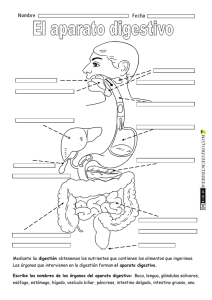
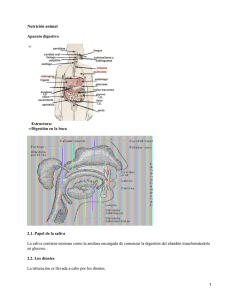
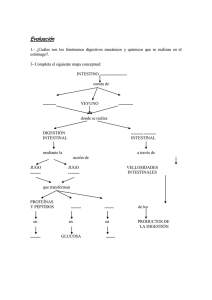
Aparato digestivo
Anuncio

Aparato digestivo 35. Digestión y absorción La digestión es el conjunto de mecanismos por los cuales se produce la degradación de los alimentos y su transformación en moléculas de tamaño reducido. El proceso supone una rotura enzimática tanto de glúcidos, como de proteínas y lípidos, por medio de enzimas de la saliva, jugo gástrico, jugo pancreático y células de las vellosidades intestinales. Los lípidos requieren además una solubilización micelar por medio de los ácidos biliares de la bilis. Los productos finales de la digestión deben a continuación atravesar la pared del tubo digestivo y pasar a la sangre (absorción), para ser posteriormente distribuídos a todas las células del organismo. La absorción intestinal se caracteriza por ser un proceso rápido, de tal forma que la digestión y absorción de una comida compleja pueden completarse en menos de tres horas. Es, además, un proceso extraordinariamente eficaz; por ejemplo, la absorción de lípidos y proteínas es superior al 95 %. Aunque en el estómago puede absorberse alcohol y algún otro tipo de moléculas (ciertos fármacos del tipo de la aspirina), sólo lo hacen en cantidades muy reducidas. Los procesos de absorción tienen lugar fundamentalmente en el intestino delgado y en el intestino grueso. El principal lugar de absorción es el intestino delgado, aunque el agua y las sales pueden hacerlo también en el intestino grueso. Superficie de absorción Superficie (m2) Incremento relativo Tubo liso 0,33 1 Pliegues de Kerckring 1 x3 Vellosidades 10 x 30 Microvellosidades (borde en cepillo) 300 x 1000 Intestino delgado La transferencia de sustancias a través de la pared del intestino es posible gracias a una estructura especialmente adaptada. La superficie interna es extraordinariamente grande (superior a los 300 m2), lo que facilita en gran medida el paso de sustancias a través de la misma y, por tanto, el proceso de absorción. Esto es posible, en primer lugar, gracias a la existencia de una serie de pliegues que incrementan tres veces la superficie (pliegues de Kerckring); y, en segundo lugar, a la presencia de un gran número de pequeñas vellosidades que la aumentan aún más. Por último, las células de la mucosa intestinal tienen un borde en cepillo, constituido por un gran número de microvellosidades que hacen todavía mayor la superficie de absorción. Estas células denominadas enterocitos, se forman a partir de células precursoras en las profundidades de las criptas del Lieberkühn y luego van emigrando hacia el extremo de la vellosidad. Los enterocitos tienen una vida media de unos pocos días y, cuando envejecen, se van descamando a la luz intestinal. El epitelio intestinal se renueva íntegramente en 6 días. 1 Digestión y absorción de glúcidos El consumo diario de glúcidos en los países occidentales es de unos 250-800 g. Más del 50 % se encuentran en forma de almidón, y en menores proporciones como los disacáridos sacarosa y lactosa y los monosacáridos glucosa y fructosa. La digestión de los hidratos de carbono se inicia por medio de la ptialina de la saliva, y continúa en el intestino delgado por la acción de la amilasa pancreática. Ambas actúan sobre los enlaces (α 1- 4) internos de las moléculas de almidón, dando lugar a la formación de maltosa (dímeros de glucosa), maltotriosa (trímeros de glucosa) y dextrinas (que contienen los puntos de ramificación del almidón). Dado que los glúcidos sólo pueden absorberse en forma de monosacáridos, los productos de la digestión o los disacáridos de la dieta tienen que continuar siendo desintegrados en la membrana del borde en cepillo, que contiene diversos tipos de oligosacaridasas (maltasas, lactasas, sacarasas e isomaltasas o dextrinasas). La acción de estas enzimas va a dar lugar a la formación de los monosacáridos glucosa, galactosa y fructosa. La absorción de los monosacáridos glucosa y galactosa, tiene lugar por un mecanismo común dependiente de sodio (transporte activo secundario), dándose fenómenos de inhibición competitiva entre ambas. Una vez que ambos monosacáridos se encuentran concentrados en el interior celular, pueden ya penetrar, por difusión facilitada, a través de la membrana basolateral, y dirigirse a la sangre. A diferencia de la glucosa y la galactosa, la absorción de la fructosa se lleva a cabo mediante un mecanismo de difusión pasiva y, una vez dentro de la célula, se produce una transformación parcial en glucosa antes del paso a la sangre. Digestión y absorción de proteínas La ingestión diaria de proteínas es de aproximadamente 70-90 g, siendo sus fuentes principales la carne y los productos lácteos. Además de la fuente dietética, una parte importante de las proteínas que llegan al intestino proceden también de las secreciones digestivas (25%) o la descamación de células epiteliales (25%). La digestión de las proteínas se inicia por acción de la pepsina del estómago, dando lugar a la formación de polipéptidos, oligopéptidos y algunos aminoácidos. La digestión se continúa en el intestino gracias a las proteasas del jugo pancreático (tripsina, quimotripsina, carboxipeptidasas, colagenasa y elastasa), formándose ya oligopéptidos (30 %) y diferentes aminoácidos (70 %). Al igual que ocurría con los glúcidos, la digestión se completa a nivel del borde en cepillo de las células intestinales o enterocitos. Estos con tiene una serie de aminopeptidasas orientadas hacia el exterior de la membrana, que hidrolizan la mayor parte de los péptidos, liberando aminoácidos. Una pequeña parte de los péptidos pueden, no obstante, pasar al interior de las células a través de un transportador ligado a H+ y se hidrolizan a aminoácidos por medio de peptidasas citoplasmáticas. Los mecanismos de absorción de los aminoácidos son transporte activo secundario acoplado al sodio, existiendo cuatro tipos de transportadores distintos, para los aminoácidos neutros, básicos, ácidos y uno específico para prolina e hidroxiprolina. 2 La mayor parte de los productos de la digestión de las proteínas se absorben en el intestino delgado. Al intestino grueso sólo llegan pequeñas cantidades que serán catabolizadas por la flora intestinal. Es importante señalar que, aunque en proporciones muy pequeñas, también es posible la absorción intestinal de proteínas por mecanismos de pinocitosis. La importancia nutritiva es mínima, pero sí puede tener interés al desencadenar una respuesta inmunológica. Digestión y absorción de lípidos El consumo diario de lípidos es de unos 60-100 g. En su mayor parte son triglicéridos y sólo una pequeña porción se encuentra en forma de lecitinas, ésteres de colesterol o vitaminas liposolubles. Emulsificación, digestión e incorporación a las micelas La solubilización sólo es posible por incorporación a las micelas de la bilis. Cuando la bilis se mezcla con las gotitas de lípidos en el intestino, los lípidos se absorben en las micelas y así se mantienen estables pasando de formar parte de gotas cuyo diámetro era de 0,5 a 1 µ, a micelas cuyo diámetro es de 4 a 6 nm (aproximadamente 1000 veces más pequeñas) La digestión de los lípidos se lleva a cabo a nivel de intestino delgado gracias a la presencia de las enzimas lipolíticas del páncreas. La lipasa pancreática, es la más importante, desdobla los triglicéridos en monogliceridos y ácidos grasos; también parece existir una lipasa gástrica, capaz de digerir triglicéridos de cadena corta, pero su actividad es muy reducida. La fosfolipasa disocia las lecitinas en lisolecitinas y ácidos grasos. La colesterol-ésterhidrolasa hidroliza el colesterol esterificado, originando ácidos grasos y colesterol libre. Al mismo tiempo, la lipasa se absorbe también, manteniéndose anclada a los ácidos biliares gracias a una proteína, la colipasa pancreática. Entonces se produce la hidrólisis de los triglicéridos, con formación de monoglicéridos y ácidos grasos, que se incorporan a las micelas ya que los productos de la hidrólisis de los lípidos son compuestos insolubles en el medio acuoso intestinal. Entrada al enterocito o célula epitelial intestinal Una vez producida la incorporación a las micelas mixtas, los productos de la digestión de los lípidos pueden ya ponerse en contacto con las microvellosidades y absorberse a través de la membrana celular por difusión. Para penetrar en el interior de los enterocitos, las moléculas lipídicas difunden primero a la zona de líquido que rodea a éstos y luego penetran a través de la membrana epitelial. Las micelas difunden entonces en sentido retrógrado y vuelven a absorber nuevos lípidos, que son transportados hacia las células de las vellosidades. La absorción intestinal de los lípidos es un proceso muy eficaz. Más del 95 % de los mismos se recuperan, fundamentalmente a nivel duodenal, y sólo una pequeña cantidad se pierde cada día a través de las heces. Metabolismo celular y formación de quilomicrones Una vez en el interior de las células intestinales, los productos de la digestión de los lípidos se unen a una proteína transportadora de bajo peso molecular, la cual los lleva hasta el retículo 3 endoplasmático liso. En éste tiene lugar la resíntesis de triglicéridos, la de lecitinas y la de colesterol esterificado. Los diferentes lípidos se agrupan posteriormente y se rodean de una cubierta de betalipoproteínas formadas en el aparato de Golgi, dando lugar a la aparición de los quilomicrones. Su composición aproximada sería: 87% de triglicéridos, 9% de fosfolípidos y colesterol libre, 3% colesterol esterificado y 1% de vitaminas liposolubles y proteínas. Recogida del sistema linfático Estos quilomicrones abandonan la célula, por exocitosis, a través de la membrana lateral y se dirigen a los quilíferos (o lactóforos) centrales de las vellosidades y al interior del sistema linfático. Los quilomicrones presentan un tamaño lo bastante grande (100 nm) como para dar a la linfa, e incluso al plasma circulante, un aspecto lechoso después de una comida rica en lípidos. Una excepción al mecanismo descrito son los ácidos grasos de cadena corta y media, que abandonan las células sin esterificación y sin incorporarse a los quilomicrones, pasando por simple difusión a los capilares sanguíneos. Los triglicéridos de cadena media (TGCM), a diferencia de los de cadena larga y corta, penetran en la célula de forma directa, sin disociación previa por medio de la lipasa. Un 30 % de los TGCM pueden ser absorbidos sin hidrólisis previa; los que son hidrolizados lo hacen por las lipasas pancreáticas, formando rápidamente micelas. Una vez en el interior de la célula no se reesterifican sino que pasan en seguida a la circulación venosa portal. Transporte de los lípidos en sangre Lípidos principales Tipo Origen Destino Quilomicrón Intestino Células TG y otros VLDL Hígado Células TG y colesterol LDL Vasos (resto de VLDL) Hígado Colesterol Transporte colesterol HDL Hígado e intestino Colesterol Elimina y degrada el colesterol Hígado y células con alto uso de colesterol Función Transporte de lípidos de la dieta Transporte de lípidos endógenos A través de la linfa se incorporan a sangre. En las células endoteliales, por medio de la lipoproteín-lipasa se hidrolizan los triglicéridos de los quilomicrones, dando ácidos grasos libres y glicerol que serán usados por las células. Los restos del quilomicrón con elevado contenido en colesterol son llevados al hígado donde son absorbidos por endocitosis. El colesterol y triglicéridos sintetizados en el hepatocito son secretados a sangre unidos a apoproteínas formando complejos denominados VLDL (lipoproteínas de muy baja densidad), que permiten el transporte de triglicéridos a diferentes órganos. Al perder triglicéridos las VLDL pasan a LDL y el excedente de colesterol se reenvía al hígado unido a proteínas formando HDL. 4 Absorción de agua e iones El agua que pasa por el intestino delgado, aproximadamente unos 9 litros diarios (2 litros procedentes de la alimentación y unos 7 litros procedentes de las secreciones digestivas), se reabsorbe en más de un 80 % en el intestino delgado. El resto lo hace en el intestino grueso, de tal modo que sólo una pequeña cantidad, alrededor del 1 %, se elimina con las heces. Los desplazamientos del agua tienen lugar por mecanismos pasivos osmóticos, y se relacionan normalmente con el transporte de solutos, siguiendo el gradiente osmótico creado por las sustancias absorbidas a la sangre. El paso del sodio al espacio intercelular produce en éste una solución hipertónica que facilita el arrastre de agua como consecuencia del gradiente osmótico creado. El agua pasa desde la luz del intestino a través de las denominadas uniones estrecha, existentes entre las membranas de células adyacentes (vía paracelular), o en menor cuantía, a través del interior celular (vía transcelular). Las uniones estrechas presentan una permeabilidad decreciente, desde el duodeno hasta el colon, de tal modo que la participación de la vía paracelular en el transporte de agua se va reduciendo a medida que se progresa a lo largo del intestino. Una vez en el espacio intercelular, el agua, gracias al gradiente de presión hidrostática creado, puede pasar a través de la membrana basal (mucho más permeable que las uniones estrechas) hacia los capilares sanguíneos. El cloro se absorbe en duodeno y yeyuno por difusión pasiva siguiendo al Na+. En el ileon y colon, realiza un cotransporte activo con el Na+, y también un intercambio con el bicarbonato. El potasio difunde a través de canales y al interior celular por la bomba Na+/K+. El calcio,se absorben diariamente de 200 a 300 mg mediante transporte activo transcelular, y transporte paracelular pasivo. El magnesio, se absorbe como el calcio El fosfato, se absorbe alrededor del 65% mediante procesos pasivos, y también por un mecanismo acoplado al Na+, utilizando un cotransportador. El hierro, en su forma hemo, penetra en las células intestinales mediante la formación de vesículas, seguido de digestión enzimática y liberación del hierro libre que se combina inmediatamente con la apoferritina para formar ferritina. El hierro en su forma no hemo, se libera de los alimentos y se solubiliza gracias a la acidez del jugo gástrico entre otros, lo cual permite el paso de hierro férrico (Fe +++ ) a ferroso (Fe++). Composición de las heces A lo largo de un día se excretan de 100 a 150 gramos de materia fecal, de la que entre 30 y 50 gramos son sólidos y de 70 a 100 gramos son de agua. La materia sólida está formada por residuos de materiales no digeribles (ej: celulosa), pigmentos y sales biliares, secreciones intestinales, principalmente mucus, células epiteliales descamadas, bacterias (hasta 1/3 de los sólidos totales), y materiales inorgánicos: Ca++, fosfatos, etc. Las heces eliminadas diariamente a través del recto contienen una cantidad de líquido inferior a los 200 ml. Cantidades superiores determinan la aparición de diarrea, que puede tener diversos orígenes. El fenómeno puede producirse, por ejemplo, por el uso de laxantes, capaces de bloquear la actividad de la ATPasa Na/ K y la absorción intestinal de sodio. El cólera o determinadas 5 infecciones bacterianas también pueden originar diarrea al estimular la secreción de agua y electrolitos por las criptas de Lieberkühn; por este mecanismo pueden llegar a perderse hasta 5-10 litros de agua cada día, con grave riesgo para la vida. 6