1 . meristemos - Atlas de Histología Vegetal y Animal
Anuncio

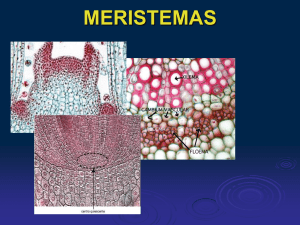
ATLAS de HISTOLOGÍA VEGETAL y ANIMAL Tejidos vegetales 1 . MERISTEMOS Pilar Molist, Manuel A. Pombal, Manuel Megías Departamento de Biología Funcional y Ciencias de la Salud. Facultad de Biología. Universidad de Vigo. (Versión: Agosto 2014) Este documento es una edición en pdf del sitio htttp://webs.uvigo.es/mmegias/inicio.html y ha sido creado con el programa Scribus (http://www.scribus.net/canvas/Scribus) Todo el contenido de este documento se distribuye bajo la licencia Creative Commons del tipo BY-NC-SA (Esta licencia permite modificar, ampliar, distribuir y usar sin restricción siempre que no se use para fines comerciales, que el resultado tenga la misma licencia y que se nombre a los autores). ÍNDICE 1. Introducción ................................ 4 2. Merisemos ......................... 6 2.1. Apical caulinar ................. 8 2.2. Apical radical ................... 9 2.3. Cambium vascular ............ 10 2.4. Cambium suberoso ........... 11 Imágenes sin marcas ............... 12 1. INTRODUCCIÓN Cuando hablamos de las características de los tejidos de las plantas tenemos que tener en mente la historia ocurrida hace 500 millones de años, cuando las plantas conquistaron la tierra. El medio terrestre ofrece ventajas respecto al medio acuático: más horas y más intensidad de luz, y mayor circulación libre de CO2. Pero a cambio las plantas tienen que solventar nuevas dificultades, casi todas relacionadas con la obtención y retención de agua, con el mantenimiento de un porte erguido en el aire y también con la dispersión de las semillas en medios aéreos. Para ello las plantas agrupan sus células y las especializan para formar tejidos con funciones determinadas que sean capaces de hacer frente a estas nuevas dificultades. A su vez los tejidos se agrupan para constituir órganos. representado por dos tejidos: colénquima y otro más especializado denominado esclerénquima. Una gran cantidad del tejido de las plantas es el parénquima, el cual realizará diversas funciones, dede la fotosíntesis hasta el almacen de sustancias. Sin embargo, uno de los hechos más relevantes en la evolución de las plantas terrestres es la aparición de un sistema conductor capaz de comunicar todos los órganos del cuerpo de la planta, formado por dos tejidos: xilema, que conduce mayormente agua, y floema, que conduce principalmente sustancias orgánicas en solución. Sólo hablamos de verdaderos tejidos conductores en las plantas vasculares. Finalmente, las plantas vasculares producen semillas, dentro de las cuales se forma el embrión, que se desarrolla y crece gracias a la actividad de los tejidos embrionarios o meristemáticos. Los meristemos , no sólo están presentes en el embrión sino que están activos a lo largo de toda la vida de la planta, permitiendo su crecimiento. Todos estos tejidos, excepto los meristemos, han derivado a lo largo de la evolución de otro tejido poco diferenciado llamado parénquima, que se mantiene en las planta actuales y que realiza múltiples funciones. Para superar un medio ambiente variable y seco, aparece un sistema protector formado por dos tejidos: la epidermis y la peridermis. Las células de estos tejidos se revisten de cutina y suberina para disminuir la pérdida de agua, y aparecen los estomas en la epidermis para controlar la transpiración y regular el intercambio gaseoso. Para mantenerse erguidas sobre la tierra las plantas tienen un sistema de sostén Clasificación de los tejidos de las plantas. 4 juntas a las células. Luego, cada célula sintetizará la pared celular primaria, a ambos lados de la lámina media, formada principalmente por hemicelulosas y celulosas. Algunas plantas, además, poseen células que pueden sintetizar la pared celular secundaria que, además de celulosa, por lo general contiene lignina. Todas las células de las plantas diferenciadas contienen lamina media y pared celular primaria más o menos gruesa pero sólo unos pocos tipos celulares tienen además pared celular secundaria. Los tejidos y sistemas de tejidos se agrupan para formar órganos que pueden ser vegetativos, como la raíz (órgano de captación de agua y sales), tallo (órgano para el transporte, sostén y a veces realiza la fotosíntesis) y hoja (órgano que capta la energía solar y realiza la fotosíntesis y es el principal responsable de la regulación hídrica de la planta), o bien reproductivos como la flor y sus derivados, la semilla y el fruto. Antes de introducirnos en el estudio de cada uno de los tejidos y órganos tenemos que entender dos conceptos característicos de las plantas: 2.- A partir del estado embrionario las plantas se desarrollan y crecen gracias a la actividad de los meristemos . El primer crecimiento de todas las plantas, y único en algunos grupos, es el crecimiento en longitud. Éste se denomina crecimiento primario, y corre a cargo de la actividad de un grupo de células meristemáticas que se sitúan en los ápices de los tallos y raíces, así como en la base de los entrenudos. Estos grupos de células son los meristemos primarios. Además, algunos grupos de plantas también pueden crecer en grosor, un tipo de crecimiento denominado crecimiento secundario, y lo hacen gracias a la actividad otro tipo de meristemos denominados meristemos secundarios. 1.- Las células de las plantas presentan una estructura denominada pared celular que recubre externamente a su membrana plasmática. Está sintetizada por la propia célula y es imprescindible para ella, puesto que aporta la rigidez necesaria en ausencia de un citoesqueleto bien desarrollado, del cuál carecen las células de las plantas. Cuando una célula de una planta se divide, lo primero que se deposita es un tabique separador denominado lámina media, formada por sustancias pécticas, que se sitúa entre las dos células hijas. Las sustancias pécticas son moléculas adherentes que tienden a mantener 5 2. MERISTEMOS Los meristemos son los responsables del crecimiento permanente de las plantas y están presentes durante toda la vida de éstas. Las células meristemáticas presentan las características citológicas de las células indiferenciadas. Son pequeñas, isodiamétricas y tienen una pared celular primaria delgada. Su citoplasma contiene características propias, como abundantes ribosomas, un retículo endoplasmático rugoso escaso, el complejo de Golgi muy desarrollado para fabricar los componentes de la pared celular, numerosos proplastidios, muchas y pequeñas vacuolas y un citoplasma desprovisto de inclusiones. El núcleo, con mucha cromatina condensada, es grande y se sitúa en posición central. Las células meristemáticas son células totipotentes, están continuamente dividiéndose por mitosis y posteriormente se diferencian para originar el espectro entero de tipos celulares de una planta adulta. La clasificación de los meristemos se realiza en base a su posición en el cuerpo de la planta. protodermis, que origina la epidermis, el procambium, que produce los tejidos conductores primarios (xilema y floema primarios), y el meristemo fundamental, que dará lugar al resto de los tejidos de la planta. Sin embargo, en las plantas monocotiledóneas, la mayor parte del crecimiento en longitud del tallo, no depende tanto de los meristemos apicales, sino que es responsabilidad de los meristemos intercalares. Como su nombre indica, se encuentran intercalados entre tejidos no meristemáticos. Los mejor conocidos son aquellos localizados en los entrenudos (principalmente en la base del entrenudo) y en la vaina de las hojas. En aquellas plantas que crecen en espesor, es decir, tienen crecimiento secundario, presentan otro tipo de meristemos denominados meristemos secundarios, también llamados meristemos laterales. Éstos son responsables del aumento de diámetro de tallos y raíces y sus células se dividen según planos periclinales (ver figura). Estos meristemos no existen en muchas plantas herbáceas, como la mayoría de las pteridofitas y monocotiledóneas, ni en determinados órganos como las hojas. Hay dos tipos de meristemos laterales: el cambium vascular, que origina los tejidos conductores secundarios (xilema y floema secundarios), y el cambium suberoso o felógeno, que origina la peridermis. Ambos meristemos se disponen como un cilindro continuo o como un anillo incompleto a lo largo del tallo o de la raíz. El primero se localiza entre el xilema secundario y el floema secundario y el segundo entre la felodermis y el súber. Los meristemos primarios son los responsables del crecimiento en longitud de la planta por lo que sus células se dividen por tabicación anticlinal (ver figura ). Dentro de ellos tenemos a los meristemos apicales que se sitúan en el ápice del tallo y de cada rama, y en el ápice de la raíz principal y de cada una de las raíces secundarias. En el primer caso están protegidos por los primordios foliares, mientras que en el segundo por la cofia o caliptra. Estos meristemos se denominan meristemos apicales caulinares y meristemos apicales radicales, respectivamente. A partir de cada meristemo apical se formarán la 6 Clasificación de los meristemos de las plantas. 2.1. Meristemo apical caulinar El meristemo apical caulinar está formado por células que tienen una gran capacidad de división, aunque la velocidad a la que esto ocurre depende de las condiciones ambientales. Por ejemplo, puede seguir ritmos circadianos. Al igual que en el meristemo de la raíz, la tabicación de las células en división sigue planos perpendiculares al eje longitudinal del tallo, es decir, la tabicación es anticlinal (ver figura). A partir de este meristemo se van a formar los tejidos primarios del tallo. Además, durante el crecimiento de la planta aparecen nuevos meristemos apicales (primordios) en el cuerpo de la planta que dan lugar a ramificaciones laterales, hojas y órganos reproductores. Inmediatamente debajo del ápice del tallo, dispuestos lateralmente, aparecen los primordios de las hojas que experimentan un rápido crecimiento, sobrepasando al ápice del tallo y recubriéndolo para protegerlo. Los primordios de las ramificaciones se sitúan un poco más apartados. Tallo: Meristemo apical caulinar Especie: Vid (Vitis vinifera) Técnica: Corte en parafina y tinción con azul de metileno (según protocolo de Xurxo Gago) 8 2.2. Meristemo apical radicular Las células meristemáticas presentan un núcleo grande de cromatina condensada y una pared celular primaria delgada que les permite dividirse continuamente por mitosis. Es el caso de las células meristemáticas de la raíz. Estas divisiones siguen una asincronía uniforme, es decir, son constantes los porcentajes de células que se encuentran en cada fase de la división mitótica. Durante la citocinesis se forma el tabique separador de las células hijas que es perpendicular al eje longitudinal de la raíz, permitiendo de esta manera el crecimiento en longitud de la misma. Este tipo de tabicación se denomina anticlinal (ver figura). A partir de este meristemo se produce el crecimiento primario de la raíz y da lugar a los tejidos primarios. todos los autores describen este centro, otros proponen que constituye una reserva de células que sirven para regenerar zonas meristemáticas dañ adas por acción mecánica. El meristemo radicular se encuentra en una posición subterminal de la raíz, protegido en su avance a través de la tierra por la cofia o caliptra, la cual está formada por células parenquimáticas secretoras que segregan un glucosaminoglucano de acción lubrificante. Algunas de las células de la cofia, llamadas estatocistes, contienen inclusiones citoplasmáticas sólidas situadas en la parte inferior de la célula, que son principalmente granos de almidón, y que están implicados en el crecimiento de la raíz con dirección geotrópica. En la parte superior de los estatocistes se sitúa el núcleo redondo, grande y de cromatina condensada. Las células más externas de la cofia se mueren y se desprenden siendo reemplazadas a partir del meristemo radicular. Además de las células que se dividen continuamente, el meristemo radicular contiene un grupo de células inactivas mitóticamente que forman el llamado centro quiescente. Aunque no Raíz: Meristemo apical radical protegido por la cofia o caliptra Especie: Cebolla (Allium cepa) Técnica: Corte en parafina y tinción con orceína acética 9 2.3. Cambium vascular Las células del cambium vascular se dividen periclinalmente, las dos células hijas se disponen en una línea perpendicualr a la superficie de la planta, originando el floema secundario hacia el exterior y el xilema secundario hacia el interior. Este meristemo presenta dos tipos celulares, las iniciales fusiformes, que se disponen verticalmente en la planta, y las iniciales radiales, que se disponen horizontalmente. Las primeras originan todos los tipos celulares del xilema y del floema secundario dispuestas verticalmente y las segundas a las que lo hacen horizontalmente. El cambium vascular procede del procambium, formando el cambium fascicular, y de la desdiferenciación del parénquima o del colénquima, formando el cambium interfascicular. De esta forma el cambium vascular se dispone en forma de anillo completo o en forma de bandas paralelas al eje principal de la planta. Tallo: Cambium vascular Especie: Girasol (Heliantthus annuus) Técnica: Corte en parafina teñido con safranina / azul alcián 10 2.4. Cambium suberoso o felógeno felodermis hacia el interior. Las tres estructuras, súber, cambium y felodermis, forman lo que se llama la peridermis, que sustituye a la epidermis cuando se produce el crecimiento secundario de la planta. La actividad estacional del felógeno origina un mayor número de capas de súber que de felodermis. Este hecho hace que las capas superficiales de súber y los restos de epidermis queden aislados por la actividad del felógeno y se desprendan anualmente. Estas capas superficiales que se desprenden es a lo que se llama el ritidoma. Las células del cambium suberoso o felógeno presentan las características citológicas típicas de las células meristemáticas, pero además son las únicas que pueden contener cloroplastos funcionales. Se originan a partir de la desdiferenciación de células parenquimáticas o de las colenquimáticas. Son células alargadas que se disponen en la corteza del tallo formando un cilindro contínuo o a distintos niveles de profundidad desde la superficie formando placas. Se dividen periclinalmente (según planos paralelos a la superficie de la planta) originando súber o corcho hacia el exterior del tallo y Tallo: Cambium suberoso o felógeno Especie: Saúco (Sambucus spp) Técnica: Corte en parafina teñido con safranina / azul alcián 11 Tallo: Meristemo apical caulinar Especie: Vid (Vitis vinifera) Técnica: Corte en parafina y tinción con azul de metileno (según protocolo de Xurxo Raíz: Meristemo apical radical protegido por la cofia o caliptra Especie: Cebolla (Allium cepa) Técnica: Corte en parafina y tinción con orceína acética 12 Tallo: Cambium vascular Especie: Girasol (Heliantthus annuus) Técnica: Corte en parafina teñido con safranina / azul alcián Tallo: Cambium suberoso o felógeno Especie: Saúco (Sambucus spp) Técnica: Corte en parafina teñido con safranina / azul alcián 13