dinamica de proteinas
Anuncio

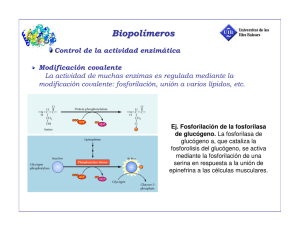
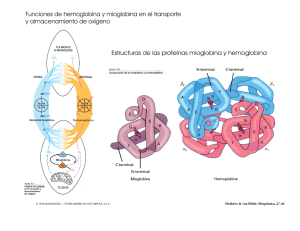
DINAMICA DE PROTEINAS Función dinámica. Las proteínas globulares tienen en común el unirse a otra molécula más pequeña llamada ligando para ser funcionales. Hay un sitio de unión perfectamente definido donde se une el ligando de manera específica y selectiva. Es una parte pequeña de la proteína pero esencial. Este sitio tendrá grupos funcionales que establezcan uniones de tipo débil con el ligando y deberá ser accesible desde fuera. Normalmente el sitio de unión y el ligando son complementarios, sólo cabe ése y no otro. Se forma un complejo proteína - ligando que es reversible excepto en el caso de antígenos y anticuerpos . Si la proteína es un enzima catalizará la reacción en que el ligando es el sustrato, la proteína se une al ligando que se transforma en producto y luego se disocia y el enzima queda libre. La función se ejerce sobre el ligando. Consideraciones generales sobre la interacción proteína - ligando. Función de saturación. 1 sitio de unión por proteína. P + L ↔ PL K es en el sentido de disociación del ligando. La actividad biológica depende de cuanto ligando hay unido, por lo tanto de la concentración de ligando. La función de saturación ν son los moles de ligando unidos partido por los moles de proteína totales: El valor máximo de la función de saturación es 1, como depende de la concentración de ligando, cuanto más haya mayor será ν . Es la ecuación de una hipérbola con una asíntota en 1: Transformando en una recta: Más de un sitio de unión por proteína. Se ocupan con el mismo ligando. Si tiene 2 sitios de unión se ocupa primero uno y luego el otro. Si todos los sitios son iguales (K iguales) hay dos posibilidades: Sitios independientes: Es el caso más sencillo. Si hay varios sitios iguales es porque la proteína tiene varias cadenas iguales con un sitio cada una. Como son iguales K1 = K2 y no se puede saber cual se ocupa primero, son independientes porque no hay conexión entre ellos. La ocupación de un sitio tiene la misma constante aunque ya haya otro ocupado. Si la proteína tiene dos sitios de unión: La asíntota estará en 2. 4Para compara proteína se usa la fracción de saturación: Y = ν/n Ahora la variación es entre 0 y 1. Sigue siendo una hipérbola. El porcentaje de saturación es Y x 100. Fracción y porcentaje de saturación van unidos a la funcionalidad de la proteína. Sitios que interaccionan: Se produce un efecto llamado cooperatividad en el que cuando se ocupa un sitio activo la unión de un segundo ligando en un sitio igual se ve afectada, siendo favorecida (cooperatividad positiva) o perjudicada (cooperatividad negativa). Para que haya cooperatividad se deben dar una serie de requisitos: - Deben haber varios sitios iguales, por lo tanto varias cadenas iguales, la proteína será oligomérica. Todas las oligoméricas presentan cooperatividad, la mayor parte de las veces positiva. Ahora ν no es una hipérbola sino una curva sigmoidea: En el caso de la unión del O2 por la hemoglobina hay cuatro sitios. Las proteínas tienen poca afinidad por el ligando al principio pero como presenta cooperatividad positiva cada vez se unen más rápido y se obtiene un salto en la curva. Explicación de la curva sigmoidea por Hill: Si la cooperatividad fuera infinita en cuanto se uniera un ligando inmediatamente se unen los demás, la proteína tendría una afinidad muy alta por el ligando. c es el coeficiente de Hill que cuantifica la cooperatividad. Si c=1 no hay cooperatividad. La cooperatividad será positiva si c varía entre 1 y n y negativa si varía entre 0 y 1. Las proteínas cooperativas al representar ν frente a [L] sale una curva sigmoidea con asíntota en n, al representar Y frente a [L] la asíntota estará en 1, y si son porcentajes la asíntota es 100. Las proteínas cooperativas son más sensibles a los cambios en la concentración de ligando porque para conseguir una variación en la saturación hay que variar menos la concentración de sustrato porque la entrada de uno favorece las de los demás. Esto es importante para la célula porque ahorra espacio. Efecto homotrópico: efecto de la entrada de un ligando sobre la entrada de otro igual. La mayor parte de las proteínas cuya función hay que controlar son cooperativas en la unión del ligando son casi siempre oligoméricas.. Efecto heterotrópico: efecto de un ligando sobre otro distinto. Tienen otros sitios específicos distintos para unir ligandos distintos llamados moduladores o efectores. La acción no se lleva a cabo sobre él sino que puede afectar a la actividad de la proteína de 2 maneras distintas: - Si el ligando se une mejor: el modulador es un activador que se une en un sitio distinto del activo y favorece la unión del ligando. - Si el ligando se une peor: el modulador es un inhibidor que perjudica la unión del ligando. Este efecto se llama alosterismo. Las proteínas que es necesario regular presentan cooperatividad en la unión del ligando y alosterismo porque son dos procesos muy ventajosos. Para que haya alosterismo la proteína no debe ser oligomérica necesariamente Al actuar un activador la curva sigmoidea se desplaza a la izquierda y el límite es la hipérbola, la cooperatividad baja porque mejora la afinidad por el ligando (el efecto del sustrato es menor) y en la hipérbola es nula.. Al actuar un inhibidor la curva se desplaza a la derecha. Explicación estructural de la cooperatividad y el alosterismo. Cuando el ligando se une se produce un cambio en la conformación. Cooperatividad: al unirse el ligando la proteína se acomoda a él produciendo u pequeño cambio en la conformación. Si la proteína presenta estructura cuaternaria las interacciones entre las subunidades cambian, con lo que varía la conformación. Si es mejor que la anterior la cooperatividad es positiva y si no negativa. Si se pierde la estructura cuaternaria se desunen las subunidades y al no comunicarse se pierde la cooperatividad. Alosterismo: igual para el activador, en el caso del inhibidor se dificulta la entrada del ligando. Modelos moleculares Modelo concertado (Monod, Wyman, Changeux). Propone que la proteína puede existir en dos conformaciones, la forma T (tensa) con poca afinidad por el ligando y la forma R (relajada) con mucha afinidad. Las dos formas existen en ausencia de ligando y la proteína es simétrica, cada subunidad no puede tener conformación distinta a la otra. Todo lo que favorezca la forma R tendrá un efecto positivo sobre la unión del ligando. Al añadir más ligando se va estabilizando la forma R y cada vez hay más.. En la cooperación positiva el ligando se une a la forma R y la estabiliza para que no vuelva a la forma T (como un activador) y en la negativa se estabiliza la forma T. Constante alostérica: relación entre la forma de baja afinidad y la de alta. Cuanta más cooperatividad hay más vale L. Modelo secuencial (Koshland, Nametry, Filmer). La proteína puede existir en dos conformaciones, una de alta afinidad por el ligando y otra de baja. El cambio en la conformación está inducido por la entrada del sustrato. Al unirse el ligando cambia la conformación de la otra subunidad, afectando a las interacciones entre las subunidades y permite la unión de otro ligando. Si K2 > K1 el efecto es positivo y si es al revés es negativo. ESTUDIO DE LAS PROTEINAS QUE UNEN O2. Mioglobina. Formada por una cadena polipeptídica. Necesita un cofactor (grupo prostético) para unir el O2 sin el cual no es funcional. Este grupo es el hemo, formado por una parte orgánica derivada de las porfirinas llamada protoporfirina IX y una parte inorgánica esencial que es el Fe2+. Cada cadena tiene su grupo hemo. El hierro tiene 6 posiciones de coordinación, 4 con la protoporfirina con la cual forma un plano y otros dos, uno por encima y el otro por debajo. Uno de ellos se une a la cadena polipeptídica por medio de la histidina F8 (cadena F, posición 8) también llamada proximal porque está cerca del O2. La otra histidina es la distal. Para que el hierro esté en forma de ferroso el entorno debe ser hidrofóbico, porque si hay agua pasa a férrico y la sexta posición de coordinación está ocupada por el agua y el O2 no puede unirse. Sólo el grupo hemo une O2, el resto de la proteína es para preservar el entorno hidrofóbico y que el Fe no se oxide completamente. Cuando el O2 se une el Fe no es oxidado sino que se oxigena, porque el O2 no cede completamente los electrones sino que cuando se va se los lleva Diferencias con la hemoglobina: - La hemoglobina tiene 4 cadenas iguales dos a dos (dos α y dos β). Cada cadena es igual a la mioglobina (grupos hemo unidos a las mismas histidinas). - La mioglobina tiene 1 cadena con 1 grupo hemo por lo que une 1 molécula de O2. - La hemoglobina tiene 4 cadenas, 4 grupos hemo en total, por lo tanto puede unir 4 moléculas de O2. Unión del O2. Mioglobina. Mb + O2 ↔ MbO2 Desoximioglobina oximioglobina A la presión parcial de O2 que el 50% de los sitios están ocupados se le llama P50 y es una medida de la afinidad de la proteína por el ligando Si la P50 sube la pO2 debe ser alta para ocupar la mitad de los sitios, y si P50 es baja es que tiene mucha afinidad. Hemoglobina Hemoglobina + 4O2 Hemoglobina(O2)4 Desoxihemoglobina hemoglobina Coeficiente de Hill, c = 2.8. Cooperatividad positiva muy alta. El primer O2 se une con poca afinidad y el cuarto con afinidad varios órdenes de magnitud más alta. Prácticamente o está vacía o llena. Representando Y para la mioglobina y la hemoglobina la asíntota está en 1. Se ve que la hemoglobina es menos afín porque está desplazada a la derecha. El P50 de la mioglobina es mucho mayor que el de la hemoglobina (26 mm Hg). La hemoglobina como tiene 4 subunidades tiene menos afinidad por el O2 que la mioglobina. La hemoglobina transporta el O2 desde los pulmones o las branquias donde la pO2 es muy alta. Si la pO2 es alta las dos proteínas se saturan. Como la pO2 es baja en los tejidos (20 - 30 mm Hg) al llegar la hemoglobina como está en torno a su P50 libera el 50% del O2. Si la mioglobina estuviera en los eritrocitos no liberaría el O2 porque a esa pO2 tiene mucha afinidad. Por eso la hemoglobina es más adecuada para el transporte. Cuando la hemoglobina suelta el O2 lo capta la mioglobina porque a esa pO2 es favorable. Cuando la pO2 en los tejidos baja lo libera la mioglobina. Causa molecular de la cooperatividad. Como el índice de cooperatividad es alto podemos tener desoxihemoglobina u oxihemoglobina , pero las formas intermedias son poco importantes. Al ocupar el O2 un grupo hemo los otros reaccionan produciendo un cambio conformacional que se comunica entre ellos por medio de la estructura cuaternaria. Las estructuras de la forma oxihemoglobina y la desoxihemoglobina son distintas. La forma T (desoxi) es más estable por la participación de puentes salinos de naturaleza iónica y muchos puentes de hidrógeno. Estas interacciones fijan la estructura e impiden la movilidad. La desoxihemoglobina se pliega dejando un hueco central entre la 4 subunidades. En la oxihemoglobina no hay puentes salinos y la cavidad es más pequeña (forma R). La hemoglobina tiene dos cadenas α y dos β. Hay interacciones muy fuertes entre αβ como si la molécula tuviera dos dímeros iguales. Cuando entra una molécula de O2 se produce un cambio en la conformación y un dímero gira con respecto al otro 15°.La conformación de la forma con O2 y la que no tiene por tanto no es la misma. El paso de la forma T a la R lo determina la entrada de O2. Esto ocurre porque el O2 se une al Fe del hemo de cada cadena, en la forma desoxi el Fe está por encima del plano que forma el grupo hemo unido a la histidina F8. Al unirse el O2 y ocupar la sexta posición de coordinación el Fe pasa a tener 6 sustituyentes y reordena sus electrones colocándose en el centro del grupo hemo. Al moverse el Fe arrastra consigo a la histidina y ésta por medio de la hélice a toda la molécula. Por ello un grupo hemo se comunica con otro por medio de la cadena polipeptídica. Al cambiar la conformación los grupos que formaban los puentes salinos ya no estarán enfrentados y no interaccionarán. La entrada de la primera molécula de O2 será más difícil porque se una a la forma T con lo cual es más difícil mover el Fe hacia el plano del hemo por impedimentos de tipo estérico entre la histidina y la valina del grupo hemo. Cuando se une el O2 la tensión es grande en la molécula y se rompen puentes salinos, la proteína pasa a la forma R y la unión de O2 es más favorable. Otros ligandos que pueden unirse a la hemoglobina. La hemoglobina puede unir otros ligandos que afecten a la unión del O2 (efecto heterotrópico).Su efecto es la disminución de la afinidad de la hemoglobina por el O2. Protones: se unen a grupos que se estén disociando a pH celular. Los grupos afectados en la hemoglobina son el imidazol de la histidina y los 4 α-amino. Al aumentar la concentración de protones disminuye la afinidad y la curva de ν se desplaza a la derecha. Como consecuencia del metabolismo aumenta la concentración de protones en los tejidos periféricos y cuando la hemoglobina llega disminuye su afinidad por el O2 y los suelta. Los protones se unen a los grupos que formaban el puente salino que al protonarse vuelven a formarlo pasando a la forma T. Al aumentar luego el pH se desprotonan y se rompen los puentes salinos. La primera molécula de O2 rompe más puentes salinos que la segunda por lo que la cooperatividad es positiva. Los protones no se unen al hemo, sino a otros grupos que son importantes porque determinan en parte la conformación. CO2: el CO2 es la forma más oxidada del C, es un producto final del metabolismo. Su concentración aumenta en los tejidos donde se degrada la materia orgánica. Se transporta en forma de ácido carbónico disuelto en la sangre hasta los pulmones donde se intercambia. Está disociado en HCO3- y H+. Esos protones tienen efecto sobre la afinidad del O2. El CO2 se une a la hemoglobina por medio del α-amino de la cadena polipeptídica: Esta reacción da lugar a carbamatos que favorecen la formación de más puentes salinos al introducir la carga negativa (se da sólo en la forma desoxi). La hemoglobina o tiene unidos H+ y CO2 o tiene el O2: Efecto Bohr: Efecto conjunto de H+ y CO2 sobre la afinidad de la hemoglobina por el O2. En los tejidos se producen H+ y CO2 y el pH baja. Hay tendencia a que la hemoglobina suelte el O2. En los pulmones la presión parcial de O2 es alta por lo que se capta el O2 y se eliminan H+ y CO2 pasando a la forma oxihemoglobina. 2,3-Bifosfoglicerato (2,3-BPG): está cargado a pH fisiológico: Presente en alta concentración en la sangre, comprable a la de la hemoglobina. Es un ligando que se une a la hemoglobina en proporción 1:1 por medio de la cavidad central del tetrámero. Tendrán que haber muchas cargas positivas para estabilizarlo, uniéndose a ellas iónicamente. Para soltarlo habrá que romper los enlaces iónicos por medio de energía. En la forma oxihemoglobina el hueco central es más pequeño y el 2,3-DPG no cabe. El efecto que produce es la disminución de la afinidad porque aporta interacciones extra que hay que romper para pasar a la forma oxi. Aumenta la cooperatividad desplazando la curva de ν a la derecha. Significado fisiológico: la sangre se acomoda a diferentes presiones parciales de O2 modificando la concentración de 2,3-DPG de modo que cuanta menos PO2 más aumenta la concentración de 2,3-DPG. Esto es para que la hemoglobina pierda bastante afinidad por el O2 para que lo libere a una PO2 menor. Otro efecto es la oxigenación de la hemoglobina fetal, que tiene composición distinta a la de los adultos. La HbA (adultos) tiene α2β2 y la HbF es α2γ2. La diferencia estriba en la secuencia de aminoácidos. La HbF debe tener más afinidad por el O2 que la HbA que la nutre. En la cadena γ un residuo de histidina cargada positivamente es sustituido por una serina (no cargada). Al eliminar una carga positiva el 2,3-DPG se une peor y es más fácil pasar a la forma oxiHb. http://www.loseskakeados.com