Metabolismo del glucógeno
Anuncio

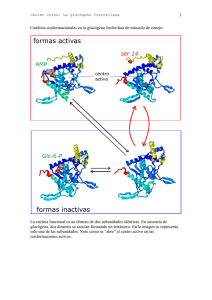
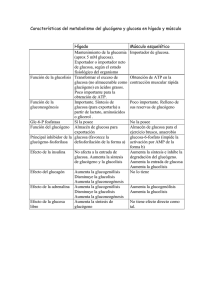
Metabolismo del glucógeno • El glucógeno representa la principal forma de almacenamiento de carbohidratos en animales. Lugares de almacenamiento El g glucógeno g se encuentra en el hígado g y músculo. • El glucógeno hepático sirve en gran parte para exportar unidades de hexosa para la conservación de la glucosa sanguínea, en particular entre comidas. Después D é de d 12 a 18 horas h d ayuno, ell hígado de hí d casii agota su reserva de glucógeno. La liberación del glucógeno en el hígado es desencadenada por niveles bajo de glucosa en sangre. • La función del glucógeno muscular es actuar como una fuente de fácil disponibilidad de unidades de hexosa para la glucólisis dentro del propio músculo. En el músculo, la glucosa-6-fosfato obtenida de la descomposición del glucógeno entra en la vía glucolítica directamente en vez de ser hidrolizada a glucosa y directamente, después exportada a la sangre. El glucógeno muscular sólo disminuye de manera significativa después de ejercicio vigoroso prolongado. Puede P ede inducirse ind cirse un n almacenaje mayor ma or de glucógeno gl cógeno muscular con dietas ricas en carbohidratos después de la depleción por el ejercicio. Descomposición del glucógeno. • Cuando existe una disminución significativa de glucosa en sangre, sangre el glucógeno es degradado por medio de una serie de enzimas para cubrir las necesidades energéticas de nuestro organismo. • Este proceso es llamado glucogenólisis. • Primero: rupturas p fosforolíticas secuenciales de los enlaces α(1-4) La glucógeno fosforilasa rompe los enlaces glucosídicos α(1-4) entre los residuos que están en los extremos no reductores por simple fosforólisis. fosforólisis La glucógeno fosforilasa es una fosfotransferasa que degrada secuencialmente las cadenas de glucógeno en sus extremos no reductores hasta q que están a ólo cuatro residuos de glucosa en la ramificación. La estructura se denomina dextrina límite y la fosforilasa no puede degradarla g más. • Segundo: las ramas del glucógeno son removidas por medio de una segunda enzima, la (α1,4 →α1,4) glucantransferasa quien cataliza dos reacciones. La primera reacción, elimina tres de los residuos de glucosa restantes y transfiere este trisacárido intacto al extremo de alguna otra ramificación externa. externa A continuación el residuo restante de glucosa unido a la cadena en posición α(1-6), es removido hidrolíticamente liberando glucosa. glucosa Ambas actividades se encuentran en sitios separados de la misma cadena polipeptídica (enzima desramificante) • Tercero: la g glucosa-1-fosfato p producida p por la g glucógeno g fosforilasa es convertida en glucosa-6-fosfato por la fosfoglucomutasa. La reacción produce glucosa 1,6 bisfosfato como un intermediario temporal pero esencial. esencial enzima-fosfato + glucosa-1-fosfato ↔ enzima + glucosa-1,6-bisfosfato glucosa-1,6-bisfosfato + enzima↔enzima-fosfato +glucosa-6-fosfato glucosa-1-fosfato ↔ glucosa-6-fosfato • Finalmente: Esta glucosa fosforilada, para salir de las células hepáticas, debe ser hidrolizada a glucosa y ortofosfato mediante di t la l enzima i glucosa-6-fosfatasa. l 6f f t Glucosa-6-P + H2O ↔ Glucosa + Pi En cambio el g glucógeno g muscular,, se utiliza principalmente como fuente de glucosa-6-fosfato para el catabolismo en las células musculares.(no hay glucosa6-fosfatasa) Síntesis del glucógeno • La síntesis de g glucógeno g a p partir de g glucosa se llama glucogénesis y se produce gracias a la enzima glucógeno sintetasa. • La adición de una molécula de glucosa al glucógeno consume dos enlaces de alta energía: una procedente del ATP y otra que procede del UTP • Ocurre principalmente en el músculo y en el hígado • Primero: la glucosa es transformada en glucosa glucosa-66 fosfato, gastando una molécula de ATP. glucosa + ATP → glucosa-6-P glucosa 6 P + ADP La reacción es catalizada p por la enzima g glucoquinasa q en el hígado y por la enzima hexoquinasa en el músculo. • Segundo: a continuación se transforma la glucosa-6glucosa 6 fosfato en glucosa-1-fosfato glucosa-6-P ←→ glucosa-1-P La reacción es fosfoglucomutasa. catalizada por la enzima • Tercero: la glucosa glucosa-1-fosfato 1 fosfato reacciona con UTP, UTP para producir uridina difosfato glucosa (UDP-glucosa) y pirofosfato (PPi). glucosa-1-P + UTP UDP-Glucosa pirofosforilasa → UDP-glucosa + PPi • Cuarto: la enzima glucógeno sintetasa va uniendo UDP UDPglucosa a través de enlaces glicosídicos α (1-4), para formar el glucógeno. ( l (glucosa)n ) + UDP UDP-glucosa l → ((glucosa)n+1 l ) 1 + UDP • Finalmente: la enzima de ramificación transfiere un segmento de siete residuos de largo de una cadena en crecimiento, a un nuevo punto de ramificación (generalmente a cuatro residuos de otra ramificación)) a (g través de un enlace glicosídico α (1-6). Importante La enzima glucógeno sintasa, no puede formar un enlace entre dos moléculas aisladas de glucosa. Es decir debe agregarse a una cadena ya existente con enlaces α (1-4). Para lograr entonces comenzar la síntesis se necesita un cebador. En este caso el grupo hidroxilo de una tirosina específica de la glucogenina g cumple este fin. proteína g La síntesis comienza enlazando un residuo de glucosa con el hidróxilo de la tirosina y luego los otros residuos se agregan en forma sucesiva al primero. primero La propia molécula de glucogenina actúa como catalizador, hasta la unión de ocho moléculas de glucosa. Luego comienza a funcionar la glucógeno sintasa. → Una proteína glicogenina es la que une la primera molécula de glucosa. → La enzima glicógeno sintasa forma un complejo con la glicogenina. → Este complejo comienza a alargar la cadena al unirse moléculas de UDP-glucosa. Control del metabolismo del glucógeno Debido a la importancia de mantener los niveles de glucosa l sanguínea, í l síntesis la í t i y degradación d d ió d l del glucógeno están muy reguladas. En el hígado la síntesis de glucógeno se acelera durante periodos en los que el individuo está bien alimentado, por tanto la degradación del glucógeno se acelera en periodos de ayuno. ayuno En el músculo la degradación del glucógeno ocurre d durante t ell ejercicio j i i activo ti y la l acumulación l ió comienza i en cuanto el músculo entra en reposo. La regulación g de la síntesis degradación ocurre a dos niveles: y 1.- la glucógeno sintasa y glucógeno fosforilasa son controladas alostericamente. 2.- control hormonal. 1.- La regulación a través de la glucógeno sintetasa i t t y la l glucógeno l ó f f il fosforilasa La glucógeno L l ó sintetasa i t t ( ti i (participante t de d la l síntesis) í t i ) tiene ti dos formas: glucógeno sintetasa I (independiente de la presencia de glucosa 6 fosfato para su acción), que no está tá fosforilada f f il d y es activa, ti y la l glucógeno l ó sintetasa i t t D (dependiente de la presencia de glucosa 6 fosfato para su acción), que está fosforilada y es menos activa. La glucógeno fosforilasa (participante de su degradación) también tiene dos formas: glucógeno degradación), fosforilasa b, menos activa, que no está fosforilada y la glucógeno fosforilasa a, activa, que está fosforilada. DESFOSFORILADAS FOSFORILADAS 1.1.- Regulación de la síntesis de glucógeno y la degradación en estado de buena alimentación En el estado de buena alimentación, la glucógeno sintasa es activada alostericamente por glucosa-6fosfato cuando está presente en concentraciones elevadas. Por el contrario, la glucógeno fosforilasa es inhibida alostericamente por la glucosa-6-fosfato, así como por el ATP En el hígado la glucosa también sirve como un ATP. inhibidor alostérico de la glucosa fosforilasa 1.2.- Activación de la degradación del glucógeno en músculo ú l por calcio l i y AMP. AMP a efecto del Ca2+: a.• Durante la contracción muscular existe una urgencia por ATP, esta energía es aportada por la glucosa almacenada en el glucógeno. Los impulsos nerviosos causan despolarización de la membrana, lo cual a su vez promueve la liberación de Ca2+ desde el retículo sarcoplásmico. El Ca2+ se une a una subunidad subu dad de la a fosforilasa os o asa c cinasa, asa, la a ca calmodulina odu a la a act activa as sin la necesidad de su fosforilación (acción catalizada por la proteína cinasa). Cuando el músculo se relaja, el Ca2+ retorna al retículo sarcoplásmico y la fosforilasa cinasa se torna inactiva. Esta enzima,, la fosforilasa cinasa,, tiene su máxima actividad cuando está fosforilada y con Ca2+ unido. b.- efecto del AMP: b. • La glucógeno fosforilasa muscular es activa en presencia de elevados niveles de AMP, lo que ocurre en el músculo en condiciones de anoxia extrema y alto consumo de d ATP. ATP El AMP ejerce j d efectos dos f t inhibidores i hibid sobre la síntesis de glucógeno: * Fosforilación de la glucógeno sintetasa, sintetasa inactivándola * Inhibición de la fosfoproteína fosfatasa, cuya actividad tendería a restablecer la actividad de la glucógeno sintetasa. 2.- co control t o hormonal. o o a • Las hormonas adrenalina y g glucagón g activan las p proteínas q quinasas que fosforilan ambas enzimas, provocando activación de la glucógeno fosforilasa, estimulando la degradación del glucógeno; mientras que la glucógeno sintetasa disminuye su actividad, lo que g g inhibe la síntesis de glucógeno. • La hormona insulina provoca la desfosforilación de las enzimas, en consecuencia la glucógeno fosforilasa se hace menos activa, y la glucógeno sintetasa se activa, activa lo que favorece la síntesis de glucógeno. • Es decir, q que hormonas como la adrenalina y el g glucagón g favorecen la degradación del glucógeno, mientras que la insulina estimula su síntesis. Conexiones bioquímicas • Existe un grupo de enfermedades heredadas genéticamente que resultan lt en un defecto d f t en alguna l enzima i requerida id para la síntesis o degradación del glucógeno. Esto resulta en: a.- formación de un glucógeno con una estructura anormal a b.- la acumulación de grandes cantidades de glucógeno normal en tejidos específicos. Una enzima particular puede ser defectuosa en un solo tejido como el hígado o el defecto puede ser mas generalizado, afectando músculo,, riñón,, intestino y miocardio. La severidad de las enfermedades va desde fatal en la infancia hasta desordenes medianos que no afectan tan drásticamente la vida. • A continuación se mencionan las enfermedades mas frecuentes relacionadas con la síntesis y degradación del glucógeno: Enfermedad de Von Gierke (Tipo I): deficiencia en la glucógeno-6g , riñón e intestino. Presenta hipoglicemia p g fosfatasa,, afecta hígado, severa en el ayuno. La estructura del glucógeno es normal, pero se acumula en cantidades significativamente mayores que las normales. Enfermedad de Pompe (Tipo II): deficiencia en la alfa-glucógeno oxidasa lisosomal. Presenta concentraciones excesivas de glucógeno en vacuolas citoplásmias anormales. Usualmente ocurre una muerte temprana. temprana Estructura del glucógeno normal. normal Síndrome de McArdle (Tipo V): la afección es en el músculo q , el metabolismo en el hígado g es normal. Debilidad esquelético, temporal, no hay salida de lactato al torrente sanguíneo durante ejercicio atenuante. Elevados niveles de glucógeno con estructura normal en el músculo. • Para que la vida de los mamíferos pueda llevarse a cabo adecuadamente,, es necesario un aporte p constante de glucosa al torrente sanguíneo. Cuando el nivel de glucosa en la sangre es bajo (hipoglicemia), el glucagón producido por el páncreas activa la acción de la glucógeno fosforilasa, estimulando la degradación del glucógeno en el hígado liberando glucosa al torrente sanguíneo con lo que su concentración aumenta. Por el contrario, cuando el nivel de glucosa sanguínea es alto (hiperglicemia), la insulina producida también en el páncreas activa la acción de la glucógeno sintetasa que estimula la síntesis de glucógeno por lo que la concentración de glucosa libre en la sangre disminuye.