EL NUCLEO
Anuncio

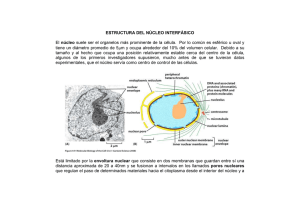
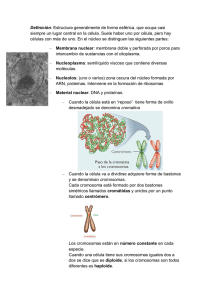
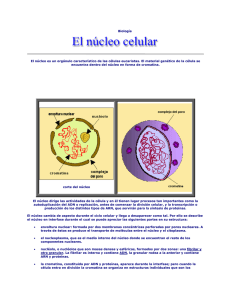
EL NUCLEO En 1830, Brown estableció que el núcleo era una constante dentro de la estructura celular; pero hoy día, aun manteniendo este concepto, hay que precisar que esto es así solo en las células eucarióticas, tanto vegetales como animales. El núcleo, como orgánulo celular, alberga en su interior la información genética en forma de ADN, y es el lugar donde se realiza la replicación del ADN y la síntesis de todos los ARN. Estructuralmente, el aspecto del núcleo depende del momento del ciclo celular en el que se encuentre la célula: se habla de núcleo interfásico (mal llamado antiguamente núcleo en reposo) cuando la célula no está en fase de división, y de núcleo mitótico cuando se diferencían los cromosomas. 1.- Características del núcleo Esta presente en todas las células eucarióticas, excepto en los glóbulos rojos de los vertebrados superiores y en las células epidérmicas del estrato córneo superficial. Componentes. El núcleo consta de una envoltura nuclear (formada por una doble membrana) y una matriz nuclear o nucleoplasma, en cuyo seno se encuentran la cromatina (ADN y proteínas asociadas) y el nucléolo (donde se sintetiza el ARN ribosómico). Forma. Es muy variable (esférica, ovalada, polilobulada); depende del tipo de célula y del momento del ciclo en el que está. Tamaño. Puede oscilar entre las 5 y las 25 µm de diámetro, pero dentro de un mismo tipo de célula es constante. Generalmente, el tamaño del núcleo es proporcional al que tiene la célula, ocupando normalmente un 10°/o del volumen total de esta. Posición. La posición del núcleo es característica de cada célula. En las células embrionarias (meristemos, blastómeros) ocupa una posición central; en las adiposas, ocupadas en su mayoría por una gran gota de grasa, el núcleo queda lateralizado; en las secretoras se sitúa basalmente, quedando el resto de los orgánulos citoplásmicos entre él y el polo secretor. Número. Suele haber un núcleo por célula, pero existen algunas excepciones. Existen células anucleadas -como por ejemplo, los eritrocitos de los mamíferos- que han perdido el núcleo durante su proceso de diferenciación, y otras como los paramecios (protozoos ciliados), que de manera constante presentan dos núcleos: un macronúcleo y un micronúcleo, En los hepatocitos también es normal la situación binucleada, mientras que los osteoblastos y las' células musculares estriadas esqueléticas son plurinucleadas. La condición plurinucleada de algunas células puede originarse mediante dos mecanismos: - División sucesiva de un primitivo núcleo sin que ocurra la consiguiente división celular, dando origen a una célula denominada plasmodio. - Fusión de varias células uninucleadas, llamándose sincitio a la resultante. 2.- Envoltura nuclear La envoltura nuclear representa una compleja organización en la frontera entre el núcleo y el citoplasma de una célula eucariótica. Con el microscopio óptico, la envoltura nuclear se observa solo como un "límite" entre el citoplasma y el núcleo, pero con el microscopio electrónico se aprecia que, en realidad, es una doble membrana con un espacio intermembranoso. • La membrana nuclear externa tiene una anchura de 7 a 8 nm, y al microscopio electrónico muestra una ultraestructura trilaminar. Sobre su cara externa o citoplásmica presenta ribosomas adosados. Esta membrana suele estar unida a la del retículo endoplásmico, sea liso o rugoso. • El espacio perinuclear o intermembranoso comprendido entre las dos membranas tiene una anchura de 10 a 20 nm, aunque en algunos lugares puede presentar dilataciones de hasta 70 nm. Se encuentra en continuidad con el espacio reticular. • La membrana nuclear interna presenta, asociado a ella y en la cara nucleoplásmica, un material electrodenso de naturaleza fibrilar denominado lámina fibrosa o corteza nuclear. Se trata de tres polipéptidos o láminas dispuestos en tres capas y con características semejantes a las de los filamentos intermedios del citoesqueleto. A la Corteza nuclear se le atribuyen funciones como servir de anclaje al material cromatínico y regular el crecimiento de la envoltura nudear. 3.- Poros nucleares En todos los núcleos, las dos membranas que forman la envoltura nuclear se fusionan en algunos lugares, dando origen a unas perforaciones circulares denominadas poros nucleares que, para cada tipo de célula, presentan el mismo diámetro. Se trata de estructuras dinámicas, capaces de formarse y desaparecer dependiendo del estado funcional de la propia célula. Son canales acuosos que regulan los intercambios de moléculas entre el núcleo y el citosol. Permiten la circulación libre de moléculas hidrosolubles, y en el caso de macromoléculas como el ARN o las proteínas, que no son hidrosolubles, regulan mecanismos de transporte activo. La cantidad de poros nucleares es muy variable. En los núcleos de eritrocitos de aves es de 2-4 poros/µm2 en la membrana nuclear de un ovocito puede haber hasta 60 poros/µm2 En general, las células que tienen mayor actividad transcripcional (hepatocitos. neuronas y fibras musculares) contienen un elevado número de poros, mientras que las de menor actividad poseen menos. Una célula de mamífero contiene, de media, unos 3000 poros nucleares. • Estructura del complejo de poro nuclear Cuando se utiliza el microscopio electrónico, se observa que los poros no son simples orificios en la envoltura nuclear sino estructuras complejas que se denominan complejo de poro. Un complejo de poro está constituido por un anillo o estructura cilíndrica que se proyecta tanto hacia la cara citoplásmica como a la nucleoplásmica, y que está constituido por ocho partículas proteicas de unos 20 nm de diámetro dispuestas en octógonos. Asociado a las partículas se encuentra un material denso, el diafragma, que disminuye la luz del poro dejando libre solo unos 10 nm. En ocasiónes se ha identificado en el centro del poro un gránulo central, que en realidad se corresponde a ribosomas recién formados o sustancias que van de un lado a otro de la membrana nuclear. Se han descrito fibrillas proteicas que se extienden a ambos lados del poro. 4.- La cromatina. En el núcleo de las células eucarióticas, el ADN está asociado a proteínas formando una estructura empaquetada y compacta denominada cromatina, que representa el genoma de las células eucarióticas. Debe su nombre (cromatina, del griego chroma, coloreado) a la fácilidad con la que se tiñe con los colorantes básicos utilizados en microscopía óptica. Características La cromatina consta de ADN y proteínas. Las proteínas pueden ser de dos tipos: • Historias. Las histonas son proteínas muy básicas, debido a la abundante presencia de los aminoácidos -cargados positivamente- arginina y lisina. Se han descrito cinco clases de histonas: H1, N2A, H2b, H3 y H4, todas ellas de bajo peso molecular. • No histonas. En contraste con el escaso número de proteínas histónicas, son muy numerosas las proteínas no histónicas que se han aislado de la crornatina. Aproximadamente, la mitad corresponden a enzimas implicadas en la replicación, la transcripción y la regulación del ADN. Tipos de cromatina Eucromatina. Es la más abundante en la interfase y se corresponde con la cromatina menos compactada, ocupando la mayor parte de los llamados espacios intercromatínicos. Heterocromatina. Es la cromatina de mayor grado de compactación y aparece en la interfase como agrupaciones condensadas teñidas: los cromocentros. Ultraestructura La observación de la cromatina al microscopio electrónico revela una constitución fibrilar. Se trata de una serie de fibras adosadas unas a otras en forma de espiral, que reciben el nombre de fibras cromatínicas o nucleosómicas de 30 nm. Si se somete la cromatina a tratamientos de descondensación, cada fibra cromatínica aislada presenta el aspecto de un "collar de cuentas". A cada "cuenta" -con forma esférica, discoidal o ligeramente cilindrica-, Dudet y sus colaboradores (1975.) le dieron el nombre de nucleosoma. Estos nucleosomas tienen un diámetro de 10 nm y están relacionadas entre si por una fibrilla de 2 µm de diámetro, que se corresponde con el espesor ,de una doble hélice de ADN. Cada nucleosoma consta de un núcleo o platisoma y de un filamento de ADN que lo rodea; cada núcleo está formado por un octámero de histonas (dos moléculas de H2A, dos moléculas de H2B dos moléculas de H3 y otras dos moléculas de H4). La histona H1 no forma parte del nucleosoma, sino que se une a los segmentos de ADN que relacionan los nucleosomas. En la actualidad, se admite que la fibra de cromatina tiene una estructura plegada en forma de solenoide con distintos grados de espiralización. En la espiralización de primer grado, las fibras cromatínicas se compactan hasta presentar el diámetro de 30 nm, localizándose la histona H1 uniendo los nucleosomas en la cara interna del solenoide. Esta fibra puede sufrir una espiralización de segundo grado con un djámetro de 300 nm, y as sucesivamente hasta llegar a la "superespiralización" en el momento de iniciar la mitosis, en el que la cromatina se compacta para formar los cromosomas. 5.- Nucleoplasma También llamado carioplasma o matriz nuclear. Es una matriz semifluida situada en el interior riel núcleo, que contiene tanto el material cromatínico (ADN y proteínas cromosomales) como el no cromatínico (proteínas). Consta de varios componentes: • Los granulos de intercromatina miden entre 20 y 28 nm de diámetro, y contienen partículas de ribonucleoproteína y diversas enzimas: ATPasa, GTPasa, B-glicerofosfatasa y pirofosfatasa. Estos gránulos se encuentran diseminados por todo el núcleo. • Los granulos de pericromatina miden entre 30 y 50 nm y se localizan en la periferia de la cromatina. Están formados por fibrillas densamente empaquetadas de ARNr de bajo peso molecular. • Las partículas de ribonucleoproteína nucleares pequeñas. 6.- Nucléolo Fue descubierto por Fontana (1781) como una estructura constante en el interior del núcleo celular. Generalmente, hay uno por núcleo, pero no son raras las células en las que se observan dos o más, como por ejemplo, en algunas neuronas o ciertos hepatocitos. Es un orgánulo más o menos redondeado, muy refringente, basófilo debido a su alto contenido en ARN y proteínas, y, generalmente, localizado próximo a la envoltura nuclear. El tamaño del nucléolo está relacionado con el grado de actividad celular. Es mayor en tas células que presentan una.gran actividad de síntesis de proteínas, llegando a ocupar hasta el 25% del volumen nuclear. Observado con el microscopio óptico y utilizando técnicas de tinción argénticas, se pone de manifiesto cómo algunos nucléolos presentan un componente filamentoso muy replegado sobre sí mismo, denominado nucleolonema, que se encuentra embutido en un componente amorfo o parte amorfa, El nucléolo solo se observa con el microscopio óptico durante el período interfásico, ya que durante la mitosis desaparece al mismo tiempo que los cromosomas van alcanzando su máximo nivel de compactación. Aparece de nuevo, posteriormente, durante la telofase. • Tipos de nucléolos: La observación detallada de los nucléolos con el microscopio electrónico permite distinguir varios tipos según se encuentren distribuidos sus componentes granulares (ribo-nucleoproteínas): Nucléolos compactos u homogéneos. Nucléolos heterogéneos o reticulados, que son los más abundantes. Nucléolos anillados, en los que el ARN es periférico y el ADN es central. • Funciones del nucléolo. En el nucléolo se realiza la síntesis del ARNr y el procesado y empaquetamiento de subunidades ribosomales, que posteriormente son exportadas al citosol. Es indispensable para el desarrollo normal de la mitosis, si bien desaparece durante la misma. • Ultraestructura del nucléolo. La microscopía electrónica ha demostrado que no existe membrana alguna que delimite el nucléolo, y ha confirmado la existencia de los dos componentes observados con el microscopio óptico: - Un componente estrictamente nucleolar en el Que se distinguen dos zonas: la zona granular, que corresponde a subunidades ribosomales en proceso de maduración, y la zona fibrilar, que corresponde a moléculas de ARNr asociadas a proteínas. - Un componente nuclear o cromatina asociada. Las fibrillas cromatínicas pueden encontrarse como cromatina perinucleolar rodeando al nucléolo, o como cromatina intranucleolar en el interior del núcleo. Ambos tipos de cromatina están en continuidad. Estas fibrillas de ADN se corresponden con unas zonas concretas de los cromosomas -denominadas regiones organizadoras nucleolares o NOR-, dado que el ADN de esta zona es el portador de los genes que codifican el nucléolo. En humanos, las NOR se encuentran en los cromosomas 13, 14, 7.- Los cromosomas Los cromosomas representan la máxima compactación de la cromatina. Fueron descritos, por primera vez, por Hoffmeister en 1848 en las células madre de los granos de polen; aunque el nombre de cromosoma fue dado por Waldeyer en 1888. Cada molécula de ADN es hasta 50 000 veces más corta que en su forma extendida. Esta máxima condensación es la que permite el reparto del material genético entre las células hijas. Según las hipótesis actuales, los cromosomas estarían formados por varios dominios estructurales en forma de bucle (bucles de cromatina), que se extenderían a partir de un eje principal (formado por proteínas no histónicas) alrededor del cual se dispondría la fibra nucleosómica de manera espiralada, dando lugar a los sucesivos bucles, • Estructura del cromosoma metafásico El cromosoma metafásico es el más estudiado y del que mejor se conoce su estructura. Está constituido por dos cromátidas paralelas entre sí, resultado de la duplicación del material genético, y separadas -excepto en el ámbito del centrómero- por donde permanecen unidas En cada cromosoma se identifican las siguientes estructuras: • El centrómero o constricción primaria. Divide al cromosoma en dos brazos, que pueden ser del mismo o de diferente tamaño; ocupa una posición variable, pero fija para cada uno de ellos a lo largo del cromosoma. Los centrómeros contienen heterocromatina constitutiva, es decir, cromatina compactada y genéticamente inactiva en todas las células A ambos lados del centrómero, y sobre cada una de las dos cromátidas, se localiza una estructura de naturaleza proteica denominada cinetocoro, que constituye los puntos desde los cuales polimerizan los microtúbulos que intervienen en la separación de los cromosomas durante la anafase de la mitosis y de la meiosis. Por cada centrómero aparecen dos cinetocoros. • Las constricciones secundarias u organizadores nucleolares. Son zonas más estrechas identificables en los brazos, y que están relacionadas con la formación .del nucléolo al final de cada mitosis. • Los telómeros. Son estructuras protectoras, situadas en cada uno de los extremos del cromosoma eucariótico que evitan que se pierda información de los extremos en cada ciclo de replicación. En el ser humano, los telómeros contienen la secuencia TTAGGG repetida miles de veces. Los telómeros desempeñan importantes funciones: son esenciales para la duplicación del cromosoma. protegen a los cromosomas contra las nucleasas (enzimas que digieren el ADN), evitan que los extremos de los cromosomas se fusionen entre sí, y facilitan la interacción entre los extremos y la envoltura nuclear. En ocasiones, a uno de los extremos se le une un fragmento de ADN denominado satélite, que consiste en un cuerpo más o.menos redondeado, no más ancho que el brazo. unido al extremo por un filamento fino. • Las bandas son segmentos de cromatina que se colorean con diferente intensidad y que permiten una identificación inequívoca de los cromosomas mediante el llamado método de patrón de bandas. Este método se basa en le utilización de diversas técnicas de coloración tras digestión enzimática o desnaturalización por el calor. A estas bandas coloreadas se las ha llamado C, G, Q y R, y permiten identificar los cromosomas homólogos. • Tipos de cromosomas Cuando se estudian los cromosomas, resulta muy útil utilizar dos índices de proporcionalidad que relacionan la longitud total, la longitud del brazo corto y la longitud del brazo largo del cromosoma. • Índice de proporcionalidad de brazos (i.p.b.). Indica la relación que existe entre la longitud del brazo corto y el brazo largo de un mismo cromosoma. • Índice de proporcionalidad centromérica (i.p.c.). Indica la relación entre la longitud del brazo corto y la longitud total del cromosoma. En función de la posición que ocupe el centrómero y el valor de estos índices de proporcionalidad, se distinguen cuatro tipos de cromosomas: •N ú mero de cromosomas El número de cromosomas diferentes (n) de una determinada célula es una constante para todas las que pertenecen a un mismo organismo. La mayoría de los organismos, tanto animales como vegetales, son diploides (2n), es decir, tienen en sus células dos juegos de cromosomas, uno heredado del padre y otro heredado de la madre. Los cromosomas forman parejas de homólogos que contienen información genética para los mismos caracteres. En estos organismos, sus células reproductoras o gametos, tanto el óvulo como el espermatozoide, solo presentan un juego de cromosomas. Son, por tanto, células haploides (n). También existen organismos en los que todas sus células son haploides; por ejemplo, algunas algas y la fase gametofítica de los heléchos y musgos. Además, existen organismos que tienen en sus células más de dos juegos de cromosomas. A los que tienen tres se les denomina triploides (3n); a los que tienen cuatro, tetraploides (4n), y a los que tienen más, poliploides. El número de cromosomas no guarda relación alguna con el nivel evolutivo alcanzado por la especie; la especie humana cuenta con 46 cromosomas, mientras que algunos protoctistas llegan a tener más de 300. Al conjunto de todos los cromosomas de una célula, representados fotomicrográficamente, se le denomina cariotipo. Dentro del cariotipo se distinguen dos tipos de cromosomas: • Los cromosomas somáticos o autosomas, comunes: en los dos sexos de la misma especie e implicados en desarrollar las características del soma o del cuerpo. • Los cromosomas sexuales o gonosomas, responsables por número, presencia o ausencia, de la determinación del sexo. Los cromosomas sexuales en humanos son el X y el Y, de menor tamaño.