) calcularon el pH considerando una serie de simplificaciones: 1) despreciar la concentración de H+ y OH- en la ecuación de balance de cargas, si el pH se mantiene en el rango entre 6 y 8; 2) la fracción no ionizada de ácidos grasos volátiles es despreciable comparada con la fracción ionizada y 3) la concentración de carbonatos en la solución es despreciable. Con estas simplificaciones se calcula el pH como función de las concentraciones de dióxido de carbono y bicarbonato expresado mediante la ecuación 4.2. La concentración de dióxido de carbono es simulada de forma dinámica, considerando las tasas de producción-eliminación biológica y transferencia líquido-gas, y la concentración de bicarbonato se calcula a partir de la ecuación de balance de cargas (ecuación 4.1). Consideraron un sistema continuo de mezcla completa. Costello et al. (1991), basándose en el modelo anterior, realizaron el cálculo del pH a partir de la ecuación del balance de cargas, con menos Optimización de la digestión anaerobia de purines de cerdo 85 simplificaciones. Así, consideraron que las concentraciones de H+ y OH- no eran despreciables en el balance de cargas (ecuación 4.3), y que la concentración de ácidos débiles no ionizados tampoco son despreciables. No consideró los carbonatos ni otras substancias, como amonio, fosfatos y otros aniones o cationes. Las concentraciones de bicarbonato y de ácido ionizado fueron calculadas a partir de la concentración total (en el caso del bicarbonato como suma de HCO3- y CO2 disuelto). El pH lo calcularon mediante la resolución por métodos numéricos de la ecuación 4.4, en cada paso de integración. Siegrist et al.(1993) introdujo el concepto de pH como variable temporal, integrando su simulación en el modelo general, como una variable más a la que le afectan una serie de procesos. Sin embargo el modelo hace muchas simplificaciones y calcula el pH sólo como función de la protonización del bicarbonato y desprotonización del CO2, ecuaciones 4.5 y 4.6, es decir la variación de la alcalinidad debida al bicarbonato. Considera la influencia del amonio, pero no considera la disociación de los ácidos grasos volátiles u otras variables. Ryhiner et al. (1993) utilizó también la ecuación de balance de cargas, considerando las componentes ionizadas, estimando el pH al resolver la ecuación implícita. Para las soluciones numéricas se consideró tan sólo el equilibrio entre cationes y aniones. Angelidaki et al. (1993; 1996, 1999) realizaron una modelización del sistema físico-químico, continuista con respecto a Andrews y Graef (1971) y Costello et al. (1991), pero incluyendo más variables (ecuación 4.7). El pH se calcula a intervalos fijos de tiempo, resolviendo por métodos numéricos la ecuación 4.8, que adopta diferentes expresiones en función del pH del medio (ácido o básico). Al dividir la resolución en dos casos, junto con el método numérico de resolución (método de la secante con dos puntos finales, uno por encima y otro por debajo del valor de la solución), se consiguió evitar problemas de convergencia. Este modelo introduce la dependencia de la temperatura de las constantes de disociación, de las constantes de solubilidad de los gases en la fase líquida, y de los parámetros de los procesos biológicos. La inclusión de la temperatura adquiere una relevancia especial en el caso de sistemas fuertemente influidos por la inhibición de amoníaco libre, cuya concentración es función de la temperatura. 86 Simulación dinámica del pH Tabla 4.1. Resumen de ecuaciones de simulación del pH en modelos estructurados de digestión anaerobia Autores Balance de cargas Andrews y Graef(1971) >HCO3 @ >Z @ >AH @ @ > > @ > > @> @ Costello et HCO3 ¦ AH OH al. (1991) H Na HCO3 H Siegrist et al. U 14 (1993) @ U 14 o m (4.1) (4.3) CO 2 H 2 O U15 k14 HCO3 · H ; > Cálculo del pH resolviendo la ecuación: @> @ (4.5) >HCO 3 @ 2>CO 3 @ ¦ >Ai @ >H 2 PO 4 @ 2>HPO 42 @ (4.7) i >NH 4 @ >Z @ Ch( pH ) Vavilin et al. (1995) @ > @ i k (4.9) 2·¦ >Ak2 @ ¦ >B j @ >H @ >Z @ > @ > k > Kiely et al. HCO 3 (1997) j @ >Z @ >NH 4 @ >AH @ (4.11) >CO2,D @ >HCO3 @ KA H Na > @> > @ d H dt (4.2) (4.4) @ ¦ Q j ·U j (4.6) U14 U15 Si Ch pH !0 >H @ Ch >H @ Ch2 4 K w 2 Si Ch pH 0 2K w Ch Ch2 4 K SC SC S P ¦ Ai OH ¦ HAk K1 >HCO3 @ ¦ >>AH@ @ >HK w @ 1 k15 >CO 2 @ U15 Angelidaki et al. (1993) >H @ 1 >H @ >H @2 w > @ (4.10) > @ > @ >H @ (4.8) SP H 1 KC1 KC1 KC1KC 2 SS SA H H 1 1 KA KS SN 0 H Z Kw 1 KN H > @ , > @ K1 >CO2,D @ >HCO3 @ (4.12) *SC es la concentración total de carbono; SP:concentración total de ortofosfato; SA: concentración total de acetato y propionato; SS concentración total de sulfuros, SN concentración total de amonio y Z carga neta de cationes y aniones inorgánicos Vavilin et al. (1995) realizaron el balance de cargas incluyendo los mismos compuestos que Angelidaki et al. (1993), añadiendo otros como los sulfuros (ecuación 4.9), y resolviendo en cada punto de la simulación la ecuación implícita resultante (ecuación 4.10). Optimización de la digestión anaerobia de purines de cerdo 87 Kalyuzhnyi (1997) realizó la simulación del pH siguiendo el método propuesto por Angelidaki et al. (1993), pero despreciando carbonato y amonio, por considerarlo poco influyente en sus condiciones, lo que es discutible si el pH está por encima de 8. Kiely et al. (1997) vuelven a utilizar el algoritmo propuesto por Andrews y Graef (1971), haciendo varias de las simplificaciones que proponen los primeros, incluyendo el sistema amonio/amoníaco (ecuación 4.11). Transferencia líquido-gas. Existe una clara relación entre la trasferencia líquido-gas y el pH, pudiendo verse afectados mutuamente. Un cambio en el pH condicionará la concentración de las formas no ionizadas y por tanto puede afectar a la transferencia líquido-gas. La mayoría de los modelos estructurados consideran el proceso de transferencia líquido gas gobernado por la teoría de la doble capa. El flujo de materia que pasa de la fase líquida a la fase gas es función del gradiente de concentración en el líquido y la concentración del equilibrio en la fase límite, que se expresa como el producto de la correspondiente presión parcial en la fase gaseosa y el coeficiente de Henry (Merkel y Krauth, 1999), N g ,i K La x V L x C i C *i K La x V L x C i Hei x p i , (4.13) donde Ng,i es el flujo de moles de la substancia i que pasan de la fase líquida a la fase gaseosa (Mol/d), VL es el volumen de líquido, KLa es la constante de transferencia líquido-gas, Ci es la concentración de la substancia en el líquido (Mol/L), en la forma no ionizada (en el caso del CO2 se correspondería con la concentración de CO2 disuelto), y Ci* es la concentración de equilibrio de dicha substancia con la fase gaseosa, calculada suponiendo que se cumple la ley de Henry, por lo que es proporcional a la presión parcial de la substancia i en el espacio de cabeza (pi), siendo Hei la constante de Henry, que depende de la temperatura. Algunos modelos no utilizan la teoría de la doble capa, como el modelo propuesto por Angelidaki et al. (1993, 1996, 1999), quienes consideran que existe un equilibrio casi estacionario entre las fases gas y líquido, considerando que se cumple en todo el volumen del líquido la ley de Henry, y no sólo en la capa límite. Normalmente sólo se considera limitante la transferencia líquido-gas del CO2, despreciando otras componentes del gas como el metano, o el hidrógeno, puesto que la solubilidad de estos gases es más baja (Merkel y Krauth, 1999; Costello et al., 1991, etc.). La mayoría de los modelos consideran la constante de transferencia de 88 Simulación dinámica del pH masas constante, para cada compuesto, aunque Merkel y Krauth (1999) demostraron que existe una relación entre el valor de KLa y la velocidad de flujo superficial ascendente del gas, u0gas (ecuación 4.14), teniendo especial importancia en situaciones de cambios bruscos en las condiciones de operación del reactor, k La E K u 0gas . (4.14) 4.5. Desarrollo del algoritmo de simulación dinámica del pH Puesto que el pH tiene una gran importancia en el proceso anaerobio y presenta un claro comportamiento dinámico, se decidió dar al pH categoría de variable de estado del sistema, estableciendo la ecuación diferencial que determina su variación en el tiempo. Partiendo de los algoritmos de cálculo del pH usados en modelos previos se construyó un algoritmo en el que intervienen las principales variables de los sistemas anaerobios, considerando varios sistemas tampón, la influencia de la temperatura y la existencia de procesos de transferencia líquido-gas. Además, la concentración total de determinados compuestos como CO2, ácidos grasos volátiles, amonio, etc., varia en función de los procesos biológicos propios del proceso anaerobio, tal y como se muesstra en el apartado 4.6. El poder tampón se puede medir mediante un parámetro (índice tampón), determinado por la variación de la concentración del ácido o base respecto al pH del medio, y que alcanza el valor máximo en el momento en que el pH es igual a la constante ácido-base de ese compuesto. El poder tampón de una mezcla compleja viene dado por la suma de los correspondientes índices tampón de cada uno de los compuestos que intervienen (Moosbrugger et al., 1993). El principal sistema tampón en los sistemas anaerobios es el sistema bicarbonato-carbónico, por ser el más importante cuando el pH está en torno a la neutralidad. El bicarbonato es un compuesto que interviene como substrato o producto en las diferentes fases del proceso. En el sistema del bicarbonato-carbónico se consideró también el proceso dinámico de transferencia líquido-gas, siguiendo la teoría general de transferencia de masas entre fases, aplicada por Costello et al. (1991). El sistema carbonato-bicarbonato-CO2 presenta tres posibles estados de Optimización de la digestión anaerobia de purines de cerdo 89 ionización, estimando la concentración de cada una de ellas en función de la concentración total en la fase líquida, suma de los tres estados de ionización posibles, del pH y de las constantes de ionización (ecuaciones 4.16, 4.17 y 4.18). He, K L a CO2 g o CO2 l0 m K1,CO2 CO2 l0 H 2O o H HCO3 m >CO2 @T ,L >CO32 @ >CO32 @ >HCO3 @ ª«¬H 2CO3 º»¼ >CO2 @T ,L >H @2 K1,CO2 K 2,CO2 >HCO3 @ 2 3 >CO2 @T ,L > @ >H @ K 2 ,CO2 (4.15) (4.16) 1 (4.17) > @ 2 · K 2 ,CO2 §¨ H H ¸ 1 ¸¸ H ¨¨ K1,CO2 K 2 ,CO2 K 2 ,CO2 ¹ © > @ >H CO @ K 2 ,CO 2o CO 2 H 3 m >CO2 @l0 >CO2 @T ,L > @ > @ 2 · K1,CO2 K 2 ,CO2 § H H ¨ 1¸ 2 ¸ ¨ K1,CO K 2 ,CO K 2 ,CO2 H 2 2 ¹ © > @ (4.18) A los pH habituales de los residuos ganaderos (alrededor de 8) adquiere especial importancia el sistema amonio-amoníaco. Las expresiones para la concentración de las diferentes formas se muestran en las ecuaciones 4.18 y 4.19. K NH 3 K La , He o NH 4 o H NH 3 l0 NH 3 ( g ) m m >NH 4 @T >NH 4 @ >NH 3 @ l0 (4.19) 90 Simulación dinámica del pH 4 @T > NH 4 @ K>NH NH 3 >H @ 1 (4.20) > NH 3 @ l0 >NH 4 @T >H @ K NH 3 (4.21) 1 Los ácidos grasos volátiles son los principales intermediarios del proceso anaerobio por lo que son muy utilizados como parámetros de control y diagnóstico de problemas o desequilibrios. Además de tener importancia en sí mismos, si se acumulan influirán en el pH del medio, pues a pesar de ser ácidos débiles, la concentración puede llegar a ser suficientemente alta como para afectar al pH. Los ácidos grasos volátiles que se consideran son los cuatro de cadena más corta (acético, propiónico, butírico y valérico), y no se diferencia entre los dos isómeros del butírico y del valérico, ya que ambos tienen la misma fuerza iónica. Tampoco se consideró la volatilización de dichos compuestos, por considerarla despreciable al pH normal de trabajo, ya que la forma volátil es la forma no ionizada. La concentración de las formas ionizada y no ionizada de los ácidos grasos volátiles (AGV) se establece como función de la concentración total de cada substancia, del pH y de la constante de ionización. ValH KVal o Val H m BuH K Bu o Bu H m Pr H K Pr o Pr H m AcH K Ac o Ac H m Para cualquiera de estos cuatro compuestos o, en general, para un ácido débil monoprotonado la concentración de cada una de las formas de ionización se puede expresar en función de la concentración total del ácido (ecuaciones 4.22, 4.23 y 4.24). AH KA o A H m >AH @T >AH @ >A @ (4.22) Optimización de la digestión anaerobia de purines de cerdo >A @ >>HAH @@T 1 >AH @T >AH @ (4.23) KA >H @ KA 91 (4.24) 1 También es importante la concentración de aniones y cationes presentes en la disolución, pues afectarán al balance de cargas, actuando como bases o ácidos conjugados. Se han considerado cuatro tipos genéricos de substancias, que dependen de la carga, quedando cubiertos cationes como sodio, calcio, potasio, magnesio y aniones como sulfatos, cloruros, etc. Se comportarán como ácidos o bases fuertes y no son función del pH del medio. Z Z 2 Z Z 2 Se ha incluido también el sistema tampón del ortofosfórico, que en determinados substratos, como residuos ganaderos, puede ser relativamente importante. El procedimiento de cálculo es similar a los anteriores, considerando la concentración total como suma de las cuatro formas posibles, determinándose la concentración de cada una de las formas de ionización en función de la total, del pH y de la constante de disociación (ecuaciones 4.25, 4.26, 4.27, 4.28 y 4.29). K P3 K P1 K P2 H 3 PO4 o H H 2 PO4 o H HPO42 o PO43 H m m m >H 3 PO4 @T >H 3 PO4 @ ª«¬ H 2 PO4 º»¼ >HPO42 @ >PO43 @ >PO43 @ > @ 3 H KP KP KP 1 2 3 >HPO42 @ >H 3 PO4 @T >H @2 KP KP 2 3 > @ (4.26) >H @ 1 KP 3 >H 3 PO4 @T > @ > @ 3 2 § · H H H ¸ 3 ¨ 1 ¸¸ KP KP KP H ¨¨ K P1 K P2 K P3 2 3 3 © ¹ KP > @ (4.25) (4.27) 92 Simulación dinámica del pH >H 3 PO4 @T >H 2 PO4 @ > @ > @ >H 3 PO4 @ > @ > @ (4.28) > @ (4.29) 3 2 § · H H H ¸ 2 3 ¨ 1 ¸¸ 2 ¨¨ K P K P K P K K K P2 P3 P3 H 2 3 © 1 ¹ KP KP >H 3 PO4 @T > @ 1 2 > @ 3 H > @ 3 2 · § H H H ¸ 3 ¨ 1 ¸¸ ¨¨ K K K K K K P2 P3 P3 ¹ © P1 P2 P3 KP KP KP Por último se ha considerado la disociación del agua, expresada como el equilibrio entre el agua molecular y los protones e hidroxilos. H 2O KW o OH H m >OH @* >H @ KW (4.30) Las constantes de disociación (K) son las constantes de disociación aparentes, resultado de dividir la constante de disociación termodinámica por el factor de actividad. Estas constantes dependen de la temperatura pudiendo ser ajustadas a una curva polinomial (ecuación 4.31) a partir de los datos encontrados en la bibliografía (Lide, 1993; Angelidaki et al, 1993). Los valores de los estimadores se muestran en la Tabla 4.2. pK pK 0º C ) a·( T ) b·( T ) 2 c·( T )3 (4.31) No se tuvo en consideración la posible precipitación de sales, ni la actuación de cationes o aniones con más de dos cargas, así como los sulfuros. En cualquier caso, la adición de nuevas componentes resultaría relativamente sencilla, debido a la notación matricial empleada. 4.5.1. Notación La notación utilizada juega un papel muy importante y será básica para la posterior incorporación del algoritmo de cálculo de pH a un modelo de digestión anaerobia de substratos complejos. Cada una de las substancias consideradas presenta uno o más estados de ionización posibles, cuya concentración se indicará como Sik, donde k es el correspondiente estado de ionización y i es el subíndice que identifica la substancia en el sistema general. La concentración total de cada substancia es la suma de las concentraciones de todos las formas posibles. El número Optimización de la digestión anaerobia de purines de cerdo 93 máximo de grados de ionización considerados son 6, de +2 a –3, pasando por el estado neutro (carga cero). Tabla 4.2. Coeficientes para las expresiones de constantes de disociación pK(0º) a b c Acet.* 4.78 -0.002 6.00E-05 -2.00E-07 Prop.* 4.806 -0.001 8.00E-05 -4.00E-07 Butir.* 4.895 -0.002 7.00E-05 -3.00E-07 CO2(1)* 6.579 -0.013 0.0002 -8.00E-07 CO2 (2) 10.619 -0.014 0.0001 0 PO3 (1)* 2.0607 0.0025 0 0 PO3 (2)* 7.314 -0.007 0.0001 -6.00E-07 PO3 (3)* 12.658 0 0 0 NH4+* 10.07 -0.036 9.00E-05 4.00E-08 Agua* 14.93 -0.043 0.0002 -6.00E-07 Los marcados con * se calcularon mediante análisis de regresión a partir de los datos recogidos del Lide (1993), el resto fueron tomados de Angelidaki et al. (1993). Para facilitar la escritura se ha utilizado la notación matricial, definiendo el subíndice j, que hace referencia al estado de ionización, siendo j = 3-k, donde k es la carga con su signo. La concentración total, suma de las diferentes componentes, se escribe: Si ¦ S ik k 2 ,3 ¦ Si j , con j 3-k. (4.32) j 1,6 La concentración de cada estado de ionización se calcula en función del pH y de la concentración total de cada substancia, tal y como se expuso en las ecuaciones 4.16 a 4.30. Se pueden expresar como el producto de la concentración total Si y de un factor fij, función del pH (H+) y de las constantes de disociación aparente, que, a su vez, son función de la temperatura: S ik S ij Si x f ij . (4.33) S será el vector de concentraciones totales, con tantos elementos, n, como variables intervengan en el sistema. La matriz F , estará compuesta por los factores fij , y tendrá el mismo número de filas que el vector S (n) y tantas columnas como estados de ionización posibles, m. Se define un vector adicional, C , de m elementos, de factores de ponderación de las concentraciones en función de la carga. 94 Simulación dinámica del pH S F f ij Si ; S ij S ij ; m Si ¦ Si, j ; 1d i d n . 1d j d m (4.34) j 1 C j3 Cj k. La matriz F completa, compuesta por los elementos fij se muestra en la Tabla 4.3. Este tipo de notación matricial puede resultar muy útil para facilitar la comprensión del modelo y su posterior ampliación. Se ha encontrado una referencia reciente que utiliza una notación similar en un estudio del proceso de transferencia líquido-gas en sistemas anaerobios (Merkel y Krauth, 1999). Simulación de la transferencia líquido-gas Para la simulación del proceso de transferencia de gases se utilizó la teoría de la doble capa, determinando el caudal en función del gradiente entre la concentración en el líquido y la concentración de equilibrio en la capa límite. La concentración en la capa límite o de equilibrio se expresa como el producto de la correspondiente presión parcial en la fase gaseosa y la constante de solubilidad de Henry. La ecuación general, que permite estimar el caudal de gas, está definida por: N g ,i k La x V L x Si0 S*i K La x V L x Si · f ij Hei x pi , (4.35) donde Ng,i es el flujo de moles de la substancia i que pasan de la fase líquida a la fase gaseosa; Si0 es la concentración la substancia i no ionizada o volátil; fij es el factor que relaciona la concentración del estado de ionización j (en este caso 3) con concentración total en el medio líquido (Tabla 4.3); Si* es la concentración en el equilibrio, según la ley de Henry; pi es la presión parcial de la substancia i en el espacio de cabeza y Hei es la constante de Henry, que a su vez depende de la temperatura; VL es el volumen de líquido y KLa es la velocidad de transferencia líquido-gas (d-1), considerada constante para cada substancia. A partir de la ecuación 4.35, la tasa de transferencia líquido-gas rg,i, para la substancia i, queda definida por la siguiente expresión: rg ,i N g ,i VL K La x Si x f i ,3 Hei x pi (4.36) 0 22 (CO2,L) 0 0 0 0 0 1 0 26 (Z+) 27 (Z2+) 28 (Z-) 29 (Z2-) 30-32 0 0 1 0 0 0 0 23-24 25 (PO43-) 0 0 0 0 0 >H @ >H @ K171 +1 0 i +2 1-16 0 17 0 (NH4+) 18 0 (Val.) 19 0 (But.) 20 0 (Prop) 21 (Ac.) 0 0 0 0 0 1 K 252 K 253 >H @2 K 253 >H @3 K 251 K 252 K 253 1 0 0 0 0 K 252 K 253 >H @2 K 252 K 253 >H @ 1 0 K 251 K 252 K 253 K 251 K 252 K 253 >H @3 K 222 K 253 >H @ 1 >H @ 1 K 222 K 221 K 222 >H @2 >H @2 1 K 222 >H @ 1 >H @ K 211 >H @ K 211 K 201 >H @ K 201 K191 >H @ K191 K181 >H @ K181 >H @2 K 251 K 252 K 253 K 222 0 1 0 0 0 K 252 K 253 >H @2 K 253 >H @ 0 K 253 >H @ 1 >H @ 1 1 0 0 0 0 -2 0 0 K 221 K 222 >H @3 Estados de ionización (K) -1 0 0 >H @3 K 221 K 222 >H @2 K 221 K 222 >H @ K171 >H @ >H @ K181 >H @ >H @ K191 >H @ >H @ K 201 >H @ >H @ K 211 >H @2 K 171 0 1 K 251 K 252 K 253 >H @3 0 0 0 0 0 K 252 K 253 >H @2 1 0 0 0 0 0 0 -3 0 0 > @ H 1 K 253 Tabla 4.3. Matriz F: relación entre la concentración de cada estado de ionización y la concentración total para cada componente Optimización de la digestión anaerobia de purines de cerdo 95 96 Simulación dinámica del pH El coeficiente de Henry (Hei) depende de la temperatura. Se calcula, para NH3 y CO2 mediante una expresión polinómica (ecuación 4.37), tomada de Angelidaki et al. (1993), y para CH4 mediante la ecuación 4.38, tomada de Lide (1993). Los valores de los coeficientes de ambas expresiones, se muestran en la Tabla 4.4. Hei LnHei Ai Ai Bi x T Ci x T 2 Di x T 3 (4.37) Bi § T (º C ) 273 · Ci * ln¨ ¸ T (º C ) 273 100 K © ¹ 100 K (4.38) Tabla 4.4. Coeficientes para las expresiones de la constante de Henry A 0.0697 52.9 -60.954 CO2 NH3 CH4 B -0.002 -1.454 85.8073 C 2.56*10-5 0.021 23.4487 D -1.21*10-7 -1.13*10-4 - 4.5.2. Establecimiento de las ecuaciones para la simulación dinámica del pH La mayoría de los modelos utilizan para el cálculo del pH la ecuación del balance de cargas (Tabla 4.1). En el presente trabajo la expresión del balance ha sido adaptada de Angelidaki et al.(1993). Se define una nueva variable denominada Carga neta (Cn), que se define como: >H @ >OH @ >HCO 3 @ 2>CO 32 @ ¦ >Ai @ >H 2 PO 4 @ 2>HPO 42 @ (4.39) i 3>PO 43 @ >Z @ 2>Z 2 @ >NH 4 @ >Z @ 2>Z 2 @. Cn ( pH ) Utilizando la notación propuesta en el apartado 4.5.1, la ecuación 4.39 se puede escribir como: Cn( pH ) n 1 § 6 · i 1© j 1 ¹, ¦ ¨¨ S i • ¦ f ij • C j ¸¸ (4.40) Por tanto la carga neta, Cn(pH), es un escalar resultado de multiplicar escalarmente el vector traspuesto de concentraciones totales ( S t ), la matriz F de coeficientes y el vector C , de factor de carga, Cn( pH ) S t x F xC. (4.41) Optimización de la digestión anaerobia de purines de cerdo 97 La diferencia entre cargas positivas y negativas, que define la carga neta (ecuación 4.39), se compensa mediante la diferencia entre las dos componentes de ionización del agua. De la expresión anterior se puede definir una función, \, que relaciona la concentración de H+ con la carga neta, Cn, calculada a partir de las expresiones propuestas por Angelidaki et al.(1993), en función del pH del medio (ecuación 4.7 y 4.8): ªH º »¼ «¬ ­si >H @t >OH @ Cn(pH) t 0 ° 2 °\ \ Cn(pH) Cn 4KW 1 2 ° ° ȥ Cn(pH) ® °si >H @ >OH @ Cn(pH) 0 ° 2K W °\ \ 2 °¯ Cn( pH ) Cn 2 4 K W (4.42) . (4.43) A partir de la ecuación 4.43, se calcula la derivada de [H+] con respecto al tiempo, y se llega a la siguiente expresión, > @ d H dt d\ Cn dCn x dCn Bx dt dCn , dt (4.44) donde B es la derivada de la función \ respecto a Cn, y se calcula teniendo en cuenta si las condiciones son ácidas o básicas: si >H @ t >OH @ Cn( pH ) t 0 B · 1 §¨ Cn( pH ) 1¸ , ¸ 2 ¨ Cn 2 4 KW ¹ © > @ > 1 @ si H OH Cn( pH ) 0 B 2 (4.45) >H @ · § Cn( pH ) ¨ 1¸. (4.46) 2 ¸ 2 x KW ¨ Cn 4 KW ¹ © 2 La derivada de Cn(pH) se deduce a partir de la ecuación 4.41, de la siguiente forma: dCn( pH ) dt d §¨ S © t x F x C ·¸ ¹ dt · § § dS t · ¨ t ¨ x F x C¸ ¨S ¨ dt ¸ ¨ © ¹ ¨© x > @ ¸ d H dF .(4.47) xC¸ x ¸ dt d ªH º ¸ «¬ »¼ ¹ 98 Simulación dinámica del pH > @ La derivada de la matriz F , será la derivada con respecto a H de cada componente de la matriz, y se puede expresar como dF ª d Hº «¬ F f i' j ' »¼ § df i j ¨ ¨d H © > @ · ¸. ¸ ¹ (4.48) La matriz F’ se muestra en la Tabla 4.5. > @ La derivada de la concentración H respecto al tiempo se deduce reemplazando la ecuación 4.46 en la ecuación 4.44: > @ d H dt ­§ t ° dS B x ®¨ ¨ °̄© dt x · § F xC¸ ¨St ¸ © ¹ x F ' > @ · d H ½ ¾ . (4.49) dt ¿ ¹ xC¸• > @ De aquí se deduce la ecuación diferencial ordinaria de la variable H : > @ d H dt § § dS t · · ¸ x F xC¸ B • ¨¨ ¨ ¸¸ ¨ ¨ dt ¸ ¹ ©© ¹. · § t ' 1 B x ¨S x F x C¸ ¨ © ¸ ¹ (4.50) > @ El pH se calcula, finalmente, a partir de la concentración de H , pH > @ log10 H . (4.51) Con la expresión 4.50 se puede introducir el pH como variable de estado en el sistema de ecuaciones diferenciales ordinarias del modelo general. 4.6. Inclusión del algoritmo de simulación del pH en un modelo dinámico estructurado de digestión anaerobia previo Se adaptó el modelo de digestión anaerobia de substratos complejos propuesto por Angelidaki et al. (1999), en el que se introdujo la simulación dinámica de los parámetros físico químicos, sin modificar la simulación de los procesos biológicos. Aunque, en un primer momento se adoptó el modelo de Angelidaki et al. (1993), se modificó con el publicado por los mismos autores en el año 1999, ya que este último era mucho más completo, al considerar la acidogénesis de diferentes tipos de macromoléculas. K191 K 201 K 211 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 23-24 25 (PO43-) 26 (Z+) 27 (Z2+) 28 (Z-) 29 (Z2-) 30-32 22 (CO2,L) K181 K171 > @ > @ > @ > @ 0 0 0 0 1 2 ¹ · > @2 ¸¸ 3 § H 4 2H ¨ 3H ¨ K 25 K 25 K 253 2 3 © 1 > @ ¸ ¹ 3 2 · § H H H ¨ 1¸ ¸ ¨ K 25 K 25 K 25 K 252 K 253 K 253 1 2 3 ¹ © > @ K 251 K 252 K 253 1 > @ § · H ¨ 1¸ ¨ K 22 K 22 ¸ K 222 1 2 © ¹ 2 H 2 2 1 §¨ H 2 H K 222 ¨ K 221 K 222 K 221 © > @ >H @ K 211 2 >H @ K 201 2 >H @ K191 2 > @·¸ >H @ K171 2 >H @ K181 2 0 1 0 >H @ K171 2 K171 +1 0 > @ > @ > @ > @ 0 0 0 0 0 2 ¹ · > @¸¸ 2 3 § · H H H ¨ 1¸ ¨ K 25 K 25 K 25 ¸ K 252 K 253 K 253 1 2 3 © ¹ > @ 0 > @ 4 2 § 1 H H ¨ 2H K 252 K 253 ¨ K 251 K 252 K 253 K 253 © · § H 2 H ¨ 1¸ ¸ ¨ K 22 K 22 K 22 2 1 2 ¹ © 2 § · 2 ¨ H 1¸ ¨ K 22 K 22 ¸ 1 2 © ¹ > @ K 211 >H @ K 211 2 K 201 >H @ K 201 2 K191 >H @ K191 2 K181 >H @ K181 2 > @ 1 K 22 2 1 > @ · ¸ ¸ ¹ 0 > @ 2 > @ 0 0 0 0 0 > @ > @ 2 3 2 § · H 2 H ¨ 1¸ ¨ K 25 K 25 K 25 ¸ K 252 K 253 1 2 3 © ¹ > @ · § H 2 H ¨ 1¸ ¸ ¨ K 22 K 22 K 22 1 2 2 ¹ © > @ § 2H 1 ¨ ¨ K 22 K 22 K 222 1 2 © > @ 0 0 0 0 -2 0 0 3 2 § · H H H ¨ 1¸ ¨ K 25 K 25 K 25 ¸ K 252 K 253 K 253 1 2 3 © ¹ K 253 Estados de ionización (K) -1 0 0 Tabla 4.5. Matriz F’, derivada de la matriz F respeto a H+ i +2 1-16 0 17 0 (NH4+) 18 0 (Val.) 19 0 (But.) 20 0 (Prop) 21 (Ac.) 0 Simulación dinámica del pH 0 > @ > @ 0 0 0 0 0 > @ > @ 3 2 § · H H H ¨ 1¸ ¨ K 25 K 25 K 25 ¸ K 252 K 253 K 253 1 2 3 © ¹ 3H 2 H 1 K 251 K 252 K 253 K 252 K 253 K 253 > @2 0 0 0 0 0 -3 0 0 2 99 100 Simulación dinámica del pH 4.6.1. Procesos y variables considerados Se consideran dos tipos de procesos, procesos biológicos que implican la hidrólisis de las partículas orgánicas, el crecimiento de las poblaciones de microorganismos y la lisis de la biomasa, y los procesos físico-químicos, dentro de los cuales están la transferencia líquido-gas y la variación del pH. Se han considerado un total de 22 procesos (Tabla 4.6) y 33 componentes del sistema (Tabla 4.7). Tabla 4.6. Relación de procesos considerados j Proceso j Proceso 1 Hidrólisis de carbohidratos 12 Lisis de S2 2 Hidrólisis de proteínas 13 Lisis de S3 3 Acidogénesis de carbohidratos 14 Lisis de S4 4 Acidogénesis de aminoácidos 15 Lisis de S5 5 Hidrólisis de lípidos 16 Lisis de S6 6 Acetogénesis de AGCL 17 Lisis de S7 7 Acetogénesis valérico 18 Lisis de S8 8 Acetogénesis butírico 19 Transferencia líquido-gas CO2 9 Acetogénesis propiónico 20 Transferencia líquido-gas CH4 10 Metanogénesis acético 21 Transferencia líquido-gas NH3 11 Lisis de S1 22 Variación de H+ En la Figura 4.1 se muestra el esquema de los diferentes procesos considerados, así como las relaciones de inhibición, incluyendo también los procesos físico-químicos. A partir de las reacciones biológicas que ocurren en un reactor anaerobio, presentadas en la Tabla 4.8, se pueden estimar los coeficientes de producción de biomasa, o coeficientes estequiométricos, pudiéndose expresar con notación matricial, en unidades de mol/mol. El cálculo de los coeficientes estequiométricos de los procesos biológicos se realiza de la siguiente forma: Optimización de la digestión anaerobia de purines de cerdo 101 Tabla 4.7. Relación de componente consideradas i Variables i Variables 1 M. acidogénicos carbohidratos 18 Ácido Valérico 2 M. acidogénicos aminoácidos 19 Ácido Butírico 3 M. acidogénicos triglicéridos 20 Ácido Propiónico 4 M. acetogénicos AGCL 21 Ácido Acético 5 M. acetogénicos valérico 22 CO2 total líquido 6 M. acetogénicos butírico 23 CH4 total líquido 7 M. acetogénicos propiónico 24 H2S total 8 M. metanogénicos acético 25 Ortofosfórico (PO43-)T 9 Carbohidratos insolubles 26 Aniones (Z-) 10 Carbohidratos no degradables 27 Aniones (Z2-) 11 Carbohidratos solubles 28 Cationes (Z+) 12 Triglicéridos 29 Cationes (Z2+) 13 AGCL 30 Presión parcial CO2 (PCO2) 14 Proteínas insolubles 31 Presión parcial CH4 (PCH4) 15 Proteínas no degradables 32 Presión parcial NH3 (PNH3) 16 Proteínas solubles 33 H+ 17 Amonio total líquido Para un proceso j : b1S 2 b2 B2 ... bi Bi o b j X j ... bm Bm b j • PM Xj bi 1 PM S i Yi , j · Q i, j bi • PM S i Yi , j PM Xj b j (4.52) La matriz de coeficientes estequiométricos se completa con los coeficientes que relacionan la tasa de los procesos físico-químicos, con las diferentes variables, y se muestra en la Tabla 4.9. 4.6.2. Cinética de los procesos biológicos considerados La cinética utilizada para la simulación de los procesos biológicos se adaptó de Angelidaki et al. (1999). Para la mayoría de los procesos biológicos se utilizó la cinética de Monod para describir la influencia del substrato, modificada por una o varias funciones de inhibición, y una función del pH. Angelidaki et al. (1999) consideran dos substratos limitantes para el crecimiento de microorganismos, el substrato orgánico 102 Simulación dinámica del pH que corresponda y el nitrógeno amoniacal como principal nutriente. MATERIA ORGÁNICA INSOLUBLE LÍPIDOS CARBOHIDRATOS PROTEINAS MATERIA ORGÁNICA NO DEGRADABLE HIDRÓLISIS ACIDOS GRASOS DE CADENA LARGA CARBOHIDRATOS SOLUBLES AMINOÁCIDOS ACIDOGÉNESIS AGV (Valer,Prop., But.) ACETOGÉNESIS ACÉTICO LISIS H2 METANOGÉNESIS CO2-HCO3--CO32(LIQ) METANO (LIQ) NH4+ CH4 (g) TRANSFERENCIA GAS-LÍQUIDO ANIONES y CATIONES CO2 (g) NH3 pH VARIACIÓN pH Proceso de inhibición Variables que influyen sobre el pH Procesos Figura 4.1. Diagrama de flujo del modelo general de digestión anaerobia Optimización de la digestión anaerobia de purines de cerdo 103 Tabla 4.8. Reacciones biológicas consideradas Nº Proceso 1 Hidrólisis de carbohidratos 2 Hidrólisis de proteínas 3 Fermentación carbohidratos solubles Reacción C 6 H 10 O5 is o YC C 6 H 10 O5 S 1 YC · C 6 H10 O5 in Pr oteínas is o Y P aminoácidos S 1 Y P · Pr oteínas in C 6 H 10 O5 S 0.1115 NH 3 o 0.1115C 5 H 7 NO2 0.744CH 3 COOH 0.5CH 3 CH 2 COOH 0.4409CH 3 CH 2 CH 2 COOH 0.6909CO 2 0.0254 H 2 O 4 Fermentación de aminoácidos CH 2.03O0.6 N 0.3S0.001 0.3006H 2O o 0.017013C5 H 7 NO2 0.29742CH 3COOH 0.02904CH 3CH 2COOH 0.02283CH 3 CH 2 2 COOH 0.013202CH 3 CH 2 3 COOH 0.0753CO2 0.283NH 3 0.001H 2 S 5 Degradación de triglicéridos C 57 H 104 O6 1.90695 H 2 O 0.04071NH 3 0.0291CO2 o 0.04071C 5 H 7 NO2 0.941843CH 3 CH 2 COOH 3C18 H 34 O2 6 Degradación de AGCL (oleico) C18 H 34 O 2 7.7401H 2 O 0.2537 NH 3 4.0834CO 2 o 0.2537C 5 H 7 NO 2 8.6998CH 3 COOH 3.4139CH 4 7 Acetogénesis de valérico CH 3 CH 2 3 COOH 0.0653 NH 3 0.5543CO2 0.8045 H 2O o 0.0653C5 H 7 NO2 0.8912CH 3COOH 0.02904CH 3CH 2COOH 0.4454CH 4 8 Acetogénesis a partir de butírico CH 3 CH 2 2 COOH 0.0653NH 3 0.5543CO2 0.8038 H 2O o 0.0653C5 H 7 NO2 1.8909CH 3COOH 0.4452CH 4 9 Acetogénesis a partir de propiónico CH 3CH 2COOH 0.06198 NH 3 0.314 H 2O o 0.06198C5 H 7 NO2 0.9345CH 3COOH 0.6604CH 4 0.1607CO2 10 Metanogénesis acetoclástica 11- Lisis de 18 microorg. CH 3COOH 0.022 NH 3 o 0.022C 5 H 7 NO 2 0.945CH 4 0.945CO2 0.066 H 2 O C5 H 7 NO2 1,3896 H 2O o 3,33CH 2.03O0.6 N 0.3S0.001 is C6 H10O5 is En el presente modelo no se ha considerado el nitrógeno como limitante, puesto que los residuos ganaderos, substrato para los que se ha ideado el presente modelo, son muy ricos en nitrógeno. La hidrólisis de carbohidratos y proteínas se simuló mediante cinética de primer 104 Simulación dinámica del pH orden, de forma que la velocidad de reacción depende de la concentración de carbohidratos y proteínas en el medio. El proceso se consideró afectado por la concentración de ácidos grasos volátiles (AGV), mediante una función de inhibición no competitiva reversible (INCR), que afecta a la tasa específica máxima de hidrólisis. La acidogénesis de carbohidratos solubles se simuló mediante la cinética de Monod, inhibida tan solo por la concentración de ácidos grasos de cadena larga (AGCL), con una cinética de tipo INCR. No se considera sensible al pH. La hidrólisis-acidogénesis de triglicéridos se simula mediante cinética de Monod, con inhibición no competitiva reversible por AGCL. La acetogénesis a partir de AGCL se simuló mediante una cinética de Haldane, considerando la inhibición por el substrato. La acetogénesis de ácidos grasos volátiles (AGV) se simuló mediante la cinética de Monod, y se consideró inhibición no competitiva reversible por AGCL y por la concentración de acético total. Se realizó la simplificación de considerar la metanogénesis a partir de hidrógeno englobada dentro del proceso de acetogénesis, al considerar que no es una etapa limitante, siguiendo a Angelidaki et al. (1999). La metanogénesis se simuló mediante cinética Monod y con inhibición no competitiva reversible por amoníaco libre y AGCL. Las tasas específicas máximas de los procesos acetogénicos y metanogénicos y acidogénesis de triglicéridos dependen del pH, relacionándose mediante la función F(pH), que es una función de tipo Michaelis normalizada (Angelidaki et al., 1993), F ( pH ) 1 2 x 10 0.5 pK l pK h 1 10 pH pK h 10 pK l pH , (4.53) donde pKl y pKh son los valores de pH para los cuales la tasa de crecimiento es un 50% de la máxima. El valor de estos parámetros son iguales para todos los procesos, pKl=6 y pKh=8,5 (Angeldiaki et al., 1993). Por último, se consideró la lisis de microorganismos como un proceso más, simulado mediante cinética de primer orden, función de la concentración de biomasa. Como producto de la lisis se obtiene materia orgánica insoluble, con una fracción de carbohidratos y proteínas, que entran a formar parte de la materia orgánica disponible para la degradación. Las tasas máximas de reacción se consideraron dependientes de la temperatura, siguiendo la función de variación propuesta por Angelidaki et al. (1993). 0 0 0 0 0 0 1.00 0 0 0 0 0 0 0 0 0 -1.00 0 0 -16.134 15.077 2.593 10.665 0 0 0 0 0 0 0 0 0 0 9 Proceso 11 12 13 14 15 16 17 18 19 20 21 22 0 -1.00 0 0 0 0 0 0 0 0 0 0 0 0 0 -1.00 0 0 0 0 0 0 0 0 0 0 0 0 0 -1.00 0 0 0 0 0 0 0 0 0 0 0 0 0 -1.00 0 0 0 0 0 0 0 0 0 0 0 0 0 -1.00 0 0 0 0 0 0 0 0 0 0 0 0 0 -1.00 0 0 0 0 0 0 0 0 0 0 0 0 0 -1.00 0 0 0 0 0 1.00 0 0 0 0 0 0 0 -1.00 0 0 0 0 0 0.2783 0.2783 0.2783 0.2783 0.2783 0.2783 0.2783 0.2783 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 3.3333 3.3333 3.3333 3.3333 3.3333 3.3333 3.3333 3.3333 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 -1.00 0 0 0 0 0 0 0 0 0 0 1.00 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 -45.455 0 0 0 0 0 0 0 0 0 0 0 0 42.955 0 0 0 0 0 0 0 0 1.00 0 0 0 42.955 0 0 0 0 0 0 0 0 0 1.00 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 -CGL 0 0 0 0 0 0 0 0 0 0 0 0 0 -CGL 0 0 0 0 0 0 0 0 0 0 0 0 0 -CGL 0 0 0 0 0 0 0 0 0 0 0 0 0 1.00 10 Tabla 4.9. Matriz de coeficientes estequiométricos Variable 1 2 3 4 5 6 7 8 1 0 0 1.00 0 0 0 0 0 2 0 0 0 1.00 0 0 0 0 3 0 0 0 0 1.00 0 0 0 4 0 0 0 0 0 1.00 0 0 5 0 0 0 0 0 0 1.00 0 6 0 0 0 0 0 0 0 1.00 7 0 0 0 0 0 0 0 0 8 0 0 0 0 0 0 0 0 9 -1 0 0 0 0 0 0 0 10 1-Yc 0 0 0 0 0 0 0 11 Yc 0 -8.969 0 0 0 0 0 12 0 0 0 0 -24.562 0 0 0 13 0 0 0 0 73.687 -3.942 0 0 14 0 -1.00 0 0 0 0 0 0 15 0 1-Yp 0 0 0 0 0 0 16 0 Yp -58.778 0 0 0 0 17 0 0 -1.000 16.633 -1.00 -1.00 -1.00 -1.000 18 0 0 0 0.769 0 0 -15.314 0 19 0 0 3.954 1.342 0 0 0 -15.314 20 0 0 4.484 1.707 23.134 0 0.445 0 21 0 0 6.673 17.482 0 34.292 13.648 28.957 22 0 0 6.196 4.424 -0.715 -16.095 -8.489 -8.489 23 0 0 0 0 0 13.456 6.821 6.818 24 0 0 0 0.059 0 0 0 0 25 0 0 0 0 0 0 0 0 26 0 0 0 0 0 0 0 0 27 0 0 0 0 0 0 0 0 28 0 0 0 0 0 0 0 0 29 0 0 0 0 0 0 0 0 30 0 0 0 0 0 0 0 0 31 0 0 0 0 0 0 0 0 32 0 0 0 0 0 0 0 0 33 0 0 0 0 0 0 0 0 Simulación dinámica del pH 105 106 Simulación dinámica del pH Tasas de reacción A partir de las expresiones de la tasa de crecimiento de los diferentes microorganismos (Tabla 4.10) y de los coeficientes estequiométricos se pueden establecer las expresiones de tasas de producción o consumo biológico de cada componente. De forma general las tasas correspondientes a los procesos biológicos, de crecimiento de microorganismos, vendrán dadas por: rj P j x X k , (4.54) donde Xk es la concentración de microorganismos responsables del proceso j. La tasa específica de crecimiento sigue diferentes expresiones dependiendo del proceso, tal y como se muestra en la Tabla 4.10. En esta tabla se muestran, además, las tasas correspondientes a los procesos físicoquímicos, como transferencia líquido-gas y variación del pH. Los valores de los diferentes parámetros cinéticos se tomaron del modelo de Angelidaki et al. (1993, 1996 y 1999), y se muestran en la Tabla 4.11. 4.6.3. Establecimiento del ordinarias sistema de ecuaciones diferenciales A partir de la tasa de reacción de los diferentes procesos físico-químicos y biológicos, y de los coeficientes estequiométricos que relacionan el consumo o producción de una componente con la velocidad de los procesos, se pueden establecer las tasas de variación en el tiempo de cada variable. En función del tipo de reactor las ecuaciones generales serán diferentes. En el trabajo actual se ha adaptado el modelo para sistemas de mezcla completa continuo y discontinuo (Figura 4.2). Reactores continuos de mezcla completa Aplicando la ecuación del balance de masas a cada una de las variables (Si) de la fase líquida, se obtiene la siguiente expresión: dS i dt 1 · S i S i ¦ Q i , j x r j ; TR TR 0 j 1,m VL , QL i=1...29 (4.66) donde Si0 es la concentración de la variable i en el influente, TR es el tiempo de retención hidráulico, QL es el caudal de influente, VL es el volumen de la fase líquida, que se mantiene constante a lo largo del tiempo. La ecuación 4.65 se complementa con las condiciones iniciales. Optimización de la digestión anaerobia de purines de cerdo 107 Tabla 4.10. Cinéticas de reacción de los procesos biológicos y físico-químicos considerados Proceso (j) Modelo Cinético 1.-Hidrólisis de carbohidratos r1 r2 2.- Hidrólisis de proteínas P m ,3 r3 3.- Acidogénesis de glucosa 4.- Acidogénesis de aminoácidos P m ,5 5.- Paso lipolítico 6.- Acetogénesis de AGCL 7.- Acetogénesis de AGV (j=7,8,9) 10.- Metanogénesis acetoclástica 11.- Lisis celular (j=11, 12, 13, 14, 15, 16, 17 y 18) 19.- Transferencia líquido-gas (j=19, 20 y 21) 22.- Variación del pH r6 r11 k i ¦ AGV S11 • K S11 S11 S12 • K S12 S12 1 x S9 (4.55) x S14 (4.56) 1 S13 • S1 (4.58) 1 1 S13 K S13 S13 K La , j x • F pH • S 4 (4.60) K I , AGCL 1 S13 1 1 1 S13 K I , AGCL 1 • F pH • S j 2 (4.61) K I ,AH 1 • 1 >NH 3 @ K I , NH • F pH • S 8 (4.62) 3 0,05 x P m, j 8 x S j 10 (4.63) si j 19 k 22 ; i 30 si j 20 k 23 ; i 31 (4.64) si j 21 k 17 ; i 32 §§ t · · ¨ dS ¸ ¸ B • ¨¨ x F xC¸ ¨ ¸ ¨ dt ¸ ¹ ©© ¹ § t · ' 1 B x ¨¨ S x F x C ¸¸ © >AH @ • K I ,AGCL S k x f k ,3 Hei x S i ; r22 (4.59) K I , AGCL S13 S j 11 • K S S j 11 1 j 10 S 21 • P m ,11 K S S 21 21 • F pH • S 3 rj rj (4.57) K I , AGCL 1 P m ,6 Pm ki k0 1 rj ki ¦ AGV P m ,4 • S16 • S 2 r4 r5 ki k0 ¹ (4.65) 108 Simulación dinámica del pH Tabla 4.11. Valores de los parámetros cinéticos utilizados en el modelo µ max (d-1) 1a KS (g/L) KI,a (g/L) - 0.33 a (AGV) 1a - 0.33 a (AGV) 3 Acidogénesis de glucosa 5.1 a 0.5 a (Glc) 4 Acidogénesis de aminoácidos 6.38 a - 5 Hidrólisis de lípidos 2.86 b 0.02 b (Lip) 6 Acetogénesis de AGCL 0.57 b 0.04 b (AGCL) 7 Acetogénesis del valérico 0.69 a 0.175 a (Val) 0.40 a (Ac) 0.20b 8 Acetogénesis del butírico 0.67 a 0.176 a (But) 0.72 a (Ac) 0.20b 9 Acetogénesis del propiónico 0.49 a 0.96 a (Ac) 0.20b Proceso 1 Hidrólisis de carbohidratos 2 Hidrólisis de proteínas KI,b (AGCL) (g/L) 0.20b 0.259 a (Pr) 0.20b 0.20b,c 10 Metanogénesis acetoclástica 0.6 a 0.120 a (Ac) 0.26 a (NH3) 0.20b b c : de Angelidaki et al. (1999); : de Angelidaki et al. (1996); : Inhibición tipo Haldane ,resto: tipo no competitiva reversible. a Biogas Influente Efluente Biogas VG VG VL VL Sistema continuo Sistema discontinuo Figura 4.2. Esquema de sistemas continuos y discontinuos La simulación de la fase gaseosa se tomó de Costello et al. (1991), aplicando también un balance de materia, quedando de la siguiente forma: dS i dt Qg Vg x Si ¦Q i , j x r j ; i=30...32 (4.67) j 1,m donde Si es la presión parcial, en atmósferas, de la componente i, Qg es el caudal de gas generado, en L gas/día y Vg el volumen del espacio de cabeza (L). La relación entre la tasa del proceso de transferencia líquido-gas, en unidades de concentración en la fase líquida (M/d), y la variación de la Optimización de la digestión anaerobia de purines de cerdo 109 presión parcial en la fase gaseosa (atmósferas/día), CGL, se calcula asumiendo el cumplimiento de la ley de los gases perfectos, a partir de la siguiente expresión: CGL VL Vg xRx 273 Temp(º C ) . (4.68) El caudal de gas se estima a partir de las tasas de transferencia líquido-gas, considerando que la presión total del sistema se mantiene constante e igual a 1 atmósfera, mediante la siguiente expresión: Qg § · ¨ ¸ x r Q ¦ i, j j¸; ¨¨ 1 PH 2O ¸ j 1,m ¹ i 30,32 © Vg ¦ (4.69) La matriz de coeficientes estequiométricos se muestra en la Tabla 4.9. La ecuación 4.66 se puede generalizar mediante notación matricial a:. §Q· ¨¨ ¸¸ ©V ¹ dS dt dS § dS i · , ¸ ¨ dt ¨© dt ¸¹1di dn S 0 S 0i ,S Q 1didn Q ij 1didn , 1d j d m r t x S 0 S Q x r S i 1didn , r j 1d jdm , ­ QL ° VL Q °° Qg =® V ° Vg °0 °¯ (4.70) if 1d i d n 3 , if n 2 d i d n 1 , if i n . donde Q es la matriz de coeficientes estequiométricos generalizada, que considera tanto los procesos biológicos, como los procesos físico-químicos y r es el vector de tasas de reacción de todos los procesos considerados (Tabla 4.10). Reactor discontinuo En el sistema discontinuo no hay entrada y salida de masas durante el proceso, por lo que la ecuación para las variables de la fase líquida se expresa mediante, dS i dt ¦Q i , j x r j j 1,m . (4.71) Para la simulación de la fase gaseosa no se considera salida de gas, por lo que la presión total del sistema va aumentando a medida que se desarrolla el proceso. Por tanto, las ecuaciones para las variables de la fase gaseosa siguen la misma expresión que en las de las fase líquida (ecuación 4.71). 110 Simulación dinámica del pH La ecuación general en notación matricial (ecuación 4.70) puede ser usada también para el reactor discontinuo, considerando que el caudal de influente y de gas es igual a cero. 4.6.4. Método numérico aplicado para la aproximación de las soluciones La resolución numérica del sistema de ecuaciones diferenciales ordinarias se realizó utilizando el método de Runge-Kutta-Fehlberg, que consiste en calcular la solución por dos métodos diferentes, con orden de error diferentes. Si el paso de integración es pequeño, la diferencia entre los resultados obtenidos es una buena aproximación al error global. Se ha utilizado el método RKF45, que combina métodos Runge Kutta de 4 y 5 pasos, en el que la función se evalúa 6 veces. Se utilizó una variante de esta rutina, denominada adaptativa, que adapta el tamaño del paso de integración en función del error cometido. De esta forma, se consigue aumentar tanto la rapidez de la simulación, en caso de sistemas estables, como la precisión en las soluciones, en el caso de sistemas con grandes variaciones. El pH se considera como una variable de estado, y la ecuación diferencial que define su variación es una más del sistema de ecuaciones diferenciales ordinarias, utilizándose el método RFK45 para su resolución. Los modelos tradicionales, al considerar el pH como un parámetro y no como una variable, lo calculaban en cada paso de integración, utilizando algún algoritmo de resolución numérica, por ejemplo el método de la secante (Angelidaki et al., 1993). Esto hace que si se producen cambios importantes en el pH del reactor y el paso de integración no es suficientemente pequeño, se pueden producir problemas de convergencia, además de hacer disminuir la velocidad de cálculo. Para realizar las simulaciones se han escrito los programas en lenguaje FORTRAN 77, compuesto de varias subrutinas. Los programas completos se muestran en los Anejos 1 y 2. 4.7. Validación del algoritmo de simulación dinámica del pH Dada la complejidad de los sistemas anaerobios, la correcta validación del algoritmo de simulación de una variable concreta, como el pH, se hace muy complicada, pudiendo quedar enmascarada con otros efectos. Por ello se realizó la validación de la parte físico-química por separado del resto del Optimización de la digestión anaerobia de purines de cerdo 111 modelo estructurado. Se desarrolló un modelo muy sencillo, en el que se conociera con exactitud la concentración y variación de la concentración de cada una de las substancias consideradas en cada momento. Se trata de una solución acuosa a la que se van añadiendo, a una determinada y conocida velocidad, una serie de soluciones concentradas de diferentes substancias, ácidos o bases, que influyen en el pH (Figura 4.3). Se han seleccionado las variables que mayoritariamente intervienen en el proceso anaerobio, como los ácidos grasos volátiles, dióxido de carbono, amonio, ortofosfórico, etc. Medida on-line pH y temperatura Q Influente Si0 V Si Figura 4.3. Esquema del dispositivo experimental utilizado para la validación del algoritmo de simulación dinámica del pH 4.7.1. Establecimiento de las ecuaciones de simulación Se desarrolló el algoritmo de simulación y el programa informático, teniendo en cuenta su posterior adaptación a un modelo dinámico estructurado digestión anaerobia (modelo de Angelidaki et al., 1999). Por tratarse de una solución acuosa sencilla no se consideraron procesos biológicos, ni la posible precipitación de sales. Sí se consideraron procesos de transferencia líquidogas, ya que resulta fundamental para simular la variación del pH en presencia de bicarbonato. Se consideraron amonio, acetato, fosfato, carbonato y aniones. La variación de la concentración de cada substancia en el tiempo depende de la concentración del influente (Si0), del volumen actual de la disolución (VL) y de los procesos que se estén desarrollando en el sistema. Se consideró la variación de la concentración de las substancias añadidas (ácidos y bases) lineal a lo largo del tiempo, siguiendo la siguiente expresión: d V L x S i dt Qi x S i 0 i 1,.., n . (4.72) A partir de la ecuación 4.72 y teniendo en cuenta las ecuaciones 4.73 y 112 Simulación dinámica del pH 4.74, se establecen las ecuaciones diferenciales ordinarias para las variables de la fase líquida (ecuación 4.75), incluyendo el efecto de determinados procesos físico-químicos (transferencia líquido-gas) sobre la concentración: dV L dt (4.73) QT V0 QT x t VL QT (4.74) ¦ Qi (4.75) i 1,n dS i dt S i0 x Qi S i x QT V0 QT x t ¦Q i , j r j i j 1,m 1,.., n ; (4.76) dónde V0 es el volumen inicial de disolución, QT es el caudal de influente, rj es la tasa del proceso j, Qi,j es un coeficiente que relaciona la variación de la substancia i con el proceso j, Qi es el caudal de la componente i, n es el número total de componentes del sistema y m es el número total de procesos considerados. Las variables de la fase gaseosa, ideadas más para el modelo de digestión anaerobia que para este modelo sencillo, son la presión parcial de las tres componentes consideradas, CH4, CO2 y NH3. Los únicos procesos físicoquímicos considerados fueron la transferencia líquido-gas de las citadas variables. Para obviar el resto de procesos, se hicieron 0, los coeficientes estequiométricos Qi,j.. La concentración de [H+] se consideró como la variable n del sistema, y su ecuación diferencial se escribió utilizando la ecuación 4.50. Utilizando la notación matricial, la ecuación 4.76, se puede expresar como: dS dt Q VL x S 0 S Q x r (4.77) En un sistema cerrado como el anaerobio discontinuo (Figura 4.2), la simulación de la variación de la presión parcial en el espacio de cabeza puede ser decisiva para la correcta simulación de todo el proceso. Sin embargo en un sistema abierto, como el utilizado en la validación del algoritmo del pH, la presión parcial en el espacio de cabeza, de los gases transferidos desde la fase líquida, será aproximadamente cero, por lo que se podría considerar constante. 4.7.2. Dispositivo experimental para la validación del algoritmo El esquema del dispositivo experimental utilizado para la validación del Optimización de la digestión anaerobia de purines de cerdo 113 algoritmo se muestra gráficamente en la Figura 4.3. El sistema se probó para mezclas secuenciales de las substancias siguientes: hidróxido amónico, ortofosfato y acético. Se realizaron cinco experimentos (a-e), en los que se alternó la adición de los citados compuestos para provocar cambios fuertes de tendencia y estudiar el ajuste del modelo en estados transitorios. El caudal, Q, de introducción de las soluciones concentradas fue fijo, de 1 mL/min, y el volumen inicial fue, en todos los casos, de V0 de 100 mL de agua desionizada, y las concentraciones iniciales, Si, fueron cero. El experimento se realizó a temperatura ambiente, alrededor de 20ºC. Las concentraciones de las tres soluciones concentradas, Si0, se muestran en la Tabla 4.12. En los casos en los que se adicionaron varios compuestos simultáneamente, se hizo adicionando todas las soluciones correspondientes, de forma que el caudal total fue la suma de los caudales de cada una de las soluciones individuales. El pH y la temperatura se midieron de forma continua, conforme se fueron adicionando las diferentes soluciones. Los resultados de estos cinco experimentos se muestran en la Figura 4.4a a 4.4e. El orden de introducción de las diferentes soluciones concentradas es el que se indica en la figura. Tabla 4.12. Componentes utilizadas para los diferentes experimentos realizados (Concentraciones expresadas en Moles/L). Resultados y momento de aplicación en Figura 4.4 Conc. Influente Conc. Inicial NH4OH HCl Acético PO4H3 Exp. a 0 0,5 0,50 1,00 Exp. b 0 0,50 0,50 1,00 Exp. c 0 0,50 0,50 1,00 Exp. d 0 0,50 0,50 1,00 Exp. e 0 0,50 0,50 1,00 1,00 Exp. f 0,23 (CaCO3) Se realizó un experimento adicional, en la que se partió de una solución de carbonato cálcico, de 0,23 M, a la que se añadió HCL, con una concentración 1N. El caudal utilizado en este experimento no fue constante, sino que se fue añadiendo el ácido conforme se fue estabilizando el pH. Se anotaron los tiempos, y se realizó una regresión para establecer la función polinómica que determina el caudal. Fue necesario realizar una ligera modificación del programa para poder tener en cuenta la función que determina el caudal. La comparación de los valores de pH reales y simulados se muestra en la Figura 4.4f. 114 Simulación dinámica del pH pH Como se observa en la Figura 4.4, los valores de pH simulados mediante el algoritmo se ajustaron muy bien a los valores medidos, con valores del coeficiente de regresión (r2) muy altos, por encima de 0,9. PO4 H3 10 9 Acético 8 2 7 r =0,995 6 NH4 OH 5 4 3 2 1 0 -4 6 16 26 12 NH4 OH 5 8 1 0 36 Tiempo PO4 H3 2 0 10 15 20 25 0 e 5 10 15 20 25 Tiempo d NH4 OH+Acético 10 9 8 7 6 5 4 3 2 1 0 30 Tiempo 6 2 r =0,940 0 2 4 6 8 HCl 14 10 Tiempo f 12 PO4 H3 +Acético 5 2 r =0,998 2 4 5 NH4 OH 3 6 0 Acético 4 Acético 2 b 6 c r =0,964 PO4 H3 +NH4 OH + Acético NH4 OH 8 PO4 H3 7 Acético PO4 H3 10 pH 9 a r2=0,963 10 4 8 3 2 6 r =0,986 2 4 1 2 0 0 0 2 4 6 8 10 Tiempo pH medido 0 5 10 15 Tiempo pH simulado Figura 4.4. Comparación de los valores de pH medidos y simulados, correspondientes a las condiciones de la Tabla 4.12 y a los momentos de introducción indicados de cada componente Optimización de la digestión anaerobia de purines de cerdo 115 Se puede concluir, por tanto que el algoritmo de simulación de pH es válido para simular el pH de soluciones acuosas en estados transitorios, por lo que puede ser incluido en el sistema anaerobio complejo. 4.8. Simulaciones con el modelo general de digestión anaerobia. El modelo del proceso anaerobio con inclusión de la simulación dinámica de los parámetros físico-químicos no ha sido calibrado en el marco del trabajo actual, por lo que no se ha podido realizar la validación del mismo. Sin embargo, sí se puede utilizar como herramienta cualitativa para prever el funcionamiento de un reactor anaerobio que trabaje con substratos complejos. El objetivo general del trabajo de tesis es la optimización del proceso anaerobio mediante la introducción de residuos orgánicos de la industria agroalimentaria como cosubstrato. Poder tener una aproximación, aunque sea cualitativa, de la respuesta del sistema anaerobio frente a la introducción de dichos residuos es, por tanto, muy interesante, ayudando en la toma de decisiones y a la planificación de los experimentos. Se realizaron algunas simulaciones para estudiar el funcionamiento del modelo frente a la introducción de los dos cosubstratos que se utilizarán posteriormente en la fase experimental (capítulos 5 y 6). La composición de los diferentes substratos utilizados se muestra en la Tabla 4.13. El purín 1 se utilizó como base de las mezclas. Los tratamientos Rpera 5%, 12,5% y 20%, consisten en un 5%, 12,5% o 20%, respectivamente, de residuo de pera y un 95%, 87,5% o 80% de purín 1. De la misma forma, los tratamientos TDO 1%, 2,5% o 5%, significa la mezcla de purín 1 con un 1%, 2,5% o 5%, respectivamente, de tierras decolorantes de aceite de oliva. El cosubstrato Rpera, produce un aumento en la concentración de carbohidratos, mientras que TDO produce un incremento en la concentración de lípidos. Se utilizó también un nuevo tipo de purín de las mismas características que el primero, pero con un mayor contenido de amonio (purín 2), para realizar simulaciones adicionales. Para conseguir el estado estacionario, se impusieron unas condiciones iniciales, especialmente en la concentración de biomasa, resultado de una larga simulación con Purín 1. 116 Simulación dinámica del pH Tabla 4.13. Composición de los diferentes substratos utilizados en las simulaciones (Concentraciones en g/L) Purín 1 Rpera Rpera Rpera TDO TDO TDO Purín 2 5% 12,5% 20% 1% 2,5% 5% Carbohidratos 44.49 56.30 73.97 91.67 45.10 46.02 47.55 44.49 Lípidos 2.93 2.93 2.93 2.93 5.44 9.21 15.48 2.93 Proteínas 11.43 11.43 11.43 11.43 11.35 11.22 11.01 11.43 N-NH4+ 3.43 3.43 3.43 3.43 3.20 3.20 3.20 4.50 Valérico 0.50 0.50 0.50 0.50 0.50 0.50 0.50 0.50 Butírico 0.60 0.60 0.60 0.60 0.60 0.60 0.60 0.60 Propiónico 1.50 1.50 1.50 1.50 1.50 1.50 1.50 1.50 Acético 3.10 3.10 3.10 3.10 3.10 3.10 3.10 3.10 PO430.80 0.80 0.80 0.80 0.80 0.80 0.80 0.80 Z 2.00 2.00 2.00 2.00 2.00 2.00 2.00 2.00 Z21.20 1.20 1.20 1.20 1.20 1.20 1.20 1.20 Z+ 4.30 4.30 4.30 4.30 4.30 4.30 4.30 4.30 Z2+ 1.20 1.20 1.20 1.20 1.20 1.20 1.20 1.20 El primer grupo de simulaciones consistieron en la introducción en un sistema estable, alimentado con purín 1 durante 10 días, de los diferentes substratos que se muestran en la Tabla 4.13. Para no provocar sobrecargas, en el caso de TDO 5%, se simuló la introducción de este substrato de forma gradual. En todos los casos, el tiempo de retención de trabajo fue 15 días y la temperatura de 35ºC. Los diferentes valores de producción volumétrica de biogás y de producción de metano por unidad de carga obtenidos se muestran en la Tabla 4.14. Tabla 4.14. Resultados obtenidos mediante el modelo de simulación, para diferentes tipos de cosubstratos en el rango mesofílico (35ºC) M B Substrato (L Biogás/L*d) (L CH4/g DQO) Purín1 1,86 0,199 Rpera 5% 2,17 0,193 Rpera 12,5% 2,64 0,187 Rpera 20% 3,11 0,182 2,16 0,212 TDO 1% TDO 2,5% 2,57 0,228 TDO 5% 3,27 0,248 Purín 2 1,72 0,196 La producción volumétrica de biogás, aumenta siempre que se introduce un Optimización de la digestión anaerobia de purines de cerdo 117 cosubstrato, debido al aumento de la materia orgánica en el influente. Sin embargo, la producción de metano por unidad de DQO depende del tipo de cosubstrato utilizado, de forma que los lípidos la hacen aumentar, mientras que los materiales lignocelulósicos la hacen disminuir (Tabla 4.14). No se ha observado ningún efecto grave de sobrecarga al introducir materiales lignocelulósicos, que sin embargo sí sucede en los experimentos en discontinuo con muy alta concentración de residuo de pera (capítulo 5). Esta discrepancia se debe a la no consideración del grupo metanogénico hidrogenotrófico de forma independiente. El efecto de sobrecarga consiste principalmente en un desequilibrio entre la velocidad de producción de H2, a partir de propiónico, butírico y valérico, y la velocidad de consumo del mismo por parte de los metanogénicos hidrogenotróficos. En futuros desarrollos debería introducirse este proceso en el modelo. Al comparar la producción de biogás y de metano con dos purines con diferente concentración de amonio, purín 1 y purín 2, en el rango mesofílico, se observa también una ligera disminución de la producción de gas (Tabla 4.14), y una ligera acumulación de ácidos grasos volátiles (datos no mostrados), aunque poco importante. Estos mismos substratos en el rango termofílico obtuvieron muy diferentes producciones de gas, debido a la inhibición por amonio del Purín 2, que obtuvo una menor producción de metano y de biogás que el Purín 1, y una concentración relativamente alta de ácidos grasos volátiles (Figura 4.5). El sistema, a pesar de la inhibición, mantiene una cierta producción de metano, aunque menor que para el purín 1 (0.093 L CH4/g DQOini, frente a 0,183 L CH4/g DQOini). Al introducir el purín 2, con una mayor concentración de amonio, el pH aumenta, por la mayor alcalinidad de dicho substrato, debida precisamente a la mayor concentración de amonio. Sin embargo, conforme pasa el tiempo se van acumulando AGV, especialmente acético, y se produce una ligera bajada del pH. Esta bajada del pH hace que se pueda mantener el proceso con una cierta producción de metano, ya que disminuye ligeramente la concentración de amoníaco libre. Así pues, este complejo fenómeno, descrito en la bibliografía (Angelidaki et al., 1993 y Bonmatí, 1998), y comprobado en los experimentos llevados a cabo en el presente trabajo (capítulo 6), ha sido correctamente simulado con el modelo desarrollado, siendo de especial interés la correcta simulación del pH. El pH, aunque disminuye, se mantiene relativamente alto a pesar de la acumulación de ácidos grasos volátiles (Figura 4.5). Este hecho se ha constatado en los experimentos en continuo realizados, en los cuales, a pesar de fuertes acumulaciones de AGV, incluso por encima de las obtenidas en la 118 Simulación dinámica del pH simulación con el modelo, el pH se mantiene siempre alto. 0.5 Purin 2 0.45 Conc. AGV y NH4+ (g/L) 8 0.4 7 pH 0.35 6 NH4+ 5 0.3 0.25 4 CH4 Acético 0.2 3 0.15 2 0.1 1 0.05 Propiónico 0 0 20 40 60 80 Tiempo (días) 100 120 Prod. CH4 (L/g DQO) 9 0 140 Figura 4.5. Simulación del proceso con dos tipos de purín en el rango termofílico: el purín 2 con mayor concentración de amonio que el purín 1, utilizado hasta el día 30 El modelo tiene en cuenta la inhibición por ácidos grasos de cadena larga libres (AGCL). Esta inhibición puede provocar el fracaso del proceso en casos de gran acumulación de estos compuestos. La forma más habitual de encontrar las sustancias lipídicas en un substrato es en forma de triglicéridos, que han de ser hidrolizados a glicerol y AGCL. La inhibición por AGCL depende de que la concentración de microorganismos acetogénicos sea suficiente para irlos consumiendo conforme se van hidrolizando los triglicéridos (Angelidaki et al., 1992). Por ello es especialmente importante la introducción gradual de substratos con alto contenido en grasas. En la Figura 4.6, se observan los resultados de la simulación del proceso al introducir TDO (residuo con alto contenido en grasas), en dos o en un paso. Si la introducción se hace en dos pasos, introduciendo en el día 10 TDO 2,5% (Figura 4.6a) o TDO 1% (Figura 4.6b) y en el día 20 TDO 5%, no hay apenas acumulación de AGV, y no disminuye la producción de metano. Sin embargo, si se introduce directamente el día 10, en un sistema alimentado previamente con Purín 1, el substrato TDO 5%, se produce una situación completamente inhibida (Figura 4.6c). Optimización de la digestión anaerobia de purines de cerdo 0.5 TDO 2.5% 3.5 0.6 3 0.5 Biogas 2.5 0.4 TDO 1% 2.5 0.4 Acético 1.5 0.2 1 Propiónico 0.1 0.5 AGCL 0 0 10 20 30 Tiempo (días) Conc. AGV y AGCL (g/L) 0 5 4.5 4 3.5 3 2.5 2 1.5 1 0.5 0 Propiónico 0.2 1.5 1 AGCL 0.1 0.5 0 40 0 0 10 20 30 Tiempo (días) 3.5 TDO 5% 2 Acético 0.3 b 3 Biogas 2 0.3 3.5 TDO 5% 40 c 3 AGCL 2.5 2 1.5 Acético Biogas 1 Propiónico 0.5 Prod. Biogas (L/L*d) Conc. AGV y AGCL (g/L) TDO 5% Prod. Biogas (L/L*d) a 0.6 119 0 0 10 20 Tiempo (días) 30 40 Figura 4.6. Evolución de AGV y producción de biogás, según la forma de introducción de TDO como cosubstrato: a: Introducción gradual 2,5%-5%; b: 1%-5%; c: Introducción única 5% La expresión de inhibición por AGCL se ha tomado directamente de Angelidaki et al. (1996), que trabajan en rango termofílico, considerando sólo un único valor de la constante de inhibición para todos los grupos de microorganismos y no dependiente de la temperatura. El valor de dicha constante es relativamente bajo, lo que provoca el fracaso total del sistema en el rango mesofílico, con un 5% de TDO. Esto no se ha constatado en los experimentos realizados (capítulo 5 y 6), aunque sí ha sucedido en el rango termofílico. La inhibición por AGCL depende de la temperatura, siendo más sensibles los sistemas termofílicos que los mesofílicos a la inhibición por AGCL (Hwu et al., 1997), diferencia que no se ha introducido en el modelo, y que es la causa de la discrepancia observada entre los datos experimentales y los simulados. 120 Simulación dinámica del pH 4.9. Conclusiones El algoritmo de simulación dinámica del pH desarrollado es capaz de predecir, con coeficientes de regresión elevados, el comportamiento del pH en soluciones acuosas sencillas, por lo que es una herramienta válida, útil para ser introducida en cualquier modelo de simulación de procesos dinámicos en los que el pH tenga un papel importante. El algoritmo de simulación del pH ha sido introducido en un modelo dinámico estructurado de digestión anaerobia de substratos complejos. Se ha utilizado notación matricial lo cual posibilita la introducción de nuevas variables al modelo, en caso de que se considere oportuno. La introducción de la simulación dinámica de las variables correspondientes a las componentes de la fase gaseosa supone una mejora importante del modelo original. Se ha comprobado la utilidad del modelo como herramienta cualitativa para prever situaciones de estrés y sobrecargas provocadas por la introducción de un cosubstrato con alto contenido en materia orgánica, o por la introducción de algún tóxico. Estos resultados son de interés para guiar los ensayos experimentales de los capítulos posteriores e interpretar resultados. 5. CODIGESTIÓN DE PURÍN CON RESIDUOS INDUSTRIALES. ENSAYOS DE VIABILIDAD EN DISCONTINUO. EFECTO DE LAS MEZCLAS. EFECTO DEL AMONIO. 122 Ensayos de viabilidad de codigestión en discontinuo Optimización de la digestión anaerobia de purines de cerdo 123 5.1. Resumen La digestión anaerobia de purines de cerdo puede ser una buena opción para la revalorización económica de estos residuos. Aunque la digestión anaerobia termofílica es más eficiente que la mesofílica, presenta algunas limitaciones, como menos estabilidad y un mayor efecto de algunos inhibidores. El principal inhibidor de la digestión anaerobia de purines de cerdo es el amoníaco libre. Con el objetivo de mejorar la producción de metano, se realizaron ensayos de viabilidad en discontinuo de mezclas de purín con residuos de la industria agroalimentaria, como residuos de la elaboración de zumos de frutas (pulpa de pera) y del refinado de aceite de oliva (tierras decolorantes). Se llevaron a cabo una serie de experimentos en discontinuo, con el objetivo de determinar el máximo potencial de metano y la biodegradabilidad de estos residuos. El experimento fue desarrollado en los rangos termofílico y mesofílico (35º y 55º). Los resultados en mesofílico fueron mejores que en termofílico, ya que a 55ºC mostraron una importante inhibición por amonio. En ambos rangos de temperatura la producción de metano mejoró por la adición de residuo de pera como cosubstrato. La máxima producción de metano se obtuvo con la codigestión de purín con tierras decolorantes de aceite de oliva, en el rango mesofílico (proporción 95% y 5% de purín y tierras, respectivamente). La producción de metano fue 340 mL CH4/g SVini, que es 2,3 veces la producción de metano respecto a sólidos volátiles para el purín sólo (150 mL CH4/g SVinitial). Sin embargo, hubo signos de ligera inhibición, aumentando al aumentar la proporción de cosubstratos. En el rango termofílico, en contraste, la adición de tierras decolorantes de aceite de oliva resultó en producciones de metano más bajas que el control. 5.2. Introducción 5.2.1. Codigestión de residuos orgánicos. El término codigestión se emplea para expresar la digestión anaerobia conjunta de dos o más substratos de diferente origen. La principal ventaja radica en el aprovechamiento de la sinergia de las mezclas, compensando las carencias de cada uno de los substratos por separado. 124 Ensayos de viabilidad de codigestión en discontinuo La digestión anaerobia de mezclas de diferentes tipos de residuos ha dado buenos resultados a escala laboratorio y en algunos casos a escala industrial, tal y como se recoge en la literatura internacional. Se han encontrado buenos resultados para mezclas de varios tipos de residuos de industrias cárnicas y mataderos, ricos en grasas, consiguiendo altas producciones de metano, del orden de 47 m3/m3 de residuo introducido (Brinkman, 1999). También ha dado buenos resultados la codigestión de lodos de depuradora y la fracción orgánica de residuos municipales, FORM (Di Palma et al., 1999; Hamzawi et al.,1998) y la mezcla de residuos sólidos urbanos, principalmente a base de restos de vegetales, y aguas residuales urbanas (Edelmann et al., 1999), así como de lodos de depuradora y residuos de frutas y vegetales (Dinsdale et al., 2000). La codigestión de residuos ganaderos y residuos orgánicos en sistemas de mezcla completa es una metodología exitosa tanto en régimen termofílico como en el mesofílico (Brinkman, J., 1999). En Dinamarca funcionan alrededor de 35 plantas centralizadas de producción de biogás, que se empezaron a implantar en los años ochenta, lo que ha posibilitado el tratamiento combinado de residuos ganaderos y residuos orgánicos procedentes de la industria alimentaria, de plantas depuradoras de aguas residuales urbanos, residuos de mataderos y la fracción orgánica de residuos sólidos urbanos (Angelidaki y Ahring, 1997a). Los residuos urbanos e industriales suelen contener altas concentraciones de materia orgánica fácilmente degradable, por lo que presentan un mayor potencial de producción de biogás que los residuos ganaderos, de 30 a 500 m3/ton (Ahring et al., 1992; Bardiya et al., 1996; Angelidaki y Ahring, 1997a). Sin embargo, estos residuos pueden presentar problemas para su digestión, como deficiencia en nutrientes necesarios para el desarrollo de los microorganismos anaerobios, baja alcalinidad, o excesivo contenido en sólidos que provoque problemas mecánicos (Banks y Humphreys,1998). Los residuos ganaderos, y en concreto el purín de cerdo, pueden ser una buena base para la codigestión porque, generalmente, presentan un contenido de agua más alto que la mayoría de residuos industriales, una mayor capacidad tampón y aportan una amplia variedad de nutrientes necesarios para el crecimiento de los microorganismos anaerobios (Angelidaki y Ahring, 1997a). Los efectos beneficiosos de la introducción de mezclas de residuos ganaderos con residuos industriales se han puesto de manifiesto en plantas a escala real en Dinamarca. La producción media de las plantas que utilizan mezclas fue en el mes de septiembre del año 1999 de 38.5 m3 de gas/m3 de Optimización de la digestión anaerobia de purines de cerdo 125 biomasa, con un valor máximo para la planta de Vegger, que trata mezclas de residuos ganaderos e industriales, de 90 m3 gas/m3 de biomasa introducida en el reactor, mientras que la producción de las plantas que trabajan sólo con residuos ganaderos fue siempre menor de 26,6 m3 de gas/m3 de biomasa, con un valor medio de 14,5 m3 de gas/m3 de biomasa (Danish Energy Agency, 1999). Diferentes tipos de residuos, principalmente de la industria agroalimentaria, se han testado ya como posibles cosubstratos para la digestión de residuos ganaderos. A continuación se exponen algunos ejemplos. Ahring et al. (1992) estudiaron la viabilidad de la codigestión de estiércol con residuos de la elaboración de piensos. Debido al alto contenido en nitrógeno de este producto, inicialmente se produce la inmediata inhibición del proceso de digestión, aunque finalmente los microorganismos son capaces de aclimatarse, disminuyendo la concentración de ácidos grasos volátiles y produciendo una alta y constante producción de biogás. Diversos trabajos se han desarrollo teniendo como base la codigestión de estiércol bovino con tierras residuales procedentes del proceso de refinado de aceite, BBO (bentonite bound oil). En general, la adición de este residuo a plantas a escala real produce una mayor estabilidad del proceso, produciendo un aumento en la producción de gas, debido a la conversión en metano de la mayoría del carbono añadido, aunque no se observa una mejora en la tasa de conversión del estiércol en sí mismo. La adición de BBO produce una mayor producción de metano respecto a los sólidos volátiles añadidos, debido al mayor potencial de producción de biogás de la grasa contenida en la BBO que del estiércol, de 0,2 a 0,23 L de CH4/g SV (Ahring et al., 1992). Una hipótesis planteada para explicar la mejora en la producción de metano es la disminución de problemas de inhibición por amonio, debido a la capacidad de adsorción superficial de este material. Sin embargo, la adición de bentonita sola (sin grasa) no tuvo ningún efecto sobre la producción de gas, aunque sí contribuyó a una más rápida recuperación después de la introducción de amonio a niveles inhibidores (Angelidaki y Ahring, 1993b), así mismo la adición de bentonita reduce la inhibición por lípidos (Angelidaki et al., 1990). La codigestión de alpechín, residuo acuoso de la producción de aceite de oliva virgen, y estiércol bovino hace posible el tratamiento del primero mediante digestión anaerobia. La fermentación de alpechín solo presenta problemas, debido a la alta concentración de compuestos tóxicos (polifenoles), o baja concentración de nutrientes esenciales (N) y baja 126 Ensayos de viabilidad de codigestión en discontinuo alcalinidad. La codigestión aumenta, además, el índice de producción de biogás del estiércol (Angelidaki et al., 1997). La mezcla de alpechín y purín de cerdo ha mostrado buenos resultados en otros estudios, alcanzando niveles de degradación de DQO del orden del 65% (Schmidt et al., 1999). La mezcla de purín con lodos de depuradora, tanto en el rango termofílico como mesofílico, ha proporcionado resultados positivos (Wong, 1990; Flotats et al.,1999). Se han obtenido también resultados positivos al mezclar estiércol de bovino y residuos lignocelulósicos, hojas machacadas, paja de trigo, restos vegetales, pretratados con hidróxido sódico (Dar y Tandon, 1987), así como la mezcla de purín con paja (Masciandaro et al., 1994). Con residuos de tomate mejora la digestión del estiércol (Trujillo et al., 1993), así como la mezcla de residuos bovinos y residuos de frutas y verduras (Callaghan et al., 1999). Otros muchos residuos se exponen en la bibliografía como responsables de mejora de las producciones de biogás de los residuos ganaderos, tales como residuos de lechería (Gavala et al., 1996; Desai y Madamwar, 1994; Desai et al., 1994), y residuos de pescados y lodos de la industria cervecera (Callaghan et al., 1999). En general, la mezcla de residuos ganaderos de diferentes tipos de ganado puede mejorar la producción de metano debido, principalmente, al mayor aporte de sólidos orgánicos (Callaghan et al., 1999), o a la dilución de algún efecto inhibitorio, como la concentración de amonio (Hansen et al., 1998). A pesar de los buenos resultados recogidos en la bibliografía, al mezclar diferentes tipos de residuos se corre el riesgo de introducción de sustancias tóxicas o inhibidoras para el proceso anaerobio, siendo preciso determinar la viabilidad de la mezcla, así como la proporción adecuada de cada substrato, y la optimización de otros parámetros del proceso, como la temperatura, velocidad de carga, etc. Por otro lado, la introducción de substratos altamente degradables, característica apreciada para mejorar la producción de gas, puede provocar problemas de sobrecargas orgánicas en el reactor, y liberar compuestos inhibidores del crecimiento de los microorganismos. Por ejemplo, el alto contenido en lípidos de algunos residuos industriales proporciona altos potenciales teóricos de producción de biogás, pero, en función de la concentración y composición de ácidos grasos, pueden resultar altamente tóxicos para el crecimiento microbiano (Galbraith et al., 1971; Hanaki et al., 1981; Koster y Cramer, 1987; Angelidaki y Ahring, 1992; Hwu et al., 1997). Es, por tanto, necesario realizar estudios de viabilidad de las mezclas, determinando la presencia de tóxicos o inhibidores que puedan Optimización de la digestión anaerobia de purines de cerdo 127 invalidar el nuevo residuo como cosubstrato. 5.2.2. Substratos posibles en el área de Lleida Tal y como se expuso en el capítulo 1, la concentración de explotaciones porcinas en las comarcas de Lleida, en especial, en las comarcas de El Segrià, Pla d’Urgell, Urgell, La Noguera y Les Garrigues, hace que los residuos generados en dichas comarcas constituyan el principal problema ambiental de la zona. El principal limitante para su aplicación como fertilizante orgánico es el elevado contenido en nitrógeno. Este también es el principal limitante para la digestión anaerobia, sobre todo en el rango termofílico, tal y como se ha expuesto en el capítulo 2. Lleida se caracteriza por ser una zona eminentemente agrícola, donde la industria agroalimentaria tiene un elevado peso sobre la economía global. El Segrià y el Pla d’Urgell son dos de las principales zonas frutícolas de España, produciendo especialmente pera, manzana y, en menores cantidades, melocotón y otras frutas. Los excedentes de producción han ocasionado algunos años problemas importantes de manejo, siendo necesario su vertido controlado. Por otra parte, la industria de procesado de frutas genera una importante cantidad de residuos sólidos, que aunque son considerados como subproductos, podrían llegar a suponer un problema de gestión. Sin embargo, podrían tener un aprovechamiento para la mejora de la digestión anaerobia de residuos ganaderos. El residuo de fruta es el residuo sólido resultante de la extracción del zumo, y está compuesto fundamentalmente por la pulpa, presentando un alto contenido en compuestos lignocelulósicos. Las frutas son muy ricas en carbohidratos y en azúcares, substratos fácilmente degradables por los microorganismos acidogénicos, de rápido crecimiento y poca sensibilidad a tóxicos e inhibidores, por lo que se generan muy rápidamente ácidos que hacen bajar el pH a un nivel al que los microorganismos anaerobios no pueden sobrevivir, sobre todo los organismos metanogénicos. La digestión anaerobia de residuos de frutas, sobre todo en el caso de épocas de excedentes, puede ser una interesante opción para la estabilización de los mismos. Sin embargo, el proceso presenta graves problemas, sobre todo debido a la baja alcalinidad y posible deficiencia de determinados nutrientes, a pesar de presentar un alto potencial de producción de biogás debido al alto contenido en materia orgánica. Por otro lado, España es el principal productor mundial de aceite de oliva, centrándose su mayor producción en Andalucía, pero con importantes producciones en Cataluña, especialmente en la comarca de Les Garrigues. 128 Ensayos de viabilidad de codigestión en discontinuo El proceso de producción de aceite de oliva genera unos residuos acuosos, conocidos como alpechín, que suponen un grave problema ambiental, por su elevada carga orgánica. El aceite de oliva se puede comercializar directamente, como aceite virgen si alcanza un determinado nivel de calidad en sus propiedades físicas, químicas y organolépticas. El aceite que no alcanza este nivel, se conoce como aceite lampante, y debe ser refinado, sometido a una serie de procesos físico-químicos, para poder comercializarlo. Lo que comercialmente se conoce como “aceite de oliva” es una mezcla de aceite de oliva refinado y de aceite de oliva virgen. El proceso de refinado de aceites comestibles consiste en una serie de procesos, entre los que están la neutralización, la winterización y la decoloración. La decoloración consiste en la adsorción sobre la superficie de un material inerte, de elevada relación superficie/volumen, de los compuestos que dan color al aceite crudo, básicamente pigmentos y clorofilas. El procedimiento consiste en mezclar el volumen de aceite en unos tambores con tierras decolorantes, material inerte con una gran superficie adsorbente y, posteriormente, realizar la separación por vacío, actuando las propias tierras como filtro, formando una capa espesa junto a la membrana que retiene las tierras y deja pasar el aceite a través de pequeños canales. Las tierras se van saturando con las sustancias adsorbidas, llegando un punto en que es necesario reemplazarlas. Las tierras una vez utilizadas presentan un alto contenido en sustancias orgánicas, con una importante fracción de grasas, que puede llegar a superar el 30% en peso. Las tierras decolorantes residuales presentan graves problemas de manejo, siendo un residuo con un elevado contenido en materia orgánica, con tendencia a la autocombustión espontánea, convirtiéndose en un residuo no sólo contaminante, sino también peligroso, que debe ser vertido en vertedero de residuos especiales, con el consiguiente coste. El alto contenido en lípidos del substrato proporciona altos potenciales teóricos de producción de biogás, por lo que se plantea el estudio de viabilidad de la digestión de la mezcla de purín con tierras decolorantes. La codigestión con tierras decolorantes de aceites comestibles ha proporcionado muy buenos resultados tanto en experimentos de laboratorio como en plantas a nivel industrial, haciendo económicamente viables plantas de gestión centralizada en Dinamarca (Ahring et al., 1992; Angelidaki y Ahring, 1997a). En función del tipo de aceite, la composición de ácidos grasos es distinta, por lo que puede tener importancia la procedencia de las tierras. El aceite de oliva constituye una importante fracción de los aceites que se procesan en las refinerías españolas. El aceite de oliva tiene entre un 65 y un Optimización de la digestión anaerobia de purines de cerdo 129 80% de riqueza de ácido oleico, característica que le hace tener buenas propiedades dietéticas. Sin embargo el ácido oleico es un importante inhibidor de los microorganismos anaerobios (Galbraith et al., 1971; Hanaki et al., 1981; Koster y Cramer., 1987; Angelidaki y Ahring, 1992; Hwu et al., 1997). 5.2.3. Metodología a utilizar para realizar estudios de viabilidad La estandarización del método de estudio, definiendo un test previo sencillo, rápido y ajustado a la realidad será fundamental para el desarrollo futuro de la tecnología de codigestión. Tradicionalmente los ensayos de digestión anaerobia han seguido siempre un esquema que pasa en primer lugar por test sencillos de biodegradabilidad y/o toxicidad en discontinuo, posteriormente, ensayos de biodegradabilidad en continuo y/o ensayos en continuo a escala laboratorio, y finalmente ensayos en planta piloto (Lema, 1995). Los ensayos en discontinuo, que se plantean en la bibliografía, son ensayos de biodegradabilidad, para medir el potencial de degradación anaerobia del substrato, o ensayos de toxicidad, que consiste en mantener la concentración de tóxico en el medio, y son útiles para conocer el potencial de producción de metano o la toxicidad de un determinado componente del substrato (Field et al., 1988), pero no proporcionan información acerca del comportamiento en condiciones reales del substrato, con la posible presencia de tóxicos no conocidos, o con una concentración relativamente baja de microorganismos. Para estudiar la viabilidad de las mezclas de residuos, diversos autores han utilizado otro tipo de experimentos en discontinuo, similares a los ensayos de biodegradabilidad y toxicidad empleados en la bibliografía, pero con menor concentración inicial de inóculo, de forma que no se enmascaran los posibles efectos inhibidores. Así se consigue, con un único ensayo, una primera aproximación a la viabilidad del proceso, y realizando determinaciones analíticas de ácidos grasos volátiles a lo largo del experimento, se puede llegar a establecer la posible causa de la existencia del problema, observado mediante la producción de gas, pudiendo determinar cual es la etapa limitante del proceso. El tipo de ensayo de viabilidad que se propone realizar en el presente estudio, ha sido utilizado previamente, habiendo obtenido resultados satisfactorios para estudios de mezclas (Callaghan et al., 1999; Flotats et al., 1999), permitiendo definir, además, el rango de proporción de mezcla que mayores producciones de metano obtiene. Para validar los resultados obtenidos en discontinuo, así como para definir 130 Ensayos de viabilidad de codigestión en discontinuo una serie de parámetros adicionales de gran importancia, como la velocidad de carga orgánica, velocidad en que se debe introducir el cambio de substrato en el reactor, es necesario realizar ensayos en continuo a escala piloto laboratorio, metodología que se aborda en el capítulo siguiente. Antes del paso a escala industrial, será conveniente realizar una validación de los resultados obtenidos a escala piloto industrial, que presenta la ventaja de poder trabajar con los mismos residuos que la planta industrial, con los mismos problemas de variabilidad, de falta de homogeneización, etc. La planta piloto industrial permite, además, ensayar diferentes parámetros operacionales que pueden tener gran importancia en el reactor real, como parámetros de diseño y forma del reactor, eficiencia del sistema de agitación, necesidades energéticas, etc. (Lema et al., 1995). 5.3. Objetivos particulares discontinuo. de los ensayos en El objetivo general de esta serie de experimentos fue el estudio del proceso de digestión anaerobia de purines de cerdo y de la codigestión de éste con residuos orgánicos de la industria agroalimentaria en discontinuo. Para la consecución de este objetivo general se plantearon los siguientes objetivos particulares: 1. Estudiar el efecto de la temperatura sobre la digestión anaerobia de purines de cerdo: Comparación de rangos mesofílico y termofílico. 2. Estudiar la viabilidad de la codigestión de purines de cerdo con residuos de la industria agroalimentaria. Efecto del tipo de cosubstrato. Optimización de la proporción de mezcla. Estudio de posibles efectos de inhibición y sobrecargas. 3. Estudiar la interacción entre los parámetros temperatura, concentración de amonio y adición de un cosubstrato. 4. Estudiar el efecto del amonio sobre la digestión anaerobia de purines de cerdo. Efecto de la dilución. Interacción de la temperatura, la concentración de amonio y el pH. 5. Evaluar la metodología de ensayos en discontinuo para estudiar la viabilidad de la digestión anaerobia de substratos complejos. Optimización de la digestión anaerobia de purines de cerdo 131 5.4. Materiales y Métodos 5.4.1. Diseño del experimento El diseño de los experimentos fue el que se muestra en la Tabla 5.1. Tabla 5.1. Diseño de los experimentos. (5 repeticiones por cada tratamiento) Experimento Trat. Tª % en peso de cada componente % Purín % Rpera % TDO % H2O Experimento 1: Diluciones de purín. Efecto del amonio Experimento 11.1 35ºC 100 0 35ºC 1.2 35ºC 95 5 1.3 35ºC 87.5 12.5 1.4 35ºC 80 20 Experimento 11.1 55ºC 100 0 55ºC 1.2 55ºC 95 5 1.3 55ºC 87.5 12.5 1.4 55ºC 80 20 Experimento 2: Codigestión de purín con residuo de pera Experimento 22.1 35ºC 100 35ºC 2.2 35ºC 95 5 2.3 35ºC 87.5 12.5 2.4 35ºC 80 20 2.5 35ºC 20 80 Experimento 22.1 55ºC 100 2.2 55ºC 95 5 55ºC 2.3 55ºC 87.5 12.5 2.4 55ºC 80 20 2.5 55ºC 20 80 Experimento 3: Codigestión de purín con tierras decolorantes de aceite de oliva Experimento 33.1 35ºC 100 35ºC 3.2 35ºC 95 5 3.3 35ºC 87.5 12.5 3.4 35ºC 12.5 87.5 Experimento 33.1 55ºC 100 55ºC 3.2 55ºC 95 5 3.3 55ºC 87.5 12.5 3.4 55ºC 12.5 87.5 Blancos: Efecto del inóculo Blanco-35ºC 4.1 35ºC 100% Blanco -55ºC 4.1 55ºC 100% 132 Ensayos de viabilidad de codigestión en discontinuo Se desarrollaron tres experimentos simultáneamente a dos temperaturas (35º y 55º). Para cada temperatura se realizó un blanco consistente en la digestión del inóculo con agua para estimar la producción de gas a partir de la materia orgánica contenida en el inóculo. Los tratamientos control de los experimentos 2 y 3, fue el purín 100% del experimento 1. 5.4.2. Métodos analíticos Se realizaron según los métodos expuestos en el capítulo 3. 5.4.3. Desarrollo y seguimiento de los ensayos discontinuos Se utilizaron viales de vidrio de 118 mL de capacidad, con 50 gramos de substrato y 5 gramos de inóculo. Se realizaron cinco repeticiones de cada tratamiento. Los viales llenos se burbujearon con un gas inerte, mezcla de N2 y CO2 en proporción 80%-20% (Figura 5.1) durante dos minutos y se cerraron inmediatamente con un septum y una cápsula metálica. Se utilizaron dos incubadores (Figura 5.2), uno para cada temperatura, equipados con termostato de seguridad. Por no disponer de un sistema en continuo en el momento de comenzar el ensayo, el inóculo se preparó digiriendo en discontinuo una mezcla de purín y residuo de manzana, inoculado a su vez con efluente de un reactor a escala industrial, tipo flujo pistón, que trata purines de cerdo. Este digestor está funcionando ininterrumpidamente desde hace 18 años en la granja Mas El Cross, de ciclo cerrado, situada en Olot (Girona). La mezcla se digirió a las dos temperaturas (35ºC y 55ºC) en discontinuo hasta encontrar que la proporción de CH4/CO2 era aproximadamente un 60%/40%, (30 días), conservándose en nevera hasta el momento de ser utilizada. Figura 5.1. Burbujeo de viales antes de comenzar la digestión Optimización de la digestión anaerobia de purines de cerdo 133 Figura 5.2. Detalle del interior de uno de los incubadores Periódicamente se analizó la composición del espacio de cabeza y se tomaron muestras de la fracción líquida con jeringa hipodérmica, para el posterior análisis de ácidos grasos volátiles (Figura 5.3). Tanto al inicio de la digestión como al final de la misma se realizó una analítica completa de las muestras incluyendo: Sólidos Totales (ST) y Volátiles Figura 5.3. Elementos (SV), Demanda Química de Oxígeno (DQO), Nitrógeno Kjeldahl (Nk), Nitrógeno amoniacal (Nde muestreo. NH4+), pH y alcalinidad. Para caracterizar el substrato inicial se analizaron también el contenido en grasas y la fibra ácido detergente, fibra neutro detergente y lignina, el contenido en cationes y aniones de la fracción soluble. 5.4.4. Cálculo de la producción de gas acumulada. La producción de gas acumulada se calculó a partir de la composición y presión del mismo. Las muestras de gas se tomaron con una jeringa con válvula de cierre, de forma que se pudieron tomar las muestras a la presión real, estimando el número de moles reales contenidos en el volumen de muestra. Se conocía la composición inicial y por tanto el número de moles contenido en el espacio de cabeza de nitrógeno y CO2. Conforme se fue produciendo el proceso de la digestión aumentó la cantidad de CO2 y CH4 produciéndose un aumento de presión. Se asumió que se cumplía la ley de los gases perfectos y se pudo calcular la presión a partir del número de 134 Ensayos de viabilidad de codigestión en discontinuo moles, analizado mediante cromatógrafo de gases. Periódicamente se procedió al vaciado de gas de estos viales, analizándose antes y después la composición del gas de cabeza para poder calcular el gas perdido. Se observó un fenómeno no carente de interés y es que en las horas posteriores al vaciado se producía una mayor producción de gas. La hipótesis planteada es que el aumento de la presión del espacio de cabeza hace que el equilibrio líquido-gas se desplace hacia la fase líquida pudiendo, incluso, provocar la inhibición de la acetogénesis por CO2 (Hansson y Molin, 1981), de forma que al vaciar, baja la concentración de CO2 y se reactiva el proceso de metanogénesis. La producción de metano se expresó en unidades de mL de CH4 a 20ºC y presión 1 atmósfera. Se puede hacer fácilmente la transformación a volumen estándar (0ºC y 1 atmósfera), multiplicando el valor obtenido por el factor 0.932. La producción acumulada neta de gas se estimó sustrayendo la producción acumulada de metano obtenida para el blanco (digestión del inóculo con agua). 5.4.5. Cálculo de índices de producción de metano La producción de metano por si sola no resulta un buen parámetro de comparación, ya que dependerá de la forma en que se ha realizado el experimento, del tipo de substrato empleado, etc. Por ello se utilizarán varios índices que permitirán la comparación y el estudio de la eficiencia del proceso. Volumen de gas acumulado (M). Este es el parámetro más utilizado, a pesar de no proporcionar información por sí sólo, pero tiene una gran importancia como medida del potencial de producción. Dado que todos los tratamientos se realizaron utilizando una misma metodología, es un índice de valor a nivel comparativo dentro de este conjunto de experimentos. Para compararlo con resultados de otros autores u otros estudios se deberá hacer siempre referencia a la metodología del ensayo y sobre todo al volumen del reactor y cantidad de substrato e inóculo empleado. El cálculo de este índice se realizó substrayendo la media del volumen acumulado de biogás/metano del blanco (Vblanco), para cada temperatura, al volumen acumulado (V) de cada una de las repeticiones de cada tratamiento. El cálculo de este índice se efectuó cada día en que se midió el gas, obteniéndose la evolución de la producción de gas acumulada (biogás, metano y dióxido de carbono). Optimización de la digestión anaerobia de purines de cerdo Vt VBlanco ,t M 135 (5.1) Volumen de gas respecto a la masa inicial de substrato (M’). Este parámetro estima la producción de gas (metano o biogas) por unidad de masa de substrato, y es necesario para comparar los resultados con los obtenidos por otros autores. M' M Masa de substrato (5.2) Volumen de gas respecto a la cantidad de materia orgánica inicial (B). Este es un parámetro mucho más universal ya que se elimina el factor tamaño del reactor o cantidad de substrato añadido, pero sobre todo es interesante para comparar la biodegradabilidad de cada substrato, independientemente del contenido de materia orgánica. Consiste en calcular el volumen de biogás o metano producido por cada gramo de materia orgánica añadida (medida como sólidos volátiles o DQO). Este índice se calcula como el volumen neto de producción de biogás o metano (M) dividido por la cantidad total de sólidos volátiles añadidos (resultado de multiplicar la concentración por la masa total de substrato en el reactor). Se calculó tanto par el biogás acumulado como para el metano y el dióxido de carbono. Bt M Masa x >C @ M' >C @ , (5.3) donde C es la concentración de SV o DQO inicial. Producción de metano potencial, velocidad específica de producción de metano y duración del retardo inicial en el inicio de la producción de gas. La estimación de los parámetros producción potencial y velocidad específica de producción de metano tiene gran interés para comparar diferentes tratamientos y para relacionar la evolución de estas variables con otros parámetros del proceso, por ejemplo para comparar la velocidad específica de producción de metano con la concentración de amonio o amoníaco libre (Hansen et al., 1998; Angelidaki y Ahring, 1993b). Con el objetivo de comparar las curvas de producción de gas para los diferentes tratamientos se buscó un modelo simple capaz de simular la producción de gas en discontinuo en la mayoría de los casos y poder realizar comparaciones. Varios modelos han sido empleados en la bibliografía con este objetivo, la mayoría basados en cinéticas de primer orden (Hashimoto, 136 Ensayos de viabilidad de codigestión en discontinuo 1989; Chen y Hashimoto, 1996; Hansen et al., 1998), siguiendo la siguiente expresión: dM k ·M 0 M , (5.4) dt donde M0 es la producción potencial y K es la constante de velocidad. Al integrar, la curva de producción de metano acumulada viene dada por: M M 0 * 1 e k * t , (5.5) La misma ecuación podría aplicarse al índice de producción acumulada respecto al volumen de substrato inicial o respecto al contenido de sólidos volátiles iniciales (B y B0). La utilización de este tipo de expresiones puede ajustarse suficientemente bien a los datos experimentales de ensayos en discontinuos de substratos complejos, si no existe desfase inicial en la producción de gas (fase lag), pero en el caso contrario podría dar valores erróneos en la estimación de la velocidad de producción de metano (Veeken y Hamelers, 1999). En el presente trabajo, fue necesario emplear otro tipo de expresiones que incluyen como un parámetro más la duración del desfase inicial, eligiéndose la ecuación de Gompertz, K* e O t 1 ) M0 ( e ), M0 * e ( (5.6) con M producción acumulada de metano, en mL CH4; M0, producción potencial acumulada de metano, en mL CH4; K, velocidad de producción de metano, en mL CH4/día; O duración de la fase lag, en días; t es tiempo, en M días; y e =exp(1). Lay et al. (1997) utilizaron esta expresión para estimar la producción potencial y la velocidad de producción de metano en ensayos discontinuos con substratos de alto contenido en sólidos, encontrando que la ecuación de Gompertz era la que mejor simulaba el comportamiento de estos sistemas, alcanzando unos coeficientes de regresión muy altos. La misma expresión se ajustó a los datos del índice de producción acumulada respecto a sólidos volátiles (B) para anular el efecto de contenido inicial de materia orgánica, cambiando las unidades: B y B0 en ml de CH4/g SV, R en ml de CH4/g SV*día. R* e O t 1 ) B0 ( e ) B0 * e ( B (5.8) Optimización de la digestión anaerobia de purines de cerdo 137 Para eliminar el efecto de la concentración inicial de substrato sobre la velocidad de producción de metano, se estimó un nuevo parámetro, que denominaremos velocidad específica de producción de metano (R’), en unidades de T-1, calculado de la siguiente forma: R' R* e B0 (5.9) Se determinaron los parámetros para cada una de las repeticiones de cada tratamiento, y posteriormente se realizó un análisis de varianza de los coeficientes obtenidos. El análisis de regresión se realizó mediante el paquete estadístico Stat-Graphics, realizando el ajuste por mínimos cuadrados con un nivel de confianza del 95%. El análisis se varianza se realizó siguiendo el mismo método que el resto de parámetros analizados. Indice de acidificación, metanogénesis y biodegradabilidad. El índice de biodegradabilidad es un índice que permite determinar la fracción de DQO de un determinado residuo que puede ser degradada por vía anaerobia (Field et al., 1988). La biodegradabilidad se determina mediante un test de biodegradabilidad, que requiere unas determinadas concentraciones de DQO, de cantidad mínima de lodo y de ausencia de tóxicos, que no se cumplieron en el presente estudio. Sin embargo los parámetros que determinan la biodegradabilidad pueden ser calculados, teniendo especial interés para la comparación de los diferentes substratos y proporciones de mezclas entre ellos. Los índices de acidificación y metanización pueden ser útiles para comparar la eficiencia de dichos procesos, y para estudiar la influencia de otros factores, como la presencia de tóxicos. El cálculo se basa en la utilización de unidades de DQO para la expresión de todos los parámetros. Los índices calculados son (Field et al., 1988): Índice de metanización (% M): %M donde DQOCH 4 100· DQOCH 4 , DQOinf mLCH 4 ( 20º C ) 376 ; (5.10) 138 Ensayos de viabilidad de codigestión en discontinuo Índice de acidificación (%A): % A 100· DQOCH 4 DQO AGV ,efl ; (5.11) DQOinf Biodegradabilidad (%BD): % BD % A %Cel % BD %A § DQOAGV ,inf YA * ¨ % A 100 1 YA ¨© DQOinf · ¸ YM * % M , (5.12) ¸ 1 YM ¹ donde YA=0.196 g DQOcel/g DQOcons. y YM=0.028 g DQOcel/g DQOcons. Los parámetros DQOAGV, ef y DQOAGV,inf, se calculan a partir de la composición en AGV, a partir de las relaciones dadas en la Tabla 5.2. Los valores de DQO son los correspondientes a la materia total en el vial. Los valores de concentración medidos se han de multiplicar por la masa total de substrato (50 g). Tabla 5.2. Factores para la conversión de la concentración de AGV a unidades de DQO Tipo AGV mg DQO/mMol mg DQO/mg C2 (acético) 64 1.07 C3 (propiónico) 112 1.51 C4 (iso-butírico y n-butírico) 160 1.82 C5 (iso-valérico y n-valérico) 208 2.04 5.4.6. Caracterización de los substratos utilizados El purín utilizado procedió de una granja de ciclo cerrado situada en el término municipal de Palau d’Anglesola, comarca de Pla d’Urgell, provincia de Lleida. Se trata de una granja que cuenta con una capacidad para 2000 animales, con 1500 plazas de engorde, divididas en dos naves de 750 animales. La alimentación de los animales es a base de un pienso fabricado en una instalación aneja a la granja, que contaba con una alto contenido en fibra y en proteína, lo que influye grandemente en las características del purín. El purín utilizado fue recogido de una fosa bajo una nave de engorde. Se trata de un purín con alto contenido en sólidos, rico en nitrógeno y con alto contenido en fibra. El residuo de pera procedió de la industria dedicada a la extracción de Optimización de la digestión anaerobia de purines de cerdo 139 zumos de fruta, situada en el término municipal de Alguaire, comarca de Segriá, provincia de Lleida. Se trata de un residuo de aspecto sólido con un alto contenido en humedad (alrededor del 65%) y un alto contenido en sólidos volátiles, del orden de un 98% de los sólidos totales. El residuo de pera fue triturado antes de mezclarlo con el purín. Las tierras decolorantes procedían de una industria dedicada al refinado y envasado de aceites comestibles, y son un residuo del proceso de decoloración de aceite de oliva. El contenido de humedad es muy bajo, del orden de un 3%, y la materia orgánica (sólidos volátiles) es el 43% de ST. La concentración de los compuestos nitrogenados fue mucho más alta en el purín que en los otros dos residuos, especialmente la concentración de nitrógeno amoniacal. Sin embargo la concentración de materia orgánica, medida tanto en unidades de DQO como de SV fue mucho más alta en los residuos industriales que en el purín, especialmente la DQO de las tierras decolorantes (TDO). Una de los principales consecuencias de añadir estos tipos de cosubstratos fue el aumento de la materia orgánica “disponible” para la degradación, y susceptible de ser convertida en metano. Sin embargo, el tipo de materia orgánica añadida fue muy diferente para cada uno de los dos cosubstratos, siendo en el residuo de pera básicamente carbohidratos, un 79% sobre la materia seca, y un 80,9% sobre SV, con una alta proporción de lignina, mientras que en TDO la fracción más importante fueron las grasas, que supusieron un 27,7% de la materia seca, y un 70% sobre SV (Tabla 5.3). El alto contenido en sólidos de los cosubstratos hizo que el porcentaje de la materia orgánica de las mezclas aumentara de forma importante, pudiendo asociar la mayor parte del incremento al aporte del cosubtrato. En el caso de las mezclas con residuo de pera, la fracción de sólidos volátiles procedentes del cosubstrato llegó al 60% del total para el tratamiento 2.4 (20% en peso de residuo de pera), y en el caso de las mezclas con TDO el tratamiento 3.3 (12,5% en peso de TDO) el aporte de SV del cosubstrato supuso el 46,64% del total. Puesto que el contenido en nitrógeno de los residuos industriales utilizados fue mucho menor que en el purín, la relación carbono/nitrógeno, medido en unidades de g SV/g N-Nk aumentó considerablemente al incrementarse la proporción de cosubstrato añadida, pasando de 11,1 para el purín a 23,16 en el caso del tratamiento 2.4 (20% de residuo de pera) y a 20,92 para el tratamiento 3.3 (12,5% de TDO). La relación carbono/nitrógeno amoniacal aumentó proporcionalmente más, debido a que la mayor parte del nitrógeno de los residuos industriales es nitrógeno 140 Ensayos de viabilidad de codigestión en discontinuo orgánico, pasando de 17 g SV/g N-NH4+ para el purín a 42,73 y 31,72 g SV/g N-NH4+ para el tratamiento 2.4 y 3.3, respectivamente. Tabla 5.3. Caracterización físico química de los substratos utilizados* Parámetro Purín Residuo de Pera TDO Nk (g N-Nk/kg) 5,26 3,30 0,81 Amonio (g N-N-NH4+/kg) 3,43 0,43 0,32 N.Orgánico (g N-Norg/kg) 1,83 2,87 0,49 Proteína (g/kg) 11,43 17,97 3,06 Proteína (% sbMS) 14,43 5,08 0,33 S.T. (g/Kg) 79,16 354,00 916,96 S.V. (g/Kg) 58,40 346,50 362,62 S.V. (% sb MS) 73,77 97,88 39,55 DQO (g/Kg) 80,92 453,0 1090,40 Celulosa (% sb MS) 16,90 37,60 n.a.** Hemicelulosa (% sb MS) 13,00 10,20 n.a. ** Lignina (% sb MS) 26,3 31,40 n.a** Total Fibra (% sb MS) 56,20 79,2 n.a. ** Grasa (% sb MS) 3,70 n.a. ** 27,70 pH 8,06 3,45 6,93 Alcalinidad Total (g CaCO3/L) 10,55 0 0,95 Alcalinidad Parcial (g CaCO3/L) 7,50 0 0,20 Alcalinidad Intermedia (g AcH/L) 3,66 0 0,90 Relación de Alcalinidades 0.29 0 0.79 *valores medios de tres repeticiones;** n.a.: no analizado. El pH del purín fue bastante alcalino, en torno a 8, mucho más alto que el pH de los cosubstratos. El pH del residuo de pera estuvo en 3,45, y al añadirlo al purín hizo que el pH de la mezcla descendiera de forma notable. Debido al bajo pH se puede considerar que la alcalinidad al bicarbonato es nula. Al mezclar con purín se observó el consecuente descenso de la alcalinidad de las mezclas. La relación de alcalinidades (RA) aumentó con el Optimización de la digestión anaerobia de purines de cerdo 141 porcentaje de cosubstrato añadido, indicando la menor estabilidad de las mezclas que del purín. El residuo TDO mostró un pH próximo a la neutralidad, aunque la alcalinidad debida al bicarbonato se mantuvo en niveles bastante bajos (Tabla 5.3). La relación de alcalinidad de este residuo mostró un valor muy alto (0,79), lo que hizo aumentar considerablemente la RA de las mezclas. El contenido de aniones y cationes de la fracción soluble de las mezclas se muestra en la Tabla 5.4. El contenido de fósforo total del purín fue bastante alto 2107 mg/kg, aunque el medido por electroforesis sobre el sobrenadante fue mucho más bajo, mostrando que la mayor parte del fósforo se encontraba en forma orgánica. Tabla 5.4. Análisis de cationes y aniones por electroforesis capilar (unidades mg/L)* Tratamiento K Ca 1.1(control) 1598,82 22,44 Na Mg Cl- SO42- PO43- 298,56 4,25 594,2 124,6 216,7 2129,12 Ptotal 1.2 1558,67 35,71 282,87 3,365 581,1 119,0 214,9 2024,82 1.3 1508,37 20,08 268,32 2,125 555,9 99,8 172,0 1784,59 1.4 1153,38 79,52 205,47 2,805 447,3 79,9 148,4 1704,92 2.1(control) 1598,82 22,44 298,56 4,25 594,2 124,6 216,7 2129,12 2.2 1671,32 27,61 274,47 10,175 610,0 106,8 109,0 2006,69 2.3 1926,01 47,12 312,71 5,46 587,1 114,1 75,6 1967,53 2.4 2125,09 67,71 314,64 4,46 598,1 86,3 70,4 na** 2.5 181,67 22,15 26,46 12,92 28,3 20,2 22,6 na** 3.1(control) 1598,82 22,44 298,56 4,25 594,2 124,6 216,7 2129,12 307,93 16,28 615,9 93,7 32,5 760,47 442,79 173,46 814,1 137,9 4,9 122,85 52,14 13,9 137,9 4,9 17,77 3.2 1526,0 3.3 1749,91 48,44 3.4 24,84 33,22 115,73 126,89 *valores medios de tres repeticiones;** n.a.: no analizado. Las concentraciones de potasio, y en menor medida de sodio, fueron bastante altas, y además la adición del cosubstrato pareció que hizo aumentar esta concentración. Las tierras decolorantes aportaron bastante magnesio, haciendo subir la concentración de la mezcla. También aumentó algo la concentración de calcio, pero de forma muy poco importante, de forma que 142 Ensayos de viabilidad de codigestión en discontinuo probablemente no tuvo influencia en la inhibición por ácidos grasos de cadena larga. 5.4.7. Cálculo de las concentraciones finales de los parámetros físicoquímicos La caracterización inicial se realizó para cada uno de los substratos y mezclas utilizadas. Sin embargo, las muestras obtenidas al final de la digestión estaban compuestas por el substrato digerido más la parte proporcional de inóculo. Para poder comparar los valores de los diferentes parámetros al inicio y al final del experimento se eliminó el efecto del inóculo. Dado que se realizaron blancos a las dos temperaturas (inóculo y agua, experimentos 4.1-35ºC y 4.1-55º) se pudo conocer la concentración final de los parámetros físico-químicos correspondientes al inóculo. Puesto que se sabe la proporción exacta de inóculo añadido al inicio se corrigieron los valores finales con los valores medios obtenidos para la digestión del blanco, de forma similar al cálculo de la producción de gas. La cantidad de substrato y de inóculo iniciales fueron 50 g y 5 g respectivamente. La concentración de cada parámetro al final de la digestión (P) se determinó de la siguiente forma: >Ps @ >P@ >PB @* 55 / 50 , (5.13) siendo [Ps] la concentración del parámetro P final corregida por el blanco; [PB] la concentración del parámetro P del “blanco” al final de la digestión (inóculo + agua digerido) y [P] la concentración del parámetro P al final medida (substrato + inóculo). Para calcular la concentración de amoníaco libre en el medio durante el proceso se utilizó el promedio de los valores iniciales y finales, tanto para la concentración de amonio como para el pH. 5.4.8. Análisis estadístico El análisis estadístico se realizó para cada experimento por separado, considerando un diseño de tipo factorial doble, siendo los efectos principales la temperatura y el tratamiento (% de dilución o % de cosubstrato) y para algunos parámetros factorial triple, al comparar también concentraciones iniciales y finales. Se realizó el análisis de la varianza mediante el paquete estadístico SAS. Si la interacción entre los diferentes efectos resulta significativa (a un 95% de nivel de confianza) se realiza un test Lsmeans para detectar diferencias entre los diferentes tratamientos. Si la interacción no es Optimización de la digestión anaerobia de purines de cerdo 143 significativa, pero sí lo son los efectos principales, la separación de medias se realiza mediante el test de Duncan. En todos los casos se utilizó un nivel de confianza del 95%. Para marcar las diferencias entre los diferentes tratamientos se asignaron letras diferentes. 5.5. Resultados y Discusión La presentación de resultados se ha dividido en función de los diferentes experimentos realizados, aunque todos los tratamientos se han comparado con el purín sin diluir, considerado como control. 5.5.1. Experimento 1. Diluciones de purín a dos temperaturas: efecto del amonio Los resultados se exponen según como han variado los principales factores considerados. Todos los valores expuestos en las tablas corresponden a los valores medios. Los valores finales de los parámetros físico-químicos fueron corregidos con la concentración correspondiente del blanco. Evolución de los compuestos nitrogenados. El contenido de nitrógeno en el purín utilizado estuvo en torno a los 5,20 g N-Nk/L, con un 65% en forma de nitrógeno amoniacal (Tabla 5.5). Tabla 5.5. Concentración de nitrógeno Kjeldahl (mg N/Kg) en el experimento 1 Tratamiento Inicial Final 35ºC Final 55ºC Dif. Temp 1.1-100% 5,26 d 5,36 d 5,35 d - 1.2- 95% 4,86 c 4,89 c 5,19 cd - 1.3- 87,5% 4,43 b 4,36 a 4,87 c * 1.4- 80% 4,12 a 4,06 a 4,08 a - Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significación del 95%). * o ** diferencias significativas para los valores finales entre temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. Hubo diferencias significativas entre los diferentes niveles de dilución en la concentración de nitrógeno Kjeldahl inicial. Sin embargo, para la mayoría de los niveles de dilución no hubo diferencias entre la concentración inicial y la final, así como entre los valores finales para cada temperatura. La única 144 Ensayos de viabilidad de codigestión en discontinuo posible pérdida de nitrógeno total del sistema sería la debida a la volatilización de amoníaco, pero es muy poco importante, ya que no se detectaron diferencias significativas en el contenido de nitrógeno total al inicio y al final. Sin embargo, el comportamiento del nitrógeno amoniacal fue diferente, encontrándose diferencias significativas para todos los niveles de dilución entre el inicio y el final de la digestión (Tabla 5.6), con un nivel de confianza del 5%. Los valores finales fueron diferentes para cada tratamiento en función de la temperatura y siempre mayores que en el inicio. Para casi todos los niveles de dilución se detectaron diferencias entre los valores finales a cada temperatura, siendo mayores en el rango termofílico que en el mesofílico. Tabla 5.6. Concentración de nitrógeno amoniacal (g N-NH4+ /L) en el experimento 1. Tratamiento Inicial Final 35ºC Final 55ºC Dif. Temp 1.1-100% 3,43 de 3,83 gh 4,09 i * 1.2- 95% 3,28 cd 3,66 fg 3,87 h * 1.3- 87,5% 2,97 b 3,44 de 3,58 ef - 1.4- 80% 2,63 a 3,11 bc 3,36 d * Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significación del 95%). * o ** diferencias significativas para los valores finales entre temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. Durante el proceso de digestión anaerobia se produce la mineralización del nitrógeno orgánico mediante la hidrólisis enzimática de las proteínas. La disminución de nitrógeno orgánico ocurrirá sólo en el caso de que la tasa de consumo de nitrógeno por parte de la biomasa sea menor que la tasa de hidrólisis de las proteínas y la degradación de los aminoácidos, que es lo normal en un sistema anaerobio, caracterizado por la baja formación de biomasa. Este hecho se constató al observar el aumento de nitrógeno amoniacal (Figura 5.4). El porcentaje de nitrógeno amoniacal sobre el nitrógeno total se puede considerar como un índice de degradación del substrato, por lo que el estudio de este parámetro puede proporcionar información acerca del proceso de hidrólisis de proteínas. Optimización de la digestión anaerobia de purines de cerdo 145 5000 + mg NH4 /L 4000 Inicial 3000 Final 35ºC 2000 Final 55ºC 1000 0 1.1 1.2 1.3 1.4 Figura 5.4. Comparación de la concentración de amonio antes y después de la digestión en el experimento 1. El índice de amonio sobre nitrógeno amoniacal aumentó durante la digestión (Tabla 5.7). El aumento de la concentración de nitrógeno amoniacal fue mayor en el rango termofílico en todos los tratamientos, es decir la hidrólisis de proteínas fue mayor a 55ºC, resultados conformes con los obtenidos por Gallert et al. (1998). Tabla 5.7. Proporción de nitrógeno amoniacal sobre nitrógeno total. Incremento sufrido durante la digestión en el experimento 1. Fracción N-NH4+/NK (%) Tratamiento Inicial Final 35ºC Final 55ºC 1.1-100% 64,7 71,91 1.2- 95% 66,1 1.3- 87,5% 1.4- 80% ' N-NH4+ /NK (%) 35ºC 55ºC Dif. Temp. 76,74 7,68 a 12,28 bc ** 73,74 77,69 7,80 a 11,82 bc * 65,6 75,31 78,53 10,20 ab 13,32 c * 64,3 76,15 82,25 11,41 bc 17,77 d ** Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significación del 95%). * o ** diferencias significativas para los valores finales entre temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. La interacción de los efectos principales, dilución y temperatura, no fue significativa para el incremento de la fracción inorgánica sobre la orgánica, aunque sí lo fueron los efectos principales. El incremento fue mayor en el rango termofílico, lo que concuerda con la afirmación de Gallert et al. (1998), que situó la biodegradabilidad en el rango termofílico por encima del 146 Ensayos de viabilidad de codigestión en discontinuo mesofílico. El efecto de dilución hizo aumentar el incremento de la fracción inorgánica, es decir la hidrólisis de proteínas fue mayor cuanto más diluido era el tratamiento, lo que indica un efecto de inhibición de la hidrólisis en los más concentrados. Esto está de acuerdo con resultados previos, ya que está descrito que las altas concentraciones de amonio en el medio pueden inhibir el crecimiento de organismos proteolíticos (Krylova et al., 1997; Gallert et al., 1998). Por otra parte, el que la hidrólisis fuera mayor en términos absolutos en el rango termofílico que en el mesofílico, y que por tanto, aumentara más la concentración de amonio, pudo tener un efecto negativo añadido sobre la metanogénesis, que como se sabe es un proceso biológico muy sensible a la concentración de amoníaco libre (Angelidaki y Ahring, 1993b). Evolución del contenido en materia orgánica Al diluir el purín se provocó la correspondiente dilución de materia orgánica, medida como sólidos volátiles o como demanda química de oxígeno. El análisis estadístico se realizó como un factorial triple en el que los factores principales son el nivel de dilución, la temperatura y el tiempo. El efecto tiempo se introdujo para comparar las concentraciones iniciales y finales. Durante la digestión se produjo la disminución de los sólidos totales y volátiles (Tabla 5.8 y Tabla 5.9), encontrando que a todos los niveles de dilución y a las dos temperaturas las diferencias fueron significativas a un nivel de significación del 1%. Al final de la digestión se detectaron diferencias significativas entre temperaturas para todos los niveles de dilución, excepto para el nivel 1.2 (95% de purín), siendo mayor la eliminación en el rango termofílico que en el mesofílico. En el rango mesofílico no se detectaron diferencias significativas en el porcentaje de eliminación de sólidos totales para los tres niveles de dilución 1.1, 1.2 y 1.3, mientras que en el termofílico, excepto los dos más concentrados, el resto fueron diferentes (Tabla 5.8). Los mismos resultados, pero con diferencias más claras, se obtuvieron al analizar la eliminación de sólidos volátiles (Tabla 5.9), indicando que no hubo ningún efecto de inhibición. Sin embargo, a 55ºC hubo un importante incremento en la eliminación conforme aumentó la dilución, lo que confirma la inhibición en el rango termofílico, atribuida al nitrógeno amoniacal. Para los dos tratamientos más concentrados (1.1 y 1.2), las eliminaciones de sólidos totales y volátiles fueron estadísticamente iguales, pero diferentes de los dos más diluidos, que a su vez fueron diferentes entre sí (Tabla 5.8 y Tabla 5.9). Optimización de la digestión anaerobia de purines de cerdo 147 Tabla 5.8. Concentraciones medias y eliminación de sólidos totales en el experimento 1. Sólidos Totales (ST) g/kg Eliminación ST(%) Tratamiento Inicial Final 35º C Final 55º C 35ºC 55ºC 1.1 – 100% 79,16 h 72,70 fg 68,30 de 8,00 A 12,81 BC 1.2 – 95% 75,21 g 66,65 cde 65,23 cd 10,42 AB 11,49 ABC 1.3 – 87.5% 69,26 ef 63,29 c 56,96 b 9,43 A 17,65 D 1.4 – 80% 63,33 cde 57,05 b 50,81 a 15,19 CD 22,26 E Letras diferentes significan valores medios diferentes. (Test Lsmeans a un nivel de significación del 95%). Tabla 5.9. Concentraciones medias y eliminación de sólidos volátiles en el experimento 1. Sólidos Volátiles (SV) g/kg Eliminación SV(%) Tratamiento Inicial Final 35º C Final 55º C 35ºC 55ºC 1.1 – 100% 58,14 a 50,27 d 45,82 ef 13,54 A 21,19 B 1.2 – 95% 54,91 b 46,23 e 43,41 f 15,80 A 20,95 B 1.3 – 87.5% 51,31 c 43,50 f 37,48 g 15,23 A 26,95 C 1.4 – 80% 48,20 d 38,60 g 32,81 h 19,92 B 31,92 D Letras diferentes significan valores medios diferentes. (Test Lsmeans a un nivel de significación del 95%). Los valores iniciales y finales de DQO (Tabla 5.10) fueron diferentes a todos los niveles de dilución en el rango mesofílico, pero no en el termofílico, en el que tan sólo se encontraron diferencias en los dos tratamientos más diluidos. Por otro lado, para todos los tratamientos, no se encontraron diferencias significativas en la DQO final, ni en la eliminación de DQO, entre las dos temperaturas. La DQO inicial no se pudo diferenciar entre los diferentes tratamientos, debido a la gran variabilidad de las repeticiones. La mayor eliminación de SV y ST a 55ºC no se corresponde con los valores de eliminación de DQO obtenidos (Tabla 5.8, Tabla 5.9 y Tabla 5.10), ni, como se verá más adelante, con los valores de producción de metano, que fueron mayores en el rango mesofílico que en el termofílico. Además, todos los valores de eliminación de materia orgánica son más bajos de lo que correspondería a la producción de metano medida. Las causas de estas 148 Ensayos de viabilidad de codigestión en discontinuo discrepancias pueden estar, por un lado, en el error analítico, que puede ser diferente en función de la temperatura debido a la diferente viscosidad. También pudo influir la extracción periódica de muestras de la fracción líquida, que pudo haber provocado una concentración del substrato a lo largo del experimento. Las diferencias entre ST/SV y DQO en los rangos mesofílico y termofílico, podrían explicarse, al menos en parte, por la fuerte acumulación de ácidos grasos volátiles del rango termofílico, fruto de la inhibición de la metanogénesis, que no estarían contabilizados en los parámetros ST y SV, debido al procedimiento analítico, y sí en el parámetro DQO. Tabla 5.10. Valores medios y eliminación de DQO en el experimento 1. DQO (g O2/kg) Eliminación DQO(%) Tratamiento Inicial Final 35º C Final 55º C 35ºC 55ºC 1.1 – 100% 80,04 f 62,01 cde 69,73 efg 23,37 AB 13,83 A 1.2 – 95% 76.87 fg 59,49 cd 67,47 def 22,61 AB 12,22 A 1.3 – 87.5% 70,79 efg 56,53 bc 56,27 abc 20,16 AB 20,52 AB 1.4 – 80% 65,44 cdef 49,48 a 44,79 ab 23,55 AB 30,81 B Letras diferentes significan valores medios diferentes. (Test Lsmeans a un nivel de significación del 95%). Evolución del pH El purín utilizado mostró un pH bastante alto (Tabla 5.11), lo que pudo condicionar el desarrollo de todo el proceso. El alto pH unido al alto contenido de amonio provocó altas concentraciones de amoníaco libre (importante inhibidor de la etapa metanogénica). Los valores de la alcalinidad fueron bastante altos, suficiente para el desarrollo de la digestión anaerobia, como muestra la pequeña variación de pH durante el proceso. Optimización de la digestión anaerobia de purines de cerdo 149 Tabla 5.11. Valores medios iniciales de pH y alcalinidad en el experimento 1 Tratamiento pH Alc. Total (g Alc. Parcial (g Alc. Int CaCO3/L) CaCO3/L) (g AcH/L) 1.1 – 100% 8,06 10,55 7,5 3,66 0,29 1.2 – 95% 7,96 10,05 7,3 3,3 0,27 1.3 – 87.5% 8,03 8,975 6,4 3,09 0,29 1.4 – 80% 7,95 8,2 5,8 2,88 0,29 RA La relación de alcalinidad mostró un valor relativamente bajo, lo que indica que se trata de un purín relativamente poco degradado. A partir de los valores iniciales y finales de pH y de amonio y de la temperatura de cada tratamiento, se calculó la concentración de amoníaco libre medio durante la digestión (como media del inicial y final), obteniéndose los valores que se muestran en la Tabla 5.12. Los valores obtenidos de amoníaco libre superan los valores descritos en la bibliografía como inhibidores tanto para el rango mesofílico como para el termofílico, aunque algunos trabajos han conseguido buenos rendimientos con concentraciones más altas (Hansen et al, 1998). Tabla 5.12. Valores medios de pH al inicio y al final de la digestión, y concentración media de amoníaco libre durante el proceso, en el experimento 1 N-NH3 (mg/L) pH Tratamiento Inicial Final-35ºC Final-55ºC 35ºC 55ºC 1.1 – 100% 8,06 d 7,70 bc 7,99 d 305,8 1106,2 1.2 – 95 % 7,96 d 7,59 ab 7,77 c 235,9 807,2 1.3 – 87,5% 8,03 d 7,54 a 7,65 abc 227,9 716,0 1.4 – 80 % 7,95 d 7,54 a 7,73 c 182,8 641,6 Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significación del 95%). Evolución de la producción de biogás Al realizar el análisis estadístico de los resultados obtenidos al final de la digestión se observó que la interacción entre los dos efectos principales (nivel de dilución y temperatura) fue significativa para la producción 150 Ensayos de viabilidad de codigestión en discontinuo acumulada, M (ecuación 5.1), de biogás, metano y dióxido de carbono (Tabla 5.13), así como para los índices de producción, B (ecuación 5.3), de biogás, metano y CO2 respecto a sólidos volátiles iniciales (Tabla 5.14). Tabla 5.13. Valores medios de la producción acumulada de gas (M) al final del proceso, para las variables biogás, metano y dióxido de carbono, en el experimento 1. Trat. M (mL de biogás) M (mL de CH4 ) 35º 35º 55º 55º M (mL de CO2) 35º 55º 1.1 658,68 d 436,99 a ** 443,13 D 195,18 A ** 215,55 BC 241,81 C - 1.2 622,44 cd 439,86 a ** 427,70 D 212,26 A ** 194,74 AB 227,60 BC - 1.3 533,55 b 494,27 ab - 367,99 C 256,73 AB ** 165,57 AB 237,54 C ** 1.4 546,59 bc 533,78 b - 370,84 C 297,41 B ** 175,75 AB 236,37 C ** Letras diferentes significan valores medios diferentes. (Test Lsmeans a un nivel de significación del 95%). * o ** diferencias significativas para los valores finales entre temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. La producción de metano fue significativamente mayor en el rango mesofílico que en el termofílico para todos los tratamientos, tanto en unidades de producción acumulada como el índice de producción respecto a los sólidos volátiles iniciales. Sin embargo la producción de CO2 fue mayor a 55ºC, existiendo diferencias significativas entre temperaturas para los tratamientos 1.3 y 1.4 (los más diluidos). La consecuencia inmediata es la producción de un biogás con un menor contenido en metano. En el rango mesofílico está cercano al 70%, mientras que en el termofílico varía según la dilución, pero está entre un 45% y un 56%. La existencia de diferencias significativas a favor del termofílico en la producción de dióxido de carbono hace que los tratamientos 1.3 y 1.4 resulten iguales para las dos temperaturas en cuanto a la producción acumulada de biogás total. Al comparar la producción acumulada, de biogás y de CH4, entre los diferentes tratamientos se observa una tendencia diferente para los dos rangos de temperatura considerados. A 35ºC la producción acumulada de metano fue mayor en los tratamientos más concentrados, mientras que a 55ºC la máxima producción se alcanzó para el tratamiento más diluido, siendo éste diferente a un nivel de significación del 5% de los otros tres (Tabla 5.13). Al comparar el índice de producción de metano respecto a los sólidos volátiles iniciales se eliminaron las diferencias entre niveles de dilución a Optimización de la digestión anaerobia de purines de cerdo 151 35ºC, demostrando que la menor producción acumulada alcanzada por los más diluidos se produjo por la menor cantidad de materia orgánica disponible para la degradación. En cambio, a 55ºC las diferencias se acrecentaron, pudiéndose distinguir claramente los dos tratamientos más concentrados de los dos más diluidos, existiendo, también , diferencias entre ellos (Tabla 5.14). Tabla 5.14. Valores medios de los índices de producción acumulada de gas respecto al contenido inicial de sólidos volátiles (B), para las variables biogás, metano y dióxido de carbono, en el experimento 1. B (mL biogás/g SV) 55º B (mL CH4/g SV) B (mL CO2/g SV) 35º 35º Trat. 35º 1.1 223,16 c 148,05 a ** 150,13 D 66,13 A ** 73,0 ABC 81,93 CD - 1.2 221,98 c 156,86 a ** 152,53 D 75,69 A ** 69,45 AB 81,17 BCD - 1.3 206,59 c 191,38 bc - 142,48 CD 99,41 B ** 64,11 A 91,97 DE ** 1.4 231,48 c 226,06 c 55º 55º - 157,05 D 125,95 C ** 74,43ABC 100,10 E ** Letras diferentes significan valores medios diferentes. (Test Lsmeans a un nivel de significación del 95%).* o ** diferencias significativas para los valores finales entre temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. Tanto en el rango termofílico, como en el mesofílico, no se detectaron diferencias significativas entre los diferentes niveles de dilución para la variable producción acumulada de CO2. A 55ºC el índice de producción de CO2 respecto a SVini fue mayor para los tratamientos más diluidos, pudiendo establecerse diferencias significativas con los más concentrados, con una tendencia similar a la del índice de producción de metano. La evolución de la producción acumulada de metano para todos los tratamientos de este experimento se muestra en la Figura 5.5. La producción acumulada de metano en el rango mesofílico fue bastante similar entre los diferentes niveles de dilución durante los primeros días, hasta el día 20, momento a partir del cual se comenzaron a diferenciar. Este comportamiento inicial similar implica que la concentración de substrato no fue limitante del crecimiento bacteriano hasta ese momento. La evolución de los tratamientos a 55ºC fue también muy similar hasta el día 13, momento en el que el tratamiento más diluido se separó del resto, encontrándose diferencias significativas con nivel de confianza del 99% a partir del 15. 152 Ensayos de viabilidad de codigestión en discontinuo mL de CH4 500 450 400 35º 1.1 350 35º 1.2 300 35º 1.3 250 35º 1.4 200 55º 1.1 150 55º 1.2 100 55º 1.3 50 55º 1.4 0 0 10 20 30 40 50 60 70 80 Día Figura 5.5. Evolución de la producción de metano acumulado en el experimento 1 La mayor producción de metano alcanzada en el rango mesofílico, y la mayor producción de los tratamientos más diluidos en el rango termofílico, indica la presencia de un tóxico en el rango termofílico. Los valores obtenidos de producción de metano respecto a sólidos volátiles a 55ºC son del mismo orden de magnitud que los obtenidos al digerir purín de cerdo en un sistema de mezcla completa por Hansen et al. (1998), 67 mL de CH4/d·gSV, sin embargo en mesofílico los valores obtenidos fueron algo menores que los obtenidos por el citado autor. En el rango mesofílico se observa en la curva de evolución de dióxido (Figura 5.6) de carbono un salto cualitativo en la producción acumulada en el rango mesofílico alrededor del día 13. La causa fue la acumulación de una gran presión de biogás en el espacio de cabeza de 140 mL de metano, que unido al volumen inicial se convirtió en una presión de más de 3 atmósferas, lo que probablemente desplazó el equilibrio líquido-gas del CO2 hacia la fase líquida. El día 13 se procedió al vaciado del espacio de cabeza de los viales, por lo que disminuyó la presión parcial del CO2 y se desplazó el equilibrio líquido-gas hacia la fase gas, dada la alta concentración presente en ese momento en el volumen de líquido. A partir de ese momento no se permitió que aumentara tanto la presión en el vial, de forma que no se obtuvieron saltos tan importantes. Optimización de la digestión anaerobia de purines de cerdo 153 mL de CO2 300 35º 1.1 250 35º 1.2 200 35º 1.3 35º 1.4 150 55º 1.1 55º 1.2 100 55º 1.3 50 55º 1.4 0 0 10 20 30 40 50 60 70 80 Día Figura 5.6. Evolución de la producción acumulada de CO2, en el experimento 1. A partir del día 13 hubo un aumento de la pendiente en el tratamiento 1.4 (80% de purín) a 55ºC. Este cambio de pendiente coincide con el cambio de pendiente en la producción acumulada de dióxido de carbono y un cambio de tendencia en la evolución de los ácidos grasos, en especial del acético. A partir de este momento comienza a disminuir de forma apreciable la concentración de ácidos grasos, por lo que se puede considerar que la metanogénesis a partir del acético se empieza a producir a un ritmo adecuado. Análisis de parámetros de velocidad, producción potencial y duración del desfase inicial El ajuste a la cinética de primer orden (ecuación 5.4) resultó en coeficientes de regresión bajos y gran variabilidad en los valores calculados para los diferentes parámetros. Sin embargo el ajuste mediante la ecuación Gompertz (ecuaciones 5.5 y 5.6) resultó en coeficientes de regresión altos, comprendidos entre 0.95-0.99 para todas las repeticiones. Una vez realizado el cálculo de los estimadores para cada vial, se realizó el análisis de la varianza de los estimadores obtenidos. Se aplicó la ecuación al cálculo tanto de la producción potencial M0, como del índice de producción potencial respecto a los sólidos volátiles iniciales B0, (ecuaciones 5.5 y 5.6). El ajuste para el índice de producción de metano sobre SVini se ilustra en la Figura 5.7. Los valores de M0 (producción potencial acumulada de metano) y B0 (índice de producción de metano sobre SVini) alcanzaron valores más altos en el régimen mesofílico que en el termofílico, para todos los niveles de 154 Ensayos de viabilidad de codigestión en discontinuo dilución. El análisis estadístico se realizó considerando como factores principales el nivel de dilución y la temperatura, analizando también la interacción entre ambos. Para todas las variables, excepto la duración del desfase (O) la interacción entre los dos factores principales fue significativa a un nivel del 5%, por lo que se procedió al análisis de las interacciones mediante un test Lsmeans (Tabla 5.15). Ni la interacción entre los dos efectos principales, ni el nivel de dilución resultaron significativos para la duración del desfase inicial (O). El efecto temperatura si fue significativo y con un nivel de confianza muy alto, pudiéndose decir que el desfase es mayor en el rango termofílico que en el mesofílico con una probabilidad de error menor del 1%. Puesto que no se ha relacionado con el grado de dilución en el rango termofílico, puede que en todos los tratamientos se haya superado el nivel inhibitorio por amonio, o bien que no sea consecuencia de la presencia de tóxicos, pudiendo atribuirse a una menor adaptación del inóculo. mL CH4/gSVini mL CH4/gSVini 180 180 1.4-35ºC 160 160 1.2-35ºC 140 140 120 120 100 100 1.1-35ºC 1.3-35ºC 80 1.4-55ºC 1.3-55ºC 80 60 60 40 40 20 20 0 1.2-55ºC 1.1-55ºC 0 0 20 1.1-35ºC 1.2-35ºC 1.3-35ºC 1.4-35ºC 40 60 Ajuste 1.1 sim-35º 1.2 sim-35º Gompertz 1.3 sim-35º 1.4 sim-35º 80 Día 0 20 1.1-55ºC 1.2-55ºC 1.3-55ºC 1.4-55ºC 40 60 1.1 sim-55º Ajuste 1.2 sim-55º Gompertz 1.3 sim-55º 1.4 sim-55º 80 Día Figura 5.7. Evolución temporal de el índice de producción acumulada respecto a sólidos volátiles iniciales, en el experimento 1. Los valores de R2 estuvieron en todos los casos entre 0,95 y 0,99 Optimización de la digestión anaerobia de purines de cerdo 155 Tabla 5.15. Valores medios de parámetros estimados mediante la ecuación de Gompertz (ecuaciones 5.5 y 5.6), en el experimento 1. K ml CH4/d O d B0 mL CH 4 g SV R mL CH 4 g SV·d R’ d-1 1.1 35ºC 428,83 d 11,08 cd 4,04 abc 144,98 e 3,76 bc 0,072 bc 1.2 35ºC 391,36 cd 12,32 d 3,99 abc 139,32 cde 4,43 c 0,086 c 1.3 35ºC 339,75 bc 10,66 cd 3,07 ab 131,33 cde 4,16 bc 0,087 c 1.4 35ºC 339,73 bc 10,24 c 2,57 a 143,59 de 4,38 c 0,083 c 4,2 a 6,3 bcd 70,21 a 1,43 a 0,056 ab 1.2 55ºC 265,12 ab 4,03 a 9,63 d 94,62 ab 1,44 a 0,046 a 1.3 55ºC 293,43 ab 4,43 a 7,18 cd 113,63 bcd 1,72 a 0,042 a 1.4 55ºC 292,02 ab 8,15 b 7,49 cd 123,67 bcd 3,45 b 0,077 c Trat. 1.1 55ºC M0 mL CH4 207,2 a Letras diferentes significan valores medios diferentes. (Test Lsmeans a un nivel de significación del 95%). Análisis estadístico realizado por separado para cada variable (cada columna). El parámetro M0 (producción potencial acumulada de metano) mostró diferencias significativas a 35ºC entre tratamientos, siendo mayor en los más concentrados. Sin embargo, el índice de producción respecto a sólidos volátiles (B0) no mostró diferencias. A 55ºC, el parámetro M0 no mostró diferencias significativas entre tratamientos, y sin embargo para el índice B0 hubo diferencias significativas entre el tratamiento 1.1 y los dos más diluidos (1.3 y 1.4). Al comparar entre temperaturas se observa que tanto para la producción potencial (M0) como para el índice de producción (B0) hubo diferencias significativas entre las dos temperaturas para los dos tratamientos más concentrados, mientras que para los dos más diluidos no se pudieron diferenciar. Al estudiar los parámetros de velocidad de producción de metano, medido como K (ml CH4/día) o R (mL CH4 /g SVini* dia) o R’ (dia-1), se vio que en el rango mesofílico no existieron diferencias significativas a un nivel del 5% entre tratamientos para ninguna de las variables. Sin embargo en el rango termofílico en todos los casos hubo una clara diferencia entre el tratamiento más diluido (1.4) y el resto. La velocidad de producción de metano (K) fue en todos los casos diferente a 35º y a 55ºC, siendo el factor temperatura significativo a un nivel de significación del 1%. El tratamiento 1.4, pese a presentar una velocidad 156 Ensayos de viabilidad de codigestión en discontinuo superior al resto de tratamientos en el rango termofílico no llegó al nivel alcanzado a 35ºC. Al eliminar el efecto de la concentración de substrato (variable R) las diferencias fueron menores, y al comparar la variable R’ (en unidades de T-1) no pudo ser separado de su homólogo a 35ºC. En el rango termofílico sí se observaron importantes diferencias entre tratamientos, tanto al comparar velocidad de producción, como producción real y potencial acumulada respecto a sólidos volátiles. En valor absoluto la producción de metano fue mayor en el tratamiento más diluido (1.4), pero no se encontraron diferencias significativas entre los valores obtenidos mediante simulación por Gompertz para los diferentes tratamientos. El efecto de inhibición quedó contrarrestado por la mayor cantidad de substrato disponible. Índice de metanización, acidificación y biodegradabilidad El índice de metanización y acidificación se calcularon según las ecuaciones 5.8 y 5.9, mostrándose los valores obtenidos en la Tabla 5.16. En el rango mesofílico no se detectaron diferencias para ninguno de los tres índices. Indicando que no estuvieron influidos, ni la metanogénesis, ni la cidogénesis por ningún inhibidor. Sin embargo, en el rango termofílico, sí hubo importantes diferencias entre tratamientos, aumentado conforme aumentó el grado de dilución, y por tanto influido por la presencia de inhibidores. Tabla 5.16. Indice de metanización, acidificación y biodegradabilidad en el experimento 1. Metanización (%M) Acidificación 55ºC 35ºC (%A) 55ºC Biodegradabilidad (%BD) Trat. 35ºC 35ºC 1.1 29,44 d 12,96 a ** 29,54 B 19.68 A ** 36,63 B 1.2 29,60 d 14,72 a ** 29,69 B 22.36 A ** 36,83 B 27.30 A ** 1.3 27,66 d 19,30 b ** 27,75 B 22.74 A ** 34,37 B 27.86 A ** 1.4 30,12 d 24,20 c ** 30,23 B 26.90 B - 37,53 B 55ºC 23.94 A ** 33.20 B - Letras diferentes significan valores medios diferentes. (Test Lsmeans a un nivel de significación del 95%). Análisis estadístico realizado por separado para cada uno de los días y cada variable. * o ** diferencias significativas para los valores finales entre temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. No sólo el índice de metanización mostró diferencias, sino que también el Optimización de la digestión anaerobia de purines de cerdo 157 índice de acidificación fue diferente para todos los tratamientos, excepto para el más diluido, y lo mismo ocurre con el índice de biodegradabilidad. Las diferencias entre temperaturas, y entre tratamientos, disminuyeron, al comparar el índice de acidificación, pero continuaron siendo significativas. La existencia de diferencias significativas en el índice de acidificación indican que no sólo la fase de metanogénesis resultó afectada, sino también los procesos de acidogénesis e hidrólisis. La inhibición de la fase metanogénica puede provocar la inhibición de fases previas, puesto que la acumulación de acético puede provocar la inhibición de la acetogénesis (Fukuzaki et al., 1990, Ahring y Westermann, 1988) e incluso la inhibición de la hidrólisis/acidogénesis (Angelidaki et al., 1993), lo que puede explicar los resultados obtenidos. El amonio puede inhibir la hidrólisis de proteínas, tal y como se deduce de los resultados diferentes de incremento de nitrógeno amoniacal sobre el total (Tabla 5.7), lo que podría haber tenido influencia sobre el índice de acidificación. Evolución de ácidos grasos volátiles (AGV). En la Figura 5.8 se observa la evolución de la concentración total de ácidos grasos volátiles en mM. Tanto a 35º C como a 55ºC se observó una fuerte acumulación inicial de ácidos, más acentuada en el régimen termofílico. La evolución de la concentración de AGV en ambos ensayos fue similar hasta el día 6 para las dos temperaturas, momento a partir del cual se comenzaron a diferenciar (Tabla 5.17), mostrando diferencias significativas. Dentro del régimen termofílico la mayor acumulación de ácidos se produjo para los dos tratamientos más concentrados (55º 1.1 y 1.2), lo que concuerda con la menor producción de metano observada. No sólo llegaron a unos niveles máximos más altos (153 mM), sino que se mantuvo durante un período más largo, no observándose una reducción importante hasta pasado el día 21, y aún así la concentración al final del experimento era alta (Figura 5.8 y Tabla 5.17). El tratamiento más diluido (1.4) se diferenció del resto desde el inicio, acrecentándose las diferencias a lo largo del tiempo. Entre los otros tres tratamientos, inicialmente no se apreciaron diferencias, aunque al final, el tratamiento 1.3 fue significativamente diferente de los tratamientos 1.1 y 1.2. En todos los casos, al final del experimento la concentración se mantuvo a niveles relativamente altos, indicando la poca estabilidad del proceso. En el rango mesofílico todos los tratamientos mostraron un comportamiento similar a lo largo del tiempo, tal y como se observa en la Figura 5.8. En ninguno de los tres puntos analizados (día 6, día 13 y final) se 158 Ensayos de viabilidad de codigestión en discontinuo observaron diferencias significativas entre los diferentes tratamientos, a un nivel de confianza del 95% (Tabla 5.17). La concentración de ácidos total alcanzó el máximo absoluto el día 6, y un máximo relativo el día 13, momento a partir del cual comenzó una clara tendencia a disminuir, estando en el día 20 a un nivel de 1 g/L, y a partir del día 30 a unas concentraciones muy bajas, al límite de la detección. AGVt (mM)-35ºC 175 AGVt (mM)-55ºC 175 150 150 125 125 100 100 75 75 50 50 25 25 0 0 0 10 20 30 40 Día 50 55º 1.1 35º 1.1 60 70 80 55º 1.2 35º 1.2 0 10 55º 1.3 35º 1.3 20 30 40 Día 50 60 70 80 55º 1.4 35º 1.4 Figura 5.8. Evolución de la concentración total de Ácidos Grasos Volátiles (C2-C5), en el experimento 1 El ácido acético fue el AGV que mayor acumulación sufrió en todos los casos (Figura 5.9). La concentración de acético en todos los tratamientos a 55ºC se mantuvo por encima de los homólogos a 35ºC a partir del día 6 y hasta el final del experimento. En el rango mesofílico la concentración máxima de acético para los cuatro tratamientos estuvo por debajo de 4 g Ac/L. A partir del máximo inicial, la concentración bajó rápidamente estando el día 20 por debajo de los 0.5 g/L para el purín 100% (1.1) y por debajo de 0,1 g/L para el purín 80% (1.4). En el rango termofílico la acumulación de acético en los primeros días mostró una tendencia similar a la observada en el rango mesofílico. Para los tratamientos más concentrados (1.1 y 1.2) se encontraron diferencias significativas respecto a los mesofílicos, ya en el día 6 (Tabla 5.17). A 55ºC la acumulación de acético continuó aumentando hasta alcanzar un máximo el día 13, con unas concentraciones muy altas, en torno a 8 g Ac/L para los tratamientos 1.1 y 1.2 a 55ºC y algo más bajo, 5.4 g Ac/L para el tratamiento 1.4 a 55ºC. Optimización de la digestión anaerobia de purines de cerdo 159 Tabla 5.17. Diferencias entre los valores de AGV total en el día 6 (máximo mesofílico), 13 (máximo termofílico), y final (valores en mM), en el experimento 1 Día 6 Trat. 35º C Día 13 55ºC 35º C Final 55ºC 35º C 55ºC Concentración total de ácidos grasos volátiles (mM) 1.1 81,5 a 96,76 b * 63,99 C 153,17 A ** 1,07 C 62,23 A ** 1.2 84,53 ab 95,77 b - 62,70 C 151,40 A ** 1,11 C 62,95 A ** 1.3 73,73 a 86,22 ab - 53,84 C 135,18 A ** 1,06 C 19,71 B ** 1.4 78,92 a - 41,35 C 107,41 B ** 0,86 C 17,35 B * 82,73 a Concentración de acético (mM) 1.1 63,03 ab 79,60 c ** 26,17 C 133,12 A ** 1,05 B 46,28 A ** 1.2 65,78 ab 79,13 c * 22,30 C 130,83 A ** 1,11 B 43,63 A ** 1.3 57,00 a 71,62bc - 18,95 C 116,99 A ** 1,06 B 10,90 B - 1.4 61,03 ab 69,42bc - 12,43 C 90,41 B ** 0,86 B 7,91 B - Concentración de propiónico (mM) 1.1 10,26 b 5,82 a ** 29,36 C 7,32 A ** nd 6,05 B ** 1.2 10,56 b 5,71 a ** 31,22 C 7,42 A ** nd 7,58 C ** 1.3 9,46 b 5,05 a ** 27,30 BC 7,08 A ** nd 4,11 A ** 1.4 10,38 b 5,07 a ** 6,16 A ** Nd 5,55 B ** 23,44 B Letras diferentes significan valores medios diferentes. (Test Lsmeans a un nivel de significación del 95%). Análisis estadístico realizado por separado para cada uno de los días y cada variable. * o ** diferencias significativas para los valores finales entre temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. nd: no detectado. A 55ºC, la eliminación de acético fue más rápida para los tratamientos más diluidos, así el día 37 el tratamiento 1.4 mostró una concentración de 1,1 g Ac/L, frente a los 3,73 g Ac/L del Purín 100%. Al final del experimento se observaron diferencias entre los dos tratamientos más concentrados y los dos más diluidos, aunque entre cada uno de estos grupos no se pudieron diferenciar y continuó siendo importante hasta el final del experimento en los tratamientos más concentrados a 55ºC, aunque con una ligera tendencia a disminuir, lo que indica actividad microbiológica después de 79 días de 160 Ensayos de viabilidad de codigestión en discontinuo digestión. Acético (mM)-55ºC 140 120 Acético (mM)-35ºC 140 120 100 100 80 80 60 60 40 40 20 0 20 0 0 10 20 30 40 50 60 70 0 80 10 20 30 40 50 60 70 80 40 Día 50 60 70 80 Propiónico (mM)-55ºC 50 Propiónico (mM)-35ºC 50 40 40 30 30 20 20 10 10 0 0 0 10 20 30 40 50 60 70 80 0 10 Día 55º 1.1 35º 1.1 55º 1.2 35º 1.2 55º 1.3 35º 1.3 20 30 55º 1.4 35º 1.4 Figura 5.9. Evolución del ácido acético y propiónico en mesofílico y termofílico, en el experimento 1 La acumulación inicial de acético, ocurrida en todos los tratamientos, a las dos temperaturas, se pudo deber a un desequilibrio inicial entre las poblaciones de microorganismos, ya que la velocidad específica máxima para los microorganismos acidogénicos es mayor que para los metanogénicos (Henze, 1996). La rápida disminución de acético a partir del día 6 en el rango mesofílico indica que el proceso metanogénico se reactivó y comenzó a funcionar adecuadamente a partir de este momento. El hecho de no detectar diferencias significativas entre los diferentes niveles de dilución, tanto para la concentración de acético como para la de AGV total, unido a que no se detectaron diferencias en el índice de producción de metano respecto a sólidos volátiles (Tabla 5.14), indica que no hubo procesos de inhibición bacteriana. Por el contrario, en el rango termofílico la hipótesis formulada sobre los Optimización de la digestión anaerobia de purines de cerdo 161 resultados de producción de metano se confirma con la gran acumulación de AGV, en especial de acético. Al diluir y por tanto, al disminuir la concentración de tóxico en el sistema, el proceso metanogénico se vio favorecido, motivo por el cual los tratamientos más diluidos (1.3 y 1.4) obtuvieron mayores producciones de metano respecto a la materia orgánica inicial (Tabla 5.14) y menores acumulaciones de AGV (Tabla 5.17). La evolución de los ácidos grasos volátiles C3-C5 fue muy diferente a las dos temperaturas testadas (Figura 5.10). En el rango mesofílico la concentración de propiónico y de las formas “iso” del butírico y el valérico continuaron aumentando hasta alcanzar un máximo absoluto el día 14, con un valor muy alto para el propiónico (mucho mayor al obtenido en el régimen termofílico), especialmente para los tratamientos más concentrados (Tabla 5.17), así el tratamiento 1.2 –35ºC superó los 30 mM (2200 mg /L). A partir de este máximo se produjo una eliminación muy rápida del propiónico, de forma que en el día 37 no se detectó para ningún tratamiento. Las formas iso del butírico y el valérico mostraron también el máximo absoluto el día 14, momento a partir del cual comenzaron a disminuir. Los máximos alcanzados fueron de mucha menor cuantía que en el caso del isobutírico (3,8 mM) y el isovalérico (5,2 mM). La evolución de los diferentes tratamientos a 35ºC fue muy similar, siendo tan sólo diferente el tratamiento más diluido, tal y como se observa en la Figura 5.10. Las formas “n”, tanto del butírico como del valérico (Figura 5.10) siguieron, sin embargo, una tendencia similar a la tendencia del acético, y completamente diferente que la del propiónico y las formas iso, alcanzando el máximo absoluto el día 6, momento a partir del cual disminuyeron rápidamente. Al correlacionar la concentración de acético con las concentraciones de las formas n del butírico y el valérico (considerando todos los datos de los tres experimentos en el rango mesofílico) se obtuvieron coeficientes de correlación relativamente altos (alrededor de 0,8). Concentraciones más bajas de acético que las obtenidas en el presente experimento se han postulado como inhibidoras para la degradación de propiónico y butírico (Ahring y Westermann, 1988; Fukuzaki et al., 1990; Hyun et al., 1998). Por tanto, la causa más probable de la acumulación de las formas n de butírico y valérico fue la inhibición de la acetogénesis por acético, y por tanto debido a la inhibición de los microorganismos metanogénicos acetoclásticos por amonio. La acumulación de propiónico a partir del día 6 indica la inhibición de la fase acetogénica, de degradación del propiónico. Diversos autores han 162 Ensayos de viabilidad de codigestión en discontinuo relacionado el aumento de la presión parcial de hidrógeno (pH2), con la acumulación de propiónico y butírico (Fukuzaki, et al., 1990, Harper y Pohland, 1986). En el presente experimento la relación propiónico/acético superó durante varios días el valor propuesto por Hill et al. (1987) como indicador de fracaso (1,4) de la digestión anaerobia en sistemas tipo CSTR. La presencia de las formas iso de los ácidos butírico y valérico se ha relacionado con el mal funcionamiento de reactores anaerobios en continuo (tipo CSTR), a unos niveles de concentración por encima de 15 mg/L (0,2 mM) (Cobb y Hill, 1991), y el aumento de la fracción iso sobre la concentración total (suma de los dos isómeros) se ha asociado con el aumento de la presión parcial de hidrógeno (Hill y Cobb, 1993). Por tanto, una posible causa de la acumulación de propiónico y de las formas iso del butírico y del valérico podría ser el aumento de la presión parcial de hidrógeno, debido a un desequilibrio entre la formación y el consumo, siendo un efecto tipo sobrecarga orgánica. Otra posible causa podría haber sido el aumento de la presión parcial de CO2, ya que coincidió con una importante acumulación de gas en el vial, llegándose a sobrepresiones muy importantes (del orden de 3 bares), aumentado, por tanto, la concentración de CO2 en la fase líquida. Aumentos en la presión parcial de CO2 han sido relacionados con acumulaciones de propiónico y en menor medida acumulaciones de butírico e iso-butírico, aunque no provoca la inhibición de los consumidores de hidrógeno (Hansson y Molin, 1981). En el rango termofílico, el nivel de propiónico se mantuvo aproximadamente constante durante todo el proceso, a la concentración alrededor de 5 mM (350-400 mg /L). Se produjo una ligera acumulación coincidiendo con la máxima acumulación de acético. Igualmente, la concentración de los isómeros iso del butírico y el valérico se mantuvieron a un nivel relativamente constante. Tan sólo el ácido iso-valérico en el tratamiento más diluido mostró cierta tendencia a disminuir. El isómero n del butírico en el rango termofílico, de forma similar a lo ocurrido en el rango mesofílico, sufrió una evolución influida por la evolución del acético, mostrando un valor más alto que en el rango mesofílico y con una tendencia a disminuir cuando la concentración de acético baja de un nivel de 60 mM. Sin embargo el n-valérico pareció no estar afectado por la concentración de acético, con unos valores medios inferiores a los máximos alcanzados en mesofílico, y de un orden de magnitud entre 10-15 veces menor que el n-butírico. Optimización de la digestión anaerobia de purines de cerdo Iso-butírico (mM )-35ºC 5 163 Iso-butírico (mM )-55ºC 5 4 4 3 3 2 2 1 1 0 0 0 10 20 30 40 50 60 70 0 80 N-butírico (mM )-35ºC 5 10 20 30 40 50 60 70 80 40 50 60 70 80 40 50 60 70 80 40 Día 50 60 70 80 N-butírico (mM )-55ºC 5 4 4 3 3 2 2 1 1 0 0 0 10 20 30 40 50 60 70 0 80 Iso-valérico (mM )-35ºC 10 10 20 30 Iso-valérico (mM )-55ºC 10 8 8 6 6 4 4 2 2 0 0 0 10 20 30 40 50 60 70 0 80 N-valérico (mM)-35ºC 1.0 10 20 30 N-valérico (mM)-35ºC 1.0 0.8 0.8 0.6 0.6 0.4 0.4 0.2 0.2 0.0 0.0 0 10 20 30 40 Día 50 55º 1.1 35º 1.1 60 70 80 55º 1.2 35º 1.2 0 10 55º 1.3 35º 1.3 20 30 55º 1.4 35º 1.4 Figura 5.10. Evolución de los dos isómeros del ácido valérico y butírico para las dos temperaturas, en el experimento 1 En el rango termofílico la acumulación de ácido acético no provocó una 164 Ensayos de viabilidad de codigestión en discontinuo acumulación importante de los ácidos grasos de cadena larga, al contrario de lo ocurrido en el rango mesofílico. Esto es, a pesar de que en el rango termofílico la fracción potencialmente inhibidora de acético (la fracción no ionizada) (Fukuzaki et al., 1990) es mayor que en el rango mesofílico a igualdad de pH y concentración total. Según los resultados obtenidos en este experimento los microorganismos acetogénicos termofílicos serían menos sensibles a la acumulación de acético que los mesofílicos. La alta concentración de acético en los tratamientos en el rango termofílico, junto con el bajo valor de la relación propiónico/acético (por debajo de 0,13), indica la inhibición de la fase metanogénica acetoclástica. Aunque a partir del día 21 se observó tendencia a disminuir, la concentración al final, sobre todo para los dos tratamientos más concentrados, continuó siendo muy alta (por encima de 60mM). La causa de la acumulación de acético se puede atribuir a la inhibición por amoníaco, dada la alta concentración de amonio (Tabla 5.6) y el alto pH del medio (Tabla 5.12), pudiendo haber influido la poca adaptación del inóculo termofílico a las altas concentraciones de amoníaco. La baja producción de metano obtenida en el intervalo inicial (hasta el día 6 para el rango mesofílico y hasta el día 25 para el termofílico) se pudo deber principalmente a la ruta metanogénica a partir de Hidrógeno, que es más rápida (presentan mayores velocidades de crecimiento) y menos sensible a la inhibición por amoníaco (Hansen et al., 1998; Robbins et al., 1989; Angelidaki y Ahring, 1993b). El alto nivel de acético junto con la baja, pero existente producción de metano, es el proceso que Angelidaki y Ahring (1993b) definieron como estado estacionario inhibido, en el cual hay actividad de los microorganismos anaerobios, pero con unos niveles de producción mucho más bajos que los potenciales. Relación entre amoníaco libre y producción de metano De la comparación entre los diferentes tratamientos en el régimen termofílico se observa que el proceso se encuentra influido por el grado de dilución, pudiendo relacionarse con el nivel de concentración de amonio y de amoniaco libre, presente en los diferentes tratamientos. En el rango mesofílico, no se observó dependencia de la producción de metano respecto al grado de dilución, no pudiendo correlacionarse la producción de metano con la concentración de amoníaco libre. Esto indica que la concentración de amoníaco libre que inhibe la metanogénesis está por encima de 305 mg N-NH3/L. Este valor es superior al propuesto por algunos autores, por ejemplo Gallert et al. (1998) lo cifraban en 92 mg de N- Optimización de la digestión anaerobia de purines de cerdo 165 NH3/L en el rango mesofílico, y Hashimoto (1986) encontró el límite en 30 mg de N-NH3/L en el rango mesofílico para cultivos no aclimatados, utilizando como substrato estiércol bovino. Puesto que los metanogénicos termofílicos son menos sensibles a la inhibición por amoníaco libre que los mesofílicos (Gallert et al., 1998) es de suponer que el nivel de inhibición para los termofílicos deberá estar, al menos, por encima de los 300 mg N-NH3/L. Al comparar el índice de producción de metano respecto a los sólidos volátiles iniciales con el amoníaco libre, considerando los datos experimentales correspondiente a las dos temperaturas, se observó una clara correlación entre las dos variables, que podría ajustarse relativamente bien a una recta, con un coeficiente de regresión r2 de 0,81 (Figura 5.11). Los datos del rango termofílico considerados de forma independiente se pueden ajustar a varios modelos, aunque con coeficientes de regresión más bajos (Figura 5.11). mL CH4/g SVini 200 Zona M esofílica Zona termofílica 180 160 140 CH 4 = -0.1125NH 3 + 183.76 R2 = 0.53 120 100 CH 4 = -0.1025NH 3 + 175.28 80 R2 = 0.81 60 CH4=EXP(3.24+968/NH 3) 40 R2=0.62 20 0 0 200 400 600 800 1000 1200 mg N-NH 3 Figura 5.11. Correlación valores experimentales de producción de metano respecto a sólidos volátiles (B) y concentración media de amoníaco libre (Experimento 1) Los dos tratamientos más concentrados a 55ºC no fueron estadísticamente diferentes a un nivel del 10%, aunque sí lo fueron de los dos más diluidos, con una producción de metano mucho mayor. Tal y como se observa en la Figura 5.11, al considerar sólo los datos en termofílico, el modelo se ajusta mejor a una curva de tipo exponencial que al modelo lineal, ya que la variación en la producción de metano es mucho más acusada en el tramo de concentraciones de amoníaco libre que va de 600 a 800 mg de NH3/L, lo 166 Ensayos de viabilidad de codigestión en discontinuo que está de acuerdo con resultados de la bibliografía (Angelidaki y Ahring, 1993b). Se realizó también la comparación de los parámetros obtenidos mediante el ajuste a la ecuación de Gompertz (Tabla 5.15) con la concentración de amoníaco libre con el objetivo de analizar que parámetro resulta más afectado, si la velocidad de producción de metano o la producción potencial. Tal y como se observa en la Figura 5.12, tanto la producción de metano, como la velocidad de producción dependen de la concentración de amoníaco libre en el rango termofílico. La forma de la curva fue, sin embargo, muy diferente. Mientras que la producción potencial (B0) mostró una tendencia casi lineal, aunque con bajos coeficientes de regresión, aumentado conforme disminuye la concentración de amoníaco, la velocidad de producción, se mostró prácticamente constante para los tres tratamientos más concentrados, aumentando fuertemente al disminuir el amoníaco libre por debajo de 700 mg N-NH3/L. Como se observa en las Figuras 5.11 y 5.12, faltan resultados en el rango de concentración de nitrógeno amoniacal entre 300 y 600 mg N/L, por lo que no se puede establecer el valor de la constante de inhibición. En futuros trabajos, sería interesante realizar experimentos en este rango de concentraciones. R mL CH4 /g SVini/día 6 B0 mL CH4 /g SVini 200 5 150 4 3 100 2 50 B0 R 1 0 0 200 400 600 800 mL NH3 /L 1000 0 1200 Figura 5.12. Comparación de la producción potencial de metano respecto a sólidos volátiles iniciales y la velocidad de producción de metano con la concentración de amoníaco libre Según Angelidaki y Ahring (1993b) el límite a partir del cual la concentración de amoníaco libre inhibe el proceso anaerobio en el rango termofílico está sobre los 650 mg/L. Sin embargo en el experimento realizado, probablemente debido a la menor adaptación del inóculo, el límite Optimización de la digestión anaerobia de purines de cerdo 167 de inhibición estuvo por debajo de 640 mg/L, sin poder decir a que nivel exacto se encuentra. Si se compararan los resultados mesofílicos y termofílicos conjuntamente se podría llegar a deducir una aproximación a este valor. Con los valores de los parámetros de esta recta se podría establecer que la concentración de amoníaco libre necesaria para la reducción de un 25% de la producción de metano es de 427,5 mg N-NH3/L. Con esta consideración, se tendría que el rango mesofílico está inhibido también por amoníaco libre, con una reducción en la producción de metano del orden de un 14% sobre el potencial teórico. La utilización de todos los resultados para la descripción no es completamente lícita, ya que se ha demostrado que los microorganismos termofílicos son menos sensibles a la inhibición por amoníaco libre que los mesofílicos (Gallert et al., 1998). Además en ausencia de inhibidores los microorganismos termofílicos presentan mayores velocidades de crecimiento, y mayor tasa de hidrólisis, por lo que la curva se desplazaría hacia arriba. 5.5.2. Experimento 2. Codigestión de purín de cerdo con residuos de pera. Se desarrolló un estudio de viabilidad de la mezcla de purín de cerdo con residuo de elaboración de zumo de pera. Se estudió la variación en la composición inicial y final de una serie de parámetros, así como la evolución durante la digestión de la producción de gas y de la concentración de AGV. Como se verá en la exposición de resultados se ha eliminado el tratamiento 2.5, que se proponía en el apartado de materiales y métodos. Aunque se realizó el seguimiento de este tratamiento, exactamente igual que de los otros, no fue en absoluto viable, con una producción de metano nula, debido a la baja alcalinidad de este substrato, que provocó un pH tan bajo que no permitió la supervivencia de los microorganismos metanogénicos. La inclusión de resultados nulos, y con muy baja variabilidad, podría provocar errores en el estudio estadístico por lo que se decidió no considerarlos. Todos los valores expuestos en las tablas corresponden a los valores medios. Los valores finales de los parámetros físico-químicos fueron corregidos con la concentración correspondiente del blanco (digestión del inóculo con agua). Evolución de compuestos nitrogenados El residuo de pera utilizado como cosubstrato produjo una dilución de la concentración de nitrógeno amoniacal, detectándose diferencias significativas entre los diferentes niveles de mezcla para el momento inicial (Tabla 5.18). 168 Ensayos de viabilidad de codigestión en discontinuo Sin embargo, la variable nitrógeno Kjeldahl no mostró este efecto de dilución, debido básicamente a la alta relación nitrógeno orgánico/nitrógeno mineral del cosubstrato residuo de pera. La concentración de nitrógeno Kjeldahl se mantiene constante a lo largo del experimento, sin que se observen diferencias significativas entre el momento inicial y el final, ni entre las dos temperaturas (Tabla 5.18). Tabla 5.18. Concentración de compuestos nitrogenados en el experimento 2. Nitrog. Kjeldahl % (g N/Kg) Residuo Trat. de pera Inicial Final 35ºC Final 55ºC Nitrog. amoniacal (g N-NH4+/Kg) Inicial Final 35ºC Final 55ºC 2.1 0 5,26 a 5,36 a 5,35 a 3,43 E 3,83 G 4,09 I 2.2 5 5,08 a 5,14 a 5,04 a 3,21 D 3,67 F 4,16 I 2.3 12,5 5,16 a 5,15 a 5,24 a 3,01 BC 3,39 E 3,93 H 2.4 20 5,01 a 5,02 a 5,49 b 2,71 A 3,09 C 3,36 E Letras diferentes significan valores medios diferentes. (Test Lsmenas a un nivel de significación del 95%). El nitrógeno amoniacal, no sólo mostró diferencias en el inicio entre los diferentes niveles de mezcla, sino que para todos los tratamientos la concentración final fue mayor que la inicial, observándose, también, diferencias entre las dos temperaturas (Tabla 5.18), siendo mayor en el rango termofílico que en el mesofílico. El incremento de la concentración de nitrógeno amoniacal fue debida a la hidrólisis de proteínas, aumentando al final del proceso la fracción de nitrógeno mineral sobre el total (Tabla 5.19), especialmente en el rango termofílico. El efecto dilución del nitrógeno amoniacal al aumentar la proporción de cosubstrato pudo haber influido en la mayor tasa de hidrólisis obtenida en los tratamientos 2.2 y 2.3 del rango termofílico. No explica, sin embargo, lo ocurrido en el tratamiento 2.4. Evolución de la materia orgánica El principal efecto observado al añadir residuo de pera como cosubstrato fue el importante aumento observado en la concentración de materia orgánica, tanto en unidades de sólidos totales y volátiles, como de DQO (Tabla 5.20, Tabla 5.21 y Tabla 5.22). Este aumento de la materia orgánica hizo aumentar la relación C/N o sólidos volátiles/Nitrógeno total de 11,10 g Optimización de la digestión anaerobia de purines de cerdo 169 SV/g N para el control a 23 g SV/g Nk para la mezcla con un 20% de residuo de pera. Los contenidos iniciales de materia orgánica fueron diferentes para todos los tratamientos. El cosubstrato residuo de pera aportó hasta un 60% del total de los sólidos volátiles iniciales (tratamiento 2.4). Tabla 5.19. Proporción de nitrógeno amoniacal sobre nitrógeno total. Incremento sufrido durante la digestión, en el experimento 2. Trat. % Residuo Fracción N-NH4+/NK (%) Inicial Final 35ºC Final 55ºC Inc. % N-NH4 35ºC 55ºC Dif. Tª de pera 2.1 0 64,47 71,94 75,64 7,48 a 11,17 b * 2.2 5 63,00 72,10 80,66 9,10 a 17,65 c * 2.3 12,5 56,94 64,18 73,30 7,25 a 16,36 c * 2.4 20 52,36 59,80 64,02 7,44 a 11,66 b * Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significación del 95%). * o ** diferencias significativas para los valores finales entre temperaturas, a un nivel de significación del 95% o del 99%, respectivamente.. Para todos los tratamientos, la concentración de sólidos totales al final del proceso fue significativamente menor que al inicio, fruto de la descomposición de materia orgánica operada por microorganismos anaerobios, pero los porcentajes de eliminación variaron en función del tratamiento y de la temperatura. Tanto en el rango mesofílico como en el termofílico la eliminación de sólidos totales fue mayor para los tratamientos con un 5 y un 12,5% de residuo de pera, que para el control. El valor de la eliminación fue mayor en el rango termofílico que en el mesofílico. Las diferencias entre los valores de sólidos volátiles siguieron un patrón similar a los sólidos totales (Tabla 5.21), encontrando valores finales mayores para el rango mesofílico que para el termofílico. Los valores de eliminación fueron mayores también en el rango termofílico, para cada uno de los niveles de mezcla. La relación SV/ST fue mayor al final del tratamiento mesofílico que al final del termofílico. La mayor eliminación de SV y ST a 55ºC no se corresponde con los valores de eliminación de DQO obtenidos (Tabla 5.22), ni con los valores de producción de metano, que fueron mayores, excepto para el tratamiento 2.4, en el rango mesofílico. Las causas de estas discrepancias ya se relataron en apartado 5.5.1. 170 Ensayos de viabilidad de codigestión en discontinuo Tabla 5.20. Concentraciones medias y eliminación de sólidos totales en el experimento 2 Trat. % Residuo Sólidos Totales (ST) g/kg Inicial Final 35ºC Final 55ºC Eliminación ST(%) 35ºC 55ºC 8,00 A 12,81 B 73,12 b 12,54 B 21,10 C de pera 2.1 0 78,33 c 72,06 b 2.2 5 92,68 e 2.3 12,5 112,27 g 99,40 f 89,92 d 11,46 B 19,91 C 2.4 20 136,74 i 121,72 h 99,89 f 10,99 AB 26,95 D 81,51 c 68,30 a Letras diferentes significan valores medios diferentes. (Test Lsmeans a un nivel de significación del 95%). Tabla 5.21. Concentraciones medias y eliminación de sólidos volátiles en el experimento 2 % Residuo de pera Inicial 2.1 0 58,15 c 50,27 ab 2.2 5 72,73 d 2.3 12,5 2.4 20 Trat. Sólidos Volátiles (SV) g/kg Final 35ºC Final 55ºC Eliminación SV(%) 35ºC 55ºC 45,8 a 13,5% A 21,2% C 60,78 c 51,1 b 16,4% B 29,7% D 93,51 f 79,66 e 68,0 d 14,8% AB 27,3% D 118,55 h 103,07 g 80,0 e 13,1% A 32,6% E Letras diferentes significan valores medios diferentes. (Test Lsmeans a un nivel de significación del 95%). Tabla 5.22. Concentraciones medias y eliminación de DQO en el experimento 2 Trat. % Residuo de pera Inicial 2.1 0 80,04 bc 62,01 a 2.2 5 98,79 d 2.3 12,5 2.4 20 DQO g O2/kg Eliminación DQO (%) Final 35ºC Final 55ºC 35ºC 55ºC 69,73 ab 23,37 A 13,83 A 77,78 b 76,16 b 21,84 A 23,47 A 127,01 ef 93,79 cd 102,98 d 26,39 A 19,18 A 157,59 g 136,96 f 119,84 e 11,82 A 22,84 A Letras diferentes significan valores medios diferentes. (Test Lsmeans a un nivel de significación del 95%). Optimización de la digestión anaerobia de purines de cerdo 171 En los valores de eliminación de DQO no se pudieron observar diferencias significativas, debido a la gran variabilidad de los resultados obtenidos para este parámetro. En la Figura 5.13 se observan los mismos datos de forma gráfica. El residuo de pera está compuesto en gran parte por materiales lignocelulósicos, alrededor del 80% sobre materia seca. El contenido en materiales lignocelulósicos del purín utilizado fue relativamente alto, estando en torno a un 56% de los sólidos totales (76% de SV). La lignina es un tipo de compuesto altamente recalcitrante, difícil de degradar, habiéndose descrito como el principal limitador de la velocidad del proceso global al degradar residuos lignocelulósicos (Pavlostathis y Giraldo-Gómez, 1991). La mezcla de purín con residuo de pera provocó un aumento del contenido en fibra total, pero, sobre todo, un aumento en la concentración de lignina y una disminución del contenido de celulosa (Tabla 5.23). Eliminación ( ) DQO g O2 /Kg 180 160 30% 25% 140 120 20% 100 80 15% 60 10% 40 5% 20 0 0% 2.1 Inicial Eliminación 2.2 2.3 Final 35ºC Eliminación 2.4 Final 55ºC Figura 5.13. Valores de DQO inicial y final y eliminación en el experimento 2 Evolución del pH La adición de residuo de pera como cosubstrato supuso una bajada del pH de la mezcla obtenida. Esta bajada del pH tuvo una importancia clave sobre el proceso, puesto que el principal limitante de la digestión en el rango termofílico es la inhibición por amoníaco, cuya concentración depende del pH. El pH inicial fue más bajo cuanto mayor fue la proporción de cosubstrato añadido. El pH del residuo de pera diluido con agua, fue muy bajo, sobre 3,5, lo que imposibilitó el desarrollo del proceso biológico con este substrato. 172 Ensayos de viabilidad de codigestión en discontinuo Tabla 5.23. Contenido en fibra y en el substrato inicial del experimento 2 (% sobre sólidos volátiles). Trat. % Residuo de pera Lignina Celulosa Hemicelulosa 2.1 0 22,91% 17,62% 35,65% 2.2 5 26,54% 15,95% 34,84% 2.3 12,5 30,06% 14,31% 34,03% 2.4 20 32,14% 13,29% 33,53% La alcalinidad total es menor cuanto mayor es la proporción de cosubstrato, pero además aumenta la relación de alcalinidad, que mide la relación entre la alcalinidad debida a los ácidos grasos y la debida al bicarbonato (Tabla 5.24). El alto contenido en ácidos grasos volátiles desde el inicio se debe al alto contenido de azúcares de este tipo de substratos, que hace que la fermentación comience muy rápidamente. Tabla 5.24. Alcalinidad total, alcalinidad al bicarbonato y relación de alcalinidad, en el experimento 2. Tratamiento % Residuo Alc. Total Alc. Parcial de pera (g CaCO3/L) (g CaCO3/L) Alc. Int. (g AcH/L) RA 2.1 0 10,55 7,5 3,66 0,29 2.2 5 9,75 6,7 3,66 0,31 2.3 12,5 8,725 5,775 3,54 0,34 2.4 20 7,575 4,75 3,39 0,37 El bajo pH inicial observado (Tabla 5.25), junto con el efecto dilución de la adición de residuo de pera, supuso una bajada importante de la concentración inicial de amoníaco libre respecto al purín sin mezclar (Tabla 5.26), y, por tanto, una mejora en el proceso de digestión anaerobia. Sin embargo, el pH final, especialmente en el rango termofílico, fue bastante más alto que el inicial, lo que podría haber provocado una inhibición posterior del proceso. El aumento del pH al final de la digestión, se atribuye al consumo de ácidos grasos por parte de los microorganismos acetogénicos y metanogénicos. En general, el aumento del pH fue mayor en el rango Optimización de la digestión anaerobia de purines de cerdo 173 termofílico que en el mesofílico. En el caso del tratamiento 2.4 en mesofílico el aumento del pH apenas llegó a las dos décimas, indicando la pobre recuperación operada en este tratamiento (ver producción de gas y evolución de ácidos grasos). Tabla 5.25. Valores medios de pH al inicio y al final de la digestión en el experimento 2. Trat. % Residuo de pera Inicial Final-35ºC Final-55ºC 2.1 0 8,06 7,70 7,99 2.2 5 7,55 7,69 7,92 2.3 12,5 7,25 7,58 7,83 2.4 20 6,88 7,11 7,87 La gran influencia de la variación del pH sobre la concentración de amoníaco libre queda de manifiesto en la Tabla 5.26, donde se muestran los valores de amoníaco libre calculados al inicio y al final de la digestión. La concentración de NH3 pasa de niveles muy bajos a superar los valores inhibidores propuestos en la bibliografía. Tabla 5.26. Concentración de amoníaco libre en el experimento 2. % Residuo de pera Inicial Final Media Inicial Final Media 2.1 0 404,00 207,59 305,80 1094,29 1118,18 1106,.23 2.2 5 126,04 197,62 161,83 402,47 1024,94 713.70 2.3 12,5 60,49 141,89 101,19 202,00 837,37 519.69 2.4 20 23,23 45,30 34,27 79,79 770,01 424.90 Trat. 35ºC 55ºC Evolución de la producción de biogás. La evolución de la producción acumulada de CH4 y CO2 siguió una tendencia diferente en función de la cantidad de co-substrato añadido y de la temperatura del ensayo. En general la producción de metano aumentó, sobre todo en el rango termofílico, pero en algún tratamiento resultó un fracaso con producciones de metano muy bajas (Tabla 5.27). 174 Ensayos de viabilidad de codigestión en discontinuo Tabla 5.27. Valores medios de producción acumulada de gas (M) al final del proceso, para las variables biogás, CH4 y CO2, en el experimento 2. M( mL Biogás) Trat. 35º 55º 2.1 658,7 bc 437,0 a 2.2 M (mL de CH4) 35º M (mL de CO2 ) 55º 35º 55º ** 443,1 BC 195,2 A ** 215,5 A 241,8 A - 852,8 de 781,3 cd - * 314,9 B 357,2 B - 2.3 1372,7 g 984,8 ef ** 831,3 E 521,5 CD ** 541,3 E 463,4 CD ** 2.4 625,2 b 1058,5 f ** 195,7 A 429,5 C 518,0 DE ** 537,9 CD 424,1 B 540,4 D ** Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significaión del 95%). * o ** diferencias significativas para los valores finales entre las dos temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. La digestión del residuo de pera sin adición de purín no fue viable debido a la baja alcalinidad y el bajo pH del residuo. Para su digestión se debería modificar artificialmente el pH. Al comparar los índices de producción de metano sobre sólidos volátiles las diferencias entre los tratamientos se atenuaron (Tabla 5.28), ya que el principal efecto del aporte de cosubstrato sobre las características del substrato fue el aumento de materia orgánica (Tabla 5.22). Rango termofílico En el régimen termofílico se observó un aumento considerable en la producción acumulada de metano al añadir un 5% de cosubstrato, tal y como se observa en la Figura 5.14 y en la Tabla 5.27, pasando de una producción total de 195 mL de metano para el Purín 100% a 424.1 mL. La producción al añadir un 12,5% de cosubstrato aumentó también aunque la diferencia con el tratamiento 2.2 fue menor y no hubo diferencias significativas entre este tratamiento y el tratamiento 2.4 (20% de residuo de pera). Los índices de producción de metano respecto a SV iniciales fueron, para todos los tratamientos con adición de residuo de pera, significativamente mayores que para el control, aunque al aumentar la proporción de cosubstrato tendieron a disminuir. Aunque los índices fueron mayores que para el control, estuvieron por debajo del índice obtenido para el purín diluido con agua al 20% (1.4), indicando que la metanogénesis pudo verse afectada por la presencia de algún tóxico. Optimización de la digestión anaerobia de purines de cerdo 175 Tabla 5.28. Valores medios del índice de producción acumulada de gas respecto a sólidos volátiles iniciales (B) al final del proceso, para las variables biogás, metano y CO2, en el experimento 2. B( mL Biogás/gSV) B (mL de CH4/g SV) Trat. 35º 55º 35º 55º 2.1 223,2 d 148,1 b ** 150,1 E 66,1 B 2.2 2 32,8 d 213,3 cd - 146,8 E 115,8 D 2.3 290,2 e 208,2 cd ** 175,7 F 110,2 CD ** 2.4 107,83 a 182,6 c 33,7 A ** 93,2 C B (mL de CO2/g SV) ** 35º 55º 73,0 A 81,9 AB - 97,9 C - 114,4 D 97,5 C * 74,1 A 89,4 BC * ** 86,0 ABC ** Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significaión del 95%). * o ** diferencias significativas para los valores finales entre las dos temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. mL de CH4 900 35º 2.1 800 35º 2.2 700 35º 2.3 600 35º 2.4 500 55º 2.1 400 55º 2.2 300 55º 2.3 200 55º 2.4 100 0 0 10 20 30 40 Día 50 60 70 80 Figura 5.14. Producción acumulada de CH4 en el experimento 2 En el régimen termofílico se observaron importantes diferencias entre los tres tratamientos de codigestión con residuo de pera. Los tratamientos 2.3 y 2.4 siguieron prácticamente la misma evolución, diferente del 2.2, con una pendiente mayor en el período inicial, estancándose a partir del día 20. Los valores de producción de metano no se pudieron diferenciar ni el día 6, ni el 15 ni al final del experimento. Al obtener un valor de producción acumulada parecido, el índice de producción sobre sólidos volátiles fue muy diferente, debido a la gran diferencia en el contenido inicial de materia orgánica de los dos tratamientos. El tratamiento 2.2 mostró una menor pendiente en el 176 Ensayos de viabilidad de codigestión en discontinuo inicio reactivándose después. En cualquier caso el comportamiento en cuanto a producción acumulada de metano mejoró ostensiblemente en todos los tratamientos respecto al obtenido para el tratamiento control (2.1 Purín 100%). mL de CO2 600 35º 2.1 500 35º 2.2 400 35º 2.3 35º 2.4 300 55º 2.1 55º 2.2 200 55º 2.3 100 55º 2.4 0 0 10 20 30 40 Día 50 60 70 80 Figura 5.15. Evolución de la producción acumulada de CO2, en el experimento 2 Rango mesofílico En el rango mesofílico la producción acumulada de metano para un 5 y un 12,5% de cosubstrato superó los valores obtenidos en termofílico y los obtenidos para el control. La máxima producción de metano se obtuvo para la mezcla con un 12,5% de cosubstrato, llegando a alcanzar 831 mL de metano. Sin embargo, el tratamiento con un 20% de residuo de pera obtuvo un valor muy bajo de producción de metano (Tabla 5.27). En el régimen mesofílico el tratamiento con un 12.5% de pera obtuvo un índice de producción de metano respecto a sólidos volátiles superior al observado para purines, sin embargo, el tratamiento con un 5% de pera no mejoró dicho índice (Tabla 5.28 y Figura 5.16). El tratamiento 2.3 (pera 12.5%) en mesofílico tuvo un comportamiento diferente del resto de tratamientos tanto a 35 como a 55ºC, mostrando un desfase inicial en la producción de metano, que se prolongó hasta el día 18, momento en que comenzó la fase de producción exponencial. El tratamiento 2.4 en mesofílico estuvo inhibido durante todo el período, con momentos en que la producción diaria fue incluso menor que la obtenida para el blanco Optimización de la digestión anaerobia de purines de cerdo 177 (inóculo más agua), lo que hizo que la curva de producción acumulada bajara. Al final del experimento, a partir del día 60 de digestión se activó ligeramente la producción de metano, aunque con una pendiente muy pequeña. Al estudiar la evolución de la producción acumulada de metano (Figura 5.14), se observa que el tratamiento 2.2 (5% de residuo de pera) se comportó de forma muy similar hasta el día 15 a las dos temperaturas probadas, pero a partir de este punto el tratamiento mesofílico produjo mayores cantidades de gas. De la misma forma que ocurría en el experimento anterior, se observa una mayor producción de CO2 en el régimen termofílico que en el mesofílico (Figura 5.15), es decir el gas obtenido tuvo una menor riqueza en metano. En el régimen mesofílico el tratamiento 2.4 (20% de pera) obtuvo una gran producción de CO2 en los primeros días de digestión, estando en el día 4 en 230 mL de CO2 acumulado. A partir de este momento sufrió una parada en la producción de CO2, mostrando tan sólo una producción residual, probablemente debido a la inhibición de la fase hidrolítica/acidogénica por la alta concentración de ácidos grasos. La máxima producción de CO2 de todos los tratamientos fue para el 2.3 del régimen mesofílico, similar a lo ocurrido con el metano. mL de CH 4/gSVini mL de CH 4/gSVini 200 200 2.1-55ºC 180 160 180 160 2.3-55ºC 140 2.2-55ºC 140 120 2.3-55ºC 120 100 2.2-55ºC 100 80 80 60 60 40 40 20 20 0 2.1-55ºC 2.4-55ºC 0 0 20 2.1-35ºC 2.3-35ºC 2.2-35ºC 2.4-35ºC 40 60 1.1 sim-35º Ajuste 2.3 sim-35º Gompertz 2.2 sim-35º 2.4 sim-35º 80 Día 0 20 2.1-55ºC 2.2-55ºC 2.3-55ºC 2.4-55ºC 40 60 1.1 sim-55º Ajuste 2.2 sim-55º Gompertz 2.3 sim-55º 2.4 sim-55º 80 Día Figura 5.16. Comparación de los datos experimentales media de las cinco repeticiones (puntos) y la curva de la simulación del modelo Gompertz, en el experimento 2, para el índice producción de metano respecto a sólidos volátiles iniciales 178 Ensayos de viabilidad de codigestión en discontinuo Análisis de parámetros de velocidad, producción potencial y duración de la fase lag. La ecuación de Gompertz (ecuación 5.5), en general se ajustó bien a los datos experimentales excepto en aquel tratamiento que resultó fuertemente inhibido (tratamiento 2.4-35ºC), para el cual no se pudo realizar el ajuste. Los coeficientes de regresión obtenidos estuvieron en todos los casos por encima de 0,93 y para la gran mayoría de las repeticiones entre 0,96 y 0,99. En la Figura 5.16 se muestran los valores del índice de producción de metano respecto a sólidos volátiles y la curva del modelo de Gompertz ajustado, para cada tratamiento. Los valores de los parámetros de las ecuaciones 5.5 y 5.6 se estimaron por mínimos cuadrados para cada una de las repeticiones desarrolladas. Una vez obtenidos los parámetros se realizó el análisis de la varianza, cuyos resultados se muestran en la Tabla 5.29. Tabla 5.29. Parámetros estimados mediante la ecuación de Gompertz en el experimento 2. M0 Tratamiento mL CH4 K ml CH4/d O d B0 mL CH 4 g SV R mL CH 4 g SV·d R’ d-1 2.1 35ºC 428,83 b 11,08 b 4,04 bc 144,98 d 3,76 bcd 0,07 a 2.2 35ºC 493,44 b 12,92 b 1,60 ab 134,57 cd 3,54 bcd 0,07 a 2.3 35ºC 846,64 c 21,46 c 12,64 d 178,72 e 4,55 d 0,07 a 2.4 35ºC - - - - - - 2.1 55ºC 207,20 a 4,21 a 6,35 c 70,21 a 1,43 a 0,06 a 2.2 55ºC 419,58 b 10,68 b 2,08 b 114,56 bc 2,91 b 0,07 a 2.3 55ºC 475,98 b 18,69 c -0,56 a 100,42 ab 3,96 cd 0,11 b 2.4 55ºC 398,14 b 28,13 d 1,93 b 85,57 a 3,22 bc 0,10 b Letras diferentes significan valores medios diferentes. (Test Lsmeans a un nivel de significación del 95%). Análisis estadístico realizado por separado para cada variable (cada columna). La producción potencial acumulada de metano (M0) sólo fue diferente para el tratamiento 2.3-35ºC y el 2.1–55ºC (control). El resto de tratamientos, excepto el 2.4-35ºC, que debido a los bajos coeficientes de regresión obtenidos no se incluyó en el análisis, tuvieron producciones acumuladas potenciales que no se pudieron diferenciar. Optimización de la digestión anaerobia de purines de cerdo 179 El valor máximo de velocidad de producción de metano en el rango mesofílico (mL de CH4/día), se obtuvo para el tratamiento 2.3 a 35ºC. Sin embargo no se pudo diferenciar del 2.3 a 55ºC y fue menor que el obtenido para el tratamiento 2.4 en el rango termofílico. Este parámetro fue el único de todos los analizados en que la interacción entre los dos efectos principales (temperatura y tratamiento) no fue significativa, por lo que se analizaron los efectos principales. La velocidad de producción de metano (K) es mayor cuanto mayor es el contenido de cosubstrato aportado, mostrando diferencias significativas a un nivel de significación del 5% entre todos los tratamientos, sin que se observaran diferencias entre temperaturas (salvo para el control). En la duración del desfase o fase lag (O), la interacción entre temperatura y nivel de mezcla, sí fue significativa, observándose que el tratamiento 2.3 a 35ºC fue el que mayor desfase sufrió, estimándose en 12,64 días. Sin embargo en el rango termofílico la duración del desfase fue pequeña, yendo de 0 a 2 días, del mismo orden de magnitud que la del 2.2 a 35ºC o que la del control a 35ºC. En general, se puede decir que en el rango termofílico la duración del desfase disminuye al añadir el cosubstrato. Sin embargo el alto valor obtenido para el tratamiento 2.3 en mesofílico indica un proceso de inhibición inicial o de sobrecarga, que en este caso fue posible superar. Las diferencias fueron menores en la velocidad de producción de metano (R) en unidades de mL de CH4/g SV*día, que en el parámetro K. Así, en el rango mesofílico no se pudieron diferenciar ninguno de los tres tratamientos considerados. En el termofílico el control mostró una velocidad menor que el resto de los tratamientos, siendo el máximo el 2.3, aunque del mismo orden de magnitud que el 2.4 y los tratamientos en mesofílico. El parámetro R’, velocidad específica de producción de metano (día–1), resultado de dividir R por la producción potencial de metano respecto a los sólidos volátiles fue estadísticamente igual para todos los tratamientos excepto para los dos tratamientos con mayor contenido de cosubstrato en el rango termofílico. Índice de metanización, acidificación y biodegradabilidad Los índices de metanización y acidificación se calcularon según las ecuaciones 5.8 y 5.9, mostrándose los valores obtenidos en la Tabla 5.30. Hubo diferencias significativas entre temperaturas en el índice de metanización, para todos los tratamientos, siendo para todos mayor a 35ºC, excepto en el tratamiento 2.4 (20% de residuo de pera). En el rango mesofílico el índice de metanización aumentó sólo para el 180 Ensayos de viabilidad de codigestión en discontinuo tratamiento 2.3 (12,5% de residuo de pera), sin que se detectaran diferencias entre el control y el 2.2 (5% de pera). El 2.4 (20%), resultó fuertemente inhibido, lo que hizo que obtuviera el índice más bajo de todo el experimento. Dentro del rango termofílico, el índice de metanización mejoró respecto al control, con la adición de residuo de pera, aunque disminuyó al aumentar la proporción de residuo de pera utilizada. Las menores diferencias entre el índice de acidificación y el de metanización, dentro del rango termofílico, se produjeron para el tratamiento con más proporción de residuo de pera, ya que fue el que menor concentración de AGV mostró al final, indicando por tanto que fue el que menor grado de inhibición de la metanogénesis tuvo. Sin embargo, el índice de acidificación fue menor que el obtenido para los tratamientos 2.2 y 2.3 a la misma temperatura (Tabla 5.30), y del mismo orden de magnitud que su homólogo a 35ºC, que, sin embargo, produjo muy poco metano. El descenso del índice de acidificación indica la menor biodegradabilidad del residuo de pera, debido probablemente al alto contenido en lignina, compuesto altamente recalcitrante. Sin embargo el tratamiento 2.3 mostró unos mejores índices tanto de metanización como de acidificación. Este tratamiento, a 35ºC, sufrió una importante fase lag, con una evolución de AGV inicial similar al tratamiento 2.4. Esta acumulación, extraordinariamente fuerte de AGV pudo hacer bajar el pH de forma muy importante, y quizá podría haber mejorado por ello la hidrólisis, lo que habría hecho aumentar el índice de BD, sobre todo frente al tratamiento a 55ºC que debió aumentar rápidamente su pH, debido a la rápida eliminación de AGV. Es probable, que si se hubiera continuado con el experimento del tratamiento 2.4 a 35ºC, se hubiera observado algo similar. El pH final del tratamiento 2.4 a 35ºC, fue bastante más bajo que para el resto de tratamientos. El índice de acidificación no mostró diferencias significativas entre temperaturas para un 5% de residuo de pera, aunque el de metanización fue menor en el rango termofílico, indicando la inhibición de la fase metanogénica, de forma similar, aunque menos acentuada, que en el tratamiento control (2.1), y probablemente provocada, también como en el control, por el amoníaco libre. Optimización de la digestión anaerobia de purines de cerdo 181 Tabla 5.30. Indice de metanización, acidificación y biodegradabilidad en el experimento 2. Trat. Metanización (%M) Acidificación (%A) 35ºC 35ºC 55ºC Biodegradabilidad (%BD) 55ºC 35ºC 55ºC 2.1 29,44 e 12,96 b ** 29,54 C 19,68 A ** 36,62 C 23,94 A ** 2.2 28,97 e 22,86 d ** 29,03 BC 28,08 BC - 35,90 BC 34,50 BC - 2.3 34,82 f 21,82 cd ** 34,92 D 24,82 B ** 43,56 D 30,64 B ** 2.4 6,60 a 18,26 c ** 17,64 A 19,96 A 21,44 A 24,68 A - - Letras diferentes significan valores medios diferentes. (Test Lsmeans a un nivel de significación del 95%). Análisis estadístico realizado por separado para cada uno de los días y cada variable. * o ** diferencias significativas para los valores finales entre temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. Evolución de ácidos grasos volátiles. La evolución de la concentración de ácidos grasos volátiles total siguió tendencias diferentes en función de la temperatura y de la proporción de cosubstrato. Inversamente a lo ocurrido en el experimento anterior (dilución de purín), las mayores acumulaciones de AGV se obtuvieron en el rango mesofílico, con unos valores máximos muy por encima de los máximos del rango termofílico, llegando a alcanzar valores extremadamente altos en el caso de los tratamientos 2.3 (378.5 mM) y 2.4 (526.7 mM) (Figura 5.17 y Tabla 5.31). AGVt (mM )-55ºC AGVt (mM )-35ºC 700 700 600 600 500 500 400 400 300 300 200 200 100 100 0 0 0 10 20 30 40 Día 50 60 55º 2.1 35º 2.1 70 80 55º 2.2 35º 2.2 0 10 55º 2.3 35º 2.3 20 30 40 50 Día 60 70 55º 2.4 35º 2.4 Figura 5.17. Evolución de los Ácidos Grasos Volátiles totales en el experimento 2 80 182 Ensayos de viabilidad de codigestión en discontinuo Tabla 5.31. Valores de concentración de Ácidos grasos Volátiles (mM) obtenidos en diferentes momentos de la digestión, en el experimento 2. Concentración media de ácidos grasos volátiles totales (mM) Día 6 Día 13 Trat. 35º C 55ºC 2.1 81,5 a 96,76 a 35º C 55ºC 64,31 A 35º C 55ºC 153,17 B * 1,07 A 62,23 B ** 2.2 164,08 c 133,14 bc - 142,63 B 163,45 B - 1,38 A 43,44 AB - 2.3 271,21 d 1,77 A 31,41 AB - 2.4 286,51 d 100,17 a ** 526,69 D 34,80 A ** 170,34 C 21,71 AB ** 83,25 a - Final ** 378,49 C 50,18 A ** Concentración media de acético (mM) Día 6 Día 13 Trat. 35º C 55ºC 35º C 2.1 63,03 A 79,9 a 2.2 126,93 B 111,68 ab - 2.3 212,82 C 2.4 207,12 C Final 55ºC 35º C 55ºC 1,05 A 46,28 B * 134,98 C ** 1,34 A 24,1 AB - 51,81 a ** 266,16 D 24,19 AB ** 1,74 A 7,16 AB - 57,39 a ** 332,45 E ** 92,55 C 3,51 A ** - 26,27 AB 133,12 C ** 60,27 B 11,3 A Concentración media de propiónico (mM) Día 6 Día 13 35º C 55ºC 29,36 B 7,32 A Final Trat. 35º C 55ºC 2.1 10,26 B 5,82 a * 2.2 25,96 c 8,74 ab ** 66,70 C 12,11 A ** 0,02 A 10,83 BC - 2.3 29,20 c 9,48 b ** 68,00 C 13,2 A 0,03 A 15,93 C ** 2.4 28,45 d 10,8 b ** 106,36 D 17,14 AB ** 34,35 D 11,88 BC ** ** ** 35º C 55ºC 0A 6,05 AB - Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significación del 95%). Análisis estadístico realizado por separado para cada uno de los días y cada variable. * o ** diferencias significativas para los valores finales entre las dos temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. Rango mesofílico El tratamiento 2.2, 5% de residuo de pera, a 35ºC sufrió, desde el primer momento, un fuerte incremento en la concentración de ácidos, superando claramente en el día 6 la concentración obtenida para el tratamiento control, Optimización de la digestión anaerobia de purines de cerdo 183 siendo este día el que alcanzó la concentración máxima, 164 mM (Tabla 5.31). La concentración de AGV, sin embargo, se mantuvo a un nivel bastante alto hasta el día 14, momento a partir del cual disminuyó rápidamente. La evolución de la concentración de acético del tratamiento 2.2 siguió una tendencia similar a la observada para la concentración de AGV, con el máximo el día 6, en 127 mM. Sin embargo, el acético comenzó a disminuir más rápidamente que los AGV totales, debido a que el ácido propiónico comenzó a acumularse a partir del día 8 (Figura 5.18), llegando a un máximo de 66.7 mM el día 13 (Tabla 5.31). A partir de este momento el propiónico comenzó a descender, aunque más lentamente que en el experimento 1, manteniéndose a un nivel relativamente alto durante bastante tiempo (por encima de 20 mM el día 37). Al final del proceso la concentración de ácidos fue muy baja, pudiéndose detectar sólo el acético. Acético (mM)-35ºC 400 350 300 250 200 150 100 50 0 0 10 20 Acético (mM )-55ºC 400 350 300 250 200 150 100 50 0 30 40 50 60 70 0 80 10 20 30 40 50 60 70 80 30 40 Día 50 60 70 80 Propiónico (mM )-55ºC Propiónico (mM )-35ºC 120 120 100 100 80 80 60 60 40 40 20 20 0 0 0 10 20 30 40 Día 50 60 55º 2.1 35º 2.1 70 80 55º 2.2 35º 2.2 0 10 55º 2.3 35º 2.3 20 55º 2.4 35º 2.4 Figura 5.18. Evolución de la concentración de acético y propiónico, en el experimento 2 Las formas n del butírico y el valérico sufrieron una ligera acumulación 184 Ensayos de viabilidad de codigestión en discontinuo coincidiendo con la máxima acumulación de acético, aunque muy por debajo de los valores obtenidos para los tratamientos con mayor proporción de residuo de pera. Sin embargo las formas iso de dichos compuestos alcanzaron niveles relativamente altos, y del mismo orden de magnitud que para los tratamientos con mayor proporción de residuo de pera, diferenciándose sólo a partir del día 37 (Figura 5.19). Aunque el valor máximo de concentración de acético fue mayor en el tratamiento 2.2 que para el control, la evolución de este parámetro fue relativamente parecida entre los diferentes tratamientos, mostrando una rápida eliminación, lo que hace suponer que no hubo problemas de inhibición importantes de la fase metanogénica acetoclástica. La acumulación inicial se puede explicar por la diferente velocidad de crecimiento de las poblaciones acidogénica y metanogénica; las diferencias con respecto al tratamiento control vendrían determinadas por la mayor concentración de carbohidratos solubles en el cosubstrato, que hace que se acumulen ácidos grasos volátiles, fruto de la fermentación de los carbohidratos. La acumulación de propiónico, especialmente importante a partir del día 8, podría, en teoría, haber estado causada por la propia acumulación de acético, ya que este compuesto se ha descrito como un inhibidor de la acetogénesis (Fukuzaki et al., 1990). Sin embargo, en este caso la concentración de acético mostraba una clara tendencia a disminuir en este momento. Por otra parte, la acumulación de propiónico, junto con la acumulación de las formas iso de butírico y valérico, se han descrito como síntomas del aumento de la presión parcial de hidrógeno (pH2) en el reactor (Hill y Cobb, 1993), que parece que es lo que ocurrió en el tratamiento 2.2. La concentración de propiónico y de las formas iso de butírico y valérico se mantuvieron a un nivel bastante alto durante muchos días, indicando que el problema persistía. Al observar la gráfica de producción acumulada media de metano se observa que a partir del día 55 hubo un cambio de pendiente, indicando la reactivación del proceso; este cambio de pendiente coincidió con la disminución de la concentración de propiónico (Figura 5.17 y Figura 5.18), y por tanto, con el correcto funcionamiento de la metanogénesis hidrogenotrófica. En el tratamiento 2.3 se observó una muy fuerte acumulación inicial de AGV hasta el día 13, disminuyendo acusadamente a partir de este momento, llegando a un nivel del mismo orden de magnitud que el tratamiento 2.2, aunque por encima del control durante bastantes días. La concentración al final del experimento no se pudo diferenciar estadísticamente del control y del tratamiento 2.2. El aumento inicial de AGV fue básicamente debido a la Optimización de la digestión anaerobia de purines de cerdo 185 acumulación de acético, que en el día 13, se situó en 266 mM (17 g Ac/L). A partir de este momento comenzó a disminuir de forma muy acusada, y el día 19 ya estaba en 109 mM y el 26 en 15.6 mM. El ácido propiónico se comportó de forma casi idéntica al tratamiento anterior, con una fuerte acumulación a partir del día 8 y hasta el 14, disminuyendo después de forma pronunciada, aunque volvió a aumentar después, observándose un nuevo máximo relativo el día 37. Al final del proceso, la concentración de propiónico fue despreciable. Las formas n, tanto del butírico como del valérico mostraron una tendencia similar a la del acético, llegando a niveles de concentración bastante altos, aunque por debajo de los obtenidos en el tratamiento 2.4. Las formas iso del butírico y el valérico, mostraron una tendencia similar a la del propiónico, pero el nivel de concentración fue mucho menor que el obtenido para las formas n. La producción de metano para este tratamiento mostró una fuerte fase lag, estimada en 12,64 días (Tabla 5.29). Esta fase lag coincidió con la fuerte acumulación de acético observada (Figura 5.18). La duración de la fase lag se ha relacionado en la bibliografía con la relación Substrato/Microorganismos (Hashimoto et al., 1989). Dado el alto contenido de materia orgánica del residuo de pera, y que la cantidad de inóculo añadido fue la misma que para el resto de tratamientos, la relación S/M aumentó con el contenido de cosubstrato. Puesto que los microorganismos acidogénicos son más rápidos y menos sensibles que los metanogénicos una alta relación S/M favorece a los primeros, y podría, como en este caso perjudicar a los segundos. La gran acumulación de acético producida podría haber sido causa de la inhibición del grupo metanogénico, ya que las altas concentraciones de acético han sido descritas como capaces de inhibir los microorganismos que lo degradan, que se conoce como inhibición por el substrato (Andrews y Graef, 1971; Hill, 1982; Angelidaki et al., 1993, etc.). A pesar de esta larga fase lag, la metanogénesis pudo funcionar correctamente una vez superada, como demuestra la rápida eliminación de acético y la gran producción de metano obtenida. El mecanismo de reactivación habría pasado por el descenso de la concentración de acético, debido a la inhibición de la acetogénesis, y la consiguiente acumulación de propiónico y otros AGV (formas n de butírico y valérico), observada fundamentalmente a partir del día 8 y con un máximo el día 14, igual que el acético. La fuerte acumulación de acético puede haber inhibido la acetogénesis, resultado de la acción conjunta de la alta concentración y la presumible bajada del pH (se llegó a una concentración de unos 260 mM de AGV), puesto que la forma de acético descrita como 186 Ensayos de viabilidad de codigestión en discontinuo inhibidora es la forma no ionizada (Fukuzaki et al., 1990), cuya concentración aumenta conforme baja el pH. Isobutírico (mM)-35ºC 5 Isobutírico (mM)-55ºC 5 4 4 3 3 2 2 1 1 0 0 0 10 20 30 40 50 60 70 80 N-butírico (mM)-35ºC 80 0 10 20 30 40 50 60 70 80 N-butírico (mM)-55ºC 80 60 60 40 40 20 20 0 0 0 10 20 30 40 Iso-valérico (mM)-35ºC 7 6 5 4 3 2 1 0 0 10 20 30 40 50 50 60 60 70 70 40 50 60 70 80 80 Iso-valérico (mM)-55ºC 7 6 5 4 3 2 1 0 0 10 20 30 40 50 60 70 80 80 N-valérico (mM)-55ºC 30 25 20 15 10 5 0 0 10 20 30 50 60 70 80 80 N-valérico (mM)-35ºC 30 25 20 15 10 5 0 0 10 20 30 40 50 60 70 0 10 Día 20 30 40 Día 55º 2.1 35º 2.1 55º 2.2 35º 2.2 55º 2.3 35º 2.3 55º 2.4 35º 2.4 Figura 5.19. Evolución de los isómeros de butírico y valérico en el experimento 2 Optimización de la digestión anaerobia de purines de cerdo 187 Aunque las formas iso del butírico y valérico (Figura 5.19) sufrieron una acumulación coincidiendo con la acumulación de propiónico, fue de mucha menor cuantía que la sufrida por las formas n, que se ha relacionado con la inhibición de la acetogénesis por acético, y no por hidrógeno. A medida que la metanogénesis comenzó a funcionar adecuadamente y el acético disminuyó, las formas n del butírico y valérico disminuyeron también, comenzando a aumentar la fracción iso sobre el total (suma de n + iso), indicando que la acetogénesis continuaba estando limitada, pero ahora por la presión parcial de hidrógeno (Hill y Cobb, 1993). La producción de metano se reactivó alrededor del día 20 con una gran producción diaria hasta el día 30, que coincidió con un fuerte descenso en la concentración de ácidos. A partir del día 30, sin embargo, la producción diaria de metano se redujo, continuando con un crecimiento constante aunque menor hasta casi el final de la digestión, debido probablemente a que a partir de ese momento el principal limitante de la velocidad del proceso debió ser la hidrólisis de los compuestos orgánicos complejos. El tratamiento 2.4 (20% de residuo de pera) a 35ºC, produjo muy poca cantidad de metano. La concentración de AGV llegó a valores máximos del orden de 527 mM (31,6 gAc/L), indicando que la metanogénesis estuvo fuertemente inhibida. Inicialmente siguió un comportamiento similar al 2.3, pero no fue capaz de reactivarse, al menos durante la duración del experimento. Aunque a partir del día 13 disminuyó algo la concentración de AGV, esta estuvo siempre por encima de los 150 mM, nivel mucho más alto que los citados en la bibliografía como inhibidores del proceso anaerobio (Ahring et al., 1995). El acético fue el ácido que mayor concentración alcanzó, llegando a valores de 20 g Ac/L. El valor máximo se detectó el día 13, momento a partir del cual hubo un importante descenso. El propiónico también se acumuló los primeros días, llegando al máximo el día 13, descendiendo después acusadamente, aunque a partir del día 20 la concentración se mantuvo aproximadamente constante, o con una ligera tendencia a aumentar. Las concentraciones de los isómeros n e iso del butírico y el valérico, mostraron tendencias similares a la del acético y el propiónico, respectivamente. La relación S/M en este tratamiento fue mucho mayor que en el anterior, y eso debió provocar de hecho una fase lag que se prolongó durante todo el experimento. La concentración de AGV aumentó mucho al inicio del proceso alcanzando, como en los otros tratamientos, un máximo alrededor del día 14. La acumulación fue tan grande que se produjo la inhibición de la acetogénesis y la consiguiente bajada en la concentración, pero al contrario que en el tratamiento 2.3, el consumo de acético no se 188 Ensayos de viabilidad de codigestión en discontinuo reactivó, continuando a muy alta concentración, de forma que el proceso continuó estando inhibido hasta el final. Una posible razón de la mayor inhibición en este tratamiento pudo ser el pH que desde el principio estuvo a un nivel relativamente bajo (por debajo de 7). El pH bajo puede potenciar el efecto inhibidor del acético sobre la acetogénesis y sobre la metanogénesis (Fukuzaki et al., 1990), lo que puede explicar que, a pesar de mostrar una concentración de acético del mismo orden de magnitud que el tratamiento 2.3 (Tabla 5.31), el sistema no fuera capaz de reactivarse. La tenue disminución en la concentración de acético al final del proceso coincidió con una ligera tendencia a aumentar la producción de metano, indicando la posible reactivación de la metanogénesis. Puesto que el ensayo se cortó en este punto, esta “reactivación” no es más que una hipótesis sin contrastar. Rango termofílico En el régimen termofílico el comportamiento del tratamiento 2.2 fue bastante similar al homólogo a 35ºC, con un fuerte incremento inicial de AGV (el día 1 ya tenía un nivel de 106 mM) y una más lenta acumulación hasta el día 13, llegando a alcanzar los 163 mM, momento a partir del cual comenzó a disminuir. El mismo comportamiento que los AGV totales siguió el acético, con un máximo el día 13 de 135 mM (8,1 g Ac/L), disminuyendo rápidamente inicialmente, manteniéndose después en un nivel relativamente alto (24 mM) hasta el final del experimento, indicando un estado de inhibición relativa de la metanogénesis. La concentración de propiónico se mantuvo aproximadamente constante durante todo el tratamiento, con una tenue acumulación alrededor del día 13. La forma n-butírico mostró una tendencia similar a la del acético, aunque con un nivel de concentración muy bajo; el n-valérico, sin embargo, mantuvo la concentración a un nivel muy bajo durante todo el experimento (Figura 5.19). Las formas iso, tanto del butírico como del valérico mostraron una concentración creciente al principio y aproximadamente constante hasta el final del experimento, siendo del mismo orden que los valores máximos del mesofílico y similares para todos los tratamientos en termofílico. La disminución de la concentración de acético ocurrió antes en el tratamiento 2.2 que en el control, indicando que la fase exponencial de producción de metano comienza antes. De hecho se observó una disminución de la duración de la fase lag sobre el control. La producción acumulada de metano, así como la velocidad de producción de metano (K) fueron mayores en este tratamiento (Tabla 5.27 y Tabla 5.29). La Optimización de la digestión anaerobia de purines de cerdo 189 acumulación de acético al inicio del experimento se debió, como ya se ha comentado, a la mayor rapidez de los microorganismos acidogénicos por tratarse de un substrato rico en azúcares, lo que provocó un rápido aumento de AGV (Figura 5.16). A partir del día 13 la concentración de AGV disminuyó de forma bastante rápida, manteniéndose durante el resto del proceso por debajo de la concentración de AGV del control, a pesar de que al final no se detectaron diferencias. Esta menor acumulación de ácidos coincidió con una mayor producción de metano En los tratamientos 2.3 y 2.4 se observó también una acumulación inicial de AGV inicial, pero de muy corta duración, y el valor máximo fue mucho más bajo que los mismos tratamientos a 35ºC e incluso que el máximo del 2.2 a 55ºC. A partir del día 3 comenzó a disminuir la concentración hasta el día 19, momento a partir del cual se mantuvo aproximadamente constante a un nivel de 25-40 mM, por debajo del control, hasta el final del experimento. La concentración de propiónico siguió una evolución similar al tratamiento 2.1-55ºC (control) y al 2.2- 55ºC, con un valor aproximadamente constante, en torno a 15 mM. El isómero n-butírico se acumuló los primeros días, coincidiendo con la acumulación de acético, llegando a concentraciones bastante altas, y mayores en el tratamiento con mayor proporción de residuo de pera, aunque a partir del día 13 la concentración detectada fue muy baja, del orden de unos pocos ppm (menos de 1 mM). A partir del día 20 aumentó la concentración de la forma iso del ácido butírico, estando en torno a los 4,5 mM durante bastantes días. Estos dos tratamientos (2.3 y 2.4) obtuvieron una mayor producción de metano, una menor duración del desfase inicial y una menor acumulación de ácidos grasos volátiles, que el control y que el tratamiento 2.2. La producción de metano mostró una mayor pendiente, mayor producción diaria en el período inicial que los otros tratamientos a esta temperatura, e incluso que el control y el 2.2 a 35ºC. El tratamiento 2.4 no mejoró al 2.3, y si se comparan índices de producción respecto a contenidos iniciales de materia orgánica, se podría decir que fue peor. La producción por gramo de sólidos volátiles fue igual (con un nivel de significación del 5%) que la producción del tratamiento 2.2. La velocidad específica de producción de metano (R’) fue mayor para este tratamiento que para el anterior. La mejora en el comportamiento inicial de los tratamientos con mayor proporción de residuo de pera se ha relacionado con una menor inhibición de la fase metanogénica por amoníaco, lo que explicaría el comportamiento intermedio observado para el tratamiento 2.2. La menor inhibición por amoníaco habría sido debida fundamentalmente a la bajada del pH 190 Ensayos de viabilidad de codigestión en discontinuo experimentada al mezclar con este tipo de cosubstrato y en menor medida a la menor concentración inicial de nitrógeno amoniacal de la mezcla (Tabla 5.25 y Tabla 5.18). Las concentraciones de propiónico y de las formas iso de butírico y valérico no disminuyeron, indicando una inhibición relativa de la acetogénesis durante todo el experimento por acumulación de hidrógeno, probablemente debido a la inhibición relativa de la metanogénesis hidrogenotrófica. En el rango termofílico no se produjo la acusada acumulación inicial de ácidos del rango mesofílico, que podría haber estado motivado por varios motivos a) un peor funcionamiento de la etapa acidogénica, que hizo que la generación de ácidos fuera más lenta y por tanto más equilibrada con las otras etapas; b) un mejor funcionamiento de la etapa metanogénica, con una velocidad de crecimiento muy superior al rango mesofílico, que no habría permitido la acumulación de ácidos o c) una mayor sensibilidad de los organismos metanogénicos y/o acetogénicos mesofílicos a la inhibición por acético que los termofílicos. De las tres hipótesis establecidas para explicar las diferencias observadas entre el rango termofílico y el mesofílico, la que parece más posible es la b, sobre todo al comparar entre los diferentes tratamientos en termofílico. La bajada inicial del pH con la adición del cosubstrato, hizo que disminuyera la concentración del principal tóxico para los metanogénicos, el amoníaco, pudiendo desarrollarse este cultivo libremente. La tasa específica de crecimiento de los microorganismos, en general, aumenta con la temperatura, por lo que la tasa de consumo de acético pudo ser mayor en termofílico que en mesofílico. La menor producción acumulada de metano en el régimen termofílico que en el mesofílico, sobre todo comparando el índice de producción de metano respecto a sólidos volátiles (Tabla 5.27 y Tabla 5.28), podría haberse debido al re-establecimiento de las condiciones de inhibición por amoníaco, ya que el consumo de ácidos provocó una subida del pH y por tanto un aumento en la concentración de amoníaco libre (Tabla 5.26). De hecho se detectó un aumento importante del pH al final del experimento. Este aumento, para el tratamiento 2.4 (con un 20% de pera), supuso pasar de un 2,9% a un 23% de amonio como amoníaco libre, que en g de NH3/L supuso pasar de 79,79 mg N-NH3/L a 770,01 mg N-NH3/L, y para el tratamiento de 2.3 (12,5% de pera) un aumento de 202,00 a 837,37 mg N-NH3/L . Optimización de la digestión anaerobia de purines de cerdo 191 5.5.3. Experimento 3. Codigestión de purín de cerdo con tierras decolorantes de aceite de oliva. Se realizó un experimento para determinar la viabilidad de la codigestión de purín de cerdo con tierras decolorantes de aceite de oliva (TDO), y para determinar la proporción de TDO que optimiza la producción de metano. De la mima forma que en los experimentos 1 y 2 se analizaron diversos parámetros, y básicamente la evolución de la producción de metano y de ácidos grasos volátiles. Como se verá en la exposición de resultados se ha eliminado de la discusión el tratamiento 3.4 (dilución de TDO con agua, sin purín), que se proponía en el apartado de materiales y métodos, ya que aunque se realizó el seguimiento de este tratamiento, exactamente igual que de los otros, la inclusión de resultados prácticamente nulos, y con muy baja variabilidad, podría provocar errores en el estudio estadístico. Todos los valores expuestos en las tablas corresponden a los valores medios. Los valores finales de los parámetros físico-químicos fueron corregidos con la concentración correspondiente del blanco (digestión del inóculo con agua). Evolución de compuestos nitrogenados La adición de tierras decolorantes como cosubstrato tuvo un efecto de dilución de los compuestos nitrogenados, tanto del nitrógeno Kjeldahl como del nitrógeno amoniacal (Tabla 5.32 y Tabla 5.33), debido al bajo contenido nitrógeno de este substrato. El Nk fue igual al inicio y al final del experimento, excepto para el tratamiento 3.3, atribuido a error analítico, más que a una diferencia real, no mostrando para ningún tratamiento diferencias significativas entre temperaturas. Aunque se ha realizado el test Lsmeans, que es útil básicamente para analizar interacciones entre los efectos principales, para el parámetro Nk la interacción de los efectos principales tratamiento (proporción de cosubstrato)*temperatura no fue significativa, siendo sólo significativos los efectos principales. Sin embargo, para el nitrógeno amoniacal, los tres efectos principales considerados (tratamiento, temperatura y día) fueron significativos, así como las interacciones dobles, excepto la interacción tratamiento*temperatura, ya que la concentración final fue mayor, para todos los tratamientos en el rango termofílico. La concentración final, fue también mayor que la inicial en todos los casos, fruto de hidrólisis de las proteínas (Tabla 5.33). 192 Ensayos de viabilidad de codigestión en discontinuo Tabla 5.32. Concentración de nitrógeno Kjeldahl en el experimento 3. Nitrog. Kjeldahl (mg Nk/Kg) Trat. % de TDO Inicial Final 35ºC Final 55ºC Dif. temp. 3.1 0% 5,26 d 5,36 d 5,35 d - 3.2 5% 4,92 bc 4,92 bc 5,07 c - 3.3 12,5% 4,61 a 4,89 b 4,90 b - Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significación del 95%). * o ** diferencias significativas para los valores finales entre las dos temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. Tabla 5.33. Concentración de nitrógeno amoniacal en el experimento 3. Nitrógeno amoniacal (mg N-NH4+/L) Trat. % de TDO Inicial Final 35ºC Final 55ºC Dif. temp. 3.1 0% 3,43 d 3,83 e 4,09 g ** 3.2 5% 3,20 b 3,70 e 3,92 f ** 3.3 12,5% 3,04 a 3,16 ab 3,40 c ** Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significación del 95%). * o ** diferencias significativas para los valores finales entre las dos temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. La mineralización queda también reflejada en la fracción de nitrógeno total, en forma de amonio, y en el incremento de dicha fracción (Tabla 5.34). En esta tabla se observa como el incremento es mayor en el rango termofílico que en el mesofílico en todos los casos. Tabla 5.34. Proporción de nitrógeno amoniacal sobre nitrógeno total. Incremento sufrido durante la digestión, en el experimento 3. ' N-NH4+/Nk (%) Fracción N-NH4+/NK (%) Trat. Inicial Final 35ºC Final 55ºC 35ºC 55ºC Dif. Temp. 3.1 64,70 71,94 76,74 7,48 b 12,28 cd * 3.2 64,30 74,44 78,84 10,13 bc 14,53 d * 3.3 63,30 65,86 70,83 2,52 a 7,48 b ** Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significación del 95%). * o ** diferencias significativas para los valores finales entre las dos temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. Optimización de la digestión anaerobia de purines de cerdo 193 Evolución de la materia orgánica Los parámetros de medida de materia orgánica proporcionaron, como en el resto de los experimentos, algunos valores contradictorios. El contenido de materia orgánica aumentó fuertemente por el aporte del cosubstrato tierras decolorantes (Tabla 5.35 y Tabla 5.36). El incremento de sólidos totales por la adición de tierras decolorantes fue de un 51,3% y un 138,9%. La concentración final de ST en el rango mesofílico fue menor que al inicio para todos los tratamientos, para el rango mesofílico. Sin embargo, en el rango termofílico, no hubo diferencias significativas entre el inicio y el final. No se detectaron diferencias en la eliminación de ST entre el tratamiento 3.2 y el control. Sí las hubo entre el 3.3 y el 3.2 y el control. Tabla 5.35. Concentraciones medias y eliminación de sólidos totales, en el experimento 3. Sólidos Totales (ST) g/kg Trat. % de TDO Eliminación ST (%) Inicial Final 35ºC Final 55ºC 35ºC 55ºC 8,20 C 12,75 D 3.1 0 78,33 b 72,08 a 68,30 a 3.2 5 118,50 d 106,10 c 118,80 d 3.3 12,5 187,14 f 177,80 e 189,78 f 10,40 CD -0,25 A 5,00 B -1,33 A Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significación del 95%). El incremento de sólidos volátiles por efecto de la adición de las tierras decolorantes no fue tan espectacular como el de ST, debido al relativamente bajo contenido en SV de este residuo (Tabla 5.3), siendo 23,9% y 60,7% para los tratamientos 3.2 y 3.3, respectivamente. La evolución de SV fue similar a ST. La máxima eliminación se obtuvo para el tratamiento 3.2 a 35ºC, aunque del mismo orden de magnitud que la obtenida para el 3.1 a 55ºC (ya se comentó en el apartado 4.1.2 la posible razón de la mayor eliminación de SV en termofílico). La eliminación de SV para el tratamiento 3.3 a 35ºC también fue significativamente mayor que la obtenida para el control. Los valores de DQO experimentaron un incremento aun más importante, de 55% y un 150% para los dos tratamientos (Figura 5.20). Esto se debe al gran contenido de materia orgánica de este material, dado que entre el 30 y el 40% del peso seco se trata de grasa, básicamente restos de aceite de oliva. 194 Ensayos de viabilidad de codigestión en discontinuo Tabla 5.36. Conc. medias y eliminación de sólidos volátiles en el experimento 3. Sólidos Volátiles (SV) g/kg Trat. % de TDO Eliminación SV (%) Inicial Final 35ºC Final 55ºC 35ºC 55ºC 3.1 0 58,15 b 50,26 a 45,83 a 13,80 B 21,00 CD 3.2 5 72,03 c 55,26 b 68,28 c 23,20 D 5,25 A 3.3 12,5 98,70 e 80,32 d 94,10 e 18,60 C 4,33 A Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significación del 95%). Las eliminaciones de DQO observada en el rango termofílico (Tabla 5.37 y Figura 5.20) aunque fueron menores que las obtenidas en el rango mesofílico, no se correspondieron con las eliminaciones de sólidos volátiles, ni con la producción de gas. La dificultad en la toma de muestra homogénea pudo ser la causa de estas irregularidades. Tabla 5.37. Concentraciones medias y eliminación de DQO, en el experimento 3. Trat. % de TDO DQO g O2/kg Eliminación DQO (%) Inicial Final 35ºC Final 55ºC 35ºC 55ºC 3.1 0 80,04 bc 62,00 a 69,72 ab 23,37 AB 13,82 A 3.2 5 134,22 d 71,34 abc 92,07 c 45,70 C 29,90 BC 3.3 12,5 200,00 e 124,70 d 139,66 d 39,80 C 32,60 C Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significación del 95%). Evolución del pH Los valores de alcalinidad total y intermedia disminuyeron al añadir el cosubstrato, tal y como se muestra en la Tabla 5.38. La relación de alcalinidades aumentó bastante, situándose en el tratamiento 3.3 en 0,38, indicando una alta concentración de AGV. Optimización de la digestión anaerobia de purines de cerdo DQO (g/Kg) 195 Eliminación (%) 50% 45% 40% 35% 30% 25% 20% 15% 10% 5% 0% 250 200 150 100 50 0 3.1 3.2 3.3 Inicial Final 35ºC Eliminación 35ºC Eliminación 55ºC Final 55ºC Figura 5.20. Eliminación de DQO para los diferentes tratamientos, en el experimento 3. Tabla 5.38. Comparación de la alcalinidad total y al bicarbonato (pH 5,75) inicial, en el experimento 3 Tratamiento Alcalinidad Total (g CaCO3/L) Alc. parcial (g Ca CO3/L) Relación alcalinidad 3.1 10,55 7,5 0,29 3.2 9,50 6,4 0,33 3.3 8,10 5,1 0,38 La adición del cosubstrato, supuso una importante bajada del pH, de 8 a 7,7 y 7,3 al añadir un 5% o un 12,5% de tierras (Tabla 5.39), aunque se mantuvo a niveles aceptables para el desarrollo de la digestión anaerobia. El pH al final del experimento fue diferente en función de la temperatura, observándose un aumento en el régimen mesofílico y una ligera tendencia a disminuir en el termofílico, mientras que en el 3.3 no se detectaron diferencias entre el inicio y el final, indicando que no hubo recuperación del sistema. 196 Ensayos de viabilidad de codigestión en discontinuo Tabla 5.39. Valores medios de pH y de amoníaco libre, en el experimento 3. pH Trat. % de TDO Inicial 3.1 0 8.06 d 7.70 b 3.2 5 7.69 b 3.3 12,5 7.33 a NH3 (mg N- NH3/L) Final 35ºC Final 55ºC 35ºC 55ºC 7.99 d 305.80 1106,2 7.89 cd 7.50 a 238.62 497.02 7.76 bc 7.47 a 135.26 335.20 Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significación del 95%). La concentración de amoníaco libre, al bajar el pH y el amonio, se mantuvo a unos niveles relativamente bajos (Tabla 5.39), muy por debajo de los niveles de amoníaco encontrados en el tratamiento más diluido de purín (experimento 1). Evolución de la producción de biogás En la Figura 5.21 y Tabla 5.40 se observa la gran diferencia en la producción de metano obtenida entre los dos regímenes de temperatura. La codigestión con tierras en una proporción de 5% de tierras y 95% de purín a 35ºC alcanzó el valor máximo del experimento, siendo mayor también que los máximos valores alcanzados al utilizar como cosubstrato residuo de pera. Sin embargo a 55ºC resultó en un completo fracaso, con las peores producciones de todos los experimentos, casi tan bajas como el blanco (inóculo+agua), lo que indica la presencia de un inhibidor más potente que el nitrógeno amoniacal. El aumento en la producción de metano en el rango mesofílico no fue únicamente fruto de la mayor cantidad de sólidos volátiles de este substrato, ya que aumentó mucho el índice de producción respecto a los sólidos volátiles (Tabla 5.41), alcanzando el tratamiento 35º 3.2 un valor de 340 mL/g SV, frente a los 150 mL/g SV del control. La mayor relación SV/DQO de este substrato influyó para la consecución de la mejora del índice. Optimización de la digestión anaerobia de purines de cerdo 197 mL de CH4 1400 1200 35º 3.1 1000 35º 3.2 800 35º 3.3 600 55º 3.1 55º 3.2 400 55º 3.3 200 0 0 10 20 30 40 50 60 70 80 Figura 5.21. Evolución de la producción de metano acumulada, en el experimento 3 La evolución de la producción acumulada de CO2 siguió un patrón parecido al observado para el metano (Figura 5.22). La composición del biogás fue muy rica en metano, llegando casi al 70% el tratamiento 35º 3.2. Tabla 5.40. Producción acumulada de gas (M), para las variables biogás, CH4 y CO2 en el experimento 3. Trat. M (mL de biogás) 35º 55º 3.1 658,68 c 436,99 bc - M (mL de CH4) 35º 55º 443,13 b 195,18 A 6,82 A M (mL de CO2) 35º 55º * 215,55 ab 241,81 B ** 582,89 c - 3.2 1871,93 e 171,52 a ** 1289,04 c 164,69 A ** 3.3 1595,63 d 339,65 ab ** 1050,13 d 136,51 A ** 535,23 c 203,14 AB ** Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significación del 95%). * o ** diferencias significativas para los valores finales entre las dos temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. Dentro del rango mesofílico la producción acumulada de metano y biogás fue mayor para el tratamiento 3.2 que para el 3.3. Sin embargo no se detectaron diferencias significativas en la producción acumulada de CO2 (Tabla 5.40). Las diferencias aumentaron a favor del 3.2 (5% de TDO) al comparar los índices de producción respecto a los sólidos volátiles iniciales (Tabla 5.41). Ambos tratamientos superaron los obtenidos por el control, para todos los parámetros considerados. 198 Ensayos de viabilidad de codigestión en discontinuo Tabla 5.41. Valores medios de los índices de producción acumulada de gas respecto al contenido inicial de sólidos volátiles (B), para las variables biogás, metano y dióxido de carbono, en el experimento 3. Trat. B (mL de biogás/g SVini) 35º 55º B (mL de CH4/g SVini) 35º 55º B (mL de CO2/g SVini) 35º 55º 3.1 223,16 c 148,05 b ** 150,13 C 66,13 B ** 3.2 493,33 e 45,20 a ** 339,72 E 1,80 A ** 153,62 D 43,41 A ** 3.3 315,42 d 67,14 a ** 207,59 D 26.99 AB ** 105,80 C 40,16 A ** 73,03 B 81,93 B - Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significaión del 95%). * o ** diferencias significativas para los valores finales entre las dos temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. El valor obtenido de producción de metano respecto a sólidos volátiles no fue, sin embargo muy alto comparado con valores obtenidos en la bibliografía, en donde es normal encontrar valores en torno a 650 mL CH4/g SV, o incluso más altos, aunque trabajando con otro tipo de residuos (Salminen et al.,1999). mL de CO2 700 600 35º 3.1 500 35º 3.2 400 35º 3.3 55º 3.1 300 55º 3.2 200 55º 3.3 100 0 0 10 20 30 40 Día 50 60 70 80 Figura 5.22. Evolución de la producción de CO2 acumulada en el experimento 3. El comportamiento en el rango termofílico fue inverso al observado en el mesofílico, habiendo obtenido el tratamiento 3.2 a 55ºC las producciones más bajas de todos los experimentos, excepto los blancos. El efecto fue mucho más marcado al comparar el índice respecto a los sólidos volátiles iniciales, ya que la cantidad total de materia orgánica de estos substratos fue Optimización de la digestión anaerobia de purines de cerdo 199 mayor que en el control. Debido a las grandes desviaciones obtenidas en los tratamientos 3.2 y 3.3 a 55ºC no se pudieron diferenciar estadísticamente las medias de producción de metano acumulada respecto al control, aunque el tratamiento 3.2 sí se diferenció al comparar el índice respecto a los sólidos volátiles. Al comparar la curva de producción de metano y valores a lo largo de la misma, se observa que el tratamiento 3.2 a 55ºC estuvo siempre, ya en el día 6, por debajo tanto de los otros tratamientos en el rango termofílico, como de su homólogo en el mesofílico. Sin embargo el 3.3 55ºC superó durante el período inicial (hasta el día 20) a su homólogo a 35ºC, aunque al final el tratamiento a 35ºC obtuvo una producción mucho mayor. La producción de CO2 (Figura 5.22) mostró un comportamiento algo diferente al metano y, aunque el crecimiento exponencial coincidió temporalmente con el del metano, durante la fase inicial la producción de CO2 fue bastante importante. La producción de CO2 para los tratamientos 3.2 y 3.3 en mesofílico estuvo siempre por encima del control. El 3.3 tuvo una mayor producción de CO2 inicialmente que el 3.2, siendo superado por este el día 21. En el rango termofílico, las producciones de CO2 y los índices de producción de CO2 respecto a los sólidos volátiles, estuvieron siempre por debajo del control, y fue menor para el tratamiento 3.2 que para el 3.3. Análisis de parámetros de velocidad, producción potencial y duración de la fase lag. Se aplicó la ecuación de Gompertz (ecuaciones 5.5 y 5.6), de forma similar a los experimentos anteriores, obteniendo los valores de los parámetros que se muestran en la Tabla 5.42. No se aplicó al tratamiento 3.2 en termofílico, ya que no se consiguieron coeficientes de regresión aceptables. Para el resto de tratamientos el ajuste fue muy bueno, tal y como se observa en la Figura 5.23. Los resultados del test de separación de medias se muestran en la Tabla 5.42. Los parámetros de velocidad de producción de metano, tanto K (mL CH4/día), como R (mL CH4/g SV·día) y R’ (día-1), obtuvieron los valores máximos para el tratamiento 3.2, siendo diferente de los otros tratamientos en mesofílico. En el rango termofílico, pese a los buenos resultados finales obtenidos se observó la existencia de un desfase en el inicio de la producción de gas (fase lag), que aumentó con la concentración de TDO en la mezcla. La duración del desfase para el tratamiento 3.2 fue de 14, indicando un ligero desequilibrio inicial, mientras que para el 3.3 fue de 40 días, mostrando un 200 Ensayos de viabilidad de codigestión en discontinuo mayor desequilibrio, o presencia de algún tóxico. mL de CH4 /g SVini mL de CH4 /g SVini 400 400 350 350 2.2-35ºC 300 300 250 250 200 200 3.1-35ºC 150 150 100 100 3.3-35ºC 50 3.3-55ºC 3.1-55ºC 50 0 0 0 20 3.1-35ºC 3.2- 35ºC 3.3-35ºC 40 60 Ajuste Gompertz 80 0 20 40 3.1- 55ºC 3.2-55ºC 3.3-55ºC Día 60 80 Día Ajuste Gompertz Figura 5.23. Comparación de los datos experimentales media de las cinco repeticiones (puntos) y la curva de la simulación del modelo Gompertz, en el experimento 3, para el índice producción de metano respecto a sólidos volátiles iniciales Tabla 5.42. Cálculo de parámetros mediante la ecuación de Gompertz en el experimento 3 Trat. K M0 mL CH4 ml CH4/d O d B0 mL CH 4 g SV 3.1 35ºC 428,83 ab 11,08 b 4,04 ab 144,98 ab 3.2 35ºC 1268,13 b 49,91 d 14,45 c 3.3 35ºC 3460,27 c 30,48 c 3.1 55ºC 207,21 a 4,21 a 3.2 55ºC - 3.3 55ºC 155,69 a 2,51 a R mL CH 4 g SV·d R’ d-1 3,76 b 0,07 c 334,21 b 13,15 d 0,11 d 40,24 d 692,74 c 6,07 c 0,03 a 6,35 b 70,21 a 1,43 a 0,06 bc -1,69 a 30,78 a 0,50 a 0,05 b Letras diferentes significan valores medios diferentes. (Test Lsmeans a un nivel de significación del 95%). Análisis estadístico realizado por separado para cada variable (cada columna). Índice de metanización, acidificación y biodegradabilidad Los índices de metanización y acidificación alcanzaron el valor máximo, de forma similar al resto de índices considerados, para el tratamiento 3.2 (5% de Optimización de la digestión anaerobia de purines de cerdo 201 TDO) a 35ºC, siendo significativamente diferente del resto de tratamientos. En el rango termofílico, la adición de TDO supuso la inhibición tanto de la fase metanogénica, como de la acidogénica (Tabla 5.43), como demuestran los bajos valores obtenidos para los índices de acidificación y metanización, siendo la fase metanogénica más sensible, dado que el segundo es más bajo que el primero. En el rango mesofílico, no se detectaron diferencias entre el tratamiento 3.3 (12,5% de TDO) y el control, tanto para el índice de metanización, como el de acidificación. Este último fue más alto que el primero, indicando una ligera acumulación de ácidos, es decir, un peor funcionamiento de la etapa metanogénica. La mayor inhibición del tratamiento 3.3 se manifiesta, principalmente por la mayor duración de la fase lag, y la mayor lentitud de desarrollo del proceso, no pudiendo concluir, que la biodegradabilidad es menor, sino más lenta. Tabla 5.43. Índices de metanización, acidificación y biodegradabilidad en el experimento 3 Trat. Metanización (%M) 35ºC 55ºC Acidificación (%A) 35ºC 55ºC Biodegradabilidad (%BD) 35ºC 55ºC 3.1 29,44 c 12,96 b ** 29,54 C 19,68 B ** 36,62 C 23,94 B ** 3.2 51.08 d 0.28 a ** 51.16 D 3.22 A ** 64.42 D 3.40 A ** 3.3 27.93 c 3.64 a ** 30.28 C 4.76 A ** 38.14 C 5.70 A ** Letras diferentes significan valores medios diferentes. (Test Lsmeans a un nivel de significación del 95%). Análisis estadístico realizado por separado para cada uno de los días y cada variable. * o ** diferencias significativas para los valores finales entre temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. Evolución de ácidos grasos volátiles La concentración de AGV total (Figura 5.24) en los tratamientos del experimento 3, llegó a valores bastante altos, aunque muy por debajo de las concentraciones alcanzadas por las mezclas de purín y residuo de pera (tratamiento 2.4). Rango mesofílico La concentración de AGV del tratamiento 3.2 a 35ºC aumentó en los primeros días, llegando a un máximo el día 8 de 68,9 mM. A partir de este momento comenzó a disminuir lentamente. En el período entre los días 20 y 40 experimentó un nuevo máximo relativo, de 47,5 mM, disminuyendo 202 Ensayos de viabilidad de codigestión en discontinuo después hasta unos niveles muy bajos del orden de 1-2 mM. AGVt (mM )-55ºC AGVt (mM )-35ºC 175 150 175 125 125 100 100 75 75 50 50 25 0 25 150 0 0 10 20 30 40 Día 50 55º 3.1 35º 3.1 60 70 80 55º 3.2 35º 3.2 0 10 20 30 40 Día 50 60 70 80 55º 3.3 35º 3.3 Figura 5.24. Evolución de la concentración total de Ácidos Grasos Volátiles, en el experimento 3 En el tratamiento 35º 3.2, el ácido acético sufrió una acumulación inicial, con un máximo en el día 8 de 2,8 g Ac/L (Figura 5.25). A partir de este momento disminuyó rápidamente, pero la concentración volvió aumentar el día 27, llegando a 1,5 g Ac/L, a pesar de que la producción de metano estaba en el período de crecimiento exponencial. La concentración de acético fue menor que la del control en el día 6, pero se igualaron (no diferencias significativas) el día 13 (Tabla 5.44). El propiónico se acumuló en el tratamiento 3.2 a 35ºC durante los primeros días, de forma similar al control, pero se mantuvo a un nivel alto y aproximadamente constante hasta el día 20, sobre 1,1-1,2 g Pr/L, momento a partir del cual se acumuló más, llegando a alcanzar el máximo el día 36 de 22.8 mM (1,69 g Pr/L). A partir de este máximo disminuyó muy rápidamente siendo despreciable el día 56. La concentración de los isómeros iso tanto del butírico como del valérico para este tratamiento mostraron una tendencia similar al propiónico hasta el día 30, pero a partir de aquí no reflejaron el máximo mostrado por el propiónico, mostrando una tendencia a disminuir, con una comportamiento muy similar al control (Figura 5.26). La concentración del n butírico, siguió un comportamiento bastante similar al observado por el acético, corroborando que el acético es el principal modulador de la degradación de los isómeros n del butírico. Por su lado el n valérico mantuvo un nivel de concentración bastante bajo, por debajo de 0.5 mM. Optimización de la digestión anaerobia de purines de cerdo 203 Tabla 5.44. Valores de concentración de ácidos grasos volátiles (mM) obtenidos en diferentes momentos de la digestión, en el experimento 3 Concentración media de ácidos grasos volátiles total (mM) Día 6 55ºC 35º C Día 13 55ºC 35º C 3.1 81,50 b 96,76 c 64,31 A 153,17 B ** 0,99 A 62,23 D ** 3.2 51,63 a 101,12 c ** 38,79 A 142,40 B ** 1,15 A 49,31 CD ** 3.3 83,79 b 104,73 c ** 126,21 B 126,58 B * 35º C Final 55ºC Trat. - 41,09 BC 25,37 B - Concentración media de acético (mM) Día 6 55ºC 35º C Día 13 55ºC 35º C Final 55ºC Trat. 35º C 3.1 63,03 c 79,60 d ** 26,17 A 133,12 C ** 0,99 A 46,28 C ** 3.2 36,17 a 82,18 d ** 16,96 A 121,29 C ** 1,14 A 40,12 C ** 3.3 53,90 b 90,42 e ** 88,71 B 109,11 BC - 7,67 AB 18,09 B - Final 55ºC 6,05 C ** Concentración media de propiónico (mM) Día 6 35º C 55ºC 10,26 c 5,82 a 35º C ** 29,36 C Día 13 55ºC 7,32 A ** 35º C 0A 3.2 11,20 c 7,98 b ** 16,42 B 7,38 A ** 0,01 A 3,84 B ** 3.3 24,26 d 6,59 ab ** 31,25 C 6,15 A ** 31,07 D 3,27 B ** Trat. 3.1 Letras diferentes significan valores medios diferentes (Test Lsmeans a un nivel de significación del 95%). Análisis estadístico realizado por separado para cada uno de los días y cada variable. * o ** diferencias significativas para los valores finales entre las dos temperaturas, a un nivel de significación del 95% o del 99%, respectivamente. El tratamiento 3.3 sufrió una mayor acumulación de ácidos, con un máximo el día 17 (134.6 mM) y a partir de este momento comenzó a disminuir lentamente, para a partir del día 36 mantenerse aproximadamente constante. El acético mostró una tendencia ascendente hasta el día 17, con un máximo 93,7 mM (5,63 g Ac/L). A partir del día 20 de observó una fuerte bajada en la concentración, llegando al final del experimento con un nivel muy bajo (0,13 g/L). El propiónico se acumuló durante los primeros días, de forma similar a lo ocurrido en el 3.2, pero llegando a un máximo relativo más alto, 34,9 mM el día 20. Después se produjo una tenue bajada, para volver a aumentar ligeramente al final, obteniéndose el máximo absoluto de este tratamiento al final del experimento, en el día 75 (39,9 mM). 204 Ensayos de viabilidad de codigestión en discontinuo Acético (mM )-55ºC Acético (mM )-35ºC 140 120 140 120 100 80 60 40 100 80 20 0 20 0 60 40 0 10 20 30 40 50 60 70 0 80 Propiónico (mM )-35ºC 10 20 30 40 50 60 70 80 30 40 Día 50 60 70 80 Propiónico (mM )-55ºC 50 50 40 40 30 30 20 20 10 10 0 0 0 10 20 30 40 Día 50 60 70 55º 3.1 35º 3.1 80 0 55º 3.2 35º 3.2 10 20 55º 3.3 35º 3.3 Figura 5.25. Evolución del acético y propiónico, en el experimento 3 La concentración de AGVt al final fue mayor para el tratamiento 3.3 que para el 3.2 y el control, debido, sobre todo a la alta concentración de propiónico alcanzada al final del experimento, indicando que el sistema aún estaba evolucionando. La concentración de AGV se mantuvo durante todo el experimento por encima de los otros tratamientos. La evolución del acético fue también diferente para los dos tratamientos de mesofílico, alcanzándose unos valores más altos para el tratamiento más concentrado (3.3) (Figura 5.25). El propiónico también estuvo por encima durante casi todo el experimento, alcanzándose las mayores diferencias al final. Optimización de la digestión anaerobia de purines de cerdo Iso-butírico (mM)-35ºC 50 205 Iso-butírico (mM)-55ºC 50 40 40 30 30 20 20 10 10 0 0 0 10 20 30 N-butírico (mM)-35ºC 6 5 4 3 2 1 0 0 10 20 30 40 40 50 50 60 60 70 70 80 80 Iso-valérico (mM)-35ºC 0 10 20 30 N-butírico (mM)-55ºC 6 5 4 3 2 1 0 0 10 20 30 40 50 60 70 80 40 50 60 70 80 40 50 60 70 80 60 70 80 Iso-valérico (mM)-55ºC 8 8 7 6 5 4 3 2 1 0 6 4 2 0 0 10 20 30 40 50 60 70 80 N-valérico (mM)-35ºC 0 10 20 30 N-valérico (mM)-55ºC 1.0 1.0 0.8 0.8 0.6 0.6 0.4 0.4 0.2 0.2 0.0 0.0 0 10 20 30 40 Día 50 60 55º 3.1 35º 3.1 70 80 0 55º 3.2 35º 3.2 10 20 30 40 Día 50 55º 3.3 35º 3.3 Figura 5.26. Evolución de los isómeros de butírico y valérico, en el experimento 3. 206 Ensayos de viabilidad de codigestión en discontinuo Rango termofílico En el tratamiento 55º 3.2 la concentración de AGV aumentó hasta llegar a un máximo de 142 mM el día 15. A partir de este momento la concentración comenzó a disminuir pero muy lentamente, estando el día 57 en 49,31 mM. El tratamiento 55º 3.3 se acumuló en el primer período y después tuvo una tendencia a disminuir, aunque muy lentamente. El máximo se alcanzó el día 15 con 126.6 mM, y el día 70 la concentración de AGV era de 25,37 mM. La evolución del acético para los dos tratamientos (3.2 y 3.3) fue bastante similar al control, con una acumulación desde el inicio hasta el día 15, donde se alcanzó el máximo absoluto tanto del tratamiento 3.2 como del 3.3, siendo este un poco menor que el obtenido por el control, pero no significativamente menor. A partir de este momento la tendencia predominante es la descendente, aunque muy lentamente, y en el 3.2, se observó una nueva subida, con un máximo sobre el día 40. El propiónico se mantuvo al nivel del control, y aproximadamente constante durante todo el experimento. Lo mismo ocurrió con los dos isómeros de los ácidos butírico y valérico, estando todos en unos niveles similares a los observados para el control, sin que se apreciara una clara tendencia. La producción de metano para el tratamiento con un 5% de tierras filtrantes en el rango mesofílico mostró la máxima producción de metano de todos los tratamientos de los tres experimentos realizados, con una producción acumulada que casi triplicó a la obtenida por el control. El aumento tan espectacular de la producción de metano en el régimen mesofílico se debió, al mayor potencial de producción de metano del cosubstrato, que es una mezcla de tierras con gran superficie específica, y materia orgánica adsorbida, formada principalmente por grasas (alrededor de un 30%). El potencial de metano que se puede obtener de los lípidos y grasas es más alto que para otros tipos de macromoléculas. En el rango termofílico los resultados obtenidos fueron extraordinariamente bajos, contradiciendo resultados obtenidos por otros autores como Angelidaki y Ahring (1992, 1993a), que obtuvieron una gran mejora al introducir tierras procedentes del filtrado y decolorado de aceites comestibles como co-substrato. En este caso se observa que en algunos tramos se obtuvo menor producción de metano que la obtenida por degradación de la materia orgánica residual contenida en el inóculo. El fracaso del experimento termofílico se atribuye a que la toxicidad de los ácidos grasos de cadena larga, en especial del oleico, es mayor en el rango termofílico que en el mesofílico. Las constantes de inhibición (concentración Optimización de la digestión anaerobia de purines de cerdo 207 de tóxico para una reducción del 50% de la tasa máxima) por oleico se han determinado entre 98.7 a 223 mg/L a 55ºC, mientras que el lodo mesofílico es hasta 12 veces más tolerante respecto a la concentración de AGCL, yendo de 663 a 1200 mg/L (Hwu et al., 1997). El efecto inhibitorio de glicerol trioleato se ha observado a partir de 2 g/L (Angelidaki et al., 1990). Por otro lado, el efecto inhibidor de los ácidos grasos de cadena larga provoca un aumento de la duración de la fase lag en ensayos en discontinuo (Hanaki et al., 1981), tal y como se ha constatado en el rango mesofílico (Tabla 5.42). Angeldaki y Ahring concluyeron que los ácidos grasos libres de cadena larga, oleico y estérico, inhiben todos los pasos de la digestión anaerobia termofílica, provocando, a una concentración de 0,2 g/L de oleico, el aumento en la duración de la fase lag, mientras que el crecimiento bacteriano es completamente inhibido a una concentración de 0.5 g/L de oleico y 1.0 de estereato (Angeldaki y Ahring, 1992). El potencial tóxico de los AGCL puede ser contrarrestado con la adición de sales cálcicas, básicamente porque se produce la precipitación de los ácidos grasos libres (Hanaki et al., 1981) y por la adición de arcillas con gran superficie activa, como bentonita (Angelidaki et al., 1990). Este efecto beneficioso puede explicar los buenos resultados obtenidos a escala real en plantas termofílicas de Dinamarca, al tratar mezclas de residuos ganaderos con tierras decolorantes. Además, las tierras decolorantes más utilizadas en las plantas centralizadas de Dinamarca provienen del refinado de aceites de semillas, especialmente de colza, que tiene un bajo contenido en ácido oleico (Frías et al., 1991). El oleico ha sido descrito como uno de los ácidos de mayor poder tóxico, casi tan tóxico como el más tóxico de los saturados (Koster y Cramer, 1987). Las tierras decolorantes utilizadas en el presente trabajo, procedían del proceso de refinado de aceite de oliva. La composición en ácidos grasos de cadena larga varía mucho en función del tipo de aceite, así el de oliva tiene entre un 60 y un 85% de oleico (C18:1), y entre un 4 y un 6% de linoleico (C18:2), mientras que en el aceite de colza la proporción de oleico está entre 10-20% y entre 12-29% de linoleico. Las tierras utilizadas tienen un contenido de grasa aproximado del 27% sobre materia seca (Tabla 5.3), constituido, principalmente por restos de aceite de oliva, y por tanto el principal ácido graso presente es el oleico. Haciendo un cálculo teórico a partir del contenido en grasa de las tierras añadidas y la proporción en que estas se añadieron, se estimó que la concentración de triglicéridos está en 13 g/L para el tratamiento 3.2 (5% de TDO) y 32,7g /L para el tratamiento 3.3 (12,5 de TDO). Puesto que la concentración de oleico está entre un 60 y un 208 Ensayos de viabilidad de codigestión en discontinuo 85% de los ácidos grasos, el contenido en tri-oleico glicérido estará entre 7,8 y 11 g /L para el 3.2 y entre 19,6 y 27,7 g /L para el tratamiento 3.3, valores muy superiores a los 5 g/L encontrados inhibidores de la digestión anaerobia termofílica por Angelidaki et al. (1990). Estos valores son muy superiores también a los encontrados como inhibidores por Hwu et al. (1997), que cifraba la concentración de ácido oleico que reducía la actividad metanogénica en un 50% en 1.21 g /L a 30ºC y 0,099-0,22 g /L a 55ºC, mientras que en el presente experimento la concentración de oleico ha estado en el intervalo comprendido entre 1,76 y 6,25 g de oleico/L, por tanto muy por encima de los valores inhibidores a ambas temperaturas, pero sobre todo a 55ºC. A pesar de los buenos resultados obtenidos para el tratamiento 3.2 del rango mesofílico, al analizar la curva de producción acumulada de metano (Figura 5.21) y las curvas de evolución de ácidos grasos (Figura 5.25 y Figura 5.26) se observa la existencia de inhibición. La producción acumulada de metano mostró un desfase inicial (Tabla 5.40 y Tabla 5.42), relacionado con la inhibición de la fase metanogénica. Sin embargo, el valor máximo de acético fue menor que en el control (Tabla 5.44). El hecho de que el acético no se acumulara indica que hubo inhibición de la etapa acetogénica y/o acidogénica. El propiónico sufrió una acumulación más prolongada que el acético y mayor que en el control, aunque no llegó a unos niveles tan altos como en el tratamiento 2.4 (20% de residuo de pera). La evolución de propiónico alcanzó un máximo en el momento en máxima producción de metano, sin que se produjera acumulación de acético, lo que indica, con alta probabilidad, la inhibición parcial de la ruta metanogénica a partir de hidrógeno (Hill y Cobb, 1993). Hanaki et al. (1981) demostró el efecto inhibidor de los ácidos grasos de cadena larga en el rango mesofílico sobre varias de las fases del proceso anaerobio, siendo las más afectadas la metanogénesis a partir de acético y la acetogénesis a partir de ácidos grasos de cadena larga y de n-butirato, y el principal efecto observado fue la mayor duración del desfase inicial en el inicio de la producción de gas. La metanogénesis a partir de hidrógeno está inhibida por altas concentraciones de ácidos grasos, afectando a la velocidad específica de producción de metano, y no a la duración del desfase en el curva de crecimiento (Hanaki et al., 1981). Esto explica que el propiónico se acumulase una vez finalizado el desfase en la producción de metano, ya que debió haber también un desfase en el inicio de la actividad de los acetogénicos, por lo que la producción de hidrógeno debió ser suficientemente baja como para que los microorganismos metanogénicos Optimización de la digestión anaerobia de purines de cerdo 209 consumidores de hidrógeno, a pesar de la baja velocidad de crecimiento debido a la inhibición por AGCL, fueran capaces de consumir todo el H2 producido. El tratamiento 3.3 mostró una mayor acumulación de todos los ácidos grasos volátiles y la duración del desfase inicial fue mucho mayor que en el tratamiento anterior, siendo del orden de los 40 días (Tabla 5.42), debido a la mayor concentración de ácidos grasos de cadena larga, confirmando los resultados obtenidos por autores previos, que relacionan la concentración de inhibidor (AGCL) con la duración del desfase y el grado de inhibición (Hanaki et al., 1981; Koster y Cramer, 1987; Angelidaki y Ahring, 1992). La evolución de propiónico fue similar a la ocurrida en el tratamiento 3.3, pero más retrasado, fruto de la mayor duración del desfase de la fase acetogénica y alcanzando mayores concentraciones, fruto del mayor nivel de inhibición (mayor concentración de AGCL) y del mayor aporte inicial de materia orgánica. La concentración de propiónico máxima se alcanzó, también, en el momento de mayor producción de metano. El diferente poder inhibidor de los ácidos grasos en función de la temperatura demostrado por Hwu et al., (1997) se ha constatado en el presente experimento. A 55ºC durante los 80 días de digestión no se observaron signos de recuperación del proceso, habiéndose obtenido la inhibición total del sistema. Curiosamente fue el tratamiento más diluido el que obtuvo menor producción de metano, indicando que se la inhibición total se produjo con un contenido de TDO del 5%. Se observó inhibición sobre todos las fases de la digestión anaerobia, incluyendo la acidogénesis, dado que no hubo una excesiva acumulación de ácidos grasos volátiles. 5.6. Conclusiones Las conclusiones que se pueden extraer de los resultados del presente capítulo son: 5.6.1. Sobre el método La utilización de un pequeño volumen de inóculo permite, en un único test, estimar la viabilidad de un determinado substrato para ser digerido anaeróbicamente, ya que proporciona concentraciones de los posibles tóxicos, que podrían ser desconocidos, cercanos a los valores reales en la mezcla problema. La cantidad de inóculo utilizada (10% en peso del substrato), en algunos 210 Ensayos de viabilidad de codigestión en discontinuo casos ha sido insuficiente, provocando un efecto similar a una sobrecarga orgánica, que se ha traducido en el retraso inicial de la producción de metano, aumento de la duración de la fase lag. Aunque no se pueden utilizar modelos sencillos de primer orden para la determinación de parámetros cinéticos, se han conseguido, buenos ajustes a un modelo, también sencillo, basado en la ecuación de Gompertz, que tiene en cuenta la duración de la fase lag. La combinación de los parámetros velocidad de producción de metano, duración de la fase lag, evolución de producción acumulada de metano y evolución de la concentración de los AGV individuales proporciona suficiente información para el estudio del proceso e identificación de problemas y causas. La presencia de tóxicos en el medio impide la determinación del potencial de producción de metano y la biodegradabilidad del substrato, que sí se podría determinar mediante el test estándar de biodegradabilidad. El tamaño de las muestras tomadas durante el experimento no permite hacer una analítica completa, impidiendo el estudio de la evolución de parámetros interesantes, pH, amonio, DQO soluble, etc. La extracción de muestras periódicas de la fracción líquida para el análisis de AGV, mediante jeringa hipodérmica, podría alterar la concentración de sólidos en el vial, lo que podría llevar a errores en la determinación de la eliminación de sólidos totales y volátiles y DQO. La medida indirecta de la producción de gas hace depender la exactitud de la estimación de la bondad de la calibración del método cromatográfico, y de la precisión en el volumen de muestra tomado. 5.6.2. Del experimento 1: Diluciones de purín - efecto del amonio. La digestión anaerobia de purines de cerdo con una concentración de nitrógeno amoniacal inicial de 3,43 g N-NH4+/L es posible en el rango mesofílico, sin que se haya observado inhibición por amonio, ni por amoníaco libre. La concentración de amoníaco libre que inhibe el proceso en el rango mesofílico está por encima de los 305 mg de N-NH3/L. La digestión anaerobia de purines de cerdo con una concentración de 3,43 g N-NH4+/L en el rango termofílico es posible aunque con una producción de metano mucho menor que la obtenida para el rango mesofílico. El proceso en el rango termofílico está afectado fundamentalmente por la inhibición por amoníaco libre. La concentración de amoníaco libre a partir de la cual el proceso se inhibe es ligeramente menor de 640 mg de NNH3/L, correspondiente con una concentración de nitrógeno amoniacal Optimización de la digestión anaerobia de purines de cerdo 211 inicial de 2,63 g N-NH4+/L, aunque no se pudo establecer claramente el valor límite. El principal proceso inhibido en los tratamientos más concentrados fue la metanogénesis acetoclástica, pero también resultó afectada la hidrólisis de proteínas y la acidogénesis. La hidrólisis de proteínas es mayor en el rango termofílico que en el mesofílico, produciendo un mayor incremento de la concentración de nitrógeno amoniacal. La inhibición de la ruta metanogénica a partir de hidrógeno pareció más afectada en el rango mesofílico que en el termofílico. 5.6.3. Del experimento 2: Codigestión con residuo de pera. La producción acumulada de metano aumentó al añadir como cosubstrato residuo de pera, tanto en el rango termofílico como en el mesofílico. Las producciones máximas se obtuvieron en el rango mesofílico. Los mejores resultados, dentro del rango mesofílico, se obtuvieron para el tratamiento con un 12,5% en peso de residuo de pera, aumentando no sólo la producción acumulada de metano respecto a la del purín, sino también el índice de producción de metano respecto a sólidos volátiles. La digestión del residuo de pera por sí sólo no es viable en las condiciones utilizadas, debido a su baja alcalinidad. La adición de más de un 12,5% de residuo de pera en peso añadido al purín, provoca un desfase en el inicio de la producción de metano, provocando un efecto similar a una sobrecarga orgánica en un sistema en continuo. La duración del desfase aumentó con la proporción de cosubstrato añadido. En el rango termofílico, la producción acumulada de metano aumentó conforme aumentó la proporción de cosubstrato, aunque el aprovechamiento de la materia orgánica añadida fue menor, como demuestra el menor índice de producción de metano respecto a sólidos volátiles iniciales. El principal efecto de la adición de residuo de pera fue la bajada del pH y por tanto la reducción de la inhibición por amonio. Al aumentar el pH por consumo de AGV se vuelven a restablecer las condiciones de inhibición, motivo por el cual no se alcanzaron valores tan altos como en mesofílico. En el rango termofílico no se detectó desfase en la producción de gas, indicando que los organismos termofílicos son más resistentes a las sobrecargas orgánicas. 212 Ensayos de viabilidad de codigestión en discontinuo 5.6.4. Del experimento 3. Adición de tierras decolorantes de aceite de oliva. La producción de metano acumulada y respecto a sólidos volátiles iniciales en el rango mesofílico aumenta al añadir TDO como cosubstrato. En el caso de un 5% de TDO la producción acumulada fue de 25,78 mL CH4/g substrato, frente a 8,86 mL CH4/g purín, es decir casi triple. El tratamiento con un 5% de TDO obtuvo la producción máxima de metano, no sólo de este experimento, sino de todos los experimentos realizados. La velocidad de producción de metano fue también la máxima. La digestión con tierras decolorantes sin mezclar con purín no es viable, resultando en producciones más bajas que el control, debido a la baja alcalinidad y altas concentraciones de ácidos grasos de cadena larga. Los ácidos grasos de cadena larga inhiben el proceso anaerobio tanto en termofílico como en mesofílico. En termofílico se inhibió completamente con una concentración aproximada de 7,8 g/L de trioleato, sin que fuera capaz de recuperarse. En el rango mesofílico el principal efecto observado fue la aparición de un desfase en la producción de metano, que aumentó con la proporción de TDO, y por tanto con la concentración de oleico. 5.6.5. Resumen de conclusiones La digestión de purín es posible tanto en termofílico como en mesofílico, aunque en el primer rango de temperatura está claramente inhibido por amonio, obteniéndose menores producciones de gas que en mesofílico, y observándose un aumento en la producción de gas con el grado de dilución. La codigestión de purín con otros substratos es una buena opción para aumentar la producción de metano. La digestión anaerobia de residuo de pera y de tierras decolorantes de aceite de oliva, sin mezclar con otro substrato, no ha resultado viable. El principal efecto de la adición de residuo de pera fue el mayor aporte de materia orgánica, mientras que el índice de producción respecto a sólidos volátiles mostró sólo una pequeña mejora. Se observó un efecto de sobrecarga orgánica en el rango mesofílico que hizo aumentar la duración de la fase lag. La codigestión de purín con tierras decolorantes de aceite de oliva mostró un buen comportamiento en el rango mesofílico, con una producción de metano respecto a sólidos volátiles 2,4 veces mayor que con purín sólo. Sin embargo, la adición de TDO en el rango termofílico inhibió completamente la producción de metano. En general, la digestión termofílica ha resultado menos viable que la Optimización de la digestión anaerobia de purines de cerdo 213 mesofílica debido a la acción de dos tóxicos diferentes, que potencian su poder inhibidor a altas temperaturas: amonio y ácidos grasos de cadena larga, especialmente oleico. La adaptación del inóculo es importante para el desarrollo del proceso. Aunque la concentración de inóculo utilizada en el ensayo ha sido baja, la mayoría de los ensayos pudieron llevarse a cabo, proporcionando información de respuesta a sobrecargas, y manteniendo condiciones en la composición del substrato reales. Los ensayos en discontinuo proporcionan, al menos, una aproximación cualitativa a la viabilidad del proceso anaerobio con mezclas de residuos, aún en el caso de desconocer su composición exacta, o la existencia de algún inhibidor del proceso metanogénico. El empleo de este tipo de test no es incompatible, sino complementario con la realización de tests de biodegradabilidad y toxicidad. 214 Ensayos de viabilidad de codigestión en discontinuo 6. ENSAYOS EN CONTINUO DE CODIGESTIÓN DE PURÍN Y TIERRAS DECOLORANTES DE ACEITE DE OLIVA. 216 Ensayos de viabilidad de codigestión en continuo Optimización de la digestión anaerobia de purines de cerdo 217 6.1. Resumen La digestión anaerobia con purines de cerdo es poco viable económicamente, debido a las bajas producciones de biogás obtenidas, teniendo como principales limitaciones la baja biodegradabilidad, comparada con otros tipos de residuos, y el alto contenido en amonio, importante inhibidor del proceso anaerobio. La codigestión con pequeñas cantidades de residuos de la industria alimentaria, que presentan altos contenidos en materia orgánica fácilmente degradable, puede ser una opción para mejorar la eficiencia del proceso y su viabilidad económica. Partiendo de los resultados obtenidos en ensayos previos en discontinuo, se pusieron en marcha cuatro reactores anaerobios en régimen semicontinuo, de 5 litros de capacidad, dos de ellos trabajando en el rango mesofílico (35ºC ) y otros dos en termofílico (55ºC). Se experimentaron dos metodologías para la fase de puesta en marcha, utilizando dos tipos de inóculo, lodo digerido de EDAR (mesofílico) y purín digerido (procedente de un reactor tipo flujo-pistón), que mostraron resultados similares, pudiendo concluir que el lodo de EDAR digerido en mesofílico puede ser utilizado para la puesta en marcha de reactores alimentados con purín de cerdo, tanto en mesofílico como en termofílico. Una vez superada la fase de puesta en marcha se comenzó la experimentación, comparando los resultados obtenidos en los reactores alimentados con purín, con los resultados de los reactores alimentados con la mezcla de purín y tierras decolorantes de aceite de oliva (TDO), en diferentes proporciones. En el rango mesofílico los reactores que se alimentaron con TDO mejoraron ampliamente los resultados obtenidos con purín sólo, aunque en parte se debió al aumento de la carga. A pesar de la mejora en la producción de gas, se observó acumulación de AGV, indicando la inhibición parcial del sistema. En el rango termofílico, los resultados fueron en todos los casos peores que en mesofílico, con fuerte acumulación de AGV, especialmente acético, mostrando que el sistema estaba fuertemente inhibido por las altas concentraciones de amonio. 218 Ensayos de viabilidad de codigestión en continuo 6.2. Introducción. 6.2.1. Ensayos en régimen semicontinuo a escala laboratorio. Los ensayos en continuo permiten evaluar distintos aspectos, tales como la viabilidad de la utilización de un cosubstrato, necesidad de recirculación, efecto de pretratamientos, etc. Este tipo de ensayos permite evaluar la capacidad de adaptación de la biota anaerobia a un determinado substrato, la respuesta a sobrecargas orgánicas, a la presencia de tóxicos o a cambios de temperatura. Los sistemas piloto de laboratorio permiten, además, seleccionar parámetros de operación, como velocidad de carga orgánica, tiempo de retención hidráulica, necesidad de recirculación, velocidad de agitación y diferentes estrategias de puesta en marcha (Lema, 1995). La tecnología de mezcla completa, CSTR (Continuous Stirred Tank Reactor), es la tecnología más clásica para el tratamiento de residuos semisólidos (de 10 a 200 mg ST/L), y está caracterizada por la agitación del substrato, continua o intermitente (Muñoz Valero et al., 1987). La mayoría de los autores de la bibliografía consultada utiliza este tipo de reactores para tratar residuos ganaderos, y la mayoría de las plantas a escala industrial que actualmente están funcionando se basan en este sistema. 6.2.2. Importancia de la puesta en marcha. Riesgo de sobrecargas La puesta en marcha de un reactor anaerobio, generalmente, requiere la adicción inicial de los microorganismos necesarios para llevar a cabo el proceso. La alimentación inicial de substrato se suele realizar a baja velocidad de carga para asegurar el desarrollo estable de la biomasa del digestor. La velocidad de puesta en marcha dependerá de las características del inóculo, del substrato alimentado, de la presencia de tóxicos, del tamaño y configuración del reactor y de las condiciones de operación (Fannin, 1987). El uso de un inóculo, procedente de un reactor que trabaje con el mismo tipo de substrato, implica un cierto grado de adaptación al tipo de substrato y a los posibles tóxicos. Sin embargo, no siempre es posible disponer de ese tipo de inóculo. En los ensayos de codigestión, en los que se trabaja con mezclas completamente nuevas de diferentes tipos de residuos, es muy difícil conseguir un inóculo adaptado. No se conoce ningún digestor, en el área de trabajo, que actualmente trabaje en el rango termofílico, por lo que necesariamente se ha de plantear el cambio de condiciones de temperatura, para la puesta en marcha de un reactor termofílico. Un cambio brusco de temperatura puede afectar muy negativamente al proceso, y el grado de afectación o la velocidad de Optimización de la digestión anaerobia de purines de cerdo 219 recuperación depende en gran medida de la rapidez con que se efectúen los cambios de temperatura (van Lier et al., 1993). En cualquier caso, y de acuerdo con el mismo autor, en un plazo relativamente corto (20 días), la producción de metano se recupera. La puesta en marcha de reactores anaerobios termofílicos utilizando inóculo mesofílico ha resultado viable en experiencias previas (Griffin et al., 1998). La inestabilidad del sistema anaerobio se puede producir por varios factores, que van desde un cambio brusco en la temperatura o en la carga orgánica, así como cualquier substancia o actividad que produzca la inhibición de microorganismos (Fannin, 1987). El riesgo de las sobrecargas orgánicas es una amenaza siempre presente en los reactores anaerobios, y especialmente en la fase de puesta en marcha. La principal consecuencia de las sobrecargas es la acidificación del reactor, que puede llegar a ser irreversible. Afortunadamente, los residuos ganaderos presentan elevados niveles de alcalinidad, por lo que el riesgo de acidificación por sobrecarga es mucho menor que con otros tipos de substratos. Diversos autores han estudiado los mejores parámetros de diagnóstico para detectar una sobrecarga, sin que esté claro cual es el parámetro de más rápida respuesta. La relación propiónico/acético se ha propuesto como un rápido y eficiente parámetro de diagnóstico (Marchaim et al., 1993). Sin embargo, Griffin et al. (1998) proponen como parámetro de diagnóstico la concentración de propiónico, independientemente de la de acético, y Ahring et al. (1995) concluyen que el parámetro de diagnóstico más rápido es la concentración de butírico, especialmente el isómero iso, muy por encima de la relación propiónico/acético o la de éstos ácidos por separado. Por otro lado, considerar únicamente la relación acético/propiónico puede enmascarar el problema en el caso de la existencia de fenómenos de inhibición de la metanogénesis acetoclástica, caso del amonio. Otros autores han propuesto parámetros de más fácil medida, pero de mucho menor valor como diagnóstico, como el pH o la relación de alcalinidad, que si bien pueden dar una idea de la existencia de un problema, no son exclusivos de un problema de sobrecarga y además, pueden ser de muy lenta reacción, sobre todo en residuos con alta alcalinidad (Iza, 1995). La temperatura de trabajo puede condicionar la estabilidad del reactor, considerándose, en general, que las altas temperaturas aumentan las tasas de conversión biológicas, pero provocan una mayor inestabilidad del proceso (Fannin, 1987). Otros trabajos más recientes contradicen esta teoría, afirmando que la respuesta a las altas cargas o sobrecargas es diferente en función de la temperatura de trabajo, siendo más sensibles los 220 Ensayos de viabilidad de codigestión en continuo microorganismos mesofílicos que los termofílicos (Griffin et al., 1998). La causa de la discrepancia puede estar en el tipo substrato utilizado y en la presencia de tóxicos o inhibidores, que pueden variar su grado de inhibición en función de la temperatura. La puesta en marcha de un reactor de mezcla completa puede realizarse siguiendo diversas estrategias. Generalmente, se comienza con una baja velocidad de carga orgánica, aumentándola gradualmente (Fannin, 1987), mientras que otras más arriesgadas proponen comenzar con altos niveles de carga. Es recomendable no provocar grandes cambios de las condiciones en las que originalmente trabajaba el inóculo, por lo que se propone mantener, al menos inicialmente, las condiciones de velocidad de carga, sobre todo si se cambia la temperatura. La puesta en marcha de un reactor que trate substratos con algún tóxico, como el caso de los purines de cerdo por su alto contenido en amonio, resulta especialmente complicada, ya que a las condiciones siempre estresantes de puesta en marcha e incluso cambio de temperatura, se unirá la presencia de un inhibidor del crecimiento de los microorganismos. Van Velsen (1979) comprobó que la puesta en marcha de reactores continuos de mezcla completa, trabajando con purines de cerdo, es posible utilizando como inóculo lodos de depuradora, obteniendo buenos resultados con un nivel de carga inicial relativamente alto (0,125 kg DQO/kg SV*día), que en sus condiciones equivale a una velocidad de carga orgánica de 2,7 kg de ST/m3 y día. La duración del período de aclimatación es mayor cuanto mayor es el nivel de carga orgánica inicial. La mayoría de los autores que han trabajado con residuos ganaderos han utilizado valores de carga orgánica del orden de 2,5-3 g SV/L reactor*día (van Velsen, 1979; Robbins et al., 1989; Angelidaki y Ahring, 1993b, Angelidaki y Ahring, 1994; Ahring et al., 1995, Hansen et al., 1998, Hansen et al., 1999; etc.), aunque en algunos trabajos se han utilizado mayores velocidades de carga, con buenos resultados (Hashimoto et al., 1986; Hill et al., 1987). 6.2.3. Digestión anaerobia de residuos ganaderos. A pesar del alto contenido en materia orgánica de los residuos ganaderos los índices de producción de metano son relativamente bajos (Tabla 6.1), indicando la baja biodegradabilidad de los mismos. Optimización de la digestión anaerobia de purines de cerdo 221 Tabla 6.1. Revisión bibliográfica de producción de metano sobre sólidos volátiles (Pc), en L CH4/g SV, de residuos ganaderos en función de la temperatura (Tª), velocidad de carga orgánica (VCO)en g SV/L·d, tiempo de retención (TR), en días y nitrógeno amoniacal, en g N/L Referencia Angelidaki y Ahring, 1993 Angelidaki y Ahring, 1993 Angelidaki y Ahring, 1993 Angelidaki y Ahring, 1993 Angelidaki y Ahring, 1993 Angelidaki y Ahring, 1993 Angelidaki y Ahring, 1993 Chen and Day (1985) Chen and Day (1985) Chen and Day (1985) Fischer et al., 1981 Hansen et al., 1998 Hansen et al., 1998 Hansen et al., 1998 Hansen et al., 1998 Hansen et al., 1999 Hansen et al., 1999 Hashimoto , 1986 Hashimoto , 1986 Hashimoto , 1986 Hashimoto , 1986 Hashimoto , 1986 Hashimoto , 1986 Hashimoto , 1986 Hashimoto , 1986 Hashimoto , 1986 Hashimoto , 1986 Hashimoto , 1986 Hill et al., 1987 Hill et al., 1987 Hill et al., 1987 Hill et al., 1987 Robbins et al., 1989 Robbins et al., 1989 Robbins et al., 1989 Robbins et al., 1989 Van Velsen, 1979 Van Velsen, 1979 Van Velsen, 1979 Van Velsen, 1979 Substrato Estiércol bovino Estiércol bovino Estiércol bovino Estiércol bovino Estiércol bovino Estiércol bovino Estiércol bovino Purín de cerdo Purín de cerdo Purín de cerdo Purín de cerdo Purín de cerdo Purín de cerdo Purín de cerdo Purín de cerdo Purín de cerdo Purín de cerdo Estiércol bovino Estiércol bovino Estiércol bovino Estiércol bovino Estiércol bovino Estiércol bovino Estiércol bovino Estiércol bovino Estiércol bovino Estiércol bovino Estiércol bovino Purín de cerdo Purín de cerdo Purín de cerdo Purín de cerdo Estiércol bovino Estiércol bovino Estiércol bovino Estiércol bovino Purín de cerdo Purín de cerdo Purín de cerdo Purín de cerdo Tª 55 55 55 55 55 55 55 50 55 60 35 37 45 55 60 37 55 35 35 35 35 35 35 55 55 55 55 55 35 35 35 35 37 37 37 37 30 30 30 30 VCO TR 3.00 15 3.00 15 3.00 15 2.80 15 2.80 15 2.80 15 2.80 15 3.00 3.00 3.00 3.00 3.0 3.0 4.44 4.44 4.44 4.44 4.44 4.44 7.76 8 8 8 8 3.56 5.71 6.86 8.03 2.625 2.625 2.625 2.625 4 4 2.7 2.7 15 15 15 15 15 15 9 9 9 9 9 9 5 5 5 5 5 10 10 10 10 16 16 16 16 15 15 15 15 NH4+ 1.5 4 6 2.5 3 5 6 Pc 0.2 0.05 0.05 0.2 0.2 0.15 0.15 0.13-0.38 0.26-0.36 0.2-0.27 0.06-0.30 5.9 0.19 6 0.14 6 0.07 6.1 0.02 5.9 0.19 6 0.07 1.04 0.49 2.64 0.54 4.33 0.40 6.08 0.51 0.09 7.74 9.12 0.40 0.97 0.30 4.36 0.30 4.38 0.30 5.4 0.29 5.37 0.28 0.30 0.26 0.28 0.02 0.7 0.20 4.2 0.16 1.4 0.21 2.8 0.08 2.675 0.32 2.75 0.33 2.68 0.32 2.75 0.33 222 Ensayos de viabilidad de codigestión en continuo En la Tabla 6.1 se observa una gran variabilidad, debido a las diferencias en los parámetros de operación, temperatura, concentración de amonio, tiempo de retención, carga orgánica, etc. La producción de metano por unidad de carga (Pc), va desde 0,02 a 0,54 L de CH4/g SV, y para purines de cerdo de 0,02 a 0,38 L de CH4/g SV, es decir menores que para otros tipos de residuos ganaderos. 6.2.4. Codigestión con tierras decolorantes de aceites vegetales La codigestión de residuos ganaderos con otros tipos de residuos, especialmente procedentes de la industria agroalimentaria, puede proporcionar mayores producciones de gas (capítulo 5). Los buenos resultados obtenidos se atribuyeron al alto potencial de producción de metano de los restos de aceite del cosubstrato utilizado (alrededor del 27% sobre materia seca). El efecto de la adicción de arcillas (bentonita) u otros materiales inertes de alta superficie específica (carbón activo o glauconita), parece influir positivamente en la digestión anaerobia, ya que parece disminuir del efecto tóxico de determinados compuestos, como amonio, sulfuros o metales pesados (Angelidaki y Ahring, 1993b; Hansen et al., 1999). Las formas aparentemente tóxicas son los ácidos libres (Angelidaki y Ahring, 1992). El efecto inhibidor parece estar atenuado si se encuentran ligados a alguna superficie adsorbente, caso de la bentonita con restos de aceite (bentonite bound oil-BBO, Angelidaki et al., 1990), y por extensión, el de las tierras decolorantes de aceite de oliva (TDO). Parece ser que el efecto tóxico depende directamente del área superficial de los gránulos de biomasa, de forma que el lodo suspendido o floculento de los sistemas de mezcla completa es mucho más sensible a la inhibición por AGCL que el lodo granular (Hwu et al., 1996) y más en el rango termofílico que en el mesofílico (Hwu et al., 1997). La codigestión de residuos ganaderos, principalmente bovino, a 55ºC, con bentonita con restos de aceite (BBO), a una proporción del 2%, mejora ligeramente la producción de metano por unidad de sólidos volátiles añadidos y sobre todo parece contrarrestar el efecto inhibidor del nitrógeno amoniacal (Angelidaki y Ahring, 1993). Sin embargo, estudios posteriores, utilizando mezclas de purines de cerdo y un 5% de BBO, obtuvieron una producción de metano peor que el control, sólo con purin (Hansen et al., 1999). Resultados similares se encontraron en el presente trabajo, en los ensayos realizados en régimen discontinuo (capítulo 5), con un nivel de producción de metano muy bajo para el rango termofílico, atribuido al efecto Optimización de la digestión anaerobia de purines de cerdo 223 inhibidor de los ácidos grasos de cadena larga. La influencia del período de puesta en marcha al trabajar con este tipo de substratos, potencialmente inhibidor, es más limitante que en condiciones normales. El hecho de que las formas tóxicas sean los ácidos libres implica la posibilidad de cierto grado de adaptación a la presencia de lípidos. Dicho proceso de adaptación implica el desarrollo de la población acetogénica capaz de degradar los AGCL, que son liberados por hidrólisis de las grasas neutras, a una velocidad adecuada para evitar su acumulación, lo que implica que la introducción en una planta de DA debe ser gradual (Angelidaki y Ahring, 1992). 6.3. Objetivos Los objetivos planteados con la serie de experimentos en continuo se resumen en los siguientes puntos: 1. Estudiar el efecto sobre los diferentes parámetros del proceso de la introducción de tierras decolorantes de aceite de oliva. Comparar el efecto del cosubstrato en función de la temperatura. 2. Estudiar la viabilidad de la digestión anaerobia de purines de cerdo con alto contenido en nitrógeno a escala piloto laboratorio. Estudiar el efecto de la temperatura y la inhibición por amonio en purines de cerdo. Contrastar los resultados obtenidos en ensayos en discontinuo. 3. Estudio de diferentes estrategias de puesta en marcha. Evaluar la influencia de los cambios en la composición del influente. Evaluar la bondad de los parámetros de diagnóstico propuestos en la bibliografía para el estudio de sobrecargas. 4. Validar la metodología empleada en los ensayos en discontinuo, estudiando su posible extrapolación en continuo. 6.4. Materiales y métodos. 6.4.1. Dispositivo experimental Se diseñó y construyó una planta piloto escala laboratorio de 5 L de volumen útil, en régimen semicontinuo, que cuenta con cuatro unidades completas que trabajan de forma independiente, pudiendo mantener diferentes condiciones de operación, como temperatura, velocidad de carga, velocidad 224 Ensayos de viabilidad de codigestión en continuo de agitación, etc. (Flotats et al., 1998). El esquema de cada unidad se muestra en la Figura 6.1. Residuo Temporización Registro datos PC 4 Biogas 1 4 C.G. Biogas Efluente 2 3 Sistema Calefacción Residuo tratado Control Temperatura Bomba alimentación Punto muestreo gas Punto muestreo residuo Sonda Temperatura Circuito agua calefacción Circuito substrato Circuito gas Sistemas de control 1 2 3 4 Depósito de influente Reactor Depósito desgasificación Sistemas de agitación Figura 6.1. Esquema de cada una de las cuatro unidades del banco de ensayos en continuo. Cada unidad se compone de: Depósito de influente de vidrio, dos litros de capacidad, con salida horizontal de 2,5 cm de diámetro interior, por la parte inferior. Cuenta con una agitador de varilla metálica, acoplado a la tapa del depósito, sincronizado con la bomba de impulsión (Figura 6.2). Se situó a una cota superior que la bomba para facilitar la impulsión. Bomba de alimentación de paletas que proporciona pequeños caudales, gracias al pequeño tamaño de la bomba y a la baja velocidad de giro del rotor (variador de velocidad). La potencia y velocidad de giro del motor son 0,12 H.P. y 1350 rpm, respectivamente. Periódicamente se procedió a la sustitución del cabezal de bombeo, debido a la corrosión de las paletas de caucho. El tubo que conecta la botella de influente y la bomba fue un tubo de plástico con refuerzo metálico de 2 cm de diámetro interior. El mismo Optimización de la digestión anaerobia de purines de cerdo 225 tubo de utilizó par conectar la bomba con el orificio de entrada al reactor. Hubo problemas de obturación del tubo que va desde la botella de influente a la bomba, lo que obligó a su revisión y limpiado diario, e hizo que en el inicio de los experimentos no se consiguiera mantener un tiempo de retención uniforme. En la Figura 5.2 se muestra una imagen del sistema agitador-bomba. Agitador influente Bomba de alimentación Botella de influente Figura 6.2. Vista de la botella de influente y la bomba de impulsión. Depósito de efluente-desgasificación de vidrio de dos litros de capacidad, con una entrada común de gas y substrato, y tres salidas, una inferior para el vaciado, otra superior de salida del gas y un orificio con tapón y septum, previsto para la toma de muestra de gas, para facilitar el vaciado de la botella y la limpieza de la misma. Se construyó sobre un matraz de vidrio de 2 L, de forma esférica para facilitar la salida del efluente, sin que queden sólidos depositados en el fondo de la botella (Figura 6.3). Entre el reactor y el depósito de efluente se dispuso de una válvula de bola manual, para aislar el reactor de la botella de efluente, de utilidad en el momento de vaciado de la botella de efluente. Medidor de gas. Para medir la producción de gas se utilizó un contador volumétrico de gas húmedo, que consiste en un sistema de vasos comunicantes, basado en un diseño de Mata-Álvarez et al. (1986). Consiste en dos tubos comunicados por la base, conteniendo una solución de agua destilada y negro de ericromo (líquido negro y opaco). Utiliza, como sistema de medida, un sistema de emisión y recepción de luz, mediante una bombilla y una célula fotoeléctrica. Cuando la célula no percibe luz debido a que el líquido opaco intercepta el haz luminoso, manda una señal eléctrica al sistema de adquisición de datos. Cuando llega al punto máximo se 226 Ensayos de viabilidad de codigestión en continuo reequilibran las presiones y se recupera el nivel inicial, de forma que comienza un nuevo ciclo. Cada ciclo equivale a un determinado volumen, que deberá ser estimado. El sistema de adquisición de datos está conectado a un ordenador personal, con un software capaz de traducir la señal eléctrica en un valor numérico. El software proporciona datos horarios para cada reactor, siendo capaz de hacer gráficas y resúmenes diarios (García Gispert, 1998). En las Figura 6.4 y Figura 6.5 se muestra un esquema de funcionamiento del gasómetro y una fotografía de una unidad funcionando. Salida de gas hacia el gasómetro Despresurizaciónmuestreo gas Válvula de conexión reactor-botella efluente Entrada de efluente procedente del reactor Salida de efluente. Punto de muestreo de efluente. Figura 6.3. Vista detalle de depósito de efluente- desgasificador Figura 6.4. Esquema de funcionamiento del contador de gas. Periódicamente se procedió al calibrado del contador. El volumen de gas varía en función del tamaño exacto de cada contador y, sobre todo, del nivel de llenado, estando en torno a los 100 mL de gas húmedo, a presión atmosférica. La utilización de este sistema, si bien es muy interesante por la comodidad y la gran cantidad de datos registrados, presentó diversos problemas que hicieron que se retrasara bastante la puesta en marcha, tales Optimización de la digestión anaerobia de purines de cerdo 227 como interferencias con otros aparatos eléctricos. Evacuación del gas producido Entrada de gas procedente del desgasificador Fuente de luz Célula fotoeléctrica Figura 6.5. Vista del contador de gas Reactor. Tiene forma cilíndrica y un volumen útil de 4,95 litros. Las paredes verticales son de vidrio y las tapas superior e inferior son de acero inoxidable. Cuenta con una entrada de influente y una única salida de efluente y gas por la parte superior. El líquido sale cuando dentro del reactor hay una sobrepresión suficiente en el espacio de cabeza para superar la altura del tubo de salida. Dispone de un punto de muestreo de gas. Está envuelto por una camisa por la que circula agua caliente y permite controlar la temperatura del reactor mediante una sonda conectada a un termostato. La homogeneización se realiza mediante un agitador mecánico que permite regular la velocidad de giro desde 0 a 1000 rpm. El sistema de acoplamiento del agitador al reactor permite, mediante sello mecánico, la correcta agitación manteniendo perfectamente estanco el reactor (Figura 6.6). Sistema de calefacción. El sistema de calefacción consiste en un calentador de agua eléctrico, con dos entradas de agua, una para el retorno del circuito y otra conectada a la red para la reposición de las pérdidas. La circulación del agua por el circuito se realizó mediante una bomba. Se dispusieron dos sistemas de control de la temperatura del depósito general, uno externo, en el inicio del circuito y uno dentro del propio depósito, que actuó como termostato de seguridad. En la entrada de la red se situó un sistema de control de presión. La temperatura a la que se calentó el agua fue de 60ºC, válida tanto para los reactores en termofílico (55ºC), como en mesofílico (35ºC). La calefacción de cada reactor se realiza por circulación de agua caliente por la camisa que rodea el cuerpo del reactor. El control de la temperatura se hace por medio de una sonda de temperatura conectada a un termostato, de forma que al bajar la temperatura por debajo de la marcada, se pone en marcha la bomba de circulación y se abre una válvula motorizada, 228 Ensayos de viabilidad de codigestión en continuo abriendo el circuito para ese reactor. Todas los tubos fijos de la instalación fueron convenientemente aislados, para evitar pérdidas de calor. Un esquema del funcionamiento se muestra en la Figura 6.7. Sistema agitación Alimentación reactor Punto muestreo de gas Salida circuito de calefacción Entrada circuito de calefacción Salida efluente Sonda Temperatura Camisa calefacción Purga Figura 6.6. Esquema y fotografía del reactor ManómetroRegulador presión Alimentación red Calentador eléctrico Termostato Válvula motorizada Válvula manual Bomba de circulación Circuito de agua caliente Circuito de retorno Control electrónico R1 R2 R3 Figura 6.7. Esquema del sistema de calefacción. R4 Optimización de la digestión anaerobia de purines de cerdo 229 6.4.2. Métodos analíticos Se utilizaron los métodos analíticos expuestos en el capítulo 3. 6.4.3. Operación de los reactores Diariamente se recogió el volumen de efluente y se recargó el depósito de influente. Tres veces por semana se analizó la composición del gas del espacio de cabeza y algunos parámetros básicos del efluente, como pH y ácidos grasos volátiles. Semanalmente se realizó una caracterización tanto del efluente como del influente, midiendo sólidos totales y volátiles, Demanda Química de Oxígeno (DQO) total y soluble, Nitrógeno Kjeldahl y Nitrógeno amoniacal y alcalinidad. La velocidad de agitación es un parámetro que influye en el desarrollo del proceso, siendo necesario un equilibrio entre una buena homogeneización y la correcta formación de agregados bacterianos (Fannin, 1987). La mayoría de los autores utilizan ciclos alternativos de agitación y parada y velocidades muy bajas que van de 60 a 150 rpm, aunque también hay trabajos que utilizan velocidades más altas, entre 400 y 600 rpm (Marchaim et al., 1993). La velocidad de agitación es un parámetro que por sí sólo no determina la buena homogeneización del substrato, ya que depende de la forma del elemento agitador. En el presente trabajo se fijó constate, en 200 rpm, sin embargo durante el desarrollo del experimento se observó que era insuficiente para la correcta homogeneización del reactor, produciéndose acumulación de sólidos en el reactor, por lo que se aumentó a 500 rpm. 6.4.4. Cálculo de parámetros A partir de los valores medidos de influente, efluente y producción de gas se estimaron varios parámetros que se detallan a continuación: Tiempo de retención. Aunque es un parámetro de diseño se comprobó diariamente el valor real obtenido, mediante la ecuación 6.1. El tiempo de retención (TR) hidráulico es igual al tiempo de retención celular por tratarse de un sistema de mezcla completa sin recirculación. Q es el volumen diario de alimentación y VR es el volumen del reactor. TR VR . Q (6.1) Velocidad de carga orgánica (VCO). Se calcula a partir del TR y de la concentración de materia orgánica en el influente, que se expresó tanto en unidades de sólidos volátiles como de DQO: 230 Ensayos de viabilidad de codigestión en continuo VCO Q x >C @ VR >C @ , TR (6.2) dónde [C] es la concentración de SV o DQO. Eliminación de materia orgánica. Se estima mediante la diferencia entre la concentración del influente media para cada período considerado (Tabla 6.4 y Tabla 6.5), y la concentración media del efluente, mediante: EMO 1 >C @ef . , >C @inf (6.3) EMO se calcula para ST, SV y DQO. Biogás, CH4 y CO2. El volumen de biogás producido se midió directamente con el contador de gas. A partir de la producción horaria de biogás y de la composición del mismo, se calculó la producción diaria de CH4 y CO2. Al tomar la muestra de gas hay una pequeña contaminación por aire, que se refleja en la aparición en el cromatograma de un pico de N2. Para el cálculo del metano producido se eliminó este efecto, considerando que todo el biogás está formado por metano y dióxido de carbono, expresando la concentración en porcentaje sobre el total. Tanto el biogás como el metano, se expresan en unidades de gas húmedo, ya que no se ha considerado la contribución del vapor de agua. El volumen se refiere, salvo indicación expresa, al volumen a temperatura ambiente, aproximadamente 20ºC. Para pasarlo a condiciones estándar, presión atmosférica y a 0º C de temperatura, se deberá emplear la siguiente expresión: V0ª C V20º C x 273 273 20 (6.4) Los diferentes parámetros de producción de gas considerados son: - Producción de gas, P (L gas/día). Es la medida directa de gas por unidad de tiempo. La producción de metano se calcula como el producto de la producción de biogás por el contenido de metano en el mismo. - Producción volumétrica, PV (L gas/L reactor*día). Es la producción por unidad de volumen de reactor y día, PV P . VR (6.5) - Producción específica o por unidad de carga orgánica, Pc (L gas/g de Mat. Org. Del influente): Es la producción de gas, biogás, CH4 o CO2, por unidad de carga orgánica (SV, o DQO), Optimización de la digestión anaerobia de purines de cerdo PC P VR x VCO PV VCO 231 (6.6 ) - Producción por unidad de masa de influente, PM (L gas/kg de substrato): Es la producción de biogás, CH4 o CO2, por unidad de masa tratada PM P x TR VR PV x TR (6.7 ) Índice de Metanización, M (%). Es un índice que estima la fracción de la materia orgánica introducida en el reactor convertida en metano. Es de especial interés para comparar con el índice de acidificación, A(%), para estimar la fase limitante del proceso. Se calcula expresando la producción de metano en unidades de DQO, y relacionándolo con la carga orgánica, en las mismas unidades, y está basado en el método propuesto en ensayos de biodegradabilidad por Field et al. (1988). M (%) P x 2,49 x 100 VCO x VR Pv x 2,49 x TR x 100 . DQOin (6.8) Índice de acidificación, A (%). Es la fracción de la DQO inicial transformada en ácidos grasos durante el proceso. Se considera que la materia orgánica, transformada en metano, ha pasado previamente por la fase de acidificación. Por ello, el índice de acidificación total es la suma de la fracción en forma de AGV, Vef, más la metanización, M. La concentración de AGV se expresa en unidades de DQO, utilizando los correspondientes coeficientes de transformación (Field et al., 1988), ¦ AGVef DQOin A(%) Vef (%) M (%) . (6.9) ¦ AGVef Pv x 2,49 x TR Pv x 2,49 x TR x 100 x 100 x 100 DQOin DQOin Índice de acidificación neta, AN (%). Se calcula como la acidificación total menos la fracción de DQO del influente correspondiente a los AGV contenidos en el mismo. Los substratos orgánicos son susceptibles de degradarse parcialmente durante el período de conservación de muestras, o incluso en el período previo al muestreo. Por ello, la hidrólisis-acidificación que realmente ocurre durante el proceso, es la acidificación neta, AN (%) AN (%) Vef (%) M (%) Vin (%) ¦ AGVef Pv x 2,49 x TR ¦ AGVin DQOin x 100 . (6.10) 232 Ensayos de viabilidad de codigestión en continuo 6.4.5. Programación de los ensayos. Se utilizaron cuatro reactores en continuo (reactor 1 a 4), con diferentes condiciones de operación (Tabla 6.2), para comparar el rango termofílico (reactor1 y 4) con el mesofílico (reactor 2 y 3). Fase de puesta en marcha Se utilizaron dos estrategias diferentes, tal y como se muestra a continuación: Estrategia 1. Los reactores 1 y 2 se iniciaron con una proporción de inóculo de 1/5 del volumen del reactor, 3/5 de substrato y 1/5 de agua. Se puso a digerir, con agitación y a la temperatura definitiva, en modo discontinuo hasta conseguir una producción de gas estable, momento a partir del cual se comenzó a alimentar con purín. La velocidad de carga orgánica, junto con otros parámetros, se fue incrementando gradualmente. El substrato utilizado siempre fue purín, más o menos diluido, para asegurar un nivel de amonio por debajo del umbral de inhibición. Se consideraron cuatro períodos diferentes, en los que se fue disminuyendo gradualmente el grado de dilución del purín utilizado. La duración total estuvo entre 188 días, para el reactor 1, y 125 días, para el reactor 4, hasta que se consideró el inicio de los experimentos. Tabla 6.2. Cronología de los cuatro reactores continuos utilizados Temperatura (ºC) Inóculo utilizado Fecha de inicio de la puesta en marcha Fecha de inicio de ensayos Fecha de inicio del estudio con TDO Fecha final del estudio Tiempo de retención hidráulico (días) Reactor 1 Reactor 2 Reactor 3 Reactor 4 Termofílico Mesofílico Mesofílico Termofílico (55ºC) (35ºC) (35ºC) (55ºC) Purín digerido Purín digerido Lodos EDAR Lodos EDAR mesofílico mesofílico digeridos 35º digeridos 35º 25/01/99 04/02/99 09/03/99 29/03/99 1/08/99 1/08/99 1/08/99 1/08/99 - - 17/08/99 17/08/99 22/10/99 22/10/99 22/10/99 22/10/99 15 15 20 hasta 29/7; 20 hasta 29/7; 15 hasta final 15 hasta final Estrategia 2. En los reactores 3 y 4 se utilizó como inóculo lodos de depuradora digeridos, comenzando con el régimen continuo desde el primer día. La alimentación fue variando, comenzando con una mezcla de lodos de Optimización de la digestión anaerobia de purines de cerdo 233 EDAR sin digerir y digeridos, a un tiempo de retención de 20 días y una carga orgánica de 1,84 g SV/L. Paulatinamente se fue substituyendo parte de los lodos por purín, hasta llegar al momento considerado como inicio del experimento, en que se trataba sólo de purín. La secuencia de cambios de la alimentación se muestra en la Figura 6.8. Unos días antes del inicio de la fase de experimentación se cambió el tiempo de retención de 20 a 15 días. 100% 80% 60% 40% 20% 0% -145 - -117 -116 - -99 Lodo digerido -98 - -69 -69 - -39 Lodo no digerido -39 - -20 -19 - 0 Purín Figura 6.8. Evolución de la composición de la alimentación de los reactores 3 y 4, durante la puesta en marcha. En la Figura 6.8 y en los apartados posteriores los días correspondientes al período de puesta en marcha, se indicarán en negativo y los de experimentación en positivo. El día 0 es el de inicio de los experimentos y final de la puesta en marcha. Fase de experimentación La fase de experimentación se considera desde el momento en que los cuatro reactores son alimentados exclusivamente con purín o con la mezcla de purín y el cosubstrato a un tiempo de retención de 15 días. La carga orgánica objetivo fue de 3,5 g SV/L reactor·d, aunque varió mucho debido a las variaciones en el substrato y en el tiempo de retención. La programación de los ensayos se muestra en la Tabla 6.3. 234 Ensayos de viabilidad de codigestión en continuo Tabla 6.3. Programación de los experimentos Días de ensayo 0-16 17-36 37-57 58-82 R1-55ºC Purín 100% Purín 100% Purín 100% Purín 100% R2-35ºC Purín 100% Purín 100% Purín 100% Purín 100% R3-35ºC Purín 100% TDO 1% TDO 2,5% TDO 5% R4-55ºC Purín 100% TDO 1% TDO 2,5% TDO 5% 6.4.6. Caracterización de la alimentación Se utilizó, durante la mayor parte del período de puesta en marcha y a lo largo de todo el experimento, una mezcla de purín procedente de cuatro granjas de diferente tipo, dos de ciclo cerrado, y otras dos de engorde, que se seleccionaron como representativas de las granjas de una zona de gran concentración ganadera (comarca de Les Garrigues). El purín se recogió una vez al mes, y se realizó una caracterización del influente una vez por semana. Se detectó una importante variabilidad tanto entre los diferentes muestreos en granja, como en el propio laboratorio, debido en el último caso a la dificultad para la correcta homogeneización. Inicialmente, en el reactor 1, se utilizó un purín diferente procedente de otra granja, de engorde, que tenía un menor contenido de nitrógeno. A partir del día –19, se utilizó el mismo purín para todos los reactores. En los reactores 3 y 4, a partir del día 17, comenzó la introducción de TDO como cosubstrato. La composición media del substrato para la fase de puesta en marcha y de experimentación se muestra en las Tabla 6.4 y Tabla 6.5, respectivamente. Optimización de la digestión anaerobia de purines de cerdo 235 Tabla 6.4. Caracterización del substrato utilizado durante la fase de puesta en marcha. Período -188 - -107 -106 - -69 -68 - -39 -38 – -20 -19 - 0 ST (g/kg) Reactor SV (g/kg) DQO (g/kg) Nk (g/kg) Reactores iniciados con purín de cerdo R1-control 55ºC 18,23 12,39 18,57 2,11 R2- control 35ºC 19,99 12,38 28,29 3,02 R1-control 55ºC 35,66 24,26 42,17 3,06 R2- control 35ºC 40,40 28,73 43,67 3,11 R1-control 55ºC 41,15 28,10 57,72 3,94 R2- control 35ºC 62,59 44,97 80,98 5,17 R1=R2 67,75 47,25 81,53 4,87 R1=R2 75,16 52,23 97,94 21,81 Reactores iniciados con lodos de depuradora -145- -117 R3 = R4 -116- -99 R3 = R4 -98- -69 R3 = R4 -68- -39 R3 = R4 -38- -20 R3 = R4 -19- 0 R3 = R4 =R1=R2 21,59 45,6 44,6 58,4 59,3 75,16 11,41 31,5 30,7 36,8 39,4 52,23 33,87 52,8 55,5 57,4 64,0 97,94 2,59 2,56 3,09 2,97 4,32 21,81 NH4+ (g/kg) TR (días) 1,74 2,36 2,08 2,21 2,76 3,62 3,53 5,62 18,37 22,23 15 15 15 15 15 15 1,21 0,83 0,99 1,19 2,84 5,62 20 20 20 20 20 20 Tabla 6.5. Caracterización de la alimentación para los diferentes períodos de experimentación Período Subst. NH4+ ST SV DQO DQOsol Nk (g/kg) (g/kg) (g/kg) (g/kg) (g/kg) (g/kg) TR VCO (g (días) SV/L*d) 0-16 Purín 100% 75,16 52,23 97,94 21,81 5,62 4,22 15 3,48 17-36 Purín 100% 78,79 54,08 94,46 21,81 6,37 4,96 15 3,60 TDO 1% 76,26 50,11 90,30 20,59 6,12 4,98 15 3,34 37-56 Purín 100% 62,80 42,81 78,90 24,25 5,89 4,42 15 2,85 TDO 2,5% 80,97 48,75 99,10 23,83 5,83 4,38 15 3,25 57-82 Purín 100% 64,30 44,92 77,14 9,84 5,24 3,85 15 2,99 TDO 5% 56,03 117,57 13,59 5,03 3,59 15 3,73 99,25 236 Ensayos de viabilidad de codigestión en continuo 6.5. Estrategias de puesta en marcha. Resultados y discusión. En el período inicial de la puesta en marcha, hubo problemas operacionales en el control del tiempo de retención, y también con la medida de volumen de biogás, debido a fallos del sistema. Esto hizo que no se obtuvieran medidas válidas hasta el mes de junio, a pesar de que el sistema ya llevaba funcionando unos 6 meses para los reactores 1 y 2. En la Figura 6.9 se muestran los períodos considerados, con los valores medios, tanto para la fase de puesta en marcha como para la de ensayo. g SV/kg; TR (d) VCO (g SV/L*d) R1- 55ºC g SV/kg; TR (d) VCO (g SV/L*d) R2- 35ºC 80 4.0 80 4.0 70 3.5 70 3.5 60 3.0 60 3.0 50 2.5 50 2.5 40 2.0 40 2.0 30 1.5 30 1.5 20 1.0 20 1.0 10 0.5 10 0.5 0 0.0 0 240 g SV/kg; TR (d) Día VCO (g SV/L*d) R3- 35ºC 0.0 0 70 4.0 60 3.5 50 3.0 2.5 40 2.0 30 1.5 0 VCO SV-medio 3.5 2.5 2.0 1.5 1.0 0.5 0.0 0 40 -180 -160 -140 -120 20 40 60 80 -100 -80 -60 -40 -20 0 -180 -160 -140 -120 Día 4.0 40 0.0 240 Día VCO (g SV/L*d) R4- 55ºC 60 0 200 240 3.0 20 160 200 80 0.5 120 160 100 10 80 120 120 1.0 40 80 g SV/kg; TR (d) 20 0 40 20 40 60 80 200 SV 80 120 160 200 240 Día 20 40 60 80 160 -100 -80 -60 -40 -20 0 120 -100 -80 -60 -40 -20 0 80 -180 -160 -140 -120 40 -180 -160 -140 -120 -100 -80 -60 -40 -20 0 20 40 60 80 0 TR Figura 6.9. Evolución de la carga orgánica, concentración de SV, datos individuales y medias por períodos, y tiempo de retención medio (TR), para los cuatro reactores Optimización de la digestión anaerobia de purines de cerdo 237 6.5.1. Reactores 1 y 2. Puesta en marcha directamente con purines de cerdo. En la puesta en marcha de los reactores 1 y 2 se utilizó como inóculo purín de cerdo previamente digerido en un sistema de flujo-pistón. La temperatura de trabajo fue, desde el inicio, la que se pretendía estudiar (mesofílica o termofílica), y la alimentación se realizó con purín. La carga orgánica inicial fue baja y paulatinamente se fue aumentando (Figura 6.9). El contenido de nitrógeno amoniacal en el reactor se mantuvo a unos niveles relativamente bajos desde el día –170 hasta el día –110, momento a partir del cual comenzó a aumentar llegando, al final del período de puesta en marcha, a un nivel de 4000 mg N-NH4+/L (Figura 6.10). Reactor 1-55ºC mg N/L 6000 5000 4000 3000 2000 1000 0 -200 -170 -140 -110 -80 -50 -20 Día Reactor 2 -35ºC mg N/L 6000 5000 4000 3000 2000 1000 0 -200 -170 -140 -110 -80 -50 -20 Día NH4 inf. NH4 ef. NH3 Figura 6.10. Evolución del nitrógeno amoniacal y amoníaco libre, del influente y del efluente, durante la fase de puesta en marcha, para los reactores 1 y 2. Debido a los altos valores de pH, el nivel de amoníaco libre, aumentó por 238 Ensayos de viabilidad de codigestión en continuo encima del nivel 650 mg N-NH3/L, indicado como límite de inhibición por algunos autores (Angelidaki y Ahring, 1993b), a partir del día –80. La concentración de amoníaco libre en el reactor 2 (35ºC) se mantuvo a un nivel relativamente bajo, por debajo de 650 mg N-NH3/L. El nitrógeno amoniacal aumentó porque el purín utilizado cada vez fue más concentrado, (Figura 6.11 y Figura 6.12), aumentando, también, la carga orgánica (Figura 6.9). Reactor 2 - 35ºC Reactor 1 - 55ºC 80 SV (g SV/kg) SV (g SV/kg) 80 60 40 20 60 40 20 0 -200 -170 -140 -110 -80 -50 0 -200 -170 -20 Día SV influente -140 -110 -80 -50 -20 Día SV efluente Figura 6.11. Evolución de los SV del influente y del efluente, durante la fase de puesta en marcha, para los reactores 1 y 2. 120 Reactor 1 - 55ºC 120 100 DQO (g O2/kg) DQO (g O2/kg) 100 80 60 40 80 60 40 20 20 0 -200 -170 -140 -110 Reactor 2 - 35ºC -80 -50 -20 Día DQO Influente 0 -200 -170 -140 -110 -80 -50 -20 Día DQO Efluente Figura 6.12. Evolución de la DQO del influente y del efluente, durante la fase de puesta en marcha, para los reactores 1 y 2. Los niveles de eliminación de materia orgánica fueron bastante altos para ambos reactores (Tabla 6.6), pero especialmente para el reactor 2 (35ºC), donde la eliminación de DQO fue de un 58% de media. A pesar de estos Optimización de la digestión anaerobia de purines de cerdo 239 altos niveles de eliminación, la producción de gas se mantuvo relativamente baja, por debajo del valor que corresponde a la eliminación de DQO observada. En el reactor 1, para una eliminación de DQO del 45%, la producción de gas media fue de 0,26 L de CH4/L·d, mientras que el valor teórico a partir del nivel de eliminación es 0,55 L de CH4/L·d. En el reactor 2, rango mesofílico, también se detectaron diferencias importantes, de 0,60 a 0,76 L de CH4/L·d, indicando que probablemente la medida de DQO fuera incorrecta, o como posteriormente se comprobó, estaba produciéndose una acumulación de sólidos en el reactor, lo que hacía que la concentración medida de DQO del efluente fuera menor de la real. Tabla 6.6. Eliminación de materia orgánica durante el período de puesta en marcha en los reactores 1 y 2 (Valores medios del período indicado). Período -178- -107 -106- -69 -68--39 -38- -20 -19- 0 Media Eliminación de materia orgánica (%) Reactor 1 – 55ºC Reactor 2 – 35ºC ST SV DQO ST SV DQO 26,51 45,48 40,91 18,15 33,78 60,83 45,48 51,55 48,02 44,44 53,78 52,85 38,86 46,72 51,65 49,56 58,65 65,16 46,69 52,77 49,66 53,56 61,71 59,92 51,19 55,94 40,57 44,14 48,94 46,63 37,38 48,87 45,08 35,46 46,72 58,21 Inicialmente la producción volumétrica de gas fue bastante similar entre los dos reactores, pero a partir del día -60, la producción del reactor 2 (35ºC) comenzó a aumentar, separándose de su homólogo termofílico (Figura 6.13). El reactor 1 (55ºC), experimentó una importante tendencia a disminuir la producción respecto a los sólidos volátiles, y de forma más retardada se observó el mismo efecto sobre la producción volumétrica de metano (Figura 6.13). La disminución de la producción de metano respecto a SV, experimentada a partir del día –40, se puede relacionar con el aumento de la concentración de nitrógeno amoniacal, que en ese período sobrepasó el nivel de 2,50 g NNH4+/L en el reactor 1 (Figura 6.11). El sistema se mantuvo bastante estable durante gran parte del período inicial, especialmente el reactor 2 (mesofílico), como demuestra los valores relativamente bajos de relación de alcalinidad (RA) observados (Figura 6.14). El reactor 1 mostró valores de RA entre 0,3 y 0,4, por encima del valor 240 Ensayos de viabilidad de codigestión en continuo umbral de fallo inminente (Iza, 1995). En cualquier caso, fueron valores moderados comparados con otros resultados obtenidos en el presente trabajo, aunque con tendencia a incrementarse al aumentar la concentración de amonio, a partir del día -40. El reactor 2, en cambio, mantuvo una relación de alcalinidad por debajo de 0,3 durante casi todo el período de puesta en marcha, lo que indica el buen desarrollo del proceso en el rango mesofílico. 0.5 1.2 Pc (L CH4/g SV) PV (L CH4/L reactor·día) 1.4 1.0 0.8 0.6 0.4 0.3 0.2 0.1 0.2 0.0 -120 0.4 -100 -80 -60 -40 -20 0 Día Reactor 1-55ºC 0.0 -120 -100 -80 -60 -40 -20 0 Día Reactor 2-35ºC Figura 6.13. Evolución de la producción de metano, Pv y Pc, durante la fase de puesta en marcha, para los reactores 1 y 2. RA 0.50 0.45 0.40 0.35 0.30 0.25 0.20 0.15 0.10 -200 -180 -160 -140 -120 -100 -80 -60 -40 -20 Reactor 1- 55ºC 0 Día Reactor 2 - 35ºC Figura 6.14. Evolución de la relación de alcalinidad (RA), durante la fase de puesta en marcha, para los reactores 1 y 2. La alcalinidad total y la debida al bicarbonato (Figura 6.15) mostraron una evolución similar a la de la materia orgánica, o el nitrógeno, ya que las Optimización de la digestión anaerobia de purines de cerdo 241 variaciones fueron fruto de la utilización de un purín más diluido. En todos los casos, la alcalinidad al bicarbonato se mantuvo por encima del valor recomendado en la bibliografía para asegurar la estabilidad del proceso, de 2,5 g de CaCO3/L (Fannin, 1987). AT y AP (g CaCO3/L) AT y AP (g CaCO3/L) Reactor1- 55ºC 30 30 25 25 20 20 15 15 10 10 5 5 0 -200 -170 -140 -110 -80 -50 -20 Día AT Reactor2- 35ºC 0 -200 -170 -140 -110 -80 -50 -20 Día AP Figura 6.15. Evolución de la alcalinidad total (AT) y parcial (AP), durante la fase de puesta en marcha, para los reactores 1 y 2. 6.5.2. Reactores 3 y 4. Puesta en marcha utilizando lodos de depuradora digeridos En los reactores 3 y 4 se utilizó como inóculo lodo de depuradora digerido, y se alimentó con mezcla de lodo digerido y sin digerir, substituyendo paulatinamente el lodo por purín de cerdo (la misma mezcla que la utilizada en los otros reactores). El tiempo de retención utilizado fue de 20 días, para mantener las condiciones iniciales del inóculo utilizado. El día –1 se cambió el tiempo de retención de 20 a 15 días, para igualar las condiciones de operación en todos los reactores (Figura 6.9). Los lodos de depuradora tienen un alto contenido de nitrógeno, pero mayoritariamente en forma orgánica por lo que no resulta tan problemático para el proceso de digestión anaerobia. La mineralización producida en los reactores 3 y 4, medida como el aumento de la fracción de nitrógeno amoniacal sobre el total, fue importante, especialmente en el rango termofílico, lo que hizo que la concentración de amonio en el efluente fuera relativamente alta, a pesar del bajo contenido en la alimentación. En cualquier caso, la concentración de amonio se mantuvo 242 Ensayos de viabilidad de codigestión en continuo por debajo de 2000 mg N-NH4+/L hasta el día –30, subiendo hasta 3000 mg N-NH4+/L al final del período de puesta en marcha (Figura 6.16). La concentración de amoníaco libre en el reactor 3 (35ºC) se mantuvo muy por debajo de 650 mg N-NH3/L, valor propuesto como límite de inhibición en la literatura (Angelidaki y Ahring, 1993b), pero no en el reactor 4, donde al final del período de puesta en marcha llegó a superar este valor (Figura 6.16). Reactor 3 - 35ºC mg N/L 6000 5000 4000 3000 2000 1000 0 -150 -130 -110 -90 -70 -50 -30 -10 -50 -30 -10 Día Reactor 4 - 55ºC mg N/L 7000 6000 5000 4000 3000 2000 1000 0 -150 -130 Nk Inf. -110 -90 NH4 inf. -70 NH4 ef. Día NH3 Figura 6.16. Evolución del nitrógeno amoniacal y amoníaco libre, del influente y del efluente, durante la fase de puesta en marcha, para los reactores 3 y 4. La concentración media de amonio , durante la fase de puesta en marcha, en el efluente de los reactores 3 y 4 fue mayor de un 50% más que en el influente, siendo, además, significativamente diferente entre los dos reactores. La evolución de los parámetros de materia orgánica se muestran en la Figura 6.17 y Figura 6.18. Se observa una tendencia ascendente conforme fue aumentando la proporción de purín en la mezcla. Optimización de la digestión anaerobia de purines de cerdo Reactor 3 Reactor 4 100 80 80 SV (g SV/kg) 100 SV (g SV/kg) 243 60 40 60 40 20 20 0 -150 -130 -110 -90 -70 -50 -30 0 -150 -130 -110 -90 -10 -70 -50 -30 Día SV Influente -10 Día SV Efluente Figura 6.17. Evolución de SV del influente y del efluente, durante la fase de puesta en marcha, para los reactores 3 y 4. Reactor 4- 55ºC 120 100 100 DQO (g O2/kg) DQO (g O2/kg) Reactor 3-35ºC 120 80 60 40 60 40 20 20 0 -150 -130 -110 -90 80 0 -70 -50 -30 -10 Día DQO Influente -150 -130 -110 -90 -70 -50 -30 -10 Día DQO Efluente Figura 6.18. Evolución de DQO del influente y del efluente, durante la fase de puesta en marcha, para los reactores 3 y 4. La eliminación de DQO media en el período de puesta en marcha fue del 54,5% en el reactor 3 y de 45,0% en el reactor 4. La menor eliminación media en el reactor 4, se debió a la baja eliminación obtenida en el primer período, probablemente causada por el cambio de rango de temperatura. Sin embargo, en el período siguiente se obtuvieron ya valores del orden del 50% (Tabla 6.7). 244 Ensayos de viabilidad de codigestión en continuo Tabla 6.7. Eliminación de materia orgánica durante el período de puesta en marcha en los reactores 3 y 4 (Valores medios del período indicado) Período -145- -117 -116- -99 -98- -69 -69- -39 -39- -20 -19-0 Media Eliminación de materia orgánica (%) Reactor 3 – 35ºC Reactor 4 – 55ºC ST SV DQO ST SV DQO 53.27 31.89 58.59 -71.36 -73.24 -6.34 47.46 55.48 54.83 48.54 57.27 50.84 41.40 49.93 59.70 38.81 49.39 52.01 48.80 55.90 49.16 39.11 49.73 46.31 21.85 28.99 62.22 42.38 50.61 53.05 44.20 52.67 58.57 47.35 53.18 58.58 43.75 45.79 56.91 31.05 39.06 45.86 El nivel de eliminación de DQO observado se correspondería, aplicando la relación de 0,402 l de CH4/g DQO eliminada (correspondiente a la temperatura de 20ºC), con una producción diaria media de 0,69 L CH4/L·d, el reactor 3, y 0,62 L CH4/L·d en el reactor 4. Sin embargo la producción media medida fue 0,55 L CH4/L·d en el reactor 3 (35ºC) y 0,45 L CH4/L·d el reactor 4 (55ºC). Esta diferencia se atribuye a que una agitación insuficiente para conseguir la mezcla perfecta. La producción de metano se mantuvo en los dos reactores a niveles muy similares hasta el final del período de puesta en marcha, tal y como se observa en la Figura 6.19, mostrando, por tanto poca influencia de la temperatura de proceso. 0.50 1.2 Pc(L CH4/g SV) Pv (L CH4/L reactor·día) 1.4 1.0 0.8 0.6 0.4 0.30 0.20 0.10 0.2 0.0 -125 0.40 -100 -75 -50 -25 0.00 -125 0 Día Reactor 3-35ºC D ía -100 -75 -50 -25 0 Día Reactor 4-55ºC Figura 6.19. Evolución de la producción de metano, Pv y Pc, durante la fase de puesta en marcha, para los reactores 3 y 4. La alcalinidad total y la parcial mostraron una tendencia ascendente a partir Optimización de la digestión anaerobia de purines de cerdo 245 del momento en que se introdujo purín (Figura 6.20), ya que la alcalinidad de este substrato es mucho más alta que la del lodo de EDAR. AT y AP (g CaCO3 /L) AT y AP (g CaCO3 /L) Reactor3- 35ºC 20 15 15 10 10 5 5 0 -150 Reactor4- 55ºC 20 -125 -100 -75 -50 -25 0 Día AT 0 -150 -125 -100 -75 -50 -25 0 Día AP Figura 6.20. Evolución de la alcalinidad total (AT) y parcial (AP), durante la fase de puesta en marcha, para los reactores 3 y 4. Durante todo el proceso la relación de alcalinidad (RA) se mantuvo alta, lo que indica cierto desequilibrio del proceso. En el reactor 4 (55ºC), inicialmente fue muy alta, en torno a 0,5, pero con tendencia a disminuir, situándose al final de la puesta en marcha en torno a 0,35 nivel similar al reactor 3 (35ºC) (Figura 6.21). El alto valor inicial de relación de alcalinidad del reactor 4, se explica por la situación de estrés a que se sometió a los microorganismos al cambiar la temperatura. La introducción de purín, sin embargo, no afectó a la relación de alcalinidad, probablemente debido a que el nivel de amonio se mantuvo relativamente bajo. La reacción al aumento de la carga orgánica (Figura 6.9) fue mayor en el reactor 3 que en el 4, donde se observó un importante incremento de la RA alrededor del día –115 (Figura 6.21). El cambio de rango de temperatura de mesofílico a termofílico provocó inicialmente un importante desequilibrio en el sistema, con un incremento de la relación de alcalinidad. A pesar de dicho desequilibrio, el rango termofílico parece menos sensible a los cambios de carga orgánica que el mesofílico. 246 Ensayos de viabilidad de codigestión en continuo RA 0.50 0.45 0.40 0.35 0.30 0.25 0.20 0.15 0.10 -150 -125 -100 Reactor4-55ºC -75 -50 -25 0 Día Reactor3-35ºC Figura 6.21. Evolución de la relación de alcalinidades (RA), durante la fase de puesta en marcha, para los reactores 3 y 4. 6.5.3. Comparación de estrategias de puesta en marcha. En la Figura 6.22 se muestra la producción volumétrica de metano para los cuatro reactores. Se puede observar que la producción volumétrica es mayor en el reactor 2 que en el resto, con diferencias significativas a un nivel de significación del 5% (Tabla 6.8). Sin embargo, las diferencias entre los reactores 2 y 3 para el parámetro producción por unidad de carga es mucho menor (Figura 6.23), aunque continúa siendo significativa (Tabla 6.8). Estas diferencias se explican por el diferente tiempo de retención empleado en cada uno de los de los reactores mesofílicos, 15 días en el reactor 2 y 20 en el reactor 3. Tabla 6.8. Producción de biogás y de metano media de los 16 últimos días del período de puesta en marcha para los cuatro reactores. Pc Pv Pv (L biogás/L·día) (L CH4/L·día) (L CH4/g SV) Reactor 1- 55ºC 1 0.395 C 0.180 D 0.052 D Reactor 2 – 35ºC 1 1.483 A 0.996 A 0.293 A Reactor 3 – 35ºC 2 1.027 B 0.727 B 0.258 B Reactor 4 – 55ºC 2 0.996 B 0.610 C 0.217 C Letras diferentes significan diferencias significativas (Test de Duncan D=0.05). El análisis estadístico se ha realizado para los últimos 16 días del período de puesta en marcha, para cada variable, por columnas. Reactor Estrategia Optimización de la digestión anaerobia de purines de cerdo 247 1.6 Pv (L CH4 /L reactor·día) 1.4 1.2 1.0 0.8 0.6 0.4 0.2 0.0 -125 -100 -75 Reactor 1-55ºC Reactor 2-35ºC -50 Reactor 3-35ºC -25 Reactor 4-55ºC 0 Día Figura 6.22. Producción volumétrica de metano, para los cuatro reactores. Realizando el estudio de los efectos principales, se observa que tanto la temperatura como el tiempo de retención fueron significativos a un nivel de confianza del 95%, así como la interacción de ambos. Los reactores mantenidos en el rango mesofílico obtuvieron mayores producciones de gas que los termofílicos. El tiempo de retención que resultó en una mayor producción de gas, dentro del rango termofílico, fue el de 20 días, debido a los malos resultados obtenidos para el TR de 15 días en el rango termofílico. Sin embargo, tal y como se observa en la Tabla 6.8, y en las Figura 6.22 y Figura 6.23, la máxima producción de gas se obtuvo para el reactor mesofílico con un tiempo de retención de 15 días En los reactores termofílicos (reactores 1 y 4), la producción volumétrica (Pv) y la producción por unidad de carga orgánica (Pc), fue más alta en el reactor 4 (estrategia 2) que en el reactor 1 (estrategia 1), a pesar de que el tiempo de retención fue mayor en el 4 (20 días) que en el 1 (15 días). Esta menor producción se atribuye a la mayor inhibición por amonio sufrida en el reactor 1, ya que la concentración de amonio durante este período fue mayor para el reactor 1 que en el 4 (Figura 6.10 y Figura 6.16), debido al efecto de dilución de la mezcla con lodos. Sin embargo, y a pesar de que la producción volumétrica en el reactor 4 se mantuvo aproximadamente constante, la producción por unidad de carga mostró una tendencia a disminuir, indicando ya un leve efecto inhibitorio, compensado por el aumento de la carga orgánica (Figura 6.9). 248 Ensayos de viabilidad de codigestión en continuo 0.50 Pc(L CH4 /g SV) 0.40 0.30 0.20 0.10 0.00 -125 -100 Reactor 1-55ºC -75 Reactor 2-35ºC -50 Reactor 3-35ºC -25 Reactor 4-55ºC 0 Día Figura 6.23. Producción de metano por unidad de carga (g SV), para los cuatro reactores. 6.5.4. Conclusiones parciales. La puesta en marcha con lodos de depuradora, comenzando con un nivel de carga orgánica relativamente alto fue posible, llegando a resultados similares que en los reactores alimentados con purín desde el inicio. No se observaron efectos irreversibles de sobrecarga, por lo que se podría, incluso, haberse introducido el 100% de purín y reducir el tiempo de retención antes de lo que se hizo. En el rango termofílico el parámetro que parece afectar más es la concentración de nitrógeno amoniacal, independientemente del inóculo utilizado. El lodo mesofílico se adapta bien al rango termofílico, aunque al cambiar la temperatura se produjo un ligero desequilibrio, con un incremento de la relación de alcalinidad. Este desequilibrio fue rápidamente superado, de forma que en 20 días los niveles de RA fueron iguales para los reactores mesofílico y termofílico, sin que se observaran diferencias en la producción de gas. El reactor mesofílico es más sensible a las sobrecargas orgánicas que el termofílico, que se tradujo en un ligero aumento de la relación de alcalinidad al aumentar la carga orgánica. Optimización de la digestión anaerobia de purines de cerdo 249 6.6. Resultados y discusión de la codigestión de purín con tierras decolorantes de aceite de oliva. A continuación se procederá a la exposición de los resultados y discusión para cada uno de los reactores por separado, pasando posteriormente al análisis comparativo de los diferentes reactores. Los 82 días totales de experimentación se han dividido en cuatro períodos, tal y como se muestra en la Tabla 6.3. 6.6.1. Digestión anaerobia de purín de cerdo. Rango termofílico (R1) El reactor 1 se mantuvo a 55ºC y fue alimentado con purín de cerdo, sin añadir ningún cosubstrato, sirviendo de control del efecto de la codigestión en el rango termofílico. El tiempo de retención desde el inicio fue de 15 días. Evolución de la producción de gas. Los niveles de producción de gas en el reactor 1, a 55ºC, fueron muy bajos, observándose, además, una disminución a medida que avanzaba el proceso, tanto del volumen de biogás producido, como de la concentración de metano del mismo. En la Figura 6.24 se observa la evolución de la producción volumétrica de biogás, la producción de metano por unidad de carga y la riqueza en metano del biogás. Dada la tendencia observada en el período de puesta en marcha, de fuerte disminución de la producción de gas, se decidió, durante el primer período, disminuir la carga orgánica, para evitar la posible acidificación del reactor. Así el tiempo de retención, durante este período, fue de 72 días (Figura 6.9), y la carga orgánica disminuyó de 3,3 a 0,7 g SV/L·día. Debido a esta reducción de la carga orgánica, entre los días 0 y 16 hubo un aumento de la producción de metano por unidad de SV, aunque la producción volumétrica continuó su tendencia descendente. El contenido en metano en el gas pareció estabilizarse en torno al 30%, la producción de biogás se estabilizó en 0,43 L/L*día y la producción de metano entre 44 mL CH4/g SV (Figura 6.24). Estos valores de producción de gas no podrían admitirse en una planta a escala industrial, estando muy por debajo de los valores normales para residuos ganaderos, incluso en experiencias en termofílico con altos niveles de amonio (Hill et al., 1987; Hansen et al., 1998). 250 Ensayos de viabilidad de codigestión en continuo Pv; Pc 0.6 3.30 0.7 0.5 2.86 2.80 0.4 0.3 0.2 0.1 0.0 0 10 20 30 Pv (L de Biogas/L*d) 40 50 60 Pc (L de CH4 /g SV) 70 80 % CH4 100% 90% 80% 70% 60% 50% 40% 30% 20% 10% 0% 90 Día % de CH4 Figura 6.24. Evolución de la producción volumétrica de biogás (Pv), la producción por unidad de carga (Pc) y la concentración de metano en el biogás (%) en el reactor 1. Los diferentes períodos corresponden a cambios en la VCO (g SV/d), valores indicados en la figura Evolución de formas nitrogenadas en el influente y efluente. Las concentraciones de nitrógeno total y amoniacal, en el influente, mostraron una tendencia ascendente hasta el día 35 (Figura 6.25). A partir de este día se cambió la proporción de mezclas de diferentes tipos de purín, con el objetivo de bajar la concentración de nitrógeno amoniacal del influente. La concentración en el efluente, y por tanto en el reactor, siguió la misma tendencia que en el influente, aunque desplazado en el tiempo. La concentración de amonio en el reactor llegó a un máximo de 5,40 g NNH4+/L. A pesar del cambio en la composición del influente (Tabla 6.3) la concentración de amonio continuó estando por encima de 4,00 g N-NH4+/L hasta el final del experimento. Las altas concentraciones de amonio, junto con el pH básico (Figura 6.26), debido a la alta alcalinidad del purín, hizo que la concentración de amoníaco libre se mantuviera muy alta, por encima del nivel inhibidor de 650 mg N-NH3/L (Angelidaki y Ahring, 1993b), durante casi todo el experimento, con picos que superaron los 1000 mg N-NH3/L. Evolución de la alcalinidad y ácidos grasos volátiles. Los residuos ganaderos presentan un alto nivel de alcalinidad, lo que los convierte en buenos substratos base para la codigestión anaerobia. Sin embargo, en los casos con alto contenido en nitrógeno amoniacal, la gran capacidad tampón limita la capacidad de autorregulación del sistema que Optimización de la digestión anaerobia de purines de cerdo 251 describió Angelidaki y Ahring (1993b), ya que a pesar de acumularse grandes concentraciones de ácidos, no se produce una bajada significativa del pH (Figura 6.26) y por tanto la concentración de amoníaco libre se mantiene alta. 7000 6000 mg N/L 5000 4000 3000 2000 1000 650 0 0 10 20 Nk Inf R1 NH4 Ef. 30 40 50 Nk ef. R1 NH3 60 70 80 NH4 inf. 90 Día Figura 6.25. Evolución de Nitrógeno Kjeldahl (Nk ) y amoniacal (NH4) del influente y efluente en el reactor 1. Concentración de amoníaco libre en el efluente 8.5 pH efluente 8 7.5 7 6.5 0 20 40 60 80 Día Figura 6.26. Evolución del pH del efluente del reactor 1. La alcalinidad total del influente se mantuvo durante todo el período de experimentación entre 15 y 20 g CaCO3/L, así como la del efluente (Figura 6.27), y la alcalinidad parcial, asociada al bicarbonato, entre 7,5 y 11,5 g CaCO3/L, muy por encima del valor mínimo recomendado de 2,5 g CaCO3/L (Fannin, 1987). 252 Ensayos de viabilidad de codigestión en continuo 1.00 0.90 0.80 0.70 0.60 0.50 0.40 0.30 0.20 0.10 0.00 25 g CaCO3 /L 20 15 10 5 0 0 10 20 AT 30 40 50 AP 60 70 80 RA RA (AI/AT) La relación de alcalinidad del influente mostró valores relativamente altos, indicando que el influente se encontraba parcialmente acidificado. La relación de alcalinidades en el efluente del reactor 1 se mantuvo durante todo el período de experimentación por encima de 0,4 (Figura 6.27), y por tanto por encima del umbral de alarma de desequilibrio en el proceso, 0,3 (Iza, 1996). 90 Día Figura 6.27. Evolución de la alcalinidad total (AT), parcial o al bicarbonato (AP) y relación de alcalinidad (RA) en el reactor 1 El alto valor de la relación de alcalinidad indica una fuerte acumulación de ácidos grasos volátiles. El principal ácido acumulado en el reactor fue el ácido acético (Figura 6.28), que aumentó hasta el día 50, llegando a unos niveles de concentración muy altos, en torno a los 10 g Ac/L (167 mM). Posteriormente disminuyó, al bajar la concentración de amonio, aunque se mantuvo siempre por encima de 6 g Ac/L. Aunque el propiónico alcanzó niveles de concentración relativamente altos, la relación propiónico/acético (P/A) se mantuvo baja, en torno a 0,2. Hill et al. (1987) considera que el proceso anaerobio fracasa si la producción de metano es inferior a 0,25 L CH4/g SV, relacionándolo con un nivel de acético superior a 800 mg Ac/L y/o con una relación P/A mayor de 1,4. La primera condición sobre la concentración de acético se ha superado ampliamente en el presente experimento. La importante acumulación de acético y la baja relación P/A indica la inhibición de la fase metanogénica acetoclástica. El comportamiento de los ácidos butírico y valérico se ha relacionado con el del acético y propiónico. El que mayor acumulación sufrió fue el isómero n-butírico, con una curva de forma similar a la del acético, mientras que los Optimización de la digestión anaerobia de purines de cerdo 253 isómeros iso, tanto del butírico como del valérico, sufrieron una acumulación más ligera y de forma más parecida al propiónico (Figura 6.28). mg AGV /L P/A 0.8 12000 3.30 0.7 2.86 2.80 10000 0.6 8000 0.4 6000 4000 0.2 2000 0.0 0 0 10 20 30 40 50 60 70 80 90 70 80 90 Día Día mg AGV /L 1600 3.30 0.7 1400 2.86 2.80 1200 1000 800 600 400 200 0 0 10 20 30 40 50 60 Acético Propiónico Prop/acet Iso Butírico N Butírico Iso Valérico N Valérico Figura 6.28. Evolución de la concentración de AGV en el efluente del reactor 1. Los diferentes períodos corresponden a cambios en la VCO (g SV/d), valores indicados en la figura Una concentración por encima de 1 g Ac/L se ha señalado como límite a partir del cual comienza a producirse la inhibición de los organismos que degradan el butírico, señalándose en unos 5 g Ac/L el punto en que la inhibición es completa (Ahring y Westermann, 1988). En este estudio la concentración de n-butírico comenzó a incrementarse de forma importante, sólo a partir de una concentración de acético de 6 g Ac/L, aunque 254 Ensayos de viabilidad de codigestión en continuo anteriormente se encontraba ya a niveles relativamente altos (200 mg But/L). El propiónico y las formas iso del butírico y el valérico comenzaron a acumularse a un nivel de concentración de acético similar, en torno a 6 gAc./L, pudiendo por tanto señalar este valor como límite de inhibición de la acetogénesis. El incremento porcentual de la concentración de las formas iso del butírico y valérico, así como del propiónico (100%) fue del mismo orden de magnitud que el incremento de acético (100%), pero menor que el aumento del n-butírico (500%). Sin embargo, el n-valérico apenas sufrió acumulación durante el proceso. Evolución de parámetros de materia orgánica: influente y efluente. Pocas conclusiones se pueden extraer del estudio de los parámetros de materia orgánica. Del día 0 al 37 se obtuvo una eliminación media de DQO del 44,08% y de SV del 59,19%, lo que no concuerda con los bajos valores de biogás obtenidos. Se observó que la homogeneización dentro del reactor no era completa, produciéndose acumulación de sólidos en el reactor. Para evitarlo, se aumentó la velocidad de agitación del reactor de 200 a 500 rpm, obteniéndose un aumento muy importante en la concentración de partículas en el efluente (Figura 6.29 y Figura 6.30), y por tanto una disminución en la eliminación de los parámetros de materia orgánica (Tabla 6.9). 140 3.30 0.7 2.86 2.80 DQO (g O2 /kg) 120 100 80 60 40 20 0 0 10 20 30 40 50 60 70 80 90 Día DQO Influente DQO Efluente DQOs Influente DQOs Efluente Figura 6.29. Evolución de la DQO total y soluble en el influente y efluente del reactor 1. Optimización de la digestión anaerobia de purines de cerdo 255 Camb io en la ag itació n 100 3 .3 0 0 .7 2 .8 6 2 .8 0 80 g /kg 60 40 20 0 0 10 20 ST in fluen te 30 40 ST eflu en te 50 60 SV in fluen te 70 80 SV eflu en te 90 Día Figura 6.30. Evolución de ST y SV en el influente y efluente del reactor 1.Los diferentes períodos corresponden a cambios en la velocidad de carga orgánica, VCO (g SV/L·d), valores indicados en la figura. La flecha roja indica el cambio de agitación de 200 a 500 rpm El cambio del régimen de agitación provocó la obtención de eliminaciones negativas para los diferentes parámetros de materia orgánica, lo que carece de sentido biológico. Los valores de eliminación de DQO y SV, medios de todo el período de experimentación, fueron, respectivamente, de 7,39% y 26,21% (Tabla 6.9). Al considerar también el período de puesta en marcha, durante el cual ya se produjo acumulación de sólidos en el reactor, los niveles de eliminación aumentan bastante, llegando a un nivel de 41,5% de SV y 32,81% de DQO. En general, excepto en el período final, la DQO soluble del efluente se mantuvo por debajo de la del influente. La DQO soluble del efluente se mantuvo entre 10 y 20 g O2/L durante la mayor parte del proceso, observándose un aumento, coincidiendo con la acumulación de AGV. La principal fracción de la DQO soluble en el efluente, está constituida por AGV. La relación sólidos volátiles sobre sólidos totales (SV/ST) fue menor en el efluente que en el influente, pasando de una media de 69,13%, en la alimentación a 63,59%, indicando que hubo una disminución de la materia orgánica del reactor. 256 Ensayos de viabilidad de codigestión en continuo Tabla 6.9. Eliminación de parámetros de materia orgánica en el reactor 1, calculada por períodos Período EST (%) ESV (%) EDQO (%) 0-16 50.74 54.92 52.17 17-36 57.04 62.79 37.26 37-56 -33.35 -29.84 -32.55 57-82 16.78 23.23 -10.12 Media experimento 21.02 26.21 7.39 Media global 32.05 41.49 32.81 Índices de metanización y acidificación El índice de metanización se mantuvo siempre muy por debajo del índice de acidificación (Figura 6.31), indicando que el principal problema estuvo en la fase metanogénica. 35% 3.30 0.7 30% 2.80 2.86 % DQO 25% 20% 15% 10% 5% 0% 0 10 20 30 40 % Metanización 50 60 % acidificación 70 80 90 Día Figura 6.31. Evolución de los índices de metanización y de acidificación en el reaactor 1, expresaada en % de DQO transformada. Los diferentes períodos corresponden a cambios en la velocidad de carga orgánica, VCO (g SV/L·d), valores indicados en la figura Los niveles de acidificación, pese a ser muy superiores a los de metanización, no alcanzaron los valores normales en los procesos anaerobios de este tipo de residuos, en torno al 40-50%, acorde con los niveles aceptables de eliminación de DQO (Hill et al., 1987), indicando que la fase de hidrólisis- Optimización de la digestión anaerobia de purines de cerdo 257 acidogénesis no fue muy eficiente. Los valores del índice de metanización al inicio fueron más altos debido al mayor tiempo de residencia durante este período, lo que provocó una mayor degradación de la materia orgánica aportada, aunque en cualquier caso por debajo de un nivel aceptable. 6.6.2. Digestión anaerobia de purín de cerdo. Rango mesofílico (R2) El reactor 2 ha sido utilizado como control del experimento en mesofílico. El tiempo de retención de trabajo durante todo el experimento fue de 15 días. Evolución de la producción de gas. El funcionamiento general del proceso anaerobio de purines mejoró en el rango mesofílico, respecto al termofílico. La producción volumétrica de biogás se mantuvo durante el inicio por encima de la unidad, aunque al disminuir la carga de 3,55 a 2,86 g SV/L·d se observó un importante descenso (Figura 6.32). La producción de metano por unidad de carga mostró valores comparables con los obtenidos de la bibliografía (Fischer et al., 1979; van Velsen, 1979; Hill et al., 1987). Pv; Pc 2.0 3.40 1.8 1.6 1.4 1.2 1.0 0.8 0.6 0.4 0.2 0.0 0 10 3.55 20 2.80 2.86 30 Pv (L de Biogas/L*d) 40 50 60 Pc (L de CH4 /g SV) 70 80 % CH4 100% 90% 80% 70% 60% 50% 40% 30% 20% 10% 0% 90 % de CH4 Figura 6.32. Evolución de la producción volumétrica de biogás (Pv), la producción por unidad de carga (Pc), concentración de metano en el biogás (%) en el reactor 2. Los diferentes períodos corresponden a cambios en la velocidad de carga orgánica, VCO (g SV/L·d), valores indicados en la figura. El biogás mostró, en todo momento, una elevada riqueza en metano, aunque al final del experimento disminuyó ligeramente (Figura 6.32), aunque se 258 Ensayos de viabilidad de codigestión en continuo mantuvo por encima del 60% de metano. La producción de metano por unidad de carga (L CH4/g de SV) tuvo un valor medio de 0,26 L de CH4/g SV, con puntas que llegaron a 0,40 L de CH4/g SV. Los resultados son mayores a los obtenidos en los ensayos en discontinuo (capítulo 4), donde se obtuvo una producción de metano por unidad de SV inicial, después de 75 días, de 0,15 L de CH4/g SV (capítulo 5). Evolución de formas nitrogenadas en el influente y efluente. La concentración de amonio en el efluente aumentó hasta el día 44, pasando de una concentración de 3,32 g N-NH4+/L a 4,88 g N-NH4+/L, disminuyendo posteriormente, hasta situarse alrededor de los 4 g N-NH4+/L (Figura 6.33). El aumento de la concentración de amonio hizo que aumentara también la forma libre (NH3), que durante varios días se situó por encima de 650 mg N-NH3/L, límite inhibidor de referencia (Angelidaki y Ahring, 1993b). Sólo a partir del día 60 se observó una bajada del NH3, debido al efecto conjunto de la disminución del amonio y la bajada del pH por debajo de 8 (Figura 6.34). A pesar de esta bajada en la concentración de NH3 no se observó un aumento en la producción de metano (Figura 6.32). 8000 3.40 3.55 2.80 2.86 7000 mg N/L 6000 5000 4000 3000 2000 650 1000 0 0 10 20 Nk inf. R2 NH4 ef. 30 40 50 Nk ef. R2 NH3 60 70 80 NH4 inf. 90 Día Figura 6.33. Evolución de Nitrógeno Kjeldahl (Nk ) y amoniacal (NH4) del influente y efluente y concentración de amoníaco libre en el efluente, en el reactor 2 Optimización de la digestión anaerobia de purines de cerdo 259 Evolución de alcalinidad y ácidos grasos volátiles. La alcalinidad total se situó entre 16 y 21 g CaCO3/L, y la alcalinidad al bicarbonato entre 12 y 16 g CaCO3/L, siendo por tanto más que suficiente para el correcto funcionamiento de la digestión anaerobia (Fannin, 1987). La relación de alcalinidad del efluente se mantuvo a un nivel adecuado durante la mayor parte del experimento, situándose entre 0,23 y 0,26, y por tanto por debajo de los niveles de alarma (Iza, 1995). La alta capacidad tampón del medio, propició que las variaciones de pH fueran poco importantes (Figura 6.34), manteniéndose a niveles altos, alrededor de 8, durante casi todo el experimento. Tan sólo en el período final se observó una ligera disminución del pH. La concentración de ácidos grasos volátiles, especialmente de acético, aumentó a partir del día 17, siendo especialmente importante a partir del día 21 (Figura 6.35), coincidiendo con el aumento de la concentración de amonio (Figura 6.33). El período de mayor acumulación de acético coincide con el de mayor concentración de amonio y de amoníaco libre, especialmente notable cuando la concentración de NH3 superó los 650 mg N-NH3/L. La concentración de acético llegó a superar 3 gAc/L (50 mM), valor límite a partir del cual el proceso metanogénico podría comenzar a estar afectado (Ahring et al., 1995). 8.5 pH efluente 8 7.5 7 6.5 0 20 40 60 80 Día Figura 6.34. Evolución del pH del efluente del reactor 2. La relación P/A se mantuvo siempre en valores muy bajos, indicando, por tanto, que la fase acetogénica no fue limitante. Aunque hubo una ligera acumulación de AGV de más de 3 átomos de carbono al aumentar la concentración de acético (Figura 6.35), la concentración de estos AGV no llegó a niveles tan altos como en el termofílico, aunque porcentualmente el 260 Ensayos de viabilidad de codigestión en continuo aumento fue muy importante, dados los bajos valores de los que se partía. La concentración de las formas iso del butírico y el valérico se han descrito como los mejores indicadores de la inestabilidad del proceso. Cobb y Hill (1991) estimaron en 15 mg/L el límite en su concentración en reactores funcionando correctamente. Los mismos autores encontraron una relación entre la fracción del isómero iso sobre el total y la presión parcial de hidrógeno (pH2), sirviendo por tanto como diagnóstico de problemas en la fase metanogénica hidrogenotrófica y acetogénica (Hill y Cobb, 1993). Ahring et al. (1995) observaron, también, que los isómeros iso son los más rápidos indicadores de problemas en el reactor, siendo los que más rápidamente se acumulan después de una perturbación. mg AGV /L 4000 P/A 3.55 3.40 0.8 2.80 2.86 3500 3000 0.6 2500 2000 0.4 1500 1000 0.2 500 0 0 10 mg AGV /L 140 20 30 40 3.55 3.40 50 60 70 80 90 0.0 Día 2.80 2.86 120 100 80 60 40 20 0 0 10 20 30 40 50 60 Acético Propiónico Prop/acet Iso Butírico N Butírico Iso Valérico 70 80 90 Día N Valérico Figura 6.35. Concentración de AGV en el efluente del reactor 2. Los diferentes períodos corresponden a cambios en la velocidad de carga orgánica, VCO (g SV/L·d), valores indicados en la figura. Optimización de la digestión anaerobia de purines de cerdo 261 En el reactor 2, la concentración de las formas iso se mantuvo muy baja, por debajo de 15 mg/L hasta el día 15, coincidiendo con un nivel de amonio por debajo de 4 g N-NH4+/L. Al aumentar la concentración de amonio se produjo un importante y rápido incremento en la concentración de todos los AGV, pero especialmente importante para las formas n, y no tanto para las formas iso, lo que indicaría que el acético inhibe esta forma concreta, siendo menos importante la acumulación de pH2. Sin embargo, la relación iso/total se mantiene alta durante todo el experimento, por lo que o siempre ha estado parcialmente inhibida la metanogénesis a partir de H2, o no es un buen indicador de este fenómeno, y sí lo es el aumento de concentración respecto al estado previo, lo que confirma los resultados de Ahring et al. (1995). Evolución de parámetros de materia orgánica: influente y efluente. Hubo importantes variaciones en la concentración de DQO y ST y SV del influente, con una importante disminución a partir del día 36 (Figura 6.36 y Figura 6.37), debido al cambió del purín utilizado. Esto hizo bajar la carga orgánica de forma importante (Figura 6.9), de 3,55 a 2,86 g SV/L*d. Se observó un ligero aumento en la concentración de ST y DQO en el efluente (Figura 6.36 y Figura 6.37) al aumentar el régimen de agitación aunque de mucha menor cuantía que el efecto observado en los reactores termofílicos, reactores 1 y 4 (Figura 6.30 y Figura 6.51). La temperatura tiene gran influencia, no sólo en los procesos biológicos y bioquímicos, sino que influye, también, en algunos parámetros físicos del reactor. A mayor temperatura, menor viscosidad de la mezcla, y por tanto, mejor capacidad de sedimentación del lodo. Este pudo ser el motivo de la mayor acumulación de sólidos en los reactores termofílicos que en los mesofílicos. La DQO soluble en la alimentación varió bastante, pero al final del proceso, se observó una disminución, lo que podría haber contribuido a la bajada en la producción de metano, debido a que la fase de hidrólisis de partículas es más lenta que el proceso de degradación de moléculas solubles. La DQO soluble del efluente se mantiene a un nivel relativamente constante, alrededor de unos 10 g O2/L, bastante alto, pero por debajo de la obtenida en el reactor 1 (termofílico). La eliminación de materia orgánica varió en función del período considerado (Tabla 6.10), aunque no de forma tan acusada como en el reactor 1 (Tabla 6.9). 262 Ensayos de viabilidad de codigestión en continuo 120 3.55 3.40 2.80 2.86 DQO (g O2 /kg) 100 80 60 40 20 0 0 10 20 DQO Influente 30 40 DQO Efluente 50 60 DQOs influente 70 80 DQOs efluente 90 Día Figura 6.36. Evolución de DQO total y soluble en el influente y efluente del reactor 2. Los diferentes períodos corresponden a cambios en la velocidad de carga orgánica, VCO (g SV/L·d), valores indicados en la figura. 100 3.55 3.40 90 2.80 2.86 Cambio en la agitación ST- SV(g/kg) 80 70 60 50 40 30 20 10 0 0 10 20 SV influente 30 40 SV efluente 50 60 ST influente 70 80 ST efluente 90 Día Figura 6.37. Evolución de ST y SV en el influente y efluente del reactor 2. Los diferentes períodos corresponden a cambios en la velocidad de carga orgánica, VCO (g SV/L·d), valores indicados en la figura. La flecha indica el cambio de agitación de 200 a 500 rpm. Optimización de la digestión anaerobia de purines de cerdo 263 Tabla 6.10. Eliminación de parámetros de materia orgánica en el reactor 2, calculada por períodos Período EST (%) ESV (%) EDQO (%) 0-16 28,43 34,74 44,28 17-36 37,02 43,36 43,47 37-56 33,11 41,51 37,93 57-82 13,70 21,91 -25,12 Media experimento 26,17 33,64 17,88 Media global 32,44 42,46 45,08 Índices de metanización y acidificación En cuanto a los índices de metanización y acidificación (Figura 6.38) se apreciaron pocas diferencias entre ellos, indicando que la metanogénesis no fue la fase limitante del proceso. 60% 3.40 3.55 2.86 2.80 50% % DQO 40% 30% 20% 10% 0% 0 10 20 30 % Metanización 40 50 60 70 % Acidificación 80 90 Día Figura 6.38. Evolución de los índices de metanización y de acidificación en el reactor 2, en % de la DQO inicial. Los diferentes períodos corresponden a cambios en la velocidad de carga orgánica, VCO (g SV/L·d), valores indicados en la figura Los niveles de acidificación, entre 40-60% en los períodos 2 y 3, indica un buen desarrollo del proceso anaerobio. Con la bajada en la concentración del influente y a pesar de la bajada en la concentración de amonio, se produjo una disminución en la eficiencia del proceso, con una bajada en la 264 Ensayos de viabilidad de codigestión en continuo producción de gas, y en el índice de acidificación por debajo del 40%. 6.6.3. Codigestión de purín con tierras decolorantes de aceite de oliva en rango mesofílico (R3). El reactor 3 se utilizó para realizar los experimentos de codigestión con tierras decolorantes de aceite de oliva en el rango mesofílico. Desde el día 0, el tiempo de retención de trabajo fue de 15 días, pudiendo, por tanto, ser comparado con el control (reactor 2). Evolución de la producción de gas. El reactor 3, mostró los mejores resultados en cuanto a producción de gas, de los cuatro reactores. Aunque la puesta en marcha se realizó con tiempo de retención hidráulica (TR) de 20 días, el día 0 ya estaba trabajando a 15 días (Figura 6.9). El aumento en la velocidad de carga orgánica (VCO) supuso una mejora de la producción volumétrica de biogás y de la producción de metano respecto a los sólidos volátiles añadidos (Figura 6.39). Pv y Pc 2.5 TDO 1% 3.53 TDO 5% TDO 2,5% 3.52 3.31 % CH4 100% 90% 80% 70% 60% 50% 40% 30% 20% 10% 0% 3.71 2.0 1.5 1.0 0.5 0.0 0 10 20 30 Pv (L de Biogas/L*d) 40 50 60 Pc (L de CH4 /g SV) 70 80 % de CH4 90 Día Figura 6.39. Evolución de la producción volumétrica de biogás (Pv); de la producción por unidad de carga (Pc) y concentración de metano en el biogás en el reactor 3. Los diferentes períodos corresponden a cambios en la velocidad de carga orgánica, VCO (g SV/L·d), valores indicados en la figura. A pesar del aumento de la concentración de amonio y de amoníaco libre desde el inicio hasta el día 16 (Figura 6.40), la producción de gas no disminuyó significativamente. El día 16 se comenzó a introducir por primera vez purín mezclado con tierras decolorantes de aceite de oliva (TDO), al 1% de concentración. Este Optimización de la digestión anaerobia de purines de cerdo 265 cambio en la alimentación supuso una mejora inmediata de la producción de metano, manteniéndose durante este período ligeramente por encima de la obtenida en el período anterior (Tabla 6.11). Al alimentar con un 2,5% de TDO, la producción volumétrica de metano disminuyó respecto al período anterior, aunque es de destacar que la velocidad de carga también disminuyó, debido a la dilución del purín. La producción de metano por unidad de carga orgánica no varió significativamente respecto al período anterior (Tabla 6.11). Al introducir un 5% de TDO se observó un incremento en la producción de gas, tanto en unidades de producción volumétrica como en la producción por unidad de carga (Tabla 6.11). La producción por unidad de carga se situó en 0,37 L/g SV, valor mejor que otros obtenidos en la bibliografía (Angeldiaki et al., 1994), por encima de los 0,25 L/g SV señalados por Hill et al. (1987), como indicadores del buen funcionamiento de un digestor. Tabla 6.11. Producción de metano media para los últimos días de cada período en el reactor 3. Período Substrato Pv (L biogás/L·día) Pv (L CH4/L·día) Pc (L CH4/g SV) 1-16 días Purín 1.41 A 1.02 A 0.29 A 17-36 días TDO 1% 1.55 B 1.13 B 0.32 B 37-57 días TDO 2,5% 1.47 AB 1.03 A 0.31 AB 58-82 días TDO 5% 1.91 C 1.34 C 0.37 C Letras diferentes significan diferencias significativas (Test de Duncan D=0.05). El análisis estadístico se ha realizado considerando los cinco últimos días de cada período y para cada variable, por columnas. Evolución de las formas nitrogenadas en el influente y efluente La concentración de amonio en el reactor 3 aumentó desde el inicio del experimento hasta el día 36, fruto del aumento de la concentración en el influente. Al final del período de alimentación con 1% de TDO, llegó al nivel de 5 g N-NH4+/L. La concentración de amoníaco libre aumentó también, sobrepasando el nivel de 650 mg N-NH3/L (Figura 6.40). A partir de este punto, y debido al menor contenido de amonio y nitrógeno total del influente, la concentración de amonio comenzó a descender lentamente, situándose al final del experimento por debajo de 4 g N-NH4+/L. A pesar de la acumulación de amonio, no se observó un efecto importante sobre la producción de gas, observándose tan sólo una ligera bajada en el 266 Ensayos de viabilidad de codigestión en continuo contenido de metano del biogás, que, en cualquier caso, se mantuvo por encima del 70% en todo momento. TDO 1% TDO 5% TDO 2,5% 7000 6000 mg N/L 5000 4000 3000 2000 1000 650 0 0 10 Nk Influente NH4+ Ef 20 30 40 50 Nk Efluente NH3 Ef. 60 70 NH4+ Inf. 80 90 Día Figura 6.40. Evolución de Nitrógeno Kjeldahl y amoniacal del influente y efluente del reactor 3. Concentración de amoníaco libre en el efluente Evolución de la alcalinidad y ácidos grasos volátiles La alcalinidad del reactor 3, tanto la alcalinidad total como la debida al bicarbonato, se mantuvo a niveles altos, más que suficientes para asegurar el correcto tamponamiento del reactor, situándose entre 15 y 23 g CaCO3/L la alcalinidad total y entre 10 y 15 g CaCO3/L la alcalinidad parcial. La relación de alcalinidad disminuyó al inicio del experimento, con respecto a la fase de puesta en marcha, y se mantuvo la mayor parte del experimento por debajo de 0,3, aunque al final, debido a la desestabilización ocasionada por la introducción del cosusbtrato (TDO), aumentó por encima de este valor. Los niveles, estuvieron siempre por debajo de los obtenidos en el rango termofílico (reactor 1 y 4). El pH se mantuvo durante todo el experimento, al niveles bastante altos, alrededor de 8, y sólo al final bajó por debajo de este umbral (Figura 6.41), debido a la mayor acumulación de AGV (Figura 6.42). Optimización de la digestión anaerobia de purines de cerdo 267 8.5 pH efluente 8 7.5 7 6.5 0 20 40 60 80 Día Figura 6.41. Evolución del pH del efluente del reactor 3. La disminución del TR de 20 a 15 días, realizada el día -1, pudo haber influido en la ligera acumulación de propiónico observada en el período inicial (Figura 6.42), aunque posteriormente disminuyó, bajando la relación P/A. Este aumento de la concentración de propiónico se puede asociar con un ligero efecto de sobrecarga orgánica. Los organismos mesofílicos parecen ser más sensibles que los termofílicos al aumento de la carga orgánica, mostrando mayor tendencia a sufrir acumulaciones de propiónico al aumentar dicho parámetro (Griffin et al., 1998). Sin embargo la adaptación fue rápida, y no llegó a afectar a la producción de biogás. El efecto más importante de la introducción de TDO a partir del día 17, fue sobre la concentración de AGV (Figura 6.42), especialmente sobre la concentración de propiónico, que aumentó hasta superar el nivel de las 2000 ppm, con un 2,5% de TDO. El acético sufrió, también, una fuerte acumulación, que en parte podría haber estado provocada por el alto nivel de amonio experimentado en este período (Figura 6.42). Al añadir un 2,5% de TDO continuó aumentando, a pesar de disminuir la concentración de NH3, superando los niveles de concentración alcanzados por el reactor 2 (Figura 6.35), llegando a un máximo de 5,5 g Ac/L. En el período de máxima acumulación de acético se observó también una ligera disminución de la producción volumétrica y de la producción por unidad de carga de metano, mostrando, además, una ligera disminución de la concentración de metano en el biogás (Figura 6.39). El incremento de la concentración de propiónico fue, sin embargo, más intenso que el de acético, de forma que la relación P/A (propiónico/acético) aumentó durante este período (1% TDO). 268 Ensayos de viabilidad de codigestión en continuo P/A TDO 1% 6000 3.52 3.53 TDO 5% TDO 2,5% 3.31 1.0 3.71 5000 0.8 4000 0.6 3000 0.4 2000 0.2 1000 80 0.0 90 Día 80 90 Día 0 0 10 mg AGV /L 600 20 30 TDO 1% 40 60 70 TDO 5% TDO 2,5% 3.52 3.53 50 3.31 3.71 500 400 300 200 100 0 0 10 20 30 40 50 60 Acético Propiónico Prop/acet Iso Butírico N Butírico Iso Valérico 70 N Valérico Figura 6.42. Evolución de la concentración de AGV en el reactor 3. Los diferentes períodos corresponden a cambios en la VCO (g SV/d), valores indicados en la figura. El acético, después de alcanzar el máximo, el día 52, disminuyó rápidamente, indicando que la metanogénesis se estaba reactivando. El propiónico, sin embargo, aunque disminuyó algo después de alcanzar el máximo, se mantuvo hasta el final a un nivel aproximadamente constante entre 1500 y 2000 mg/L. Consecuencia de ello la relación P/A aumentó al final del experimento, aunque no llegó a superar el valor de 1,4, nivel señalado en la bibliografía como límite para el fracaso (Hill et al., 1987). Durante el período de alimentación con un 2,5% de TDO la producción de biogás y de metano mostró un período crítico, coincidiendo con la Optimización de la digestión anaerobia de purines de cerdo 269 máxima acumulación de ácidos, pero que superó, a pesar de mantenerse relativamente altas las concentraciones de AGV, situándose el día 57 alrededor de 1,8 L de biogás/L·d, recuperándose también la producción por unidad de carga (Figura 6.39). El aporte de un 5% de TDO (día 57) no supuso un aumento en la concentración de AGV, sin que se observara un efecto especial de sobrecarga ni de inhibición. La producción de gas continuó la tendencia ascendente, estabilizándose al final del proceso alrededor de 1,9 L biogás/L·d y de 0,37 L de CH4/g SV (Figura 6.39), valores comparables e incluso mejores que los obtenidos por otros autores (Angelidaki y Ahring, 1993b; Hansen et al., 1998). Al adicionar TDO, el AGV, de más de 3 átomos de carbono, que mayor incremento porcentual sufrió fue el n-valérico. Este ácido en los otros reactores, tanto en el mesofílico (reactor 2) como en los termofílicos (reactores 1 y 4), apenas se detectó. La concentración de n-valérico se mantuvo alta, incluso cuando la del acético y la del propiónico habían disminuido, por lo que se puede decir que el cosubstrato TDO introduce un efecto inhibidor del proceso de degradación del ácido valérico. Las tierras decolorantes de aceite de oliva contienen elevadas concentraciones de grasas, restos del aceite que se pretende decolorar, cifrándose alrededor de un 25-30% en peso. Los ácidos grasos de cadena larga (AGCL), presentan elevados potenciales de producción de biogás (Angelidaki y Ahring, 1992), pero si se encuentran en un concentración muy alta pueden inhibir el proceso anaerobio, inhibiendo todas las fases de la digestión, especialmente la metanogénesis (tanto acetoclástica como hidrogenotrófica) y la acetogénesis (Hanaki et al., 1981). Puesto que son los ácidos grasos de cadena larga libres los auténticos tóxicos, la introducción de residuos con alto contenido en grasas en plantas de biogás se debe realizar lentamente, para así permitir el desarrollo de una población desarrollada de organismos acetogénicos que degraden los AGCL conforme se van liberando por la hidrólisis de los triglicéridos, de forma que no se alcancen niveles inhibidores (Angelidaki y Ahring, 1992). La evolución de la concentración de acético puede indicar un efecto inhibidor de los AGCL inicial, tipo fase lag, sobre la población metanogénica acetoclástica, ya descrito por Hanaki (Hanaki et al., 1981). Así, aunque inicialmente hay inhibición, al pasar el tiempo la población de microorganismos se reactiva, sin que se vea afectada la velocidad específica de crecimiento, disminuyendo la concentración de acético. Al incrementar la proporción de TDO del 1 al 2,5% no se observó efecto inhibitorio sobre la 270 Ensayos de viabilidad de codigestión en continuo metanogénesis acetoclástica. El mantenimiento de un nivel alto de concentración de propiónico hasta el final del experimento, indica otro tipo de efecto inhibitorio. Hanaki et al. (1981) encontraron que los AGCL provocan sobre los microorganismos metanogénicos hidrogenotróficos una disminución de la velocidad de crecimiento, lo que podría haber hecho que se acumulase H2, que a su vez habría provocado la acumulación de propiónico. Pese a la existencia de inhibición por AGCL, la introducción de TDO tuvo un efecto positivo sobre la producción de gas. Parece que con el paso del tiempo el efecto inhibitorio por AGCL va desapareciendo, fruto del establecimiento de una población de microorganismos acetogénicos que consuman AGCL a medida que se van produciendo por hidrólisis de grasas. Es posible que una introducción más lenta de TDO pudiera haber proporcionado mejores resultados. En cualquier caso el sistema estaba evolucionando correctamente, indicando la recuperación a medio plazo. Evolución de parámetros de materia orgánica: influente y efluente. La mezcla de purín y TDO mostró un mayor contenido en materia orgánica que el purín solo. Las tierras decolorantes son un residuo con muy bajo contenido de humedad, del orden del 2-3%, y, sin embargo un alto contenido de materia orgánica (Tabla 4.3). La mayor parte de materia orgánica está constituida por lípidos, que muestran un valor de la relación DQO/SV muy alta, en torno a 3 g O2/g SV, mientras que para purín está en torno a 1,8 g O2/g SV, tratándose, por tanto, de moléculas con un alto potencial energético. El porcentaje de TDO añadido es sobre la masa total, siendo la fracción de materia orgánica (SV o DQO), aportada por el cosubstrato, mucho más alta. Así pues, con la introducción de TDO como cosubstrato se produce un importante aumento de materia orgánica, lo que supone un incremento de la relación C/N, a pesar de que la concentración de nitrógeno apenas varía. Este es uno de los posibles efectos sinérgicos de la mezcla. La concentración de DQO total, los sólidos totales y volátiles del influente aumentan al introducir TDO como cosubstrato (Figura 6.43 y Figura 6.44), a pesar de que la concentración del purín disminuyó en estos períodos (Figura 6.36). La DQO soluble del influente no aumentó significativamente con la introducción del cosubstrato, indicando que las moléculas orgánicas, básicamente lípidos, están adsorbidas sobre la superficie de las tierras y no pasan a la solución hasta que no actúen sobre ellas las bacterias hidrolíticas. La DQO soluble estuvo básicamente compuesta por los AGV. Optimización de la digestión anaerobia de purines de cerdo TDO 1% 140 120 TDO 2,5% 271 TDO 5% Cambio en la agitación g /kg 100 80 60 40 20 0 0 10 20 ST Influente 30 40 ST Efluente 50 60 SV Influente 70 80 SV Efluente 90 Día Figura 6.43. Evolución de ST y SV del influente y efluente del reactor 3. Los diferentes períodos corresponden a cambios en la velocidad de carga orgánica, VCO (g SV/L·d) TDO 1% 140 TDO 2,5% TDO 5% DQO (mg O2/L) 120 100 80 60 40 20 0 0 10 20 DQO Influente 30 40 DQO Efluente 50 60 DQOs influente 70 80 90 DQOs efluente Día Figura 6.44. Evolución de la DQO total y soluble en el influente y efluente del reactor 3. La eliminación de DQO se ha determinado como la media de cada período, y se muestran en la Tabla 6.12. La eliminación de DQO disminuye a partir del día 30, debido al cambio del régimen de agitación, de forma que en el último período el porcentaje de eliminación de DQO es menor que el índice 272 Ensayos de viabilidad de codigestión en continuo de metanización (Figura 6.45). Estas discrepancias se explican por el error cometido en la medida de la eliminación de materia orgánica, con acumulación de sólidos en el reactor en el primer período y salida del efluente más concentrado al incrementar el régimen de agitación. Tabla 6.12. Eliminación de materia orgánica en el reactor 3, calculada por períodos. Período Alimentación EST (%) ESV (%) EDQO (%) 0-16 Purín 44.75 51.80 55.78 17-36 Purín + 1%TDO 36.75 42.66 37.10 36-57 Purín + 2.5%TDO 20.00 23.81 34.09 58-82 Purín + 5%TDO 12.62 13.99 24.96 Media exp. 25.87 29.90 35.64 Media global 37.08 39.86 48.97 Índices de metanización y acidificación. Los índices de metanización y acidificación fueron muy similares durante el período inicial, en el que se alimentó con purín, indicando que el proceso se producía adecuadamente (Figura 6.45). TDO 1% TDO 2,5% TDO 5% 70% 3.52 3.53 60% 3.31 3.71 % DQO 50% 40% 30% 20% 10% 0% 0 10 20 30 40 % M etanización 50 60 70 % Acidificación 80 90 Día Figura 6.45. Evolución índices de acidificación y metanización para el reactor 3. Los diferentes períodos corresponden a cambios en la velocidad de carga orgánica, VCO (g SV/L·d), valores indicados en la figura Optimización de la digestión anaerobia de purines de cerdo 273 Es curioso observar que al disminuir el TR, en el día 0, no hubo una disminución del índice de metanización, sino un ligero aumento, indicando que la hidrólisis aumentó ligeramente con la disminución del TR, es decir con el aumento de la VCO. A partir del momento en que se comenzó a aportar TDO como cosubstrato, se produjo una cierta separación de los índices debido a la acumulación de AGV, aunque al final del proceso volvió a tener tendencia a aproximarse, situándose el índice de metanización por encima del 40% y el de acidificación por encima del 45%. 6.6.4. Codigestión con tierras decolorantes de aceite de oliva - Rango termofílico (R4). El reactor 4 se utilizó para realizar los experimentos de codigestión con tierras decolorantes de aceite de oliva en el rango termofílico. Desde el día -1 el tiempo de retención de trabajo ha sido de 15 días, pudiendo, por tanto, ser comparado con el control (reactor 1). Evolución de la producción de gas. El proceso de digestión anaerobia en rango termofílico de la mezcla de purín con tierras decolorantes de aceite de oliva no dio buenos resultados, confirmando los resultados obtenidos en los experimentos en discontinuo (capítulo 5). Como en el reactor 3, la puesta en marcha se realizó con un tiempo de retención de 20 días, y el día -1 se cambió el tiempo de retención de 20 a 15 días. Contrariamente a lo ocurrido en el caso anterior, el cambio en el tiempo de retención coincidió con una acusada bajada en la producción volumétrica de gas, tal y como se observa en la Figura 6.46. Bajó la producción volumétrica y la producción por unidad de carga, así como el contenido de metano en el biogás. A partir del día 16 comenzó la adicción de tierras decolorantes de aceite de oliva a un nivel del 1%. Esto pareció estimular ligeramente la producción de gas, aunque no aumentó la concentración de metano. Al final del período de alimentación con TDO 1%, la producción de biogás volvió a bajar. Al introducir un 2,5% de TDO la concentración de metano en el biogás mostró una tendencia a disminuir, llegando a niveles extraordinariamente bajos (entre el 10 y 20%). Esto hecho, junto con la fuerte acumulación de ácidos grasos volátiles (Figura 6.49), hizo que se optara por parar la alimentación del reactor a partir del día 43, para evitar la completa acidificación del mismo. Sin embargo, el proceso de inhibición pareció ser 274 Ensayos de viabilidad de codigestión en continuo irreversible, y sólo al final del experimento pareció experimentar cierta ligera mejora, aunque no llegó a superar el 25% de contenido de metano. TDO 1% Pv y Pc 1.00 3.52 0.90 3.54 0.80 0.70 0.60 0.50 0.40 0.30 0.20 0.10 0.00 0 10 20 TDO 2,5% 30 Pv (L de Biogas/L*d) 3.33 0.55 40 50 0.43 60 Pc (L de CH4 /g SV) 70 % CH4 100% 90% 80% 70% 60% 50% 40% 30% 20% 10% 0% 80 90 día % de CH4 Figura 6.46. Evolución de la producción volumétrica de biogás (Pv), producción por unidad de carga y concentración de metano en el biogás, en el reactor 4. Los diferentes períodos corresponden a cambios en la VCO (g SV/d),valores indicados en el gráfico. Evolución de las formas nitrogenadas en el reactor 4. La concentración de N-NH4+ del efluente (del líquido mezcla) aumentó de forma muy acusada desde el inicio del experimento, hasta superar los 5 g NNH4+/L el día (Figura 6.47). El aumento se debió, al incremento gradual de la concentración en el influente. La concentración de amoníaco libre se mantuvo durante todo el experimento por encima de los 650 mg N-NH3/L, con picos de más de 1000 mg N-NH3/L. Evolución de la alcalinidad y ácidos grasos volátiles La alcalinidad total se mantuvo durante todo el experimento por encima de 15 g CaCO3/L, mientras que la parcial, asociada al bicarbonato se mantuvo entre 7,5 y 12 g CaCO3/L. Estos valores son más bajos que los encontrados en los otros reactores, aunque están por encima de los límites propuestos en la bibliografía (Fannin, 1987). La relación de alcalinidad (RA) aumentó a partir del día -1, momento en que se redujo el tiempo de retención de 20 a 15 días, y continuó aumentando hasta alcanzar valores por encima de 0,5, muy por encima de los límites propuestos en la literatura para diagnosticar la Optimización de la digestión anaerobia de purines de cerdo 275 presencia de problemas en un reactor (Iza, 1995). TDO 1% 3.54 7000 TDO 2,5% 3.52 3.33 0.55 0.43 6000 mg N/L 5000 4000 3000 2000 1000 650 0 0 10 20 Nk inf. NH4+ ef. 30 40 50 60 Nk ef. 55ºC NH3 70 80 90 NH4+ inf. Día Figura 6.47. Evolución de nitrógeno Kjeldahl y amoniacal del influente y efluente en el reactor 4. Concentración de amoníaco libre en el efluente. El pH, inicialmente, se situó en torno a 8, pero con el descenso del tiempo de retención bajó progresivamente hasta situarse en valores cercanos a 7,5, manteniéndose en ese nivel hasta el final del experimento (Figura 6.48). La bajada del pH se relacionó con la fuerte acumulación de AGV (Figura 6.49). 8.5 pH efluente 8 7.5 7 6.5 0 20 40 60 80 Día Figura 6.48. Evolución del pH del efluente del reactor 4. El descenso de la producción de metano al reducir el TR de 20 a 15 días, (período inicial) pudo estar motivado conjuntamente por el aumento de amonio, y de amoníaco libre (aumentó también el pH), y por el efecto de 276 Ensayos de viabilidad de codigestión en continuo incremento de la carga orgánica. Las sobrecargas orgánicas se caracterizan por la acumulación de hidrógeno en el sistema, que provoca la rápida acumulación de AGV de más de 3C, especialmente propiónico (Griffin et al., 1998). Aunque la concentración de propiónico y otros AGV aumentó, el principal efecto observado fue la acumulación de acético (Figura 6.49). Por ello, la causa más probable de la inhibición de la metanogénesis es el aumento de concentración de amonio y/o amoníaco libre, muy por encima del nivel límite propuesto por Angelidaki y Ahring (1993b). mg AGV /L TDO 1% P/A TDO 2,5% 12000 0.8 3.52 3.54 3.33 0.55 0.43 10000 0.6 8000 6000 0.4 4000 0.2 2000 0 0.0 0 mg AGV /L 10 20 TDO 1% 3.52 3.54 40 50 60 70 80 90 70 80 90 Día TDO 2,5% 2000 1750 30 3.33 0.55 40 50 0.43 1500 1250 1000 750 500 250 0 0 10 20 30 60 Acético Propiónico Prop/acet Iso Butírico N Butírico Iso Valérico Día N Valérico Figura 6.49. Evolución de la concentración de AGV en el efluente del reactor 4. Los diferentes períodos corresponden a cambios en la velocidad de carga orgánica, VCO (g SV/L·d), valores indicados en la figura. Optimización de la digestión anaerobia de purines de cerdo 277 A partir del día 16, con el comienzo de la adición de tierras decolorantes de aceite de oliva a un nivel del 1%, se observó una tímida recuperación de la producción de gas (Figura 6.46), pero la concentración de AGV continuó aumentando, especialmente la de acético. El día 37 se aumentó la proporción de TDO hasta el 2,5%, observándose una disminución aún mayor de la concentración de CH4, la producción de biogás se estabilizó por debajo de 0,3 L/L*d, y la concentración total de AGV continuó aumentando hasta llegar a los 15 g Ac/L (Figura 6.49). La relación P/A se mantuvo a niveles bajos durante todo el experimento. El día 43 se decidió incrementar el tiempo de retención y substituir el substrato por purín para intentar la recuperación del reactor. Hasta el día 82 se mantuvo con una muy baja alimentación, la mínima necesaria para poder extraer una muestra y realizar el control, manteniendo un tiempo de retención hidráulico de 99 días. A pesar de todo, no se consiguió la recuperación del reactor, manteniéndose el nivel de AGV entre 10 y 13 g Ac/L, hasta el final del experimento. Si bien la relación propiónico/acético se mantuvo a niveles bajos durante el experimento, la concentración de propiónico alcanzó niveles muy importantes, superando los 2000 mg Pr/L. Los ácidos butírico y valérico aumentaron mucho durante el experimento, siguiendo una evolución similar a la del acético y propiónico. Evolución de los parámetros de materia orgánica: influente y efluente. Los parámetros relacionados con la materia orgánica, DQO y SV, en el efluente aumentaron de forma importante a partir del día 30. La concentración superó a la del influente, observándose eliminaciones negativas. La causa fue el deficiente régimen de agitación operado durante los 30 primeros días y durante la puesta en marcha, que hizo que se acumularan sólidos en el reactor. Al observar el fenómeno y aumentar el régimen de revoluciones del agitador se produjo el espectacular aumento en la concentración del efluente, y una gran disminución de la eliminación de materia orgánica, que alcanzó valores negativos (Tabla 6.13). Esto fue similar a lo ocurrido, y descrito en el reactor 1 y en menor medida en los reactores mesofílicos, reactor 2 y 3. En los reactores termofílicos el proceso de acumulación de sólidos en el reactor fue más importante que en los mesofílicos, puesto que el incremento de la DQO y SV del efluente es mucho más importante en los primeros. 278 Ensayos de viabilidad de codigestión en continuo 180 3.54 TDO 1% 3.52 TDO 2,5% 3.33 0.55 0.43 160 DQO(g O2 /kg) 140 120 100 80 60 40 20 0 0 10 DQO influente 20 30 DQO efluente 40 50 60 DQOs influente 70 80 DQOs efluente 90 Día Figura 6.50. Evolución de la DQO total y soluble del influente y el efluente, reactor 4. Los diferentes períodos corresponden a cambios en la velocidad de carga orgánica, VCO (g SV/L·d), valores indicados en la figura. Tabla 6.13. Eliminación de materia orgánica en el reactor 4 calculada por períodos Alimentación EST (%) ESV (%) EDQO (%) 0-16 Purín 53.60 58.40 52.22 17-36 Purín + 1%TDO 22.83 26.75 15.73 36-57 Purín + 2.5%TDO 0.28 -1.94 -17.18 58-82 Purín -64.67 -42.47 -98.02 Media experimento -4.41 3.98 -21.25 Media global 17.33 25.49 19.90 Período La DQO soluble del efluente se mantuvo aproximadamente constante desde el día 30, aproximadamente a un nivel de 16-18 gO2/L, indicando que una importante fracción de la materia orgánica se encontraba en forma de compuestos solubles, y no pudieron ser degradados. La mayor parte de la DQO soluble está constituida por los ácidos grasos volátiles. Optimización de la digestión anaerobia de purines de cerdo 120 3.54 100 TDO 1% 3.52 279 TDO 2,5% 3.33 0.55 0.43 Cambio en la agitación g/kg 80 60 40 20 0 0 10 20 ST influente 30 40 ST efluente 50 60 SV influente 70 80 SV efluente 90 Día Figura 6.51. Evolución de los ST y SV del influente y efluente del reactor 4. Los diferentes períodos corresponden a cambios en la velocidad de carga orgánica, VCO (g SV/L·d), valores indicados en la figura. Índice de metanización y acidificación. El índice de metanización disminuyó mucho al aumentar el contenido en amonio, manteniéndose bajo al añadir el cosubstrato TDO. Sin embargo el índice de acidificación subió al final situándose en valores cercanos al 50% (Figura 6.52). Esto se debió a la parada en la alimentación y el aumento del tiempo de retención, por lo que los índices de transformación fueron mayores, a pesar de que el sistema es mucho menos eficiente. Por el mismo motivo, el índice de metanización también aumentó al final del experimento. La gran diferencia observada entre los índices de metanización y acidificación indica que la fase metanogénica fue más afectada que la acidogénica (Figura 6.52). A pesar de ello, la acidogénesis resultó inhibida, dados los bajos valores obtenidos, por debajo del 30% durante la mayor parte del experimento. 280 Ensayos de viabilidad de codigestión en continuo TDO 2,5% TDO 1% 60% 50% 3.52 3.54 3.33 0.55 0.43 % DQO 40% 30% 20% 10% 0% 0 10 20 30 40 % M eta 50 60 70 % acidifc 80 90 Día Figura 6.52. Evolución de los índices de acidificación y metanización en el reactor 4. 6.6.5. Inhibición por amonio. Influencia de la temperatura. La digestión anaerobia de purines de cerdo en el rango termofílico ha resultado poco viable de acuerdo con los resultados obtenidos en el presente experimento. El proceso ha fracasado debido, fundamentalmente, al mal funcionamiento de la etapa metanogénica acetoclástica, como indica la gran acumulación de ácido acético observada. El principal parámetro con influencia en el proceso parece haber sido la alta concentración de nitrógeno amoniacal. Ya se ha citado en el presente documento la importancia del amonio como inhibidor de la metanogénesis (Hashimoto, 1986; Robbins et al., 1989; Gallert et al., 1998, Angelidaki y Ahring, 1993b, 1994; Hansen et al., 1998, 1999; Campos et al., 1999), aunque el límite de inhibición varía mucho en función del trabajo, debido básicamente al nivel de adaptación del cultivo (van Velsen, 1979; Hashimoto, 1986). Los resultados obtenidos en el presente experimento, en el que la producción media para el reactor 1 (termofílico) fue de 53 mL de CH4/g SV, y la media del contenido de metano en el biogás de 36,25%, son más bajos que resultados encontrados en la bibliografía con valores muy altos de amonio (Hansen et al., 1998), lo que constata la importancia de la adaptación al tipo de substrato. La altas concentraciones de nitrógeno amoniacal y amoníaco libre inhiben, principalmente, la fase metanogénica acetoclástica (Angelidaki y Ahring, 1993b; Hansen et al., 1998). Otros procesos, como la metanogénesis Optimización de la digestión anaerobia de purines de cerdo 281 hidrogenotrófica y la hidrólisis de proteínas, pueden estar afectados por amonio, pero requieren concentraciones más altas de tóxico para su inhibición (Angelidaki y Ahring, 1993b; Gallert et al., 1998; Hansen et al., 1998). Por tanto, la presencia de amonio provocará, principalmente, la acumulación de acético en el líquido mezcla. Así, la concentración de acético junto con la producción de gas, son los principales parámetros de diagnóstico de inhibición de la metanogénesis acetoclástica, y han sido utilizados para estudiar el efecto del amonio por otros autores (Zeeman et al., 1985). Puesto que se dispone de gran cantidad de datos de concentración de amonio, amoníaco, ion amonio y concentración de acético, se estudiaron las correlaciones entre dichos parámetros, obteniéndose resultados interesantes, diferentes según la temperatura del reactor. En los dos reactores termofílicos se obtuvieron buenos ajustes entre el nitrógeno amoniacal total y la producción volumétrica de metano, mientras que entre la forma libre, NH3, y la producción de metano, no se encontró ninguna relación estadísticamente significativa. La relación entre la concentración del ion amonio y amonio total con el acético fue significativa para los reactores 1 y 4 a un nivel de significación del 99%, obteniendo muy altos coeficientes de regresión para el modelo lineal, por encima de 0,94 (Figura 6.53). Al correlacionar la producción volumétrica de metano con la concentración de amonio la relación, pese a ser significativa, mostró peores ajustes que para el acético. El modelo que mejor se ajustó fue el modelo exponencial: Pv exp (a b ), NH 4 (6.11) considerando sólo los puntos valores a partir de la concentración de amonio de 3000 mg N-NH4+/L. Por debajo de esta concentración no se observó correlación entre ambos parámetros. Según se observa en la Figura 6.53, a partir de 3500- 4000 mg N-NH4+/L, la disminución de la producción de gas, al aumentar la concentración de amonio es menor que para menores concentraciones de amonio, indicando que, probablemente, la ruta metanogénica acetoclástica está completamente inhibida, y la formación de metano residual es debida a otras rutas metanogénicas, principalmente la hidrogenotrófica, que aunque es sensible a la inhibición por amonio, muestra una mayor tolerancia (Angelidaki y Ahring, 1993b; Hansen et al., 1998). El acético, no obstante, continúa 282 Ensayos de viabilidad de codigestión en continuo acumulándose, indicando que la acetogénesis no se inhibe por amonio. A altas concentraciones de amonio parece cambiar la tendencia, debido a que las altas concentraciones de acético en el reactor sí inhiben la acetogénesis. mg Ac/L 12000 mL de CH4 /L*d Pv=EXP(0.34+18346/NH4 +) 800 R2 =0.873 700 2 10000 R = 0.9411 600 8000 500 400 6000 300 2 R = 0.947 4000 200 2000 100 Pv=EXP(1.77+12236/NH4 +) R2 =0.776 0 0 0 1000 2000 3000 4000 5000 6000 + mg N-NH4 /L 0 R1-CH4 R1-acético R4-CH4 1000 2000 3000 4000 5000 6000 + mg N-NH4 /L R4-acético Figura 6.53. Estudio de regresión de la concentración de acético y la producción volumétrica de metano con la concentración de nitrógeno amoniacal . Del análisis conjunto de todos los datos del rango termofílico, reactores 1 y 4, se observa un claro cambio de tendencia en torno a la concentración de 3000 mg N-NH4+/L. Para valores de amonio por debajo de 3000 mg NNH4+/L, se obtuvo una buena correlación entre acético y amoníaco libre, especialmente considerando un modelo exponencial, lo que indicaría que a concentraciones bajas la forma tóxica sí puede ser el NH3. A pesar de la acumulación de acético, por debajo de 3000 mg N-NH4+/L, no hubo una correlación significativa entre la producción volumétrica de metano y la concentración de metano en el biogás y la concentración de amonio, o amoníaco libre. Una posible explicación al mal ajuste obtenido para el amoníaco libre, supuesto el auténtico tóxico para los microorganismos metanogénicos (Hashimoto, 1986; Angelidaki y Ahring, 1993b), es que la concentración de NH3, cuando la concentración de amonio fue superior a 3000 mg NNH4+/L, se mantuvo a niveles muy altos, por encima de los 600 mg NNH3/L. Así la inhibición por amonio dependería del pH, siempre y cuando la concentración de amoníaco libre estuviera por debajo de este nivel, o la del Optimización de la digestión anaerobia de purines de cerdo 283 amonio por debajo de 3000 mg N-NH4+/L. Estos resultados están de acuerdo con los obtenidos en estudios antiguos (McCarty, 1964, citado en van Velsen, 1979). Sin embargo otros autores han encontrado respuesta del sistema hasta una concentración de amoníaco libre de 1,1 g N-NH3/L (Hansen et al., 1998). Las diferencias podrían atribuirse al diferente grado de aclimatación del inóculo. El límite establecido por Hill para el correcto funcionamiento de un digestor anaerobio de 800 mg Ac/L (Hill et al., 1987), coincide con una concentración de amonio de 2400 mg N-NH4+/L. A partir de 3000 mg NNH4+/L, la pendiente de la curva aumenta, indicando que con menores incrementos de concentración se acumula más acético. Para los datos de los reactores en mesofílico los coeficientes de regresión obtenidos fueron más bajos, pese a ser significativa la relación entre acético y todas las formas de nitrógeno amoniacal a un nivel del 99%. Los máximos coeficientes de regresión se obtuvieron utilizando un modelo exponencial, pero siempre estuvieron por debajo del 75% para el reactor 3 y del 60% para el reactor 2. El ajuste al modelo lineal proporcionó coeficientes de regresión del orden del 50% para la variable amonio total y del 30% para el amoníaco libre en los dos casos estudiados. Los niveles de acético alcanzados fueron, en todo caso, menores en el rango mesofílico que en el termofílico, sin que se detectaran situaciones claramente inhibidas, con reducción de la producción de biogás y de la concentración de metano. En el reactor 3, de codigestión con TDO se observó un importante aumento de acético al añadir un 2,5% de cosubstrato, que se sale completamente de la tendencia del reactor alimentado con purín sólo, indicando que la adicción del cosubstrato tuvo un efecto inhibitorio sobre la metanogénesis. 6.6.6. Producción de gas según los diferentes regímenes de alimentación Se ha realizado el análisis estadístico de los datos de producción de gas para los diferentes períodos en los que se ha dividido el experimento. Se han considerado los valores de los 5 últimos días de cada período, considerado como representativos del estado estacionario de cada período, para ello se puso la condición de un coeficiente de variación de la producción por unidad de carga menor del 10% (Cobb y Hill, 1991). Esta condición se aplicó a los reactores mesofílicos, ya que en los termofílicos la variabilidad fue mucho más alta. Se han comparado los resultados obtenidos para los diferentes reactores, correspondientes a la producción volumétrica de biogás, de 284 Ensayos de viabilidad de codigestión en continuo metano y producción de metano por unidad de carga, en unidades de sólidos volátiles. Los períodos considerados son los expuestos en la Tabla 6.5. A partir del tercer período, y para todas las variables, la interacción entre el efecto codigestión y temperatura resultó significativa, por lo que se realizó el análisis de la interacción mediante el test Lsmeans. En la Tabla 6.14 se muestran los valores medios obtenidos para cada reactor y cada período. Se realizó, también, la separación de medias de los efectos principales (datos no mostrados), mediante el test Duncan. Tabla 6.14. Producción volumétrica de metano (L CH4/L·d) y producción por unidad de carga (LCH4/g SVin). Medias para cada período, y cada reactor. TDO 0% TDO 1% TDO 2,5% TDO 5% 1 (1-16 d) 2 (17-36) 3 (37-57) 4 (58-82) Prod. volumétrica R1-55ºC control 0.19 C 0.26 B 0.20 C 0.43 C (Pv) de biogás R2-35ºC control 1.47 A 1.62 A 1.03 B 0.92 B (L biogás/L*día) R3-35ºC codig. 1.41 A 1.55 A 1.47 A 1.91 A R4-55ºC codig. 0.37 B 0.38 B 0.12 C 0.28 D Prod. volumétrica R1-55ºC control 0.09 b 0.08 b 0.06 c 0.12 c (Pv) de metano R2-35ºC control 1.02 a 1.10 a 0.71 b 0.60 b (L CH4/L*día) R3-35ºC codig. 1.02 a 1.13 a 1.03 a 1.34 a R4-55ºC codig. 0.11 b 0.09 b 0.01 c 0.05 d Prod. por unidad de R1-55ºC control 0.12 B 0.02 B 0.02 C 0.04 D carga (Pc) de metano R2-35ºC control 0.30 A 0.31 A 0.25 B 0.21 B (L CH4/g SV) R3-35ºC codig. 0.29 A 0.32 A 0.31 A 0.37 A R4-55ºC codig. 0.03 C 0.03 B 0.03 C 0.12 C Letras diferentes significan diferencias significativas a un nivel de significación del 5% (Test de Duncan D=0.05). El análisis estadístico se ha realizado para cada período considerado (columnas), y para cada variable Pv biogás, Pv metano, Pc. Período 1. Durante el primer período las condiciones experimentales fueron las mismas en los reactores de codigestión (3 y 4) que en los controles (1 y 2), ya que fue a partir del segundo período cuando se comenzó a adicionar el cosubstrato. El efecto codigestión, por tanto, no fue significativo para ninguna de las variables analizadas. Sin embargo, sí se detectaron diferencias significativas entre temperaturas. Todas las variables analizadas fueron significativamente mayores en el rango mesofílico que en el termofílico, que estuvo fuertemente influido por la concentración de amonio (apartado 6.6.5). Las diferencias observadas entre los dos reactores termofílicos, R1 y R4, se debieron a que el reactor 1 sufrió una variación en la carga orgánica (Apartado 6.6.1), aumentando de forma considerable el TR (Figura 6.9). La Optimización de la digestión anaerobia de purines de cerdo 285 producción volumétrica de biogás fue más alta en R4, que mantuvo el tiempo de retención de 15 días, mientras que la producción por unidad de carga fue significativamente mayor en el reactor 1, por la baja carga aportada. En los dos reactores mesofílicos no se observaron diferencias significativas, pese a que el reactor 3 llevaba poco tiempo operando con el tiempo de retención de 15 días, y pese a las diferentes estrategias de puesta en marcha. Período 2. En este período comenzó la introducción de TDO, al 1%. Al final de este período (días 31-36) el efecto codigestión no fue significativo, es decir, no se pudo diferenciar, dentro de cada temperatura, los tratamientos con y sin cosubstrato. Sí se mantuvieron las diferencias entre temperaturas. Sí se observó, sin embargo, un aumento en la concentración de AGV en el reactor 3, tanto de acético como de propiónico (Figura 6.42), lo que indica la respuesta del sistema. La justificación puede estar en el poder inhibidor de los ácidos grasos de cadena larga (capítulo 5; Hanaki et al., 1981; Angelidaki y Ahring, 1992; Hwu et al., 1997), que habría impedido la formación adicional de metano. A pesar de este fenómeno inhibitorio de la etapa metanogénica y especialmente de la acetogénica, la producción de metano no fue menor que en el control (reactor 2), debido al mayor aporte de materia orgánica con un alto potencial de producción de metano. Período 3. Durante este período se incrementó la fracción de TDO de la mezcla a un 2,5%. La evolución del reactor 4, termofílico, en el que la acumulación de AGV alcanzó niveles extraordinariamente altos (Figura 6.49), hizo que se tomara la decisión de parar la alimentación para evitar la completa acidificación del mismo (apartado 6.6.4). Todos los efectos considerados, codigestión, temperatura y la interacción de ambos fueron significativos con una probabilidad < 0,01%. Dentro del rango mesofílico la producción del reactor 3 fue significativamente mayor que la del reactor 2 (control de purín), tanto para la producción volumétrica de biogás y metano, como para la producción por unidad de carga (Pc). Sin embargo, los resultados del control fueron peores que los del período anterior, debido, fundamentalmente, al aumento de la concentración de amonio (Figura 6.33), y la consiguiente inhibición de la fase metanogénica, como se deduce también de la acumulación de acético (Figura 6.35; apartado 6.6.5). La producción de metano y biogás del reactor 3 disminuyó levemente (Figura 6.39), respecto al período anterior, coincidiendo con una fuerte acumulación de AGV, especialmente de acético, y en menor medida de propiónico, butírico y valérico, y de amonio (Figura 6.42 y Figura 6.40). La concentración de AGV de más de tres átomos de carbono se estabilizó a un nivel relativamente alto, lo que indicaría la existencia de inhibición de la 286 Ensayos de viabilidad de codigestión en continuo acetogénesis y probablemente de la metanogénesis hidrogenotrófica por AGCL. Período 4. La proporción de TDO de la mezcla introducida en el reactor 3 fue de un 5%, proporción que había resultado óptima en los ensayos en discontinuo, realizados previamente (capítulo 4). En el rango termofílico no se intentó esta proporción, dados los malos resultados obtenidos con proporciones menores. Tanto las variables codigestión, temperatura, como la interacción de ambas, resultaron significativas a un nivel de confianza del 0,01%. Dentro del rango mesofílico hubo diferencias entre el tratamiento con TDO (reactor 3) y el control (reactor 2), tanto para la variable producción volumétrica de biogás y de metano, como para la producción por unidad de carga (Tabla 6.14). La producción volumétrica casi duplicó la obtenida por el control, que continuó una tendencia descendente (Figura 6.39), debido a la disminución de la velocidad de carga (Figura 6.9). La producción de metano por unidad de sólidos volátiles del reactor 3 (con TDO) aumentó, respecto al período anterior, ampliando, por tanto, las diferencias respecto al control, llegando a valores bastante altos, por encima de del límite propuesto por Hill et al. (1987), y por encima de los obtenidos con residuos ganaderos por otros autores (Angelidaki y Ahring, 1993b), aunque mucho menores que los obtenidos con otros tipos de residuos, como los residuos de matadero (Salminen et al., 1999) o de fracción orgánica de residuos sólidos urbanos (Pavan et al., 1999). El mantenimiento de una concentración alta y un nivel aproximadamente constante de AGV como el propiónico al final del experimento (Figura 6.42), es indicador de la fase acetogénica parcialmente inhibida. Es de suponer, que con el tiempo, el nivel de inhibición debería disminuir, ya que no se observó un importante aumento de AGV al pasar de 2,5% al 5%, indicando el progreso adecuado del proceso. En cuanto al rango termofílico, se detectaron diferencias, entre los dos reactores, pero la causa fue el diferente tiempo de retención y nivel de carga. El reactor 4, a pesar de la bajada de carga experimentada, mantuvo la concentración de AGV a niveles muy altos, por encima de los obtenidos para el reactor 1, especialmente el nivel de acético. La relación P/A se mantuvo a un nivel bajo, a pesar de que el propiónico estuvo en torno a 2000 mg Prop/L (Figura 6.49). La producción volumétrica de gas fue menor en el reactor 4 que en el reactor 1, mientras que la producción por unidad de carga fue mayor en el 4 que en el 1, fruto de la menor velocidad de carga. Optimización de la digestión anaerobia de purines de cerdo 287 6.6.7. Diferencias en los niveles de acidificación. Comparación de la eficiencia de la hidrólisis-acidogénesis. Los resultados obtenidos al final del proceso muestran unos mayores índices de acidificación que de metanización, con diferencias especialmente importantes entre temperaturas. La digestión anaerobia de purines en el rango mesofílico no mostró diferencias importantes entre el índice de metanización y el de acidificación, durante las primeras etapas y sólo cuando aumentó el nivel de amonio, se incrementaron las diferencias, aunque estas siempre fueron mucho menores que en el rango termofílico (Figura 6.38). Esto indica que el limitante en el rango mesofílico no fue la metanogénesis, sino la hidrólisis-acidogénesis. En el rango termofílico el índice de metanización se mantuvo bastante bajo durante todo el experimento, observándose pequeñas subidas en los períodos en los que se aumentó el tiempo de retención (Figura 6.31 y Figura 6.52). La gran diferencia entre el índice de acidificación y metanización indican que la fase limitante del proceso fue la fase metanogénica. Sin embargo, el índice de acidificación en los reactores termofílicos se mantuvo más bajo que en los reactores mesofílicos, lo que indica que el proceso hidrolítico-acidogénico resultó afectado. En el experimento de codigestión con tierras decolorantes, la diferencia entre los índices de metanización y acidificación se incrementó conforme aumentó el porcentaje de tierras añadidas. Sin embargo, al final del experimento, experimentó un proceso de adaptación disminuyendo la diferencia (Figura 6.45). Al contrario que lo ocurrido en el rango termofílico la fase más afectada pudo haber sido la fase acetogénica, dados los altos niveles de propiónico encontrados en el reactor. 6.6.8. Influencia de la VCO sobre la producción de biogás. En el reactor 2, control a 35ºC, se observó durante los períodos 3 y 4, una bajada en el nivel de producción volumétrica de biogás y en la producción de metano por unidad de carga (Figura 6.32). La disminución de la producción volumétrica de biogás y de metano se puede relacionar directamente con la disminución de carga orgánica, y, de hecho se obtuvieron buenos coeficientes de regresión (Figura 6.54a). Sin embargo, la relación entre la producción de metano por unidad de carga y la VCO para el reactor 2, alimentado exclusivamente con purín, fue, también, significativa, a un nivel de confianza del 95% (Figura 6.54b). El reactor 3 mostró una relativamente buena correlación entre las dos variables, aunque peor que el reactor 2, sólo significativa a un nivel de confianza del 288 Ensayos de viabilidad de codigestión en continuo 10%, indicando que otros factores pudieron haber intervenido. 0.40 2.5 2 1.5 Pc (L CH4/g SV*d) Pv (L Biogas/L*d) 0.35 2.0 2 R = 0.8114 2 R = 0.9811 1.0 0.5 R = 0.7461 0.30 2 0.25 R = 0.9274 0.20 0.15 0.10 0.05 0.00 0.0 2 2.5 3 3.5 2 4 2.5 VCO (g SV/L*d) R2-Pv 3 3.5 4 VCO (g SV/L*d) R3-Pv R2- Pc R3-Pc Figura 6.54. Correlación de la producción volumétrica de biogás y de la producción de metano por unidad de carga respecto a la velocidad de carga orgánica. El efecto de incremento de producción de gas por unidad de carga al aumentar la velocidad de carga orgánica ha sido observada por otros autores. Parece que el incremento de la velocidad de carga orgánica tiene un efecto estabilizador sobre el proceso biológico, junto con una mayor resistencia a procesos de toxicidad e inhibición por amonio (Ahring et al., 1995). Así los citados autores observaron que al aumentar la velocidad de carga orgánica de 6.3 a 10.3 g SV/L*d, se incrementó tanto la producción volumétrica de metano como la producción de metano por unidad de carga. En estudios con purín sólo, el incremento de la producción por unidad de carga, sin embargo, no ha quedado demostrado, así Hill et al. (1987), observaron que se producía una disminución de la producción por unidad de carga a partir de un valor de VCO de 6,86 g SV/L*d. En el presente estudio se observó un importante efecto beneficioso del incremento de VCO entre 2,5 y 4 g SV/L*d sobre la producción de gas. 6.6.9. Comparación de la eficiencia de la hidrólisis entre los diferentes reactores, mediante el uso del parámetro NH4+/Nk. La concentración de nitrógeno amoniacal fue más alta en el reactor 1, termofílico, que en el 2, mesofílico, ambos alimentados con el mismo tipo de purín. Esta diferencia podría haberse debido a la mayor tasa de hidrólisis en Optimización de la digestión anaerobia de purines de cerdo 289 el rango termofílico. La velocidad de los procesos biológicos en general (Pavlostathis y Giraldo-Gómez, 1991), y en especial la tasa de hidrólisis de macromoléculas (Veeken y Hamelers, 1999) aumenta con la temperatura. Se ha utilizado como parámetro de diagnóstico la relación de nitrógeno amoniacal sobre el nitrógeno total, tanto en el influente como en el efluente, para los cuatro reactores (Tabla 6.15). Analizando los resultados del período de puesta en marcha para los reactores 3 y 4, alimentados con mezcla de lodos de EDAR y purín, se observa que la fracción de amonio sobre el Nitrógeno Kjeldahl aumentó más que en los reactores alimentados con purín sólo. El bajo nivel de la fracción NH4+/Nk, y sobre todo la baja concentración de nitrógeno amoniacal influyó en la hidrólisis de nitrógeno orgánico. El incremento pudo haber sido aún mayor debido al tiempo de retención más largo, empleado en estos reactores. Tabla 6.15. Fracción de nitrógeno amoniacal sobre el Kjeldahl, e incremento de dicha fracción durante la digestión. Valores medios de todo el período de ensayo. Día 0-102 Influente Efluente Incremento R1- 55ºC 75.05% r3.69% 82.27% r4.28% 7.22% R2- 35ºC 74.33% r3.81% 79.18% r3.03% 4.85% R3- 35ºC 74.44% r5.33% 77.06% r4.88% 2.62% R4- 55ºC 73.89% r5.16% 79.34% r4.41% 5.45% Período de puesta en marcha con mezcla de lodos de EDAR y purín R3 – 35ºC 35.87% r5.10% 62.08% r8.05% 26.21% R4 – 55ºC 35.14% r5.13% 65.36% r3.39% 30.23% Se observa también, aunque menos importante que en el período de puesta en marcha, una mayor hidrólisis de compuestos nitrogenados en el rango termofílico que en el mesofílico. Resultados similares se obtuvieron en el caso de los ensayos en discontinuo, en los que hubo un mayor incremento en el rango termofílico, siendo, además mayor cuanto menor era la concentración inicial de amonio (capítulo 5). Estos resultados confirman los resultados de Gallert et al. (1998) que encontraron que había una disminución en la fracción de peptona hidrolizada en función de la concentración de amonio del medio, siendo además mayor en el rango termofílico que en el mesofílico. Puesto que el efecto de inhibición por amonio es mayor en el rango termofílico que en el mesofílico, mayor degradabilidad de las proteínas a 290 Ensayos de viabilidad de codigestión en continuo 55ºC puede contribuir a agravar el problema de inhibición por amonio de los residuos ganaderos en el rango termofílico. 6.6.10. Comparación con los resultados obtenidos en discontinuo y con el modelo de simulación. Los resultados obtenidos en discontinuo y en continuo fueron bastante diferentes, debido, a la diferencia de método y a la variación en la composición del purín utilizado. En cualquier caso, cualitativamente, las conclusiones obtenidas de los experimentos en discontinuo se confirman en los experimentos en continuo. También se han confirmado, aunque sólo de forma cualitativa, los resultados obtenidos en las simulaciones realizadas con el modelo en el capítulo 4. Así, se ha observado cierta sobrecarga del sistema al introducir TDO como cosubstrato. Esta sobrecarga ha sido mayor y más permanente en los resultados experimentales que en los simulados, indicando que la modelización de la inhibición por AGCL no es suficientemente ajustada. De las simulaciones realizadas se puede concluir que la introducción gradual del cosubstrato es fundamental para asegurar el éxito del proceso. Los efectos negativos de acumulación de AGV disminuyen cuanto mayor sea el número de pasos utilizados, más que la duración de los mismos, debido a que si el paso es suficientemente pequeño, la perturbación es pequeña y la recuperación rápida, constituyéndose adecuadamente la población acetoclástica. También se ha observado, de nuevo de forma cualitativa, los efectos negativos en el rango termofílico, de la introducción de un purín con mayor contenido de amonio. Así como la diferencia del grado de inhibición por amonio en función de la temperatura. 6.7. Conclusiones de los ensayos en continuo Las principales conclusiones obtenidas de los resultados de los ensayos en continuo se resumen en los siguientes puntos: 6.7.1. Sobre el método Para la obtención de resultados concluyentes es conveniente mantener las condiciones estables durante períodos más largos que los utilizados en el presente estudio. La excesiva variación en las características de la alimentación ha podido Optimización de la digestión anaerobia de purines de cerdo 291 perjudicar la obtención de resultados, a veces enmascarados por otros efectos. La velocidad de agitación ha de ser suficiente para evitar la acumulación de sólidos en el reactor. Se han constatado diferencias importantes en la eficiencia de la agitación en función de la temperatura, habiéndose observado una mayor acumulación de sólidos en los reactores termofílicos que en los mesofílicos. 6.7.2. Sobre el período de puesta en marcha La puesta en marcha de un reactor anaerobio requiere largos tiempos, si se pretenden conseguir condiciones de adaptación adecuadas. La utilización como inóculo de lodo de depuradora de aguas residuales urbanas, incrementando gradualmente la proporción de purín en la alimentación, es una opción viable para la puesta en marcha con purines de cerdo. Es posible utilizar un inóculo de un reactor mesofílico para la puesta en marcha de un reactor termofílico, recuperándose en pocos días de la perturbación, adaptándose bien a las nuevas condiciones, sin que se observase reducción de la producción de gas. El factor más limitante sobre el proceso de digestión anaerobia de purines de cerdo en el rango termofílico, en la fase de puesta en marcha, es la concentración de amonio. 6.7.3. Sobre la viabilidad de la digestión anaerobia de purines de cerdo La producción de metano es mayor en el rango mesofílico que en el termofílico, tanto en la producción volumétrica como la producción por unidad de carga. En el rango termofílico la producción es muy baja, muy por debajo del umbral de rentabilidad propuesto en la bibliografía. La producción volumétrica y por unidad de carga de metano aumentan, en el rango mesofílico, al aumentar la velocidad de carga orgánica en el rango entre 2,5 y 4 g SV/L*d. En general, en ausencia de inhibidores, la etapa limitante de la digestión anaerobia de purines de cerdo en el rango mesofílico, es la etapa hidrolítica. La etapa limitante en el rango termofílico es la fase metanogénica, inhibida por amonio. En el rango termofílico, se obtiene una importante acumulación de ácido butírico a partir de una concentración de ácido acético de 6 g Ac/L. La hidrólisis de los compuestos proteicos depende de la concentración de 292 Ensayos de viabilidad de codigestión en continuo amonio del reactor y de la temperatura de trabajo, siendo mayor a temperaturas termofílicas que a mesofílicas, de forma que a igualdad de substrato la concentración de nitrógeno amoniacal en un reactor anaerobio es mayor en el rango termofílico que en el mesofílico. La concentración de acético puede considerarse como un importante parámetro de diagnóstico de la inhibición por amonio. Aunque se ha constatado que el grado de inhibición por amonio es diferente a temperaturas termofílicas y mesofílicas, no se puede asegurar que la causa de la mayor inhibición sea la mayor concentración de amoníaco libre, ya que la correlación entre la concentración de acético y la producción de gas con la concentración de amoníaco libre no es significativa si la concentración de amonio total es superior a 3000 mg N-NH4+/L, mientras que sí es significativa la correlación con la concentración de amonio total. Si la concentración es inferior a 3000 mg N-NH4+/L, la forma tóxica parece ser el amoníaco libre, pero por encima de este valor podría iniciarse la inhibición por la forma iónica. 6.7.4. Sobre el efecto de la codigestión con tierras decolorantes de aceite de oliva La utilización de tierras decolorantes de aceite de oliva (TDO) como cosubstrato, en el rango mesofílico, tiene un efecto claramente positivo sobre el proceso de digestión anaerobia, aumentando la producción de metano, que pasa de un valor medio de 0,6 L/L*d a 1,34 L/L*d. El incremento en la producción de metano se debe, en parte, al aumento de carga orgánica del reactor, pero la producción específica de metano también aumenta. Las tierras decolorantes de aceite de oliva introducen un importante efecto inhibidor, relacionado con el alto contenido de ácidos grasos de cadena larga. Este efecto es sobre todos los procesos, pero especialmente sobre la población metanogénica, acetoclástica y más aún la hidrogenotrófica, que a su vez inhibe la acetogénesis. Un efecto importante observado es la inhibición del proceso de degradación del ácido valérico. La inhibición de la acetogénesis es más duradera que la metanogénesis acetoclástica, como indica el mantenimiento de una alta concentración de propiónico al final del experimento, y la subida de la relación Propiónico/acético. La inhibición por AGCL es mayor a temperaturas termofílicas que a mesofílicas. No se ha observado en el rango termofílico ningún efecto de mejora sobre la inhibición por amonio debido a la mezcla con cosubstratos. Debido al efecto inhibidor de las TDO resulta fundamental la introducción Optimización de la digestión anaerobia de purines de cerdo 293 gradual de este cosubstrato, que permita la constitución de una población acetogénica suficiente para evitar la acumulación de AGCL en el reactor. En el presente experimento se realizó en dos pasos, cada uno con una duración de unos 20 días, sin que se haya evitado la acumulación de AGV. Por ello se propone utilizar un mayor número de pasos, aunque sean de menor duración. 6.7.5. Resumen de conclusiones En el rango termofílico el proceso ha estado fuertemente inhibido por la alta concentración de amonio en el reactor, siendo la población metanogénica la más afectada. Existe una clara correlación entre la concentración de nitrógeno amoniacal y el aumento de la concentración de acético y la disminución de la producción de gas. La codigestión de purín con tierras decolorantes de aceite de oliva mostró un buen comportamiento en el rango mesofílico, aumentando tanto la producción volumétrica de gas como la producción por unidad de carga. Sin embargo, la adición de TDO en el rango termofílico inhibió la producción de metano. A pesar de los buenos resultados, el cosubstrato TDO introduce un importante efecto inhibidor, relacionado con el alto contenido de ácidos grasos de cadena larga. Este efecto es sobre todos los procesos, pero especialmente sobre la población metanogénica, acetoclástica y más aún la hidrogenotrófica, que a su vez inhibe la acetogénesis. Un efecto importante observado es la inhibición del proceso de degradación del ácido valérico. La inhibición de la acetogénesis es más duradera que la metanogénesis acetoclástica. 294 Ensayos de viabilidad de codigestión en continuo 7. CONCLUSIONES GENERALES 296 Conclusiones generales Optimización de la digestión anaerobia de purines de cerdo 297 7.1. Del modelo de simulación dinámica del pH El algoritmo de simulación dinámica del pH desarrollado es capaz de simular con muy altos coeficientes de regresión el comportamiento del pH en soluciones acuosas sencillas, por lo que es un herramienta válida, útil para ser introducida en cualquier modelo de simulación de procesos dinámicos en los que el pH juegue un papel importante. El algoritmo de simulación del pH ha sido introducido en un modelo dinámico estructurado de digestión anaerobia de substratos complejos. Se ha utilizado notación matricial lo cual posibilita la introducción de nuevas variables al modelo, en caso de que se considere oportuno. La introducción de la simulación dinámica de las variables de la fase gaseosa supone una mejora importante del modelo original. Se ha comprobado la utilidad del modelo como herramienta cualitativa para prever situaciones de estrés y sobrecargas provocadas por la introducción de un cosubstrato con alto contenido en materia orgánica, o por la introducción de algún tóxico. Estos resultados son de interés para guiar los ensayos experimentales e interpretar resultados. 7.2. Experimentos en discontinuo 7.2.1. Sobre el método La utilización de un pequeño volumen de inóculo permite, en un único test, estimar la viabilidad de un determinado substrato para ser digerido anaeróbicamente, ya que proporciona concentraciones de los posibles tóxicos, que podrían ser desconocidos, cercanos a los valores reales en la mezcla problema. La cantidad de inóculo utilizada (10% en peso del substrato), en algunos casos ha sido insuficiente, provocando un efecto similar a una sobrecarga orgánica, que se ha traducido en el retraso inicial de la producción de metano, o aparición de la fase lag. Se han conseguido buenos ajustes a un modelo simple basado en la ecuación de Gompertz, que tiene en cuenta la duración de la fase lag. La combinación de los parámetros velocidad de producción de metano, duración de la fase lag, evolución de producción acumulada de metano y 298 Conclusiones generales evolución de la concentración de los AGV individuales proporciona suficiente información para el estudio del proceso e identificación de problemas y causas. Cuando la presencia de tóxicos en el medio impide la determinación del potencial de producción de metano es necesario la aplicación del test estándar de biodegradabilidad. 7.2.2. Resultados del experimento 1: Diluciones de purín - efecto del amonio. La digestión anaerobia de purines de cerdo con una concentración de nitrógeno amoniacal inicial de 3,43 g N-NH4+/L es posible en el rango mesofílico, sin que se haya observado inhibición por amonio, ni por amoníaco libre. La concentración de amoníaco libre que inhibe el proceso en el rango mesofílico está por encima de los 305 mg de N-NH3/L. La digestión anaerobia de purines de cerdo con una concentración de 3,43 g N-NH4+/L en el rango termofílico es posible aunque con una producción de metano mucho menor que la obtenida para el rango mesofílico. El proceso en el rango termofílico está afectado fundamentalmente por la inhibición por amoníaco libre. La concentración de amoníaco libre a partir de la cual el proceso se inhibe es ligeramente menor de 640 mg de NNH3/L, correspondiente con una concentración de nitrógeno amoniacal inicial de 2,63 g N-NH4+/L, aunque no se pudo establecer claramente el valor límite. El principal proceso inhibido en los tratamientos más concentrados fue la metanogénesis acetoclástica, pero también resultó afectada la hidrólisis de proteínas y la acidogénesis. La hidrólisis de proteínas es mayor en el rango termofílico que en el mesofílico, produciendo un mayor incremento de la concentración de nitrógeno amoniacal. La inhibición de la ruta metanogénica a partir de hidrógeno pareció más afectada en el rango mesofílico que en el termofílico, según se comprueba de la mayor acumulación de ácido propiónico en el rango mesofílico. 7.2.3. Resultados del experimento 2: Codigestión con residuo de pera. La producción acumulada de metano aumentó al añadir como cosubstrato residuo de pera, tanto en el rango termofílico como en el mesofílico. Las producciones máximas se obtuvieron en el rango mesofílico. Los mejores resultados, dentro del rango mesofílico, se obtuvieron para el tratamiento con un 12,5% en peso de residuo de pera, aumentando no sólo Optimización de la digestión anaerobia de purines de cerdo 299 la producción acumulada de metano respecto a la del purín, sino también el índice de producción de metano respecto a sólidos volátiles. La digestión del residuo de pera por sí solo no es viable en las condiciones utilizadas, debido a su baja alcalinidad. La adicción de más de un 12,5% de residuo de pera en peso provoca un desfase en el inicio de la producción de metano, provocando un efecto similar a una sobrecarga orgánica en un sistema en continuo. La duración del desfase aumentó con la proporción de cosubstrato añadido. En el rango termofílico la producción acumulada aumentó conforme aumentó la proporción de cosubstrato, aunque el aprovechamiento de la materia orgánica añadida fue menor, como demuestra el menor índice de producción de metano respecto a sólidos volátiles iniciales. El principal efecto de la adición de residuo de pera fue la bajada del pH y por tanto la reducción de la inhibición por amonio. Al aumentar el pH por consumo de AGV se vuelven a restablecer las condiciones de inhibición, motivo por el cual no se alcanzaron valores tan altos como en mesofílico. En el rango termofílico no se detectó desfase en la producción de gas, indicando que los organismos termofílicos son más resistentes a las sobrecargas orgánicas. 7.2.4. Resultados del experimento 3. Adición de tierras decolorantes de aceite de oliva. La producción de metano acumulada y respecto a sólidos volátiles iniciales en el rango mesofílico aumenta al añadir TDO como cosubstrato. En el caso de un 5% de TDO la producción acumulada fue de 25,78 mL CH4/g substrato, frente a 8,86 mL CH4/g purín, es decir casi tres veces. El tratamiento con un 5% de TDO obtuvo la producción máxima de metano, no sólo de este experimento, sino de todos los experimentos realizados. La velocidad de producción de metano fue también la máxima. La digestión de tierras decolorantes de aceite de oliva sin mezclar con purín no es viable en las condiciones estudiadas, resultando en producciones de metano más bajas que el control, debido a la baja alcalinidad y a las altas concentraciones de ácidos grasos de cadena larga. Los ácidos grasos de cadena larga inhiben el proceso anaerobio tanto en termofílico como en mesofílico. En termofílico se inhibió completamente con una concentración aproximada de 7,8 g/L de trioleato, sin que fuera capaz de recuperarse. En el rango mesofílico el principal efecto observado fue la aparición de un desfase en la producción de metano, que aumentó con la proporción de TDO, y por tanto con la concentración de oleico. 300 Conclusiones generales 7.3. Experimentos en continuo Las principales conclusiones obtenidas de los resultados de los ensayos en continuo se resumen en los siguientes puntos: 7.3.1. Sobre el método Para la obtención de resultados concluyentes es conveniente mantener las condiciones estables durante períodos más largos que los utilizados en el presente estudio. La excesiva variación en las características de la alimentación han podido perjudicar la obtención de resultados, a veces enmascarados por otros efectos. La velocidad de agitación ha de ser suficiente para evitar la acumulación de sólidos en el reactor. Se han constatado diferencias importantes en la eficiencia de la agitación en función de la temperatura, habiéndose observado una mayor acumulación de sólidos en los reactores termofílicos que en los mesofílicos. 7.3.2. Sobre el método de puesta en marcha La puesta en marcha de un reactor anaerobio requiere largos tiempos, si se pretenden conseguir condiciones de adaptación adecuadas. La utilización como inóculo de lodo de depuradora de aguas residuales urbanas, incrementando gradualmente la proporción de purín en la alimentación, es una opción viable para la puesta en marcha para tratar purines de cerdo. Es posible utilizar un inóculo de un reactor mesofílico para la puesta en marcha de un reactor termofílico, recuperándose en pocos días de la perturbación, adaptándose bien a las nuevas condiciones, sin que se observe reducción de la producción de gas. El factor más limitante sobre el proceso de digestión anaerobia de purines de cerdo en el rango termofílico, en la fase de puesta en marcha, es la concentración de amonio. 7.3.3. Sobre la viabilidad de la digestión anaerobia de purines de cerdo La producción de metano fue mayor en el rango mesofílico que en el termofílico, tanto en la producción volumétrica como la producción por unidad de carga. En el rango termofílico la producción fue muy baja, muy por debajo del umbral de rentabilidad propuesto en la bibliografía. La producción volumétrica y por unidad de carga de metano aumentan, en Optimización de la digestión anaerobia de purines de cerdo 301 el rango mesofílico, al aumentar la velocidad de carga orgánica entre 2,5 y 4 g SV/L*d. En general, en ausencia de inhibidores, la etapa limitante de la digestión anaerobia de purines de cerdo en el rango mesofílico, es la etapa hidrolítica. La etapa limitante en el rango termofílico es la fase metanogénica, inhibida por amonio. En el rango termofílico, se obtiene una importante acumulación de ácido butírico a partir de una concentración de ácido acético de 6 g Ac/L. La hidrólisis de los compuestos proteicos depende de la concentración de amonio del reactor y de la temperatura de trabajo, siendo mayor a temperaturas termofílicas que a mesofílicas, por lo que a igualdad de substrato la concentración de nitrógeno amoniacal en un reactor anaerobio es mayor en el rango termofílico que en el mesofílico. La concentración de acético puede considerarse como un importante parámetro de diagnóstico de la inhibición por amonio. Aunque se ha constatado que el grado de inhibición por amonio es mayor en el rango termofílico que en el mesofílico, no se puede asegurar que la causa sea la mayor concentración de amoníaco libre a temperaturas termofílicas, ya que la correlación entre la concentración de acético y la producción de gas con la concentración de amoníaco libre no es significativa si la concentración de amonio total es superior a 3000 mg N-NH4+/L, mientras que sí es significativa la correlación con la concentración de amonio total. Si la concentración de amonio es inferior a 3000 mg N-NH4+/L, la forma tóxica parece ser el amoníaco libre, pero por encima de este valor podría iniciarse la inhición por la forma iónica. 7.3.4. Sobre el efecto de la codigestión con tierras decolorantes de aceite de oliva La utilización de tierras decolorantes de aceite de oliva (TDO) como cosubstrato, en el rango mesofílico, tiene un efecto claramente positivo sobre el proceso de digestión anaerobia, aumentando la producción de metano, que pasa de 0,6 L/L*d a 1,34 L/L*d. El incremento en la producción de metano se debe, en parte, al aumento de carga orgánica del reactor, pero el índice de producción de metano sobre sólidos volátiles se incrementa también. Las tierras decolorantes de aceite de oliva introducen un importnate efecto inhibidor, relacionado con el alto contenido de ácidos grasos de cadena larga. Este efecto es sobre todos los procesos, pero especialmente sobre la población metanogénica, acetoclástica y más aún la hidrogenotrófica, que a 302 Conclusiones generales su vez inhibe la acetogénesis. Un efecto importante observado es la inhibición del proceso de degradación del ácido valérico. La inhibición de la acetogénesis es más duradera que la metanogénesis acetoclástica, como indica el mantenimiento de una alta concentración de propiónico al final del experimento, y la subida de la relación propiónico/acético. La inhibición por AGCL es mayor a temperaturas termofílicas que a mesofílicas. No se ha observado en el rango termofílico ningún efecto de mejora sobre la inhibición por amonio debido a la mezcla con cosubstratos. Debido al efecto inhibidor de las TDO resulta fundamental la introducción gradual de este cosubstrato, que permita la constitución de una población acetogénica suficiente para evitar la acumulación de AGCL en el reactor. En el presente experimento se realizó en dos pasos, cada uno con una duración de unos 20 días, sin que se haya evitado la acumulación de AGV. Por ello se propone utilizar un mayor número de pasos, aunque sean de menor duración. 7.4. Conclusiones generales. x x x x x x El algoritmo de simulación dinámica del pH desarrollado es capaz de predecir con muy altos coeficientes de regresión el comportamiento del pH en soluciones acuosas sencillas. A pesar de la utilización de parámetros cinéticos obtenidos de la bibliografía, se ha comprobado la utilidad del modelo de simulación del proceso anaerobio con substratos complejos como herramienta cualitativa para prever situaciones de estrés y sobrecargas, provocadas por la introducción de un cosubstrato con alto contenido en materia orgánica, o la introducción de algún tóxico. La digestión de purín es posible tanto en rango termofílico como en mesofílico, aunque en el primer rango de temperatura está claramente inhibido por amonio, obteniéndose menores producciones de gas que en rango mesofílico, y observándose un aumento en la producción de gas con el grado de dilución. La digestión anaerobia del residuo de pera y de las tierras decolorantes de aceite de oliva no es posible sin mezclar con otro substrato. La codigestión de purín con otros substratos es una buena opción para aumentar la producción de metano. La producción de metano, en términos absolutos, aumenta al añadir Optimización de la digestión anaerobia de purines de cerdo x x x x x 303 como cosubstrato residuo de pera. Sin embargo, el principal efecto de la adición de este residuo fue el mayor aporte de materia orgánica, por lo que el índice de producción de metano respecto a sólidos volátiles mostró, solo, una pequeña mejora. Se observó un efecto de sobrecarga orgánica, mayor en el rango mesofílico que en el termofílico. La codigestión de purín con tierras decolorantes de aceite de oliva mostró un buen comportamiento en el rango mesofílico, aumentando tanto la producción volumétrica de gas como la producción por unidad de carga, tanto en los ensayos en discontinuo como en los ensayos en continuos. Sin embargo, la adición de TDO en el rango termofílico inhibió completamente la producción de metano. Las tierras decolorantes de aceite de oliva introducen un importnate efecto inhibidor, relacionado con el alto contenido de ácidos grasos de cadena larga. Este efecto es sobre todos los procesos, pero especialmente sobre la población metanogénica, acetoclástica y más aún la hidrogenotrófica, que a su vez inhibe la acetogénesis. Un efecto importante observado es la inhibición del proceso de degradación del ácido valérico. La inhibición de la acetogénesis es más duradera que la metanogénesis acetoclástica. La adaptación del inóculo es muy importante para asegurar la viabilidad del proceso. La utilización de ensayos en discontinuo con baja concentración de inóculo proporciona información de respuesta a sobrecargas, y mantiene las condiciones de toxicidad reales. Este tipo de ensayo proporciona, al menos, una aproximación cualitativa a la viabilidad del proceso anaerobio con mezclas de residuos, aún en el caso de desconocer su composición exacta, o la existencia de algún inhibidor del proceso metanogénico. El empleo de este tipo de test no es incompatible, sino complementario, con la realización de tests de biodegradabilidad y de toxicidad. Así mismo es necesario realizar ensayos en continuo de laboratorio antes de pasar a escala industrial, para confirmar los resultados y determinar otro tipo de parámetros, como la velocidad de carga orgánica, tiempo de retención, velocidad de introducción de cosubstratos, etc. 7.5. Consideraciones de futuro. Se han cumplido la mayoría de los objetivos propuestos en el trabajo. No 304 Conclusiones generales obstante, también se han detectado algunos defectos en la metodología que deberán ser subsanados en trabajos posteriores. El método de experimentos en discontinuo es un método válido para la obtención de resultados cualitativos, siendo mucho más económico que los estudios en continuo. En cualquier caso, no evitan la realización posterior de ensayos en continuo, si los resultados son positivos, aunque sí lo evita si los resultados son negativos. La modelización del proceso anaerobio de substratos complejos, ha resultado una herramienta útil para prever el comportamiento del sistema ante la introducción de un nuevo substrato. Sin embargo, el modelo no está completamente calibrado para todas las situaciones, habiéndose detectado algunos defectos, como, por ejemplo, la no dependencia de la temperatura, de la inhibición por AGCL. Igualmente, el modelo no considera de forma independiente el proceso metanogénico hidrogenotrófico, lo que impide la simulación de sobrecargas orgánicas, ya que es éste el fenómeno que suele afectarse más en este tipo de situaciones. Es por tanto una línea de investigación abierta, en la que sería muy interesante continuar trabajando, tanto en la modelización propiamente dicha, introduciendo los procesos necesarios, como en la calibración del modelo, a partir de los datos experimentales obtenidos en los capítulos 5 y 6, y en nuevos experimentos que se lleven a cabo. 8. REFERENCIAS BIBLIOGRÁFICAS 306 Referencias bibliográficas Optimización de la digestión anaerobia de purines de cerdo 307 Ahring, B.K. (1995). Methanogenesis in thermophilic biogas reactors. Antonie van Leeuwnhoek. Vol. 67, pag. 91-102. Ahring, B.K., Angelidaki, I., Johansen, K. (1992). Anaerobic treatment of manure together with industrial waste. Water Science Technology. Vol. 25 (7), pag. 311-318. Ahring, B.K., Sandberg, M., Angelidaki, I. (1995). Volatile fatty acids as indicators of process imbalance in anaerobic digestors. Applied microbiological Biotechnology. Vol. 43 (3), pag.559-565. Ahring, B.K., Westermann, P. (1987a).Thermophilic anaerobic degradation of butyrate by a butyrate-utilizing bacterium in coculture and triculture with methanogenic bacteria. Applied and Environmental Microbiology. Vol. 53 (2), pag. 429-433. Ahring, B.K., Westermann, P. (1987b). Kinetics of butyrate, acetate, and hydrogen metabolism in a thermophilic, anaerobic, butyrate-degrading Triculture. Applied and Environmental Microbiology. Vol. 53 (2), pag. 434-439. Ahring, B.K., Westermann, P. (1988). Product Inhibition of butyrate metabolism by acetate and hydrogen in a thermophilic coculture. Applied and environmental microbiology. Vol. 54 (10), pag. 2393-2397. American Public Health Association (1995). Standard methods for examination of water and wastewater. 19th ed., APHA-AWWA-WEF, Wshington DC, USA. Andrews, J. F., Graef, S.P. (1971). Dynamic Modeling and simulation of the anaerobic digestion proccess. En Anaerobic biological treatment processes. Advances in chemistry series, 105. American Chemical Society. Washington D.C. Angelidaki, I., Ahring, B.K. (1992). Effects of free long fatty acids on thermophilic anaerobic digestion. Applied Microbiology and Biotechnology. Vol. 37 (6), pag., 808-812. Angelidaki, I., Ahring, B.K. (1993a). Effect of the clay mineral bentonite on ammonia inhibition of anaerobic thermophilic reactors degrading animal waste. Biodegradation. Vol. 3, pag. 409-414. Angelidaki, I., Ahring, B.K. (1993b). Thermophilic anaerobic digestion of livestock waste: the effect of ammonia. Applied Microbiology and Biotechnology. Vol. 38, pag. 560-564. Angelidaki, I., Ahring, B.K. (1994). Anaerobic thermophilic digestion of manure at different ammonia loads: effect of temperature. Water Research. Vol. 28 (3), pag. 727-731. Angelidaki, I., Ahring, B.K. (1997a). Anaerobic digestion in Denmark. Past, 308 Referencias bibliográficas present and future. III curso de Ingeniería Ambiental, pag., 336-342. Lleida, octubre de 1997. Angelidaki, I., Ahring, B.K. (1997b). Monitoring and controlling the biogas process. III curso de Ingeniería Ambiental, pag., 270-282. Lleida, octubre de 1997. Angelidaki, I., Ellegaard, L., Ahring, B.K. (1993). A mathematical model for dynamic simulation of anaerobic digestion of complex substrates: focusing on ammonia inhibition. Biotechnology and Bioengineering. Vol, 42, pag 159-166. Angelidaki, I., Ellegaard, L., Ahring, B.K. (1996). A mathematical model for dynamic simulation of anaerobic digestion of complex substrates: focusing on codigestion with lipid containing substrates. Management of Urban Biodegradable Wastes pag. 132-142. Angelidaki, I., Ellegaard, L., Ahring, B.K.(1997) Modelling anaerobic codigestion of manure with olive oil mill effluent. Water Science and Technology. Vol.36 (6-7), pag. 263-269. Angelidaki, I., Ellegaard, L., Ahring, B.K. (1999). A comprehensive model of anaerobic bioconversion of complex substrates to biogas. Biotechnology and Bioengineering. Vol, 63 (3), pag 363-372. Angelidaki, I., Petersen, SP, Ahring, B. (1990). Effects of lipids on thermophilic anaerobic digestion and reduction of lipid inhibition upon addition of bentonite. Applied Microbiology and Biotechnology. Vol. 33 (4), pag. 469-472. Bagley, D.M.; Brodkorb, T.S. (1999). Modeling microbial kinetics in an anaerobic sequencing batch reactor- model development and experimental validation. Water Environment Research. Vol. 71 (7), pag. 1320-1332. Baier, U., Schmidheiny, P. (1997). Enhanced anaerobic degradation of mechanically disintegrated sludge. Water Science and Technology. Vol.36 (11), pag. 137-143. Banks, C.J., Humphreys, P.N.(1998). The Anaerobic Treatment of a Lignocellulosic substrate offering little natural pH buffering capacity. Water Science and Technology. Vol. 38 (4-5), pag. 29-35. Bardiya, N., Somayaji, D., Khanna, S. (1996). Biomethanation of banana peel and pineapple waste. Bioresource Technology. Vol. 58, pag. 73-76. Barredo, M.S., Evison, L.M. (1991). Effect of propionate toxicity on methanogen-enriched sludge, Methanobrevibacter smithii, and Methanospirillum hungatii at different pH values. Applied and environmental microbiology. Vol. 57(6), pag. 1764-1769. Optimización de la digestión anaerobia de purines de cerdo 309 Bonmatí, A., Flotats, X., Mateu, L., Campos, E. (2000). Estudio de la hidrólisis térmica como pre-tratamiento de la digestión anaerobia mesofílica de purines de origen porcino. VI Seminario Latinoamericano de digestión anaerobia. Recife, Brasil, noviembre de 2000. Bonmatí, A. (1998). Digestió anaeròbia de purins amb altres residus orgànics. Pagès Editors. Lleida. Boone, D., Bryant, M. (1980). Propionate-degrading bacterium, Syntrophobacter wolinii sp. nov.gen.nov., from methanogenic ecosystems. Applied and environmental microbiology. Vol. 40(3), pag. 626-632. Boone, D., Mah, R. (1987). Transitional Bacteria. En Anaerbic digestion of biomas. Editado por Chynoweth, D.P y Isaacson, R. Elsevier applied science LTD. Boone, D., Xun, L. (1987). Effects of pH, temperature and nutrients on propionate degradation by methanogenic enrichment culture. Applied and environmental microbiology. Vol. 53(3), pag.1589-1592. Brinkman, J. (1999). Anaerobic digestion of mixed waste slurries from kitchens, slaughterhouses and meat processing industries. Proceedings of the II International symposium on Anaerobic Digestion of Soild Waste. Barcelona, Junio de 1999, pag 190-191. Bryant, M.P., Tzeng, S.F., Robinson, I.M., Joyner, A.E. (1971). Nutrient requeriments of methanogenic bacteria. Anaerobic biological treatment processes. Advances in chemistry series, 105. American Chemical Society. Washington DC. Cairó, J.J., París, J.M. (1988). Microbiología de la digestión anaerobia, metanogénesis. Actas del 4th seminario de tratamiento anaerobio de aguas residuales. Valladolid, pag. 41-51. Callaghan, F.J., Wase, D.A.J., Thayanithy, K., Forster, C.F.(1999). Codigestion of waste organic solids: batch studies. Bioresource technology. Vol. 37, pag. 117-122. Campos, E., Palatsi, J., Flotats, X.(1999). Codigestion of pig slurry and organic wastes from food industry. Proceedings of the II International symposium on Anaerobic Digestion of Soild Waste. Barcelona, Junio de 1999, Vol.II, pag. 192-195. Chen, Y.R., Hashimoto, A.G. (1978).Kinetics of methane fermentation. Biotechnology and Bioengineering Symp. Vol. 8, pag. 269-282. Chen, T.H., Hashimoto, A.G. (1996). Effects of pH and substrate: inoculum ratio on batch methane fermentation. Bioresource Technology. Vol. 56, pag. 179-186. Chiu, Y.C.,Chang, C.N., Lin, J.G., Huang, S.J. (1997). Alkaline and ultrasonic 310 Referencias bibliográficas pretreatment of sludge before anaerobic digestion. Water Science and Technology. Vol.36 (11), pag. 152-162. Clark, R.H., Speece, R.E. (1989). The pH tolerance of anaerobic digestion. Advanced water pollution research. Int. Conf. 5th, pag. 27/1-27/14. Cobb, S.A., Hill, D.T. (1991). Volatile fatty acid relationships in attached growth anaerobic fermenters. Transactions of ASAE. Vol. 34 (6), pag. 231-234. Contois, D.E. (1959). Kinetics of bacterial growth: relationship between population density and specific growth rate of continuous cultures. J. gen. Microbiol. Vol. 21, pag. 40-50. Coombs, J. (1990). “The present and future of anaerobic digestion”, en Anaerobic digestion: a waste treatment technology. Editado por Wheatley, A. Critical reports on applied chemistry. Vol. 31, pag. 93-138. Elsevier applied science LTD. Costello, D.J., Greenfield, P.F., Lee, P.L. (1991). Dynamic modelling of a single-stage high-rate anaerobic reactor- 1. Model derivation. Water Research. Vol. 25 (7), pag. 847-858. Danés, R., Molina, V., Prats, I.L., Álamos, M., Boixadera, J., Torres, E. (1996). Manual de gestió dels purins i de la seva reutilització agrícola. Editado por la Generalitat de Catalunya. Barcelona Danish Energy Agency (1999). Bio-statistik (Nyt om biogasfaellesanlaeg). Dansk Bioenergi. Vol, 48, pag.10-17. Dar, G.H., Tandon, S.M. (1987). Biogas production from pretreated wheat straw, lantan residue, apple and peach leaf litter with cattle dung. Biological wastes. Vol. 21, pag. 75-83. DARP (Departament d’agricultura, ramaderia i Pesca) (1999). Estadístiques i cojuntura agrària, 1999 pag.137-138. DARP (Departament d’agricultura, ramaderia i Pesca) (1999). Estadístiques ramaderes. Estadístiques i cojuntura agrària,1999, pag.78-91. Delgenès, J.P., Penaud, V., Torrijos, M., Moletta, R. (1999). Thermochemical pretreatment of an indsutrial microbial biomass: effect of sodium hydroxide addition on COD solubilization, anaerobic biodegradability and generation of soluble inhibitory compounds. Proceedings of II International Symposium on anaerobic digestion of solid waste. Barcelona, 15-17 de junio , 1999. pg, 121-128. Desai, M., Madamwar, D. (1994). Anaerobic digestion of a mixture of cheese whey, poultry waste and cattle dung: a study of the use of adsorbents to improve digester performance. Environmental pollution. Vol. 86, pag. 337-340 Optimización de la digestión anaerobia de purines de cerdo 311 Desai, M., Patel, V., Madamwar, D. (1994). Effect of temperature and retention time on biomethanation of cheese whey-poultry waste-cattle dung. Environmental pollution. Vol. 83, pag. 311-315 Di Palma, L., Medici, F., Merli, C., Petruci, E. (1999). Optimising gas production in the anaerobic codigestion of the organic fraction of solid waste from markets. Proceedings of the II International symposium on Anaerobic Digestion of Solid Waste. Barcelona, Junio de 1999, pag 184189. Dinsdale, R.M., Premier, G.C., Hawkes, F.R., Hawkes, D.L.(2000). Twostage anaerobic codigestion of waste activated sludge and fruit/vegetable waste using inclined tubular digesters. Bioresource technology. Vol. 72, pag. 159-168. Edelmann, W., Engeli, H., Graddenecker, M. (1999). Codigestion of organic solid wastes and wastewater. Proceedings of the II International symposium on Anaerobic Digestion of Solid Waste. Barcelona, Junio de 1999, pag. 381-388. Fannin, K.F. (1987). “Start-up, operation, stability, and control”, en Anaerobic digestion of biomass. Editado por Chynoweth, D. Y y Isaacson, R. Elsevier applied science LTD. Ferguson, T., Mah, R. (1987). “Methanogenic bacteria”, en Anaerobic digestion of biomass. Editado por Chynoweth, D. Y y Isaacson, R. Elsevier applied science LTD. Field, J., Sierra Alvárez, R-, Lettinga, G. (1988). Ensayos anaerobios. Actas del 4º Seminario de Depuración anaerobia de aguas residuales. Valladadolid, pag. 52-80. Fisher, J.R. (1979). Producing methane gas from swine manure in a pilot-size digester. Transactions of the ASAE. Vol. 22 (2), pag. 370-374. Flotats, X., Bonmatí, A., Campos, E., Antúnez, M. (1999). Ensayos en discontinuo de codigestión anaerobia termofílica de purines de cerdo y lodos residuales. Información Tecnológica. Vol. 10 (1), pag. 79-85. Flotats, X., Bonmatí, A., Campos, E., Teira, M.R. (2000). El proceso de secado de purines en el marco de gestión integral de residuos ganaderos. Residuos. Vol. 53, pag. 40-46. Flotats, X., Campos, E., Bonmatí, A. (1998). Banc d'assaigs de digestió anàerobia de residus orgànics. II Jornades sobre Energia, pag. 21-27. Flotats, X., Campos, E., Bonmatí, A. (1998). Tecnologías para la modificación de las características de los residuos: caracterización general de métodos. IV Curso de Ingeniería Ambiental. Lleida, ocubre de 1998. Frías, L., García-Ortiz, A., Hermoso, M., Jimenez, A., Llavero, M.P., 312 Referencias bibliográficas Morales. J., Ruano, T., Uceda, M. (1991). “Análisis de aceites (III)”, en Analistas de laboratorio de almazara. Junta de Anadalucía-Consejería de Agricultura y Pesca. Dirección general de investigación y extensión agrarias. Centro de información y documentación agraria. Sevilla. Fukuzaki, S., Nishio, N., Shobayashi, M., Nagai, S. (1990). Inhibition of fermentation of propionate to methane by hydrogen, acetate, and propionate. Applied and Environmental Microbiology, 56 (3), 719-723 Galbraith, H., Miller, T.B., Paton, A.M., Thompson, J.K. (1971). Antibacterial activity of long chain fatty acids and the reversal with Calcium, Magnesium, Ergocalciferol y Colesterol. Journal of applied Bacteriology. Vol. 34 (4), pag. 803-813. Gallert, C., Bauer, S., Winter, J. (1998). Effect of ammonia on the anaerobic degradation of protein by a msophilic and thermophilic biowaste population. Applied Microbiology and Biotechnology. Vol. 50, pag. 495501. García Gispert, S. (1998). Automatització del registre de la producció de gas d’un procés de fermentació anaeròbia de residus. Trabajo Final de Carrera. Enginyeria Tècnica en Informàtica de Sistemes. Escola Universitària Politècnica. Universitat de Lleida. Gavala, H.N., Skiadas, I.V., Bozinis, N.A., Lyberatos, G. (1996). Anaerobic codigestion of agricultural industries’ wastewaters. Water Science and Technology. Vol. 34 (11), pag. 67-75. Generalitat de Catalunya (Institut d’Estadística de Catalunya y Departament de Medi Ambient) (1999). Dades del medi ambient de Catalunya. URL: http://www.gencat.es/mediamb/gt/1999. Grau, P., Dohànyos, M., Chudoba, J. (1975). Kinetics of multicomponent substrate removal by activated sludge. Water Research. Vol. 9, pag. 637. Griffin, M. E.; McMahon, K. D.; Mackie, R. I.; Raskin, L.(1998). Methanogenic Population Dynamics during Start-Up of Anaerobic Digesters Treating Municipal Solid Waste and Biosolids. Biotechnology and bioengineering. Vol. 57, pag. 342-355. Hamzawi, N., Kennedy, K.J., McLean, D.D. (1998). Anaerobic digestion of co-mingled municipal solid waste and sewage sludge. Water Science and Technology. Vol 38 (2), pag. 127-132 Han, Y., Sung, S., Dague, R. (1997). Temperature-phased anaerobic digestion of wastewaters sludges. Water Science and technology. Vol. 36 (6-7), pag. 367-374. Hanaki, K., Matsuo, T., Nagase, M. (1981). Mechanism of inhibition caused by long-chain fatty acids in anaerobic digestion process. Biotechnology Optimización de la digestión anaerobia de purines de cerdo 313 and bioengineering. Vol 23, pag. 1591-1610. Hansen, K., Angelidaki, I., Ahring, B.K. (1998). Anaerobic digestion of swine manure: inhibition by ammonia. Water Research. Vol 32 (1), pag. 5-12. Hansen, K.H., Angelidaki, I., Ahring, B.K. (1999). Improving thermophilic anaerobic digestion of swine manure. Water Research. Vol. 33 (8), pag. 1805-1810. Hansson, G. Y., Molin, N. (1981). End product inhibition in methane fermentations: Effects of Carbon Dioxide on Fermentative Acetogenic Bacteria. European Journal Applied Microbiology and Biotechnology. Vol. 13, pag. 242-247. Harper, S.R., Pohland, F.G. (1986) Recent Develpoments in Hydrogen management during anaerobic biological wastewater treatment. Biotechnology and Bioengineering. Vol. 28, pag. 585-602. Hartmann, H., Angelidaki, I., Ahring, B.K.(1999) Increase of anaerobic degradtaion of particulate organic matter in full-scale biogas plants by mechanical maceration. Proceedings of II International Symposium on anaerobic digestion of solid waste. Barcelona, 15-17 de junio , 1999. pg, 129-136. Hashimoto, A.G. (1986). Ammonia Inhibition of methanogenesis from cattle wastes. Agricultural Wastes. Vol. 17, pag. 241-261. Hashimoto, A.G. (1989). Effect of inoculum/substrate ratio on methane yield and production rate from straw. Biological wastes. Vol. 28, pag. 247255. Hayes, T.D., Theis, T.L. (1978). The distribution of heavy metals in anaerobic digestion. Journal water pollution control federation. Vol. 50 (1), pag. 31-72. Henze, M., Gujer, W., Mino, T., Matsuo, T, Wentzel, M., Marais, G. (1995) Activated sludge model nº 2. IAWQ Scientific and Technical Report nº 3. Int. Assoc. Water Qual. Londres. Henze, M. (1995). Basic biological processes. En Wastewater treatment. Biological and chemical processes. Henze, M., Harremöes, P., Cour Jansen, J., Arvin, E Springer-Verlag. Henze, M., Gujer, W., Mino, T., Matsuo, T, Wentzel, M., Marais, G., van Loosdrecht, M. (1999) Activated sludge model nº 2. Water Science and Technology. Vol. 39 (1), pag, 165-182. Hill D.T., Holmberg, R.D. (1988). Long chain volatile fatty acid relationship in anaaerobic digestion of swine waste. Biological wastes. Vol. 23, pag. 195-214 314 Referencias bibliográficas Hill, D.T. (1982) A comprehensive dynamic model for animal waste methanogenesis. Transactions of the ASAE, 25. Pag 1374-1380. Hill, D.T., Cobb, S.A. (1993). Modelling predictive indicators in animal waste methanogenesis. Transactions of the ASAE, 36 (3). Pag 879-885. Hill, D.T., Cobb, S.A., Bolte, J.P. (1987). Using Volatile fatty acid relationships to predict anaerobic digester failure. Transactions of the ASAE. Vol. 30 (2), pag. 496-501. Hill, D.T., Jenkins, S.R.(1989). Measuring alkalinity accurately in aqueous systems containing high organic acid concentrations. Transactions of ASAE, vol. 32 (6), pag. 2175-2178. Hills, D.J., Nakano, K. (1984). Effects of particle size on anaerobic digestion of tomato solid wastes. Agricultural Wastes. Vol. 10, pag. 285-295. Hilpert, R., Winter, J., Kandler, O. (1987). Feed additives and desinfectants as inhibitory factors in anaerobic digestion of agricultural wastes. Biomass for energy. Elsevier applied science LTD. Hobson, P.N. (1990).“ The treatment of agricultural wastes”, En Anaerobic digestion: a waste treatment technology. Editado por Wheatley, A. Critical reports on applied chemistry. Vol. 31, pag. 93-138. Elsevier applied science LTD. Hulshoff Pol, L., Lens, P., Stams, A., Lettinga, G. (1998). Anaerobic treatment of sulphate-rich wastewaters. Biodegradation. Vol. 9, pag. 213224. Hwu, C.-S., Donlon, B., Lettinga, G.(1997) Acute toxicity of oleate to acetate-utilizing methanogens in mesophilic and thermophilic anaerobic sludges. Enzyme and Microbial Technology. Vol. 21, pag. 297-301. Hyung, S.H., Young, J.C., Kim, I.S. (1998). Inhibition kinets for propionate degradation using propionated enriched mixed cultures. Water Science Technology. Vol. 38 (8-9), pag. 443-452. Iza, J. (1995). Control del proceso anaerobio. I Curs d’enginyeria ambiental. Universitat de Lleida. Lleida, abril de 1995. Jain, S., Lala, A., Bhatia, S., Kudchadker, A.P. (1992). Modelling of hydorlysis controlled anaerobic digestion. Journal of Chemic and technology and Biotechnology, 53, 337-344. Kalyuzhnyi, S.V. (1997). Batch anaerobic digestion of glucose and its mathematical modeling. II description, verification and application of model. Bioresource technology. Vol 59, pag. 249-258. Kiely, G., Tayfur, G., Dolan, C., Tanji, K. (1997). Physical and mathematical modelling of anaerobic digestion of organic wastes. Water Research. Vol. 31 (3), pag. 534-540. Optimización de la digestión anaerobia de purines de cerdo 315 Kim, I.S., Kim, D.H., Hyun, S. (1999). Effect of particle size, sodium concnetrations on anaerobic thermophilic food waste digestion. Proceedings of the II International symposium on Anaerobic Digestion of Solid Waste. Barcelona, Junio de 1999, vol. II, pag. 13-16. Kleinstreuer, C., Poweigha, T. (1982). Dynamic simulator for anaerobic digestion processes. Biotechnology and Bioengineering. Vol. 24, Pag. 1941-1951. Koster, I. (1986) Characteristics of the pH influenced adaptation of methanogenic sludge to ammonium toxicity. Journal Chem. Techn. Biotech. Vol. 36, pag. 445-455. Koster, I. (1987). Abatement of long chain fatty acid inhibition of methanogenesis by clacium addition. Biological Wastes. Vol. 22 (4), pag. 295-301. Koster, I., Cramer, A. (1987). Inhibition of methanogenesis from acetate in granular sludge by long chain fatty acids. Applied and Environmental Microbiology. Vol. 53 (2), pag. 403-409. Koster, I.W., Lettinga, G. (1988). Anaerobic digestion at extreme ammonia concentrations. Biological Wastes. Vol. 25, pag. 51-59. Krugel, S., Nemeth, L., Peddie, C. (1998). Extending thermophilic anaerobic digestion for producing class a biosolods at the greater Vancouver regional districts annacis island wastewater treatment plant. Water Science and Technology. Vol. 38 (8-9), pag. 409-416. Krylova, N., Khabiboulline, R., Naumova, R., Nagel, M. (1997). Journal of Chem. Tech. And Biotech. Vol. 79, pag. 99-105. Kugelman, I.J., Chin, K.K. (1971). Toxicity synergism, and antagonism in anaerobic waste treatment processes. Ananerobic biological treatment processes. Advances in chemistry series, 105. American chemical society. Washington D.C. Lay, J.J., Li, Y.Y., Noike, T. (1997). Influences of pH and moisture content on the methane production in high-solids sludge digestion. Water Research, vol. 31 (10), pag., 1518-1524. Lema, J. (1995). Metodología para la obtención de parámetros de diseño de digestores anaerobios. I Curs d’enginyeria ambiental. Universitat de Lleida. Lleida, abril de 1995. Lide, D. (1993). CRC Handbook of chemistry and physics. A ready-reference book of chemical and physical data. Ed. CRC Press, 73 ed. Boca Raton. Madigan, , M.T., Martinko, J.M., Parker, J. (1998). Brock Biología de los microorganismos. Edit. Prentice Hall International (UK) Ltd. 8ª ed. Marchiam, U., Krause, C.(1993). Propionic to acetic acid ratios in overloaded 316 Referencias bibliográficas anaerobic digestion. Bioresource technology. Vol. 43, pag. 195-203. Masciandaro, G., Ceccanti, B., García, C. (1994). Anaerobic digestion of straw and piggery wastewaters:II. Optimization of the process. Agrochimica, Vol. 38 (3), pag. 195- 203. Massé, D.I., Masse, D.Lu, Droste, R.L. (2000). Effect of antibiotics on psychrophilic anaerobic digestion of swine manure slurry in sequencing batch reactors. Bioresource Technology. Vol. 75, pag. 205-211. Mata-Álvarez, J., Martínez-Viturtia, A., Torres, R. (1986). A simple device to measure biogas production in laboratory scale digesters. Biotechnology Letters. Vol. 8 (10), pag. 719-720. McCarty, P.L. (1971). Energetics and kinetics of anaerobic treatment. Anaerobic biological treatment processes. Advances in chemistry series 105. American Chemical Society. Washinton D.C., 1971. Merkel, W., Krauth, K. (1999). Mass transfer of carbon dioxide in anaerobic reactors uner dynamic substrate loading conditions. Water Research. Vol. 33 (9), pag. 2011-2020. Metzler, D.E. (1981). Catabolismo de los azúcares. Bioquímica: Las reacciones químicas en las células vivas. Ed. Omega, S.A. Barcelona. Ministerio de Agricultura, Pesca y Alimentación- MAPA (1994). Métodos oficiales de análisis de alimentos. Mundi-Prensa. Madrid. Monod, J. (1950). La technique de culture continue. Théorie et applications. Annals Institute Pasteur. Vol. 79, pag. 390-410. Moosbrugger, R.E., Wentzel, M.C., Ekama, G.A., Marais, G.R. (1993). Weak acid/bases and pH control in anaerobic systems-a review. Water SA. Vol. 19(1), pag. 1-10. Mosey, F.E. (1983). Mathematical modelling of the anaerobic digestion process: regulatory mechanisms for the formation of short-chain volatile acids from glucose. Water Science and Technology. Vol. 15, pag. 209232. Muñoz Valero, J.A., Ortiz Cañavate, J., Vázquez Minguela, J.(1987).Técnica y aplicaciones agrícolas de la biometanización. Serie Técnica- Ministerio de Agricultura Pesca y Alimentación. Madrid. Noone, G.P. (1990) “The treatment of domestic wastes”, en Anaerobic digestion: a waste treatment technology. Editado por Wheatley, A. Critical reports on applied chemistry. Vol. 31, pag.139-170. Oles, J., Ditchl, N., Niehoff, H. (1997). Full scale experience of two stage thermophilic/mesophilic sludge digestion. Water Science and Technology. Vol. 36 (6-7), pag 449-456. Omil, F., Méndez, R., Lema, J. (1995). Anaerobic treatment of saline Optimización de la digestión anaerobia de purines de cerdo 317 wastewaters under high sulphide and ammonia content. Bioresource Technology. Vol. 54, pag. 269-278. Palmowski, L., Müller, J.(1999). Influence of the size reduction of organic waste on their anaerobic digestion. Proceedings of II International Symposium on anaerobic digestion of solid waste. Barcelona, 15-17 de junio , 1999. pg, 137-144. Pavan, P., Battisoni, P., Cecchi, F., Mata-Álvarez, J. (1999). Two-phase anaerobic digestion of source sorted OFMSW: performance and kinetic study. Proceedings of II International Symposium on anaerobic digestion of solid waste. Barcelona, 15-17 de junio, 1999. pg, 91-98. Pavlostathis, S.G., Giraldo-Gómez, E. (1991). Kinetics of anaerobic treatment: a critical review. Critical reviews in environmental control. Vol. 21 (5,6), pag. 411-490. Péringer, P. (1999). Biomethanation of sorted household waste: experimental validation of a relevant mathematical model. Proceedings of II International Symposium on anaerobic digestion of solid waste. Barcelona, 15-17 de junio , 1999.pg, 10-16. Pomares, F. (1998). Los residuos orgánicos utilizables en la agricultura: origen, composición y características. 4º Curso de Ing. Ambiental. Lleida. Pomares, C., Dil Bailleul, P.J., Rivesto, J. (1999). Alimentar mejor los cerdos para proteger el medio ambiente. Anaporc, nº 193, pag. 87-102. Ratledge, C. (1992). Microbial Oxidations of fatty alcholos and fatty acids, en Mini-review compilation: Biodegradation and Biotransformations of oils and fats. Journal of Chemical Technology and Biotechnology. Vol. 55, 397-414. Rinzema, A., Boone, M., Knippenberg, K.-van, Lettinga, G., Van Knippenberg, K. (1994). Bactericidal effect of long chain fatty acids on anaerobic digestion. Water Environment Research. Vol. 66 (1), pag., 4049. Robbins, J.E., Gerhardt, S.A., Kappel, T.J. (1989). Effects of total amonia on anaerobic digestion and an example of digestor performance from cattle manure-protein mixture. Biological wastes. Vol. 27, pag. 1-4. Salminen , E., Rintala, J., Lokshina, L.Ya., Vavilin, V.A. (1999). Anaerobic batch degradation of solid poultry slaughterhouse waste. Proceedings of the II International symposium on Anaerobic Digestion of Solid Waste. Barcelona, Junio de 1999, pag. 41-48. Sanders, W.T.M., Geerink,M., Zeeman, G., Lettinga, G. (1999). Anaerobic hydrolysis kinetics of particulate substrates. Proceedings of II International Symposium on anaerobic digestion of solid waste. 318 Referencias bibliográficas Barcelona, 15-17 de junio , 1999. pg, 25-32 Schmidt, J.E., Angelidaki, I., Ahring, B.K. (1999). Anaerobic digestion of olive mill effluents together with different wastes. Proceedings of the II International symposium on Anaerobic Digestion of Soild Waste. Barcelona, Junio de 1999, Vol. 2, pag. 180-183. Shimizu, T., Kudo, K., Nasu, Y. (1993). Anaerobic waste-activated sludge digestion. A bioconversion mechanism and kinetic model. Biotechnology and Bioengineering. Vol. 41, pag. 1082-1091. Shin, H., Song, Y. (1995). A model for evaluation of anaerobic degradation characteeristics of organic waste: focusing on kinetics, rate-limiting step. Environmetnal technology. Vol., 16, pp 775-784. Siegrist, H., Renggli, D,.Gujer, W. (1993). Mathematical modelling of anaerobic mesophilic sewage sludge treatment. Water Science and Technology. Vol. 27, 2, pp 25-36. Sleat, R., Mah, R. (1987). “Hydrolytic bacteria”, en Anaerobic digestion of biomass. Editado por Chynoweth, D. Y y Isaacson, R. Elsevier applied science LTD. Speece, R.E. (1987). “Nutrient Requirements”, en Anaerbic Digestion of biomass. Editado por Chynowth D. Y y Isaacson, R. Elsevier applied science LTD. Speece, R.E. (1987). “Toxicity”, en Anaerbic Digestion of biomass. Editado por Chynowth D. Y y Isaacson, R. Elsevier applied science LTD. Stafford, D.A. (1982). The effects of mixing and volatile fatty acid concentrations on anaerobic digester performance. Biomass. Vol. 2, pag. 43-55. Stams, A.J.M. (1994). Metabolic interactions between anaerobic bacteria in methanogenic environments. Antonie van Leeuwenhoek. Vol. 66, pag. 271-294. Teira, M.R., Flotats, X., Casañé, A., Magrí, A., Martín, P., Montané, L., Tarradas, J., Campos, E., Bonmatí, A. (1999). A case study on livestock waste management: Juncosa de les Garrigues, Catalonia, Spain. Jornadas Internacionales de Ingeniería Ambiental. Cartagena, 9-10 Septiembre 1999. Tiehm, A., Nickel, K., Neis, U. (1997). The use of ultrasound to accelerate the anaerobic digestion of sewage sludge. Water Science and Technology. Vol.36 (11), pag. 121-128. Trujillo, D., Pérez, J.F., Cebreros, F.J. (1993). Energy recovery from wastes. Anaerobic digestion of tomato plant mixed with rabbit wastes. Bioresource Technology. Vol. 45, pag., 81-83. Optimización de la digestión anaerobia de purines de cerdo 319 Turner, C., Burton, C.H. (1997). The inactivation of viruses in pig slurries: a review. Bioresource Technology. Vol. 61, pag. 9-20. Valentini, A., Garuti, G., Rozzi, A., Tilche A. (1997). Anaerobic degradation kinetics of particulate organic matter: A new apporach. Water Science and Technology. Vol. 36. Nº 6-7, pp 239-246. Van Lier, J.B., Hulsbeek, J., Stams, A.J., Lettinga, G. (1993). Temperature susceptibility of thermophilic methanogenic sludge: implication for reactor start-up and operation. Bioresource technology. Vol. 43, pag. 227235. Van Lier, J.B. (1995). “Temperature optima of thermophilic methanogenic sludge: implications for reactor start-up and operation”, en Thermophilic anaerobic wastewater treatmewnt; temperature aspects and process stability. Tesis doctoral. Universidad de Wageningen. Van Velsen, A.F.M. (1979). Adaptation of methanogenic sludge to high ammonia-nitrogen concentrations. Water Research. Vol. 13, pag. 995999. Vavilin, V.A., Rytov, S.V., Lokshina, L.Y. (1996). A description of hydrolysis kinetics in anaerobic degradation of particulate organic matter. Vavilin, V.A., Vasiliev, V.B., Ponomarev, A.V. Rytov, S.V. (1994). Simulation model “methane” as a tool for effective biogas production during anaerobic conversion of complex organic matter. Bioresource technology. Vol. 48, pag 1-8. Vavilin, V.A., Vasiliev, V.B., Rytov, S.V., Ponomarev, A.V. (1995). Modeling ammonia and hydrogen sulfide inhibition in anaerobic digestion. Water Research. Vol. 29 (3), pag. 827-835. Veeken, A., Hamelers, B. (1999) Effect of temperature on hydrolysis rates of selectes biowaste components. Bioresource technology. Vol. 29, pag. 249-254 Viturtia, A.M., Mata-Alvárez, J., Cecchi, F., Fazzini, G. (1989). Two-phase anaerobic digestion of a mixture of fruit and vegetable wastes. Biological wastes. Vol 29, pag 189-199. Watanabe, H., Kitamura, T., Ochi, S., Ozaki, M. (1997). Inactivation of pathogenic bacteria under mesophilic and thermophilic conditions. Water Science and Technology. Vol. 36 (6-7), pag. 25-32. Wheatley, A.D. (1990). “Anaerobic digestion: industrial waste treatment”. En Ananerobic Digestion: a waste treatment technology. Editado por Wheatley, A.D. Critical reports on applied chemistry. Vol. 31, pag. 171223. Elsevier applied science LTD. Wong, M.H. (1990). Anaerobic digestion of pig manure mixed with sewage 320 Referencias bibliográficas sludge. Biological wastes. Vol. 31, pag. 223-230. Zeeman, G., Wiegant, W.M., Koster-Treffers, M.E., Lettinga, G. (1985). The influence of total ammonia concentration on the thermophilic digestion of cow manure. Agricultural Wastes. Vol. 14, pag. 19-35. 9. ANEJO 1. PROGRAMA DE SIMULACIÓN DE LA DIGESTIÓN ANAEROBIA DE SUBSTRATOS COMPLEJOS 322 Anejo 1 Optimización de la digestión anaerobia de purines de cerdo PROGRAM CONTINUOANG99 IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) COMMON/GRUP1/M,TR COMMON/GROUP2/VI,VL,VT,VG COMMON/GROUP3/YC,YP DIMENSION U(40),PM(40),DQO(40),XK(30),F(40),U0(40) CHARACTER NAME*12, RESP*2 EXTERNAL FSUB EXTERNAL TASAS T1=0. N=33 M=22 MM=1 7 FORMAT(A12) YC=0.5 YP=0.8 5 T0=0. WRITE(*,*)'ESCRIBE EL TIEMPO FINAL1' READ(*,*)TFINAL 6 FORMAT(A1) IF (MM.EQ.1) NAME='CONT1.DAT' IF (MM.EQ.2) NAME='CONT2.DAT' IF (MM.EQ.3) NAME='CONT3.DAT' IF (MM.EQ.4) NAME='CONT4.DAT' IF (MM.EQ.5) NAME='CONT5.DAT' IF (MM.EQ.6) NAME='CONT6.DAT' IF (MM.EQ.7) NAME='CONT7.DAT' IF (MM.EQ.8) NAME='CONT8.DAT' IF (MM.EQ.9) NAME='CONT9.DAT' IF (MM.EQ.10) NAME='CONT10.DAT' OPEN(3,FILE=NAME) PT=0.1 VL=4.95 VT=6.95 VG=VT-VL TEMP=35 WRITE(*,*)'TEMPERATURA POR DEFECTO', TEMP,'¿QUIERES CAMBIARLA?S/N' READ(*,6) RESP 323 324 Anejo 1 IF ((RESP.EQ.'S').OR.(RESP.EQ.'S')) THEN WRITE(*,*)'ESCRIBE LA NUEVA TEMPERATURA' READ(*,*)TEMP ENDIF PW=(0.148*TEMP**3-9.071428569*TEMP**2+410.1428567*TEMP& 3979.599985)/1.013250E+5 TR=15. WRITE(*,*)'TIEMPO DE RETENCION', TR,'¿QUIERES CAMBIARL0?S/N' READ(*,6) RESP IF ((RESP.EQ.'S').OR.(RESP.EQ.'S')) THEN WRITE(*,*)'ESCRIBE EL NUEVO TR' READ(*,*)TR ENDIF WRITE(*,*)'ESCRIBE EL NUMERO DE ARCHIVO DE CONC DE INFLUENTE' READ(*,*)NN IF (NN.EQ.1) NAME='CONI1.PRN' IF (NN.EQ.2) NAME='CONI2.PRN' IF (NN.EQ.3) NAME='CONI3.PRN' IF (NN.EQ.4) NAME='CONI4.PRN' IF (NN.EQ.5) NAME='CONI5.PRN' IF (NN.EQ.6) NAME='CONI6.PRN' IF (NN.EQ.7) NAME='CONI7.PRN' IF (NN.EQ.8) NAME='CONI8.PRN' IF (NN.EQ.9) NAME='CONI9.PRN' IF (NN.EQ.10) NAME='CONI10.PRN' OPEN(5,FILE=NAME) READ(5,*) (U0(I),I=1,N-1),PH U0(N)=10**(-PH) CLOSE(5) CALL MASASMOLEC(PM,DQO) DO I=1,29 U0(I)=U0(I)/PM(I) ENDDO PH0=8. HH0=U0(N) HH01=HH0 TEMP0=20. CALL SECANTE(TEMP0,U0,HH01) PH1=-DLOG10(HH01) Optimización de la digestión anaerobia de purines de cerdo 325 CALL SECANTE(TEMP,U0,HH0) PH2=-DLOG10(HH0) WRITE(*,*)'EL PH CAL. A 20ºC DEL INFLUENTE ES:',PH1 U0(N)=10**(-PH2) WRITE(*,*)'EL PH CAL. A LA TEMP', TEMP,'DEL INFLUENTE ES:',PH2 WRITE(*,*)'¿QUIERES CAMBIAR EL PH INICIAL (S/N)?' READ(*,6) RESP IF ((RESP.EQ.'S').OR.(RESP.EQ.'S')) THEN WRITE(*,*)'ESCRIBE EL PH MEDIDO' READ(*,*)PH U0(N)=10**(-PH) ENDIF DQOT=0. DO I=1,N-1 DQOT=DQOT+U0(I)*DQO(I) ENDDO DQOS=U0(11)*DQO(11)+U0(13)*DQO(13)+U0(16)*DQO(16)+U0(18)*DQO(18 & )+ U0(19)*DQO(19)+U0(20)*DQO(20)+U0(21)*DQO(21) DO I=1,29 U0(I)=U0(I)*PM(I) ENDDO OPEN(6,FILE='CONCINI.PRN') READ(6,*) (U(I),I=1,N-1),PH CLOSE(6) DO I=1,29 U(I)=U(I)/PM(I) ENDDO PH0=8. HH0=10**(-PH0) HH01=HH0 TEMP0=20. CALL SECANTE(TEMP0,U,HH01) PH1=-DLOG10(HH01) CALL SECANTE(TEMP,U,HH0) PH2=-DLOG10(HH0) WRITE(*,*)'EL PH CAL. A 20ºC ES:',PH1 U(N)=10**(-PH2) WRITE(*,*)'EL PH CAL. A LA TEMP', TEMP,'ES:',PH2 326 Anejo 1 WRITE(*,*)'¿QUIERES CAMBIAR EL PH INICIAL (S/N)?' READ(*,6) RESP IF ((RESP.EQ.'S').OR.(RESP.EQ.'S')) THEN WRITE(*,*)'ESCRIBE EL PH MEDIDO' READ(*,*)PH ENDIF U(N)=10**(-PH2) DO I=1,29 U(I)=U(I)*PM(I) ENDDO WRITE(3,*)'TFINAL,TEMP,(U(I),I=1,N-1),PH' IF (T1.EQ.INT(T1)) THEN WRITE(*,20)T1,U(17),PH,U(1),U(9),U(18),U(19),U(20),U(21),QG ENDIF HINIT=1.E-3 TOL=1.D-9 15 DO T=T0,TFINAL,PT T1=T+PT DO I=1,29 U0(I)=U0(I)/PM(I) U(I)=(U(I)/PM(I)) ENDDO CALL RFK(FSUB,N,TEMP,T,T1,HINIT,U,F,TOL,U0,QG) PH=-DLOG10(U(N)) CALL SECANTE(TEMP,U,HH01) PH1=-DLOG10(HH01) CALL CONSTANT(TEMP,XK) DQOT=0. DO I=1,N-1 DQOT=DQOT+U0(I)*DQO(I) ENDDO DQOS=U0(11)*DQO(11)+U0(13)*DQO(13)+U0(16)*DQO(16)+U0(18)*DQO(18 & )+U0(19)*DQO(19)+U0(20)*DQO(20)+U0(21)*DQO(21) DO I=1,29 U0(I)=U0(I)*PM(I) U(I)=(U(I)*PM(I)) ENDDO FNH3=1./(U(N)/XK(6)+1.) XNH3=U(17)*FNH3 Optimización de la digestión anaerobia de purines de cerdo 327 HH0=10**(-PH) YCH4=QG*U(31)*TR/(VL*DQOT) WRITE(*,20)T1,PH,PH1,U(17),U(19),U(20),U(21),U(22),QG,YCH4 WRITE(3,27)T1,(U(I),I=1,32),PH,XNH3,QG,YCH4 20 FORMAT(F7.0,7F7.2,3F8.2) 25 FORMAT(F7.2,23F10.4) 26 FORMAT (F10.2,24E8.2,3F10.3) 27 FORMAT (F10.2,32F12.4,4F12.4) ENDDO WRITE(*,*)'QUIERES CORRERLO DE NUEVO? SI/NO' READ(*,6)RESP IF ((RESP.EQ.'S').OR.(RESP.EQ.'S')) THEN WRITE(*,*)'QUIERES CONTINUAR A PARTIR DEL TIEMPO FINAL(S/N)' READ(*,6)RESP IF ((RESP.EQ.'S').OR.(RESP.EQ.'S')) THEN T0=T1 WRITE(*,*)'ESCRIBE EL NUEVO TIEMPO FINAL1' READ(*,*)TFINAL WRITE(*,*)'QUIERES CAMBIAR LAS CONC. DEL INFLUENTE(S/N)' READ(*,6)RESP IF ((RESP.EQ.'S').OR.(RESP.EQ.'S')) THEN WRITE(*,*)'ESCRIBE EL NUMERO DE ARCHIVO DE CONC INICIALES' READ(*,*)NN IF (NN.EQ.1) NAME='CONI1.PRN' IF (NN.EQ.2) NAME='CONI2.PRN' IF (NN.EQ.3) NAME='CONI3.PRN' IF (NN.EQ.4) NAME='CONI4.PRN' IF (NN.EQ.5) NAME='CONI5.PRN' IF (NN.EQ.6) NAME='CONI6.PRN' IF (NN.EQ.7) NAME='CONI7.PRN' IF (NN.EQ.8) NAME='CONI8.PRN' IF (NN.EQ.9) NAME='CONI9.PRN' IF (NN.EQ.10) NAME='CONI10.PRN' OPEN(5,FILE=NAME) READ(5,*) (U0(I),I=1,N-1),PH CLOSE(5) ENDIF WRITE(*,*)'QUIERES CAMBIAR LA TEMPERATURA? S/N' READ(*,6) RESP 328 Anejo 1 IF ((RESP.EQ.'S').OR.(RESP.EQ.'S')) THEN WRITE(*,*)'ESCRIBE LA NUEVA TEMPERATURA' READ(*,*)TEMP ENDIF PW=(0.148*TEMP**3-9.071428569*TEMP**2+410.1428567*TEMP& 3979.599985)/1.013250E+5 WRITE(*,*)'QUIERES CAMBIAR EL TIEMPO DE RETENCION', TR,'?S/N' READ(*,6) RESP IF ((RESP.EQ.'S').OR.(RESP.EQ.'S')) THEN WRITE(*,*)'ESCRIBE EL NUEVO TR' READ(*,*)TR ENDIF GOTO 15 ENDIF CLOSE(3) MM=MM+1 GOTO 5 ENDIF WRITE(*,*)'CERRAR EL PROGRAMA SI/NO(VOLVER A COMENZAR)' READ(*,6) RESP IF ((RESP.EQ.'S').OR.(RESP.EQ.'S')) GO TO 40 GO TO 5 40 RETURN END ******************************************************************** SUBROUTINE RFK(FSUB,N,TEMP,T0,TFINAL,HINIT,U,V6,TOL,U0,QG) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) PARAMETER (NMAX=33) DIMENSION U(NMAX),V1(NMAX),V2(NMAX),V3(NMAX),V4(NMAX), & V5(NMAX),V6(NMAX),U4(NMAX),U5(NMAX),UTEMP(NMAX) TK=T0 HT=HINIT 5 TKP1=MIN(TK+HT,TFINAL) HT=TKP1-TK TT=TK CALL FSUB(N,TEMP,U,V1,U0,QG) DO I=1,N UTEMP(I)=U(I)+HT*V1(I)/4 ENDDO Optimización de la digestión anaerobia de purines de cerdo TT=TK+HT/4. CALL FSUB(N,TEMP,UTEMP,V2,U0,QG) DO I=1,N UTEMP(I)=U(I)+HT*(3*V1(I)+9*V2(I))/32 ENDDO TT=TK+3.*HT/8. CALL FSUB(N,TEMP,UTEMP,V3,U0,QG) DO I=1,N UTEMP(I)=U(I)+HT*(1932*V1(I)-7200*V2(I)+7296*V3(I))/2197 ENDDO TT=TK+12.*HT/13. CALL FSUB(N,TEMP,UTEMP,V4,U0,QG) DO I=1,N UTEMP(I)=U(I)+HT*(8341*V1(I)-32832*V2(I)+29440*V3(I) & -845*V4(I))/4104 ENDDO TT=TK+HT CALL FSUB(N,TEMP,UTEMP,V5,U0,QG) DO I=1,N UTEMP(I)=U(I)+HT*(-6080*V1(I)+41040*V2(I)-28352*V3(I) & +9295*V4(I)-5643*V5(I))/20520 ENDDO TT=TK+HT/2. CALL FSUB(N,TEMP,UTEMP,V6,U0,QG) E=0.0 DO I=1,N U4(I)=U(I)+HT*(2375*V1(I)+11264*V3(I)+10985*V4(I) & -4104*V5(I))/20520 U5(I)=U(I)+HT*(33440*V1(I)+146432*V3(I)+142805*V4(I) & -50787*V5(I)+10260*V6(I))/282150 E=MAX(E,DABS((U4(I)-U5(I))/(HT))) ENDDO IF (E.GT.TOL) THEN HT=HT/2. GO TO 5 ENDIF TK=TKP1 DO I=1,N U(I)=U4(I) 329 330 Anejo 1 ENDDO IF (E.LT.(0.01*TOL)) THEN HT=HT*2. ENDIF IF (TK.LT.TFINAL) GO TO 5 RETURN 50 END ******************************************************************** SUBROUTINE FSUB(N,TEMP,U,F,U0,QG) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) COMMON/GRUP1/M,TR COMMON/GROUP2/VI,VL,VT,VG DIMENSION F(40),U(40),Y(40,40),R(30),U0(40) IF (U(N).LE.0.) U(N)=1.D-14 IF (U(N).GE.1.) U(N)=1. CALL PRODUC(N,M,TEMP,Y) CALL TASAS(N,M,TEMP,U,R) DO I=1,29 F(I)=1./TR*(U0(I)-U(I)) DO J=1,M F(I)=F(I)+Y(I,J)*R(J) ENDDO ENDDO PW=(0.148*TEMP**3-9.071428569*TEMP**2+410.1428567*TEMP& 3979.599985)/1.013250E+5 QG=-0.082*(TEMP+273)*VL/(1.-PW)*(R(19)+R(20)+R(21)) DO I=30,32 F(I)=-QG/VG*U(I) DO J=1,M-1 F(I)=F(I)+Y(I,J)*R(J) ENDDO ENDDO CALL CHFUN(N,U,TEMP,F,FH) F(N)=FH RETURN END SUBROUTINE CHFUN(N,U,TEMP,F,FH) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION U(40),F(40),F1(40,40),DF1(40,40),XK(30),C1(30) Optimización de la digestión anaerobia de purines de cerdo 331 NC=6 CALL CONSTANT(TEMP,XK) CALL MATRIZF(N,NC,U,XK,F1,DF1) CALL CARGAS(NC,C1) CH=0. DO I=1,N DO J=1,NC CH=CH+U(I)*F1(I,J)*C1(J) ENDDO ENDDO VAL1=0. DO I=1,N DO J=1,NC VAL1=VAL1+F(I)*F1(I,J)*C1(J) ENDDO ENDDO VAL2=0. DO I=1,N DO J=1,NC VAL2=VAL2+U(I)*DF1(I,J)*C1(J) ENDDO ENDDO IF (CH.GE.0) THEN B=(1.+CH/DSQRT(CH**2+4.*XK(10)))/2. ELSE B=-(U(N)**2/(2*XK(10)))*(-1.+CH/DSQRT(CH**2+4.*XK(10))) ENDIF FH=(B*VAL1)/(1.-B*VAL2) RETURN END ******************************************************************** SUBROUTINE MATRIZF(N,NC,U,XK,F1,DF1) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION F1(40,40),DF1(40,40),XK(20),U(30) DO I=1,N DO J=1,NC F1(I,J)=0. DF1(I,J)=0. ENDDO 332 Anejo 1 ENDDO F1(18,4)=XK(12)/(U(N)+XK(12)) F1(20,4)=XK(1)/(U(N)+XK(1)) F1(19,4)=XK(2)/(U(N)+XK(2)) F1(21,4)=XK(3)/(U(N)+XK(3)) F1(22,3)=(U(N)**2)/(U(N)**2+XK(4)*U(N)+XK(4)*XK(5)) F1(22,4)=(XK(4)*U(N))/(U(N)**2+XK(4)*U(N)+XK(4)*XK(5)) F1(22,5)=(XK(4)*XK(5))/(U(N)**2+XK(4)*U(N)+XK(4)*XK(5)) F1(17,2)=U(N)/(XK(6)+U(N)) F1(17,3)=XK(6)/(XK(6)+U(N)) F1(25,4)=(XK(7)*U(N)**2)/(U(N)**3+XK(7)*U(N)**2+ & XK(7)*XK(8)*U(N)+XK(7)*XK(8)*XK(9)) F1(25,5)=XK(7)*XK(8)*U(N)/(U(N)**3+XK(7)*U(N)**2+XK(7)*XK(8)* & U(N)+XK(7)*XK(8)*XK(9)) F1(25,6)=XK(7)*XK(8)*XK(9)/(U(N)**3+XK(7)*U(N)**2+XK(7)*XK(8)* & U(N)+XK(7)*XK(8)*XK(9)) F1(26,4)=1. F1(27,5)=1. F1(28,2)=1. F1(29,1)=1. DF1(18,4)=-XK(12)/(U(N)+XK(12))**2 DF1(20,4)=-XK(1)/(U(N)+XK(1))**2 DF1(19,4)=-XK(2)/(U(N)+XK(2))**2 DF1(21,4)=-XK(3)/(U(N)+XK(3))**2 DF1(22,4)=XK(4)*(-U(N)**2+XK(4)*XK(5))/(U(N)**2+XK(4)*U(N)+ & XK(4)*XK(5))**2 DF1(22,5)=-(XK(4)*XK(5)*(2.*U(N)+XK(4)))/(U(N)**2+XK(4)*U(N)+ & XK(4)*XK(5))**2 DF1(17,2)=XK(6)/(XK(6)+U(N))**2 DF1(25,4)=XK(7)*(-U(N)**4+XK(8)*XK(7)*U(N)**2+ & 2.*XK(9)*XK(7)*XK(8)*U(N))/(U(N)**3+ XK(7)*U(N)**2+ & XK(7)*XK(8)*U(N)+XK(9)*XK(7)*XK(8))**2 DF1(25,5)=XK(7)*XK(8)*(-2.*U(N)**3-XK(7)*U(N)**2+ & XK(9)*XK(7)*XK(8))/(U(N)**3+ XK(7)*U(N)**2+ & XK(7)*XK(8)*U(N)+XK(9)*XK(7)*XK(8))**2 DF1(25,6)=XK(7)*XK(8)*XK(9)*(3.*U(N)**2+ & 2.*XK(7)*U(N)+XK(7)*XK(8))/(U(N)**3+ XK(7)*U(N)**2+ & XK(7)*XK(8)*U(N)+XK(9)*XK(7)*XK(8))**2 RETURN Optimización de la digestión anaerobia de purines de cerdo 333 END ******************************************************************** SUBROUTINE CARGAS(NC,C1) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION C1(30) C1(1)=-2. C1(2)=-1. C1(3)=0. C1(4)=1. C1(5)=2. C1(NC)=3. RETURN END ******************************************************************** SUBROUTINE PRODUC(N,M,TEMP,Y) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) COMMON/GROUP2/VI,VL,VT,VG COMMON/GROUP3/YC,YP DIMENSION Y(40,40) DO I=1,N-1 DO J=1,M-1 Y(I,J)=0. ENDDO ENDDO Y(1,11)=-1 Y(2,12)=-1 Y(3,13)=-1 Y(4,14)=-1 Y(5,15)=-1 Y(6,16)=-1 Y(7,17)=-1 Y(8,18)=-1 DO J=11,18 Y(9,J)=0.27833 Y(14,J)=3.3333 ENDDO Y(9,1)=-1. Y(10,1)=1-YC Y(11,1)=YC 334 Anejo 1 Y(14,2)=-1. Y(15,2)=1-YP Y(16,2)=YP Y(1,3)=1. Y(11,3)=-8.9686 Y(17,3)=-1. Y(19,3)=3.9543 Y(20,3)=4.4843 Y(21,3)=6.67266 Y(22,3)=6.1964 Y(2,4)=1. Y(16,4)=-58.778 Y(17,4)=16.633 Y(18,4)=0.7689 Y(19,4)=1.3417 Y(20,4)=1.7069 Y(21,4)=17.482 Y(22,4)=4.4242 Y(24,4)=0.058889 Y(3,5)=1. Y(12,5)=-24.562 Y(13,5)=73.687 Y(17,5)=-1. Y(20,5)=23.134 Y(22,5)=-0.7148 Y(4,6)=1. Y(13,6)=-3.9417 Y(17,6)=-1. Y(21,6)=34.292 Y(22,6)=-16.095 Y(23,6)=13.456 Y(5,7)=1. Y(17,7)=-1. Y(18,7)=-15.314 Y(20,7)=0.4447 Y(21,7)=13.648 Y(22,7)=-8.4885 Y(23,7)=6.8208 Y(6,8)=1. Optimización de la digestión anaerobia de purines de cerdo 335 Y(17,8)=-1. Y(19,8)=-15.314 Y(21,8)=28.957 Y(22,8)=-8.4885 Y(23,8)=6.8178 Y(7,9)=1. Y(17,9)=-1. Y(20,9)=-16.134 Y(21,9)=15.0777 Y(22,9)=2.593 Y(23,9)=10.655 Y(8,10)=1. Y(17,10)=-1. Y(21,10)=-45.455 Y(22,10)=42.955 Y(23,10)=42.955 CGL=(VL/VG)*0.0821*(273+TEMP) Y(22,19)=1. Y(30,19)=-CGL Y(23,20)=1. Y(31,20)=-CGL Y(17,21)=1. Y(32,21)=-CGL Y(N,M)=1. RETURN END ******************************************************************** SUBROUTINE TASAS(N,M,TEMP,U,R) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) COMMON/GROUP2/VI,VL,VT,VG DIMENSION U(40),R(40),XKIA(20),XKIB(20), XKSA(20) DIMENSION RMAX(20),A(20),TOPT(20),TMAX(20) DIMENSION XK(20),XKX(20),F1(40,10) RMAX(1)=1. RMAX(2)=1. RMAX(3)=5.1 RMAX(4)=6.38 RMAX(5)=2.86 RMAX(6)=0.57 336 Anejo 1 RMAX(7)=0.69 RMAX(8)=0.67 RMAX(9)=0.49 RMAX(10)=0.6 TOPT(1)=55 TOPT(3)=55 TOPT(8)=60 TOPT(9)=53 TOPT(10)=55 TMAX(1)=65 TMAX(3)=65 TMAX(8)=70 TMAX(9)=65 TMAX(10)=65 A(1)=0. A(3)=0. A(8)=0.00018 A(9)=0.00017 A(10)=0.00017 IF(TEMP.LE.TOPT(1)) THEN RMAX(1)=RMAX(1)-A(1)*(TOPT(1)-TEMP) ELSE RMAX(1)=RMAX(1)*(TMAX(1)-TEMP)/(TMAX(1)-TOPT(1)) ENDIF IF(TEMP.LE.TOPT(3)) THEN RMAX(3)=RMAX(3)-A(3)*(TOPT(3)-TEMP) ELSE RMAX(3)=RMAX(3)*(TMAX(3)-TEMP)/(TMAX(3)-TOPT(3)) ENDIF IF(TEMP.LE.TOPT(8)) THEN RMAX(8)=RMAX(8)-A(8)*(TOPT(8)-TEMP) ELSE RMAX(8)=RMAX(8)*(TMAX(8)-TEMP)/(TMAX(8)-TOPT(8)) ENDIF IF(TEMP.LE.TOPT(9)) THEN RMAX(9)=RMAX(9)-A(9)*(TOPT(9)-TEMP) ELSE RMAX(9)=RMAX(9)*(TMAX(9)-TEMP)/(TMAX(9)-TOPT(9)) ENDIF Optimización de la digestión anaerobia de purines de cerdo IF(TEMP.LE.TOPT(10)) THEN RMAX(10)=RMAX(10)-A(10)*(TOPT(10)-TEMP) ELSE RMAX(10)=RMAX(10)*(TMAX(10)-TEMP)/(TMAX(10)-TOPT(10)) ENDIF CALL CONSTANT(TEMP,XK) CALL MATRIZF(N,NC,U,XK,F1,DF1) XNH3=U(17)*F1(17,3) VFA=U(18)+U(19)+U(20)+U(21) XKIA(11)=0. XKIA(1)=0.33/60. XKIA(2)=0.33/60. XKIA(3)=0. XKIA(4)=0. XKIA(5)=0. XKIA(6)=0. XKIA(7)=0.4/60. XKIA(8)=0.72/60. XKIA(9)=0.96/60. XKIA(10)=0.26/14. XKIB(1)=0. XKIB(2)=0. XKIB(3)=0.2/282 XKIB(4)=0. DO J=5,10 XKIB(J)=0.2/282 ENDDO XKSA(11)=0. XKSA(1)=0. XKSA(2)=0. XKSA(3)=0.5/160. XKSA(4)=0. XKSA(5)=0.02/884. XKSA(6)=0.04/282. XKSA(7)=0.175/102. XKSA(8)=0.176/88. XKSA(9)=0.259/74. XKSA(10)=0.12/60. FFH=(1.+2.*10**(-1.25))/(1.+(10**(-8.5))/U(N)+U(N)*10**(6.)) 337 338 Anejo 1 R(1)=RMAX(1)*XKIA(1)/(XKIA(1)+VFA)*U(9) XKX(1)=0.1/162. R(2)=RMAX(2)*XKIA(2)/(XKIA(2)+VFA)*U(14) XKX(2)=0.1/38.2 R(3)=RMAX(3)*(U(11)/(U(11)+XKSA(3)))*(1/(1+U(13)/XKIB(3)))*U(1) R(4)=(RMAX(4)*U(16))*U(2) R(5)=RMAX(5)*(U(12)/(U(12)+XKSA(5)))*(1/(1+U(13)/XKIB(5)))*U(3) R(6)=RMAX(6)*(U(13)*XKIB(6)/(U(13)*XKIB(6)+XKSA(6)*XKIB(6)+ & U(13)**2))*FFH*U(4) R(7)=RMAX(7)*(U(18)/(U(18)+XKSA(7)))* &(1/(1+U(21)/XKIA(7)))*(1/(1+U(13)/XKIB(7)))*FFH*U(5) R(8)=RMAX(8)*(U(19)/(U(19)+XKSA(8)))* &(1/(1+U(21)/XKIA(8)))*(1/(1+U(13)/XKIB(8)))*FFH*U(6) R(9)=RMAX(9)*(U(20)/(U(20)+XKSA(9)))* &(1/(1+U(21)/XKIA(9)))*(1/(1+U(13)/XKIB(9)))*FFH*U(7) R(10)=RMAX(10)*(U(21)/(U(21)+XKSA(10)))* &(1/(1+XNH3/XKIA(10)))*(1/(1+U(13)/XKIB(10)))*FFH*U(8) R(11)=0.05*RMAX(3)*U(1) R(12)=0.05*RMAX(4)*U(2) R(13)=0.05*RMAX(5)*U(3) R(14)=0.05*RMAX(6)*U(4) R(15)=0.05*RMAX(7)*U(5) R(16)=0.05*RMAX(8)*U(6) R(17)=0.05*RMAX(9)*U(7) R(18)=0.05*RMAX(10)*U(8) XKLACO2=55. XKLACH4=55. XKLANH3=1. XKHCO2=0.0697-0.002*TEMP+2.56E-5*TEMP**2-1.21E-7*TEMP**3 XKHCH4=EXP(-60.954+85.8073/((273.15+TEMP)/100.)+ & 23.4487*DLOG((273.15+TEMP)/100.)) XKHNH3=52.9-1.454*TEMP+0.021*TEMP**2-1.13E-4*TEMP**3 R(19)=XKLACO2*(XKHCO2*U(30)-U(22)*F1(22,3)) R(20)=XKLACH4*(XKHCH4*U(31)-U(23)) R(M-1)=XKLANH3*(XKHNH3*U(32)-XNH3) RETURN END ******************************************************************** Optimización de la digestión anaerobia de purines de cerdo SUBROUTINE MASASMOLEC(PM,DQO) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION PM(40),DQO(40) DO I=1,8 PM(I)=113. ENDDO PM(9)=162. PM(10)=162. PM(11)=162. PM(12)=884. PM(13)=282 PM(14)=27.862 PM(15)=27.862 PM(16)=27.862 PM(17)=14. PM(18)=102. PM(19)=88. PM(20)=74. PM(21)=60. PM(22)=44. PM(23)=16. PM(24)=34. PM(25)=31. PM(26)=35.5 PM(27)=96. PM(28)=39. PM(29)=32. DO I=1,8 DQO(I)=160. ENDDO DO I=9,11 DQO(I)=192. ENDDO DQO(12)=2560. DQO(13)=816. DO I=14,16 DQO(I)=31.424 ENDDO DQO(18)=208. 339 340 Anejo 1 DQO(19)=160. DQO(20)=112. DQO(21)=64. DO I=22,29 DQO(I)=0. ENDDO RETURN END ******************************************************************** SUBROUTINE CONSTANT(TEMP,XK) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION C(20,20),PK(20),XK(20) *1:PROPIONICO; 2:BUTÍRICO; 3:ACETICO; 4 Y 5:1ª Y 2ºCARBÓNICO; *6:AMONIO; 7,8,Y 9:1ª,2ª,3ºFOSFÓRICO; 10:AGUA; 12:VALÉRICO NC=11 C(3,1)=4.7803 C(3,2)= -0.0023 C(3,3)= 6.E-5 C(3,4)=-2.E-7 C(2,1)=4.8947 C(2,2)= -0.0024 C(2,3)= 7.E-5 C(2,4)=-3.E-7 C(1,1)=4.8063 C(1,2)= -0.0011 C(1,3)= 8.E-5 C(1,4)=-4.E-7 C(4,1)=6.5787 C(4,2)= -0.0133 C(4,3)= 0.0002 C(4,4)=-8.E-7 C(5,1)=10.619 C(5,2)= -0.010 C(5,3)= 1.01E-7 C(5,4)=0. C(7,1)=2.0473 C(7,2)=0.0019 C(7,3)=5.E-5 C(7,4)=0. Optimización de la digestión anaerobia de purines de cerdo C(8,1)=7.3144 C(8,2)=-0.0073 C(8,3)=0.0001 C(8,4)=-6.E-7 C(9,1)=12.6576 C(9,2)=0. C(9,3)=0. C(9,4)=0. C(6,1)=10.072 C(6,2)=-0.0356 C(6,3)=9.E-5 C(6,4)=4.E-8 C(10,1)=14.934 C(10,2)=-0.0425 C(10,3)=0.0002 C(10,4)=-6.E-7 DO I=1,10 C(10,5)=0. ENDDO DO I=1,NC-1 PK(I)=0. ENDDO DO I=1,NC-1 DO J=1,5 PK(I)=PK(I)+C(I,J)*TEMP**(J-1) ENDDO XK(I)=10**(-PK(I)) ENDDO C(11,1)=0.0134 C(11,2)=-0.0004 C(11,3)=8.E-6 C(11,4)=-7.E-8 C(11,5)=3.E-10 XK(11)=0. DO J=1,5 XK(11)=XK(11)+C(11,J)*TEMP**(J-1) ENDDO PK(12)=PK(1) XK(12)=XK(1) 341 342 Anejo 1 RETURN END ******************************************************************** SUBROUTINE SECANTE(TEMP,U,X) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION U(30) TOL=1.E-4 PX=-DLOG10(X) I=1 F=1.0001 5 PX1=PX*F PX2=PX/F 10 CALL FUNCH(TEMP,U,PX1,F1) CALL FUNCH(TEMP,U,PX2,F2) IF((F1*F2).GT.0.) THEN F=F*1.1 GOTO 5 ENDIF PX3=PX2-(PX1-PX2)/(F1/F2-1.) CALL FUNCH(TEMP,U,PX3,F3) IF((F1*F3).GT.0.)THEN PX1=PX3 ELSE PX2=PX3 ENDIF ERR1=DABS(PX2-PX1) ERR2=DABS(F3) ERR=MAX(ERR1,ERR2) ERR0=MIN(ERR1,ERR2) I=I+1 IF(ERR.GT.TOL) THEN IF (ERR0.LT.TOL/1.E2) GO TO 20 GO TO 10 ENDIF 20 PX=PX3 X=10**(-PX) RETURN END ******************************************************************** Optimización de la digestión anaerobia de purines de cerdo 343 SUBROUTINE FUNCH(TEMP,U,PH,FHH) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION U(40),A(40),XK(20) CALL CONSTANT(TEMP,XK) HH=10**(-PH) CALL COEF1(HH,XK,A) CH=0. DO I=17,31 CH=CH+A(I)*U(I) ENDDO IF (CH.GE.0.) THEN PH1=(-DLOG10((CH+DSQRT(CH**2+4.*XK(10)))/2.)) ELSE PH1=(-DLOG10(2.*XK(10)/(-CH+DSQRT(CH**2+4.*XK(10))))) ENDIF FHH=PH-PH1 RETURN END ******************************************************************** SUBROUTINE COEF1(HH,XK,A) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION A(40),XK(20) DO I=17,29 A(I)=0. ENDDO A(20)=XK(1)/(HH+XK(1)) A(19)=XK(2)/(HH+XK(2)) A(21)=XK(3)/(HH+XK(3)) A(18)=XK(12)/(HH+XK(12)) A(23)=0. A(22)=(XK(4)*HH+2.*XK(4)*XK(5))/(HH**2+XK(4)*HH+XK(4)*XK(5)) A(17)=-HH/(XK(6)+HH) A(25)=(XK(7)*HH**2+2.*XK(7)*XK(8)*HH+3.*XK(7)*XK(8)*XK(9))/ & (HH**3+XK(7)*HH**2+XK(7)*XK(8)*HH+XK(7)*XK(8)*XK(9)) A(26)=1. A(27)=2. A(28)=-1. A(29)=-2. RETURN 344 Anejo 1 END Optimización de la digestión anaerobia de purines de cerdo 345 10. ANEJO 2. PROGRAMA DE SIMULACIÓN DEL PH EN SISTEMAS ACUOSOS SENCILLOS. Optimización de la digestión anaerobia de purines de cerdo 347 348 Anejo 2 PROGRAM PROGPH IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) COMMON/GRUP1/M COMMON/GROUP2/U0,V0,VG DIMENSION U(40),U0(40),Q(40) CHARACTER NAME*12, RESP*2 EXTERNAL FSUB EXTERNAL TASAS T1=0. N=33 M=22 5 T0=0. 6 FORMAT(A1) 7 FORMAT(A12) V0=0.1 VG=5.*5.*5.*1000. DO I=1,N-1 U0(I)=0. ENDDO DO I=1,N-1 U(I)=0. ENDDO MM=1 WRITE(*,*)'MM=',MM IF (MM.EQ.1) NAME='PH-1.DAT' IF (MM.EQ.2) NAME='PH-2.DAT' IF (MM.EQ.3) NAME='PH-3.DAT' IF (MM.EQ.4) NAME='PH-4.DAT' IF (MM.EQ.5) NAME='PH-5.DAT' IF (MM.EQ.6) NAME='PH-6.DAT' IF (MM.EQ.7) NAME='PH-7.DAT' IF (MM.EQ.8) NAME='PH-8.DAT' IF (MM.EQ.9) NAME='PH-9.DAT' IF (MM.EQ.10) NAME='PH-10.DAT' OPEN(3,FILE=NAME) WRITE(*,*)'VOLUMEN INICIAL POR DEFECTO ES 100 ML & ¿QUIERES CAMBIARLO?(SI/NO)' READ(*,6) RESP IF ((RESP.EQ.'S').OR.(RESP.EQ.'S')) THEN Optimización de la digestión anaerobia de purines de cerdo 349 WRITE(*,*) 'ESCRIBE EL VOLUMEN INICIAL(ML)' READ(*,*)V0 ENDIF V=V0 TEMP=20. WRITE(*,*)'¿QUIERES CAMBIAR LA TEMPERATURA (20ºC)? SI/NO' READ(*,6) RESP IF ((RESP.EQ.'S').OR.(RESP.EQ.'S')) THEN WRITE(*,*)'ESCRIBE LA TEMPERATURA' READ(*,*)TEMP ENDIF PT=0.1 WRITE(*,*)'LAS CONC. INICIALES SON CERO, QUIERES CAMBIAR ALGUNA & (SI/NO)' READ(*,6) RESP IF ((RESP.EQ.'S').OR.(RESP.EQ.'S')) THEN 10 WRITE(*,*) 'ESCRIBE EL ÍNDICE Y LA CONC INICIAL' READ(*,*)I,U(I) WRITE(*,*)'QUIERES CAMBIAR OTRA (SI/NO)' READ(*,6) RESP IF ((RESP.EQ.'S').OR.(RESP.EQ.'S')) GOTO 10 ENDIF WRITE(*,*)'ESCRIBE EL PH INICIAL' READ(*,*)PH HH=10**(-PH) U(N)=HH WRITE(3,*)'TFINAL,TEMP,(U(I),I=1,N-1), PH' WRITE(3,25)TFINAL,TEMP,(U(I),I=1,N-1), PH HINIT=1.E-3 TOL=1.D-7 WRITE(*,*)'ESCRIBE EL NUMERO DE ARCHIVO DE CONC INFLUENTE' READ(*,*)NN IF (NN.EQ.1) NAME='CONI1.PRN' IF (NN.EQ.2) NAME='CONI2.PRN' IF (NN.EQ.3) NAME='CONI3.PRN' IF (NN.EQ.4) NAME='CONI4.PRN' IF (NN.EQ.5) NAME='CONI5.PRN' IF (NN.EQ.6) NAME='CONI6.PRN' 350 Anejo 2 IF (NN.EQ.7) NAME='CONI7.PRN' IF (NN.EQ.8) NAME='CONI8.PRN' IF (NN.EQ.9) NAME='CONI9.PRN' IF (NN.EQ.10) NAME='CONI10.PRN' OPEN(5,FILE=NAME) 12 CALL CARACTER(TFINAL,N,U0,Q) IF (TFINAL.EQ.0) GO TO 35 15 DO T=T0,TFINAL,PT T1=T+PT CALL RFK(FSUB,N,TEMP,T,T1,HINIT,U,TOL,Q,V) PH=-DLOG10(U(N)) WRITE(*,20)T1,PH,U(17),U(21),U(22),U(23),V WRITE(3,25)T1,(U(I),I=1,N-1),PH,V 20 25 FORMAT(F8.1,6F11.5) FORMAT(F8.3,34F10.6) ENDDO WRITE(*,*)'QUIERES CONTINUAR? SI/NO' READ(*,6)RESP IF ((RESP.EQ.'N').OR.(RESP.EQ.'N')) THEN GO TO 35 ENDIF T0=T1 GO TO 12 CLOSE(3) CLOSE(5) GOTO 5 35 WRITE(*,*)'QUIERES CORRERLO DE NUEVO SI/NO' READ(*,6) RESP IF ((RESP.EQ.'N').OR.(RESP.EQ.'N')) GO TO 40 MM=MM+1 GO TO 5 40 RETURN END *************************************************************** SUBROUTINE CARACTER(TFINAL,N,U0,Q) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION U0(40),Q(40) CHARACTER RESP*2 6 FORMAT(A1) Optimización de la digestión anaerobia de purines de cerdo 351 READ(5,*) TFINAL WRITE(*,*)'EL TIEMPO FINAL ES',TFINAL PAUSE DO I=1,N-1 READ(5,*) U0(I),Q(I) ENDDO DO I=1,N-1 Q(I)=Q(I)/1000. ENDDO DO I=1,N-1 WRITE(*,*)'CONCENTRACION MADRE', I, U0(I) ENDDO WRITE(*,*)'QUIERES CAMBIAR ALGUNA (S/N)' READ(*,6) RESP IF ((RESP.EQ.'S').OR.(RESP.EQ.'S')) THEN 10 WRITE(*,*)'ESCRIBE EL ÍNDICE Y LA CONCENRTAC' READ(*,*)I, U0(I) WRITE(*,*)'OTRA?' READ (*,6) RESP IF ((RESP.EQ.'S').OR.(RESP.EQ.'S')) THEN GOTO 10 ENDIF ENDIF RETURN END ******************************************************************** SUBROUTINE RFK(FSUB,N,TEMP,T0,TFINAL,HINIT,U,TOL,Q,V) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) PARAMETER (NMAX=33) DIMENSION U(40),V1(NMAX),V2(NMAX),V3(NMAX),V4(NMAX),V5(NMAX), & V6(NMAX),U4(NMAX),U5(NMAX),UTEMP(NMAX),Q(NMAX) TK=T0 HT=HINIT 5 TKP1=MIN(TK+HT,TFINAL) HT=TKP1-TK TT=TK CALL FSUB(N,TT,TEMP,U,V1,Q,V) DO I=1,N UTEMP(I)=U(I)+HT*V1(I)/4 352 Anejo 2 ENDDO TT=TK+HT/4. CALL FSUB(N,TT,TEMP,UTEMP,V2,Q,V) DO I=1,N UTEMP(I)=U(I)+HT*(3*V1(I)+9*V2(I))/32 ENDDO TT=TK+3.*HT/8. CALL FSUB(N,TT,TEMP,UTEMP,V3,Q,V) DO I=1,N UTEMP(I)=U(I)+HT*(1932*V1(I)-7200*V2(I)+7296*V3(I))/2197 ENDDO TT=TK+12.*HT/13. CALL FSUB(N,TT,TEMP,UTEMP,V4,Q,V) DO I=1,N UTEMP(I)=U(I)+HT*(8341*V1(I)-32832*V2(I)+29440*V3(I) & -845*V4(I))/4104 ENDDO TT=TK+HT CALL FSUB(N,TT,TEMP,UTEMP,V5,Q,V) DO I=1,N UTEMP(I)=U(I)+HT*(-6080*V1(I)+41040*V2(I)-28352*V3(I) & +9295*V4(I)-5643*V5(I))/20520 ENDDO TT=TK+HT/2. CALL FSUB(N,TT,TEMP,UTEMP,V6,Q,V) E=0.0 DO I=1,N U4(I)=U(I)+HT*(2375*V1(I)+11264*V3(I)+10985*V4(I) & -4104*V5(I))/20520 U5(I)=U(I)+HT*(33440*V1(I)+146432*V3(I)+142805*V4(I) & -50787*V5(I)+10260*V6(I))/282150 E=MAX(E,DABS((U4(I)-U5(I))/(HT))) ENDDO IF (E.GT.TOL) THEN HT=HT/2. GO TO 5 ENDIF TK=TKP1 DO I=1,N Optimización de la digestión anaerobia de purines de cerdo 353 U(I)=U4(I) ENDDO IF (E.LT.(0.01*TOL)) THEN HT=HT*2. ENDIF IF (TK.LT.TFINAL) GO TO 5 RETURN 50 END ******************************************************************** SUBROUTINE FSUB(N,T,TEMP,U,F,Q,V) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) COMMON/GRUP1/M COMMON/GROUP2/U0,V0,VG DIMENSION F(40),U(40),Y(40,40),R(30),U0(40),Q(40) DIMENSION F1(40,40), DF1(40,40) IF (U(N).LE.0.) U(N)=1.D-14 IF (U(N).GE.1.) U(N)=1. CALL PRODUC(N,M,TEMP,Y,V) CALL TASAS(N,M,TEMP,U,R) CALL MATRIZF(N,NC,U,XK,F1,DF1) QT=0. DO I=1,N QT=QT+Q(I) ENDDO V=V0+QT*T DO I=1,29 F(I)=(U0(I)*Q(I)-U(I)*QT)/(V0+QT*T) DO J=1,M-1 F(I)=F(I)+Y(I,J)*R(J) ENDDO ENDDO PW=(0.148*TEMP**3-9.071428569*TEMP**2+410.1428567*TEMP& 3979.599985)/1.013250E+5 DO I=30,32 F(I)=-U(30)/VG*U(I) DO J=2,M-1 F(I)=F(I)+Y(I,J)*R(J) ENDDO ENDDO 354 Anejo 2 CALL CHFUN(N,U,TEMP,F,FH) F(N)=FH RETURN END ******************************************************************** SUBROUTINE CHFUN(N,U,TEMP,F,FH) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION U(40),F(40),F1(40,40),DF1(40,40),XK(30),C1(30) NC=6 CALL CONSTANT(TEMP,XK) CALL MATRIZF(N,NC,U,XK,F1,DF1) CALL CARGAS(NC,C1) CH=0. DO I=1,N DO J=1,NC CH=CH+U(I)*F1(I,J)*C1(J) ENDDO ENDDO VAL1=0. DO I=1,N DO J=1,NC VAL1=VAL1+F(I)*F1(I,J)*C1(J) ENDDO ENDDO VAL2=0. DO I=1,N DO J=1,NC VAL2=VAL2+U(I)*DF1(I,J)*C1(J) ENDDO ENDDO IF (CH.GE.0) THEN B=(1.+CH/DSQRT(CH**2+4.*XK(10)))/2. ELSE B=-(U(N)**2/(2*XK(10)))*(-1.+CH/DSQRT(CH**2+4.*XK(10))) ENDIF FH=(B*VAL1)/(1.-B*VAL2) RETURN END ******************************************************************** Optimización de la digestión anaerobia de purines de cerdo 355 SUBROUTINE MATRIZF(N,NC,U,XK,F1,DF1) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION F1(40,40),DF1(40,40),XK(20),U(30) DO I=1,N DO J=1,NC F1(I,J)=0. DF1(I,J)=0. ENDDO ENDDO F1(18,3)=U(N)/(U(N)+XK(12)) F1(18,4)=XK(12)/(U(N)+XK(12)) F1(20,3)=U(N)/(U(N)+XK(1)) F1(20,4)=XK(1)/(U(N)+XK(1)) F1(19,3)=U(N)/(U(N)+XK(2)) F1(19,4)=XK(2)/(U(N)+XK(2)) F1(21,3)=U(N)/(U(N)+XK(3)) F1(21,4)=XK(3)/(U(N)+XK(3)) F1(22,3)=(U(N)**2)/(U(N)**2+XK(4)*U(N)+XK(4)*XK(5)) F1(22,4)=(XK(4)*U(N))/(U(N)**2+XK(4)*U(N)+XK(4)*XK(5)) F1(22,5)=(XK(4)*XK(5))/(U(N)**2+XK(4)*U(N)+XK(4)*XK(5)) F1(17,2)=U(N)/(XK(6)+U(N)) F1(17,3)=XK(6)/(XK(6)+U(N)) F1(25,3)=(U(N)**3)/(U(N)**3+XK(7)*U(N)**2+XK(7)*XK(8)*U(N)+ & XK(7)*XK(8)*XK(9)) F1(25,4)=(XK(7)*U(N)**2)/(U(N)**3+XK(7)*U(N)**2+XK(7)*XK(8)* & U(N)+XK(7)*XK(8)*XK(9)) F1(25,5)=XK(7)*XK(8)*U(N)/(U(N)**3+XK(7)*U(N)**2+XK(7)*XK(8)* & U(N)+XK(7)*XK(8)*XK(9)) F1(25,6)=XK(7)*XK(8)*XK(9)/(U(N)**3+XK(7)*U(N)**2+XK(7)*XK(8)* & U(N)+XK(7)*XK(8)*XK(9)) F1(26,4)=1. F1(27,5)=1. F1(28,2)=1. F1(29,1)=1. DF1(18,4)=-XK(12)/(U(N)+XK(12))**2 DF1(18,3)=XK(12)/(U(N)+XK(12))**2 DF1(20,4)=-XK(1)/(U(N)+XK(1))**2 DF1(20,3)=XK(1)/(U(N)+XK(1))**2 DF1(19,4)=-XK(2)/(U(N)+XK(2))**2 356 Anejo 2 DF1(19,3)=XK(2)/(U(N)+XK(2))**2 DF1(21,4)=-XK(3)/(U(N)+XK(3))**2 DF1(21,3)=XK(3)/(U(N)+XK(3))**2 DF1(22,3)=XK(4)*(U(N)**2+2.*XK(5)*U(N))/(U(N)**2+XK(4)*U(N)+ & XK(4)*XK(5))**2 DF1(22,4)=XK(4)*(-U(N)**2+XK(4)*XK(5))/(U(N)**2+XK(4)*U(N)+ & XK(4)*XK(5))**2 DF1(22,5)=-(XK(4)*XK(5)*(2.*U(N)+XK(4)))/(U(N)**2+XK(4)*U(N)+ & XK(4)*XK(5))**2 DF1(17,2)=XK(6)/(XK(6)+U(N))**2 DF1(17,3)=-XK(6)/(XK(6)+U(N))**2 DF1(25,3)=XK(7)*(U(N)**4+2*XK(8)*U(N)**3+ &3.*XK(8)*XK(9)*U(N)**2)/(U(N)**3+ XK(7)*U(N)**2+ & XK(7)*XK(8)*U(N)+XK(9)*XK(7)*XK(8))**2 DF1(25,4)=XK(7)*(-U(N)**4+XK(8)*XK(7)*U(N)**2+ &2.*XK(9)*XK(7)*XK(8)*U(N))/(U(N)**3+ XK(7)*U(N)**2+ & XK(7)*XK(8)*U(N)+XK(9)*XK(7)*XK(8))**2 DF1(25,5)=XK(7)*XK(8)*(-2.*U(N)**3-XK(7)*U(N)**2+ &XK(9)*XK(7)*XK(8))/(U(N)**3+ XK(7)*U(N)**2+ & XK(7)*XK(8)*U(N)+XK(9)*XK(7)*XK(8))**2 DF1(25,6)=XK(7)*XK(8)*XK(9)*(3.*U(N)**2+ &2.*XK(7)*U(N)+XK(7)*XK(8))/(U(N)**3+ XK(7)*U(N)**2+ &XK(7)*XK(8)*U(N)+XK(9)*XK(7)*XK(8))**2 RETURN END ******************************************************************** SUBROUTINE CARGAS(NC,C1) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION C1(30) C1(1)=-2. C1(2)=-1. C1(3)=0. C1(4)=1. C1(5)=2. C1(NC)=3. RETURN END ******************************************************************** SUBROUTINE PRODUC(N,M,TEMP,Y,V) Optimización de la digestión anaerobia de purines de cerdo 357 IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) COMMON/GROUP2/U0,V0,VG DIMENSION Y(40,40),U0(40) DO I=1,N-1 DO J=1,M-1 Y(I,J)=0. ENDDO ENDDO CGL=(V/VG)*0.0821*(273+TEMP) Y(23,12)=1. Y(32,12)=-CGL Y(22,13)=1. Y(31,13)=-CGL Y(17,14)=1. Y(33,14)=-CGL Y(34,15)=1. Y(22,15)=-1. Y(29,15)=-1. RETURN END ******************************************************************** SUBROUTINE TASAS(N,M,TEMP,U,R) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION U(40),R(40) DIMENSION XK(20) DIMENSION F1(40,40),DF1(40,40) CALL CONSTANT(TEMP,XK) CALL MATRIZF(N,NC,U,XK,F1,DF1) XNH3=U(17)*F1(17,3) XKLACO2=55. XKLACH4=55. XKLANH3=1. XKHCO2=0.0697-0.002*TEMP+2.56E-5*TEMP**2-1.21E-7*TEMP**3 XKHCH4=EXP(-60.954+85.8073/((273.15+TEMP)/100.)+ & 23.4487*DLOG((273.15+TEMP)/100.)) XKHNH3=52.9-1.454*TEMP+0.021*TEMP**2-1.13E-4*TEMP**3 R(19)=XKLACO2*(XKHCO2*U(30)-U(22)*F1(22,3)) R(20)=XKLACH4*(XKHCH4*U(31)-U(23)) R(M-1)=XKLANH3*(XKHNH3*U(32)-XNH3) 358 Anejo 2 RETURN END ******************************************************************** SUBROUTINE CONSTANT(TEMP,XK) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION C(20,20),PK(20),XK(20) *1: PROPIONICO;2:BUTÍRICO; 3:ACÉTICO; ; 4 Y 5: 1º Y 2º¦ CARBÓNICO,6: AMONIO, 7,8,Y 9, 1º,2ºY3º FOSFORICO,10: AGUA NC=11 C(3,1)=4.7803 C(3,2)= -0.0023 C(3,3)= 6.E-5 C(3,4)=-2.E-7 C(2,1)=4.8947 C(2,2)= -0.0024 C(2,3)= 7.E-5 C(2,4)=-3.E-7 C(1,1)=4.8063 C(1,2)= -0.0011 C(1,3)= 8.E-5 C(1,4)=-4.E-7 C(4,1)=6.539 C(4,2)= -0.010 C(4,3)= 2.56E-5 C(4,4)=-1.21E-7 C(5,1)=10.619 C(5,2)= -0.014 C(5,3)= 1.01E-4 C(5,4)=0. C(7,1)=2.0473 C(7,2)=0.0019 C(7,3)=5.E-5 C(7,4)=0. C(8,1)=7.3144 C(8,2)=-0.0073 C(8,3)=0.0001 C(8,4)=-6.E-7 C(9,1)=12.6576 C(9,2)=0. Optimización de la digestión anaerobia de purines de cerdo 359 C(9,3)=0. C(9,4)=0. C(6,1)=10.072 C(6,2)=-0.0356 C(6,3)=9.E-5 C(6,4)=4.E-8 C(10,1)=14.934 C(10,2)=-0.0425 C(10,3)=0.0002 C(10,4)=-6.E-7 DO I=1,10 C(10,5)=0. ENDDO DO I=1,NC-1 PK(I)=0. ENDDO DO I=1,NC-1 DO J=1,5 PK(I)=PK(I)+C(I,J)*TEMP**(J-1) ENDDO XK(I)=10**(-PK(I)) ENDDO C(11,1)=0.0134 C(11,2)=-0.0004 C(11,3)=8.E-6 C(11,4)=-7.E-8 C(11,5)=3.E-10 XK(11)=0. DO J=1,5 XK(11)=XK(11)+C(11,J)*TEMP**(J-1) ENDDO PK(12)=PK(1) XK(12)=XK(1) RETURN END ******************************************************************** SUBROUTINE SECANTE(TEMP,U,X) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION U(30) 360 Anejo 2 TOL=1.E-4 PX=-DLOG10(X) I=1 F=1.0001 5 PX1=PX*F PX2=PX/F 10 CALL FUNCH(TEMP,U,PX1,F1) CALL FUNCH(TEMP,U,PX2,F2) IF((F1*F2).GT.0.) THEN F=F*1.1 GOTO 5 ENDIF PX3=PX2-(PX1-PX2)/(F1/F2-1.) CALL FUNCH(TEMP,U,PX3,F3) IF((F1*F3).GT.0.)THEN PX1=PX3 ELSE PX2=PX3 ENDIF ERR1=DABS(PX2-PX1) ERR2=DABS(F3) ERR=MAX(ERR1,ERR2) ERR0=MIN(ERR1,ERR2) I=I+1 IF(ERR.GT.TOL) THEN IF (ERR0.LT.TOL/1.E2) GO TO 20 GO TO 10 ENDIF 20 PX=PX3 X=10**(-PX) RETURN END ******************************************************************** SUBROUTINE FUNCH(TEMP,U,PH,FHH) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION U(40),A(40),XK(20) CALL CONSTANT(TEMP,XK) HH=10**(-PH) CALL COEF1(HH,XK,A) Optimización de la digestión anaerobia de purines de cerdo 361 CH=0. DO I=18,31 CH=CH+A(I)*U(I) ENDDO IF (CH.GE.0.) THEN PH1=(-DLOG10((CH+DSQRT(CH**2+4.*XK(10)))/2.)) ELSE PH1=(-DLOG10(2.*XK(10)/(-CH+DSQRT(CH**2+4.*XK(10))))) ENDIF FHH=PH-PH1 RETURN END ******************************************************************** SUBROUTINE COEF1(HH,XK,A) IMPLICIT DOUBLE PRECISION(A-H,O-Z),INTEGER(I-N) DIMENSION A(40),XK(20) DO I=17,29 A(I)=0. ENDDO A(20)=XK(1)/(HH+XK(1)) A(19)=XK(2)/(HH+XK(2)) A(21)=XK(3)/(HH+XK(3)) A(18)=XK(12)/(HH+XK(12)) A(23)=0. A(22)=(XK(4)*HH+2.*XK(4)*XK(5))/(HH**2+XK(4)*HH+XK(4)*XK(5)) A(17)=-HH/(XK(6)+HH) A(25)=(XK(7)*HH**2+2.*XK(7)*XK(8)*HH+3.*XK(7)*XK(8)*XK(9))/ & (HH**3+XK(7)*HH**2+XK(7)*XK(8)*HH+XK(7)*XK(8)*XK(9)) A(26)=1. A(27)=2. A(28)=-1. A(29)=-2. RETURN END 11. ÍNDICES DE TABLAS Y FIGURAS Optimización de la digestión anaerobia de purines de cerdo 363 364 Índices de tablas y figuras ÍNDICE DE TABLAS Tabla 1.1. Generación de residuos en España en el año 1996 ................................................ 4 Tabla 1.2. Generación de residuos e incidencia de la recogida selectiva en el año 1998, en Cataluña .................................................................................................................................. 5 Tabla 1.3. Destino final de lodos de EDAR en 1998........................................................... 6 Tabla 1.4. Distribución de la contribución de los distintos sectores de actividad al Valor Añadido Bruto en Cataluña (1995).......................................................................................... 7 Tabla 1.5. Censo de ganado según la encuesta de diciembre de 1998 ....................................... 9 Tabla 1.6. Estimación de la producción anual de purín, basada en el censo de porcino del año 98 (DARP, 1999) y las equivalencias propuestas por Danés et al. (1996). ............................. 11 Tabla 1.7. Síntesis de operaciones aplicables al tratamiento de residuos ganaderos, en especial a purines de cerdo ....................................................................................................................... 15 Tabla 1.8. Ventajas del proceso de digestión anaerobia ........................................................ 16 Tabla 2.1. Componentes del biogás en función del substrato utilizado.................................... 25 Tabla 2.2. Diferentes cinéticas de crecimiento de microorganismos usadas en la bibliografía ... 28 Tabla 2.3. Tipos de inhibición y expresión matemática de la cinética ..................................... 29 Tabla 2.4. Comparación de las diferentes cinéticas utilizadas en la simulación de la fase hidrolítica en modelos de digestión anaerobia de substratos complejos ......................................... 35 Tabla 2.5. Resumen de parámetros cinéticos para las diferentes fases y diferentes grupos en el proceso de digestión anaerobia ................................................................................................. 41 Tabla 2.6. Reacciones acetogénicas que ocurren en los sistemas anaerobios ............................. 43 Tabla 2.7. Principales reacciones metanogénicas y otras consumidoras de hidrógeno................. 46 Tabla 2.8. Resumen de parámetros cinéticos de la fase metanogénica...................................... 47 Tabla 2.9. Resumen de constantes cinéticas propuestas para los diferentes procesos ................ 48 Tabla 2.10. Temperatura óptima y máxima, parámetros cinéticos de crecimiento para diferentes cultivos metanogénicos acetoclásticos .......................................................................................... 51 Tabla 2.11. Rangos de concentración de nutrientes, necesarios para el correcto crecimiento de las bacterias anaerobias................................................................................................................. 52 Tabla 2.12. Concentración de inhibición y toxicidad de metales pesados ................................ 62 Tabla 2.13. Concentración límite de cationes en sistemas anaerobios...................................... 62 Tabla 4.1. Resumen de ecuaciones de simulación del pH en modelos estructurados de digestión anaerobia................................................................................................................................ 86 Tabla 4.2. Coeficientes para las expresiones de constantes de disociación ................................ 93 Tabla 4.3. Matriz F .......................................................................................................... 95 Tabla 4.4. Coeficientes para las expresiones de la constante de Henry.................................... 96 Tabla 4.5. Matriz F’ ......................................................................................................... 99 Tabla 4.6. Relación de procesos considerados ...................................................................... 100 Tabla 4.7. Relación de componente consideradas................................................................. 101 Tabla 4.8. Reacciones biológicas consideradas ..................................................................... 103 Optimización de la digestión anaerobia de purines de cerdo 365 Tabla 4.9. Matriz de coeficientes estequiométricos............................................................... 105 Tabla 4.10. Cinéticas de reacción de los procesos biológicos y físico-químicos considerados ..... 107 Tabla 4.11. Valores de los parámetros cinéticos utilizados en el modelo ............................. 108 Tabla 4.12. Componentes utilizadas para los diferentes experimentos realizados ................ 113 Tabla 4.13. Composición de los diferentes substratos utilizados en las simulaciones.............. 116 Tabla 4.14. Resultados obtenidos mediante el modelo de simulación, para diferentes tipos de cosubstratos en el rango mesofílico (35ºC) ............................................................................... 116 Tabla 5.1. Diseño de los experimentos. (5 repeticiones por cada tratamiento)....................... 131 Tabla 5.2. Factores para la conversión de la concentración de AGV a unidades de DQO... 138 Tabla 5.3. Caracterización físico química de los substratos utilizados*................................. 140 Tabla 5.4. Análisis de cationes y aniones por electroforesis capilar (unidades mg/L)* .......... 141 Tabla 5.5. Concentración de nitrógeno Kjeldahl (mg N/Kg) en el experimento 1 ................. 143 Tabla 5.6. Concentración de nitrógeno amoniacal (g N-NH4+ /L) en el experimento 1. ..... 144 Tabla 5.7. Proporción de nitrógeno amoniacal sobre nitrógeno total. Incremento sufrido durante la digestión en el experimento 1.............................................................................................. 145 Tabla 5.8. Concentraciones medias y eliminación de sólidos totales en el experimento 1. ....... 147 Tabla 5.9. Concentraciones medias y eliminación de sólidos volátiles en el experimento 1...... 147 Tabla 5.10. Valores medios y eliminación de DQO en el experimento 1............................. 148 Tabla 5.11. Valores medios iniciales de pH y alcalinidad en el experimento 1 .................... 149 Tabla 5.12. Valores medios de pH al inicio y al final de la digestión, y concentración media de amoníaco libre durante el proceso, en el experimento 1............................................................ 149 Tabla 5.13. Valores medios de la producción acumulada de gas (M) al final del proceso, para las variables biogás, metano y dióxido de carbono, en el experimento 1..................................... 150 Tabla 5.14. Valores medios de los índices de producción acumulada de gas respecto al contenido inicial de sólidos volátiles (B), para las variables biogás, metano y dióxido de carbono, en el experimento 1. ...................................................................................................................... 151 Tabla 5.15. Valores medios de parámetros estimados mediante la ecuación de Gompertz (ecuaciones 5.5 y 5.6), en el experimento 1. ............................................................................ 155 Tabla 5.16. Indice de metanización, acidificación y biodegradabilidad en el experimento 1. .. 156 Tabla 5.17. Diferencias entre los valores de AGV total en el día 6 (máximo mesofílico), 13 (máximo termofílico), y final (valores en mM), en el experimento 1 ......................................... 159 Tabla 5.18. Concentración de compuestos nitrogenados en el experimento 2. ........................ 168 Tabla 5.19. Proporción de nitrógeno amoniacal sobre nitrógeno total. Incremento sufrido durante la digestión, en el experimento 2............................................................................................. 169 Tabla 5.20. Concentraciones medias y eliminación de sólidos totales en el experimento 2 ...... 170 Tabla 5.21. Concentraciones medias y eliminación de sólidos volátiles en el experimento 2.... 170 Tabla 5.22. Concentraciones medias y eliminación de DQO en el experimento 2................. 170 Tabla 5.23. Contenido en fibra y en el substrato inicial del experimento 2........................... 172 Tabla 5.24. Alcalinidad total, alcalinidad al bicarbonato y relación de alcalinidad, en el 366 Índices de tablas y figuras experimento 2. ...................................................................................................................... 172 Tabla 5.25. Valores medios de pH al inicio y al final de la digestión en el experimento 2.... 173 Tabla 5.26. Concentración de amoníaco libre en el experimento 2. ...................................... 173 Tabla 5.27. Valores medios de producción acumulada de gas (M) al final del proceso, para las variables biogás, CH4 y CO2, en el experimento 2.................................................................. 174 Tabla 5.28. Valores medios del índice de producción acumulada de gas respecto a sólidos volátiles iniciales (B) al final del proceso, para las variables biogás, metano y CO2, en el experimento 2. ...................................................................................................................... 175 Tabla 5.29. Parámetros estimados mediante la ecuación de Gompertz en el experimento 2... 178 Tabla 5.30. Indice de metanización, acidificación y biodegradabilidad en el experimento 2. .. 181 Tabla 5.31. Valores de concentración de Ácidos grasos Volátiles (mM) obtenidos en diferentes momentos de la digestión, en el experimento 2. ........................................................................ 182 Tabla 5.32. Concentración de nitrógeno Kjeldahl en el experimento 3.................................. 192 Tabla 5.33. Concentración de nitrógeno amoniacal en el experimento 3. .............................. 192 Tabla 5.34. Proporción de nitrógeno amoniacal sobre nitrógeno total. Incremento sufrido durante la digestión, en el experimento 3............................................................................................. 192 Tabla 5.35. Concentraciones medias y eliminación de sólidos totales, en el experimento 3. .... 193 Tabla 5.36. Conc. medias y eliminación de sólidos volátiles en el experimento 3................... 194 Tabla 5.37. Concentraciones medias y eliminación de DQO, en el experimento 3................ 194 Tabla 5.38. Comparación de la alcalinidad total y al bicarbonato (pH 5,75) inicial, en el experimento 3 ....................................................................................................................... 195 Tabla 5.39. Valores medios de pH y de amoníaco libre, en el experimento 3....................... 196 Tabla 5.40. Producción acumulada de gas (M), para las variables biogás, CH4 y CO2 en el experimento 3. ...................................................................................................................... 197 Tabla 5.41. Valores medios de los índices de producción acumulada de gas respecto al contenido inicial de sólidos volátiles (B), para las variables biogás, metano y dióxido de carbono, en el experimento 3. ...................................................................................................................... 198 Tabla 5.42. Cálculo de parámetros mediante la ecuación de Gompertz en el experimento 3.. 200 Tabla 5.43. Índices de metanización, acidificación y biodegradabilidad en el experimento 3 .. 201 Tabla 5.44. Valores de concentración de ácidos grasos volátiles (mM) obtenidos en diferentes momentos de la digestión, en el experimento 3 ......................................................................... 203 Tabla 6.1. Revisión bibliográfica de producción de metano sobre sólidos volátiles (Pc), en L CH4/g SV, de residuos ganaderos en función de la temperatura (Tª), velocidad de carga orgánica (VCO)en g SV/L·d, tiempo de retención (TR), en días y nitrógeno amoniacal, en g N/L..... 221 Tabla 6.2. Cronología de los cuatro reactores continuos utilizados ....................................... 232 Tabla 6.3. Programación de los experimentos ..................................................................... 234 Tabla 6.4. Caracterización del substrato utilizado durante la fase de puesta en marcha. ...... 235 Tabla 6.5. Caracterización de la alimentación en los diferentes períodos de experimentación . 235 Tabla 6.6. Eliminación de materia orgánica durante el período de puesta en marcha en los Optimización de la digestión anaerobia de purines de cerdo 367 reactores 1 y 2....................................................................................................................... 239 Tabla 6.7. Eliminación de materia orgánica durante el período de puesta en marcha en los reactores 3 y 4....................................................................................................................... 244 Tabla 6.8. Producción de biogás y de metano media de los 16 últimos días del período de puesta en marcha para los cuatro reactores. ....................................................................................... 246 Tabla 6.9. Eliminación de parámetros de materia orgánica en el reactor 1, calculada por períodos................................................................................................................................. 256 Tabla 6.10. Eliminación de parámetros de materia orgánica en el reactor 2, calculada por períodos................................................................................................................................. 263 Tabla 6.11. Producción de metano media para los últimos días de cada período en el reactor 3. ............................................................................................................................................ 265 Tabla 6.12. Eliminación de materia orgánica en el reactor 3, calculada por períodos............ 272 Tabla 6.13. Eliminación de materia orgánica en el reactor 4 calculada por períodos............. 278 Tabla 6.14. Producción volumétrica de metano (L CH4/L·d) y producción por unidad de carga (LCH4/g SVin). Medias para cada período, y cada reactor................................................... 284 Tabla 6.15. Fracción de nitrógeno amoniacal sobre el Kjeldahl, e incremento de dicha fracción durante la digestión. Valores medios de todo el período de ensayo............................................. 289 368 Índices de tablas y figuras ÍNDICE DE FIGURAS Figura 1.1. Distribución de los cerdos de engorde en Europa ................................................... 8 Figura 1.2. Principales productos de la producción final agraria ............................................. 9 Figura 2.1. Ilustración de la cinética de crecimiento de Monod............................................... 28 Figura 2.2. Efecto de la concentración de substrato sobre la relación tasa específica de crecimiento/tasa máxima, para diferentes valores de n .............................................................. 30 Figura 2.3. Esquema de reacciones de la digestión anaerobia de materiales poliméricos.. ......... 32 Figura 2.4. Simplificación de las rutas metabólicas de degradación de la glucosa por las bacterias acidogénicas............................................................................................................................. 38 Figura 2.5. Esquema de dependencia de la energía libre en función de la presión parcial de hidrógeno ................................................................................................................................ 44 Figura 2.6. Dependencia de la constante de crecimiento de la temperatura ............................. 49 Figura 3.1. Ejemplo de cromatograma del análisis de AGV................................................ 73 Figura 3.2. Ejemplo de rectas de calibración para los diferentes AGV.................................. 75 Figura 3.3. Rectas de calibración para los tres componentes del biogás considerados ................ 77 Figura 4.1. Diagrama de flujo del modelo general de digestión anaerobia ............................. 102 Figura 4.2. Esquema de sistemas continuos y discontinuos .................................................. 108 Figura 4.3. Esquema del dispositivo experimental utilizado para la validación del algoritmo de simulación dinámica del pH .................................................................................................. 111 Figura 4.4. Comparación de los valores de pH medidos y simulados, correspondientes a las condiciones de la Tabla 4.12 y a los momentos de introducción indicados de cada componente.... 114 Figura 4.5. Simulación del proceso con dos tipos de purín en el rango termofílico: el purín 2 con mayor concentración de amonio que el purín 1, utilizado hasta el día 30.................................. 118 Figura 4.6. Evolución de AGV y producción de biogás, según la forma de introducción de TDO como cosubstrato.................................................................................................................... 119 Figura 5.1. Burbujeo de viales antes de comenzar la digestión.............................................. 132 Figura 5.2. Detalle del interior de uno de los incubadores.................................................... 133 Figura 5.3. Elementos de muestreo. ................................................................................... 133 Figura 5.4. Comparación de la concentración de amonio antes y después de la digestión en el experimento 1. ...................................................................................................................... 145 Figura 5.5. Evolución de la producción de metano acumulado en el experimento 1 .............. 152 Figura 5.6. Evolución de la producción acumulada de CO2, en el experimento 1. ............... 153 Figura 5.7. Evolución temporal de el índice de producción acumulada respecto a sólidos volátiles iniciales, en el experimento 1.................................................................................................. 154 Figura 5.8. Evolución de la concentración total de Ácidos Grasos Volátiles (C2-C5), en el experimento 1 ....................................................................................................................... 158 Figura 5.9. Evolución del ácido acético y propiónico en el experimento 1.............................. 160 Optimización de la digestión anaerobia de purines de cerdo 369 Figura 5.10. Evolución de los dos isómeros del ácido valérico y butírico, en el experimento 1. 163 Figura 5.11. Correlación valores experimentales de producción de metano respecto a sólidos volátiles (B) y concentración media de amoníaco libre (Experimento 1) .................................... 165 Figura 5.12. Comparación de la producción potencial de metano respecto a sólidos volátiles iniciales y la velocidad de producción de metano con la concentración de amoníaco libre.............. 166 Figura 5.13. Valores de DQO inicial y final y eliminación en el experimento 2.................. 171 Figura 5.14. Producción acumulada de CH4 en el experimento 2........................................ 175 Figura 5.15. Evolución de la producción acumulada de CO2, en el experimento 2 ............... 176 Figura 5.16. Comparación de los datos experimentales media de las cinco repeticiones (puntos) y la curva de la simulación del modelo Gompertz, en el experimento 2, para el índice producción de metano respecto a sólidos volátiles iniciales .............................................................................. 177 Figura 5.17. Evolución de los Ácidos Grasos Volátiles totales en el experimento 2 ............. 181 Figura 5.18. Evolución de la concentración de acético y propiónico, en el experimento 2........ 183 Figura 5.19. Evolución de los isómeros de butírico y valérico en el experimento 2 ................. 186 Figura 5.20. Eliminación de DQO para los diferentes tratamientos, en el experimento 3. ... 195 Figura 5.21. Evolución de la producción de metano acumulada, en el experimento 3............ 197 Figura 5.22. Evolución de la producción de CO2 acumulada en el experimento 3. ............... 198 Figura 5.23. Comparación de los datos experimentales media de las cinco repeticiones (puntos) y la curva de la simulación del modelo Gompertz, en el experimento 3, para el índice producción de metano respecto a sólidos volátiles iniciales .............................................................................. 200 Figura 5.24. Evolución de la concentración total de AGV en el experimento 3.................. 202 Figura 5.25. Evolución del acético y propiónico, en el experimento 3 ................................... 204 Figura 5.26. Evolución de los isómeros de butírico y valérico, en el experimento 3. ............... 205 Figura 6.1. Esquema de cada una de las cuatro unidades del banco de ensayos en continuo. . 224 Figura 6.2. Vista de la botella de influente y la bomba de impulsión................................... 225 Figura 6.3. Vista detalle de depósito de efluente- desgasificador ........................................... 226 Figura 6.4. Esquema de funcionamiento del contador de gas. .............................................. 226 Figura 6.5. Vista del contador de gas ................................................................................ 227 Figura 6.6. Esquema y fotografía del reactor ...................................................................... 228 Figura 6.7. Esquema del sistema de calefacción. ................................................................. 228 Figura 6.8. Evolución de la composición de la alimentación de los reactores 3 y 4, durante la puesta en marcha................................................................................................................... 233 Figura 6.9. Evolución de la carga orgánica, concentración de SV, datos individuales y medias por períodos, y tiempo de retención medio (TR), para los cuatro reactores.................................. 236 Figura 6.10. Evolución del nitrógeno amoniacal y amoníaco libre, del influente y del efluente, durante la fase de puesta en marcha, para los reactores 1 y 2................................................... 237 Figura 6.11. Evolución de los SV del influente y del efluente, durante la fase de puesta en marcha, para los reactores 1 y 2............................................................................................. 238 Figura 6.12. Evolución de la DQO del influente y del efluente, durante la fase de puesta en 370 Índices de tablas y figuras marcha, para los reactores 1 y 2............................................................................................. 238 Figura 6.13. Evolución de la producción de metano, Pv y Pc, durante la fase de puesta en marcha, para los reactores 1 y 2............................................................................................. 240 Figura 6.14. Evolución de la relación de alcalinidad (RA), durante la fase de puesta en marcha, para los reactores 1 y 2.......................................................................................................... 240 Figura 6.15. Evolución de la alcalinidad total (AT) y parcial (AP), durante la fase de puesta en marcha, para los reactores 1 y 2......................................................................................... 241 Figura 6.16. Evolución del nitrógeno amoniacal y amoníaco libre, del influente y del efluente, durante la fase de puesta en marcha, para los reactores 3 y 4................................................... 242 Figura 6.17. Evolución de SV del influente y del efluente, durante la fase de puesta en marcha, para los reactores 3 y 4.......................................................................................................... 243 Figura 6.18. Evolución de DQO del influente y del efluente, durante la fase de puesta en marcha, para los reactores 3 y 4............................................................................................. 243 Figura 6.19. Evolución de la producción de metano, Pv y Pc, durante la fase de puesta en marcha, para los reactores 3 y 4............................................................................................. 244 Figura 6.20. Evolución de la alcalinidad total (AT) y parcial (AP), durante la fase de puesta en marcha, para los reactores 3 y 4......................................................................................... 245 Figura 6.21. Evolución de la relación de alcalinidades (RA), durante la fase de puesta en marcha, para los reactores 3 y 4............................................................................................. 246 Figura 6.22. Producción volumétrica de metano, para los cuatro reactores. ........................... 247 Figura 6.23. Producción de metano por unidad de carga (g SV), para los cuatro reactores.... 248 Figura 6.24. Evolución de la producción volumétrica de biogás (Pv), la producción por unidad de carga (Pc) y la concentración de metano en el biogás (%) en el reactor 1.................................... 250 Figura 6.25. Evolución de Nitrógeno Kjeldahl (Nk ) y amoniacal (NH4) del influente y efluente en el reactor 1. Concentración de amoníaco libre en el efluente...................................... 251 Figura 6.26. Evolución del pH del efluente del reactor 1..................................................... 251 Figura 6.27. Evolución de la alcalinidad total (AT), parcial o al bicarbonato (AP) y relación de alcalinidad (RA) en el reactor 1 ........................................................................................ 252 Figura 6.28. Evolución de la concentración de AGV en el efluente del reactor 1.................. 253 Figura 6.29. Evolución de la DQO total y soluble en el influente y efluente del reactor 1. .... 254 Figura 6.30. Evolución de ST y SV en el influente y efluente del reactor 1.......................... 255 Figura 6.31. Evolución de los índices de metanización y de acidificación en el reaactor 1, expresaada en % de DQO transformada. ............................................................................. 256 Figura 6.32. Evolución de la producción volumétrica de biogás (Pv), la producción por unidad de carga (Pc), concentración de metano en el biogás (%) en el reactor 2. . ...................................... 257 Figura 6.33. Evolución de Nitrógeno Kjeldahl (Nk ) y amoniacal (NH4) del influente y efluente y concentración de amoníaco libre en el efluente, en el reactor 2..................................... 258 Figura 6.34. Evolución del pH del efluente del reactor 2..................................................... 259 Figura 6.35. Concentración de AGV en el efluente del reactor 2. ....................................... 260 Optimización de la digestión anaerobia de purines de cerdo 371 Figura 6.36. Evolución de DQO total y soluble en el influente y efluente del reactor 2. ........ 262 Figura 6.37. Evolución de ST y SV en el influente y efluente del reactor 2.......................... 262 Figura 6.38. Evolución de los índices de metanización y de acidificación en el reactor 2, en % de la DQO inicial..................................................................................................................... 263 Figura 6.39. Evolución de la producción volumétrica de biogás (Pv); de la producción por unidad de carga (Pc) y concentración de metano en el biogás en el reactor 3. ......................................... 264 Figura 6.40. Evolución de Nitrógeno Kjeldahl y amoniacal del influente y efluente del reactor 3. Concentración de amoníaco libre en el efluente......................................................................... 266 Figura 6.41. Evolución del pH del efluente del reactor 3..................................................... 267 Figura 6.42. Evolución de la concentración de AGV en el reactor 3................................... 268 Figura 6.43. Evolución de ST y SV del influente y efluente del reactor 3. ........................... 271 Figura 6.44. Evolución de la DQO total y soluble en el influente y efluente del reactor 3. ... 271 Figura 6.45. Evolución índices de acidificación y metanización para el reactor 3. ................. 272 Figura 6.46. Evolución de la producción volumétrica de biogás (Pv), producción por unidad de carga y concentración de metano en el biogás, en el reactor 4.................................................... 274 Figura 6.47. Evolución de nitrógeno Kjeldahl y amoniacal del influente y efluente en el reactor 4. Concentración de amoníaco libre en el efluente..................................................................... 275 Figura 6.48. Evolución del pH del efluente del reactor 4..................................................... 275 Figura 6.49. Evolución de la concentración de AGV en el efluente del reactor 4.................. 276 Figura 6.50. Evolución de la DQO total y soluble del influente y el efluente, reactor 4......... 278 Figura 6.51. Evolución de los ST y SV del influente y efluente del reactor 4. ...................... 279 Figura 6.52. Evolución de los índices de acidificación y metanización en el reactor 4. ........... 280 Figura 6.53. Estudio de regresión de la concentración de acético y la producción volumétrica de metano con la concentración de nitrógeno amoniacal ................................................................ 282 Figura 6.54. Correlación de la producción volumétrica de biogás y de la producción de metano por unidad de carga respecto a la velocidad de carga orgánica. .................................................. 288 La digestión anaerobia de purines de cerdo puede ser una buena opción para la estabilización y valorización económica de estos residuos. Una clara opción para mejorar la producción de metano, y por tanto la viabilidad económica, es la digestión conjunta con residuos orgánicos de la industria agroalimentaria. De los resultados obtenidos en el presente trabajo se puede concluir que la codigestión de purín de cerdo con residuos del procesado de pera, o con residuos procedentes del refinado de aceite de oliva, tierras decolorantes, es una opción interesante para incrementar las producciones de biogas, posibilitando, a su vez, el tratamiento de dichos residuos industriales. Se han desarrollado mejoras en la modelización del proceso, lo cual ha permitido obtener una herramienta útil para la predecir el comportamiento del sistema. Anaerobic digestion of pig slurry can be a good option to valorise and stabilise this waste. Co-digestion with organic wastes from food industry is a clear option to improve methane production, and, therefore, the economic feasibility of anaerobic digestion. From the results of this work, it can be concluded that codigestion with pearjuice wastes, or wastes from oil refineries (olive oil bleaching earth), is an interesting option to increase biogas production, improving the treatment feasibility of some kinds of industrial wastes. Improvements in the modelling of the process have been developed, which have allowed to obtain a useful tool for predicting the system behaviour.