Potencial ecológico de frutales nativos del - CGSpace
Anuncio

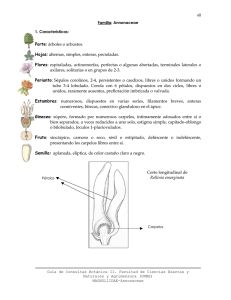
Potencial ecológico de frutales nativos del neotrópico, aguacate y anonas, en la diversificación de los paisajes cafetales en América Central como estrategia de adaptación al Cambio Climático Autor: Nekane Castillo Tutor: Maarten van Zonneveld Informe técnico Año de publicación: 2015 Citación correcta: Castillo N, van Zonneveld M. 2015. Potencial ecológico de frutales nativos del neotrópico, aguacate y anonas, en la diversificación de los paisajes cafetales en América Central como estrategia de adaptación al Cambio Climático: Informe Técnico. Programa de investigación de CGIAR en Cambio Climático, Agricultura y Seguridad Alimentaria (CCAFS). Copenhague, Dinamarca. Disponible en: www.ccafs.cgiar.org El Programa de Investigación de CGIAR en Cambio Climático, Agricultura y Seguridad Alimentaria (CCAFS) es una alianza estratégica entre el Consorcio CGIAR y Future Earth, liderado por el Centro de Investigación en Agricultura Tropical (CIAT). El programa es apoyado por los donantes del Fondo CGIAR, La Agencia de Cooperación al Desarrollo de Dinamarca (DANIDA), El Gobierno de Australia (ACIAR), Irish Aid, Environment Canadá, el Ministerio de Asuntos Exteriores para los Países Bajos, la Agencia Suiza para Desarrollo y Cooperación (SDC), El Instituto de Investigación Científica Tropical (IICT), UK Aid, el Gobierno de Rusia, la Unión Europea (EU), y el Ministerio de Relaciones Exteriores y Comercio de Nueva Zelanda, con apoyo técnico del Fondo Internacional para el Desarrollo Agrícola (Por sus siglas en ingles IFAD). Contacto: CCAFS Coordinating Unit - Faculty of Science, Department of Plant and Environmental Sciences, University of Copenhagen, Rolighedsvej 21, DK-1958 Frederiksberg C, Denmark. Tel: +45 35331046; Email: ccafs@cgiar.org Licencia de Creative Commons Este documento de trabajo es autorizado por la licencia Atribución-NoComercial-SinDerivadas 3.0 Unported. Los artículos que aparecen en esta publicación pueden citarse y reproducirse siempre que se reconozca la fuente. Ningún uso de esta publicación puede ser para reventa u otros fines comerciales. © 2015 Programa de Investigación de CGIAR en Cambio Climático, Agricultura y Seguridad Alimentaria (CCAFS) DESCARGO DE RESPONSABILIDAD: Este informe técnico ha sido preparado como un producto para el proyecto insignia: Agricultura Sostenible Adaptada al Clima bajo el programa CCAFS y no ha sido revisado por pares. Cualquier opinión expresada en este documento es la del (los) autor(es) y no refleja necesariamente las políticas u opiniones de CCAFS, los organismos donantes o socios. Todas las imágenes son propiedad exclusiva de su autor y no pueden ser utilizadas para cualquier propósito sin el permiso por escrito del mismo. La designación geográfica empleada y la presentación del material en esta publicación no implican la expresión de ninguna opinión por parte de CCAFS sobre la condición jurídica de países, territorios, ciudades o zonas, o de sus autoridades, ni respecto de la delimitación de sus fronteras o límites. 1 Resumen Aguacate (Persea) y anonas (Annona) están entre los géneros de frutales más importantes nativos del neotrópico. Sus productos tienen un potencial alto para mercados nacionales y son una fuente nutricional promisoria en sistemas de alimentación local. Con el presente trabajo se espera identificar el potencial ecológico de aguacate y anonas para la diversificación de los paisajes cafetales en American Central para mejorar la seguridad alimentaria y nutricional de familias caficulturas, y como posibles cultivos alternativos para generar ingresos. Secundariamente, y para ello, identificamos el área de distribución potencial actual y futura de las especies cultivadas y parientes de los géneros Persea y Annona en América Latina y evaluamos su estado de conservación y diversidad interespecífica. Las hipótesis que planteamos son: 1) los frutales neotropicales son ecológicamente aptos para la diversificación de los paisajes cafetales en América Central, 2) el Cambio Climático tiene un impacto negativo en las especies de frutales seleccionadas, y 3) los parientes de estos frutales pueden servir como recurso para mejoramiento de los mismos ante el Cambio Climático. Se modeló la distribución potencial de los frutales y sus parientes silvestres aplicando el Software Maxent bajo condiciones climáticas actuales y proyecciones climáticas para las 2050s considerando dos escenarios de emisión del IPCC. Los áreas de distribución potencial actual y futura obtenidos se sobrepusieron sobre el área de producción actual de café en América Central para saber cuáles frutales tenían mayor potencial para diversificación en estos areas. Contrariamente a la hipótesis que planteábamos, los resultados obtenidos sugieren que los parientes de los cultivos de aguacate no son tan prioritarios para mejoramiento de agucates a nuevas condiciones climaticas como lo es la diversidad genetica del propio cultigen P. americana Mill. Asimismo, y en base a los análisis realizados, se puede decir que entre las especies evaluadas, la especie con mayor potencial ecológico para la diversificación de los paisajes cafetales es la guanabana (A. muricata L.) 2 1. Introducción Los efectos adversos que el Cambio Climático está ocasionando sobre los cultivos agrícolas son evidentes en todo el mundo (Calzadilla, et al., 2014; Mendelsohn, 2014; Vargas-Amelin & Pindado, 2014). Se espera que los impactos sean diversos y heterogéneos afectando tanto a la calidad como cantidad de agua, incluyendo un aumento considerable en la demanda de agua para riego, acortamiento de ciclos vegetativos, aumento de plagas y especies exóticas, riesgo creciente de olas de calor o inundaciones, repercusiones directas en la calidad y producción agrícola (EEA, 2012), y por tanto, amenazando la seguridad alimentaria (Jones & Thornton, 2003, Bandara & Cai, 2014). En este sentido, los paisajes cafetales no son una excepción y también se están viendo gravemente afectados (Glenn et al., 2013; Laderach et al., 2013), observándose una disminución en la producción del café. No obstante, su demanda sigue en aumento, lo que ha llevado a la búsqueda de nuevas soluciones con el fin de evitar que los cultivos tengan que desplazarse a otras zonas. Ciertamente, se está desarrollando nuevas variedades, pero especialmente, se está impulsando el establecimiento de sombríos o coberturas vegetales en los cafetales (Borkhataria et al., 2012; Nonato de Souza et al., 2012). En efecto, son habituales en los cultivos especies de leguminosas, cacaos, bananos, así como frutales nativos del nuevo mundo o neotropicales, siendo estos últimos los que más beneficios aportan: Son fuente de seguridad alimentaria (Kehlenbeck et al., 2013; Bacon et al., 2014), lo que es especialmente importante en los meses con falta de ingresos y escasez de producción del cultivo de café (junio-agosto) (Bacon et al., 2014), periodo que se espera que se alargue con el Cambio Climático. Por ejemplo, cabe destacar que Annona muricata L. provee de frutos todo el año (Cordero & Boshier, 2003). Poseen un altor valor nutricional (Kehlenbeck et al., 2013). Han experimentado un fuerte crecimiento en las exportaciones como consecuencia del aumento de la demanda en otros países (Regmi, 2001; FAO, 2003), y se espera que su demanda siga creciendo (Scheldeman et al., 2007), con lo que constituyen un mercado creciente. La creación de estos sistemas agroforestales regula la temperatura y mejora las condiciones edáficas, aumenta la biodiversidad y protege a los cultivos de plagas y enfermedades, entre otros servicios ecosistémicos (Somarriba et al., 2004; Borkhataria et al., 2012). Lo que ocurre es que los frutales neotropicales tampoco están exentos de problemas ante el Cambio Climático y es ante esta situación donde los parientes de 3 los cultivos cobran importancia. Los parientes de los cultivos son especies estrechamente ligadas a los cultivos que juegan un papel esencial como fuente de recursos genéticos para la mejora de los cultivos y adaptación de los mismos a condiciones abióticas y bióticas cambiantes (Cordero & Boshier, 2003; Hajjar & Hodgkin, 2007). Por otro lado, presentan un elevado valor comercial ya que existen especies con características muy similares a las comercializadas que están siendo poco explotadas o que son subutilizadas (Bost, 2009; Barbalho et al., 2012); y por último, desde un punto de vista más conservacionista, son especies muy valoradas. Sin embargo, las poblaciones naturales de los parientes silvestres también están en riesgo creciente (Maxted, 2003; Kell et al., 2012), lo que unido a la disminución de la heterogeneidad genética de las variedades cultivadas, supone un aumento de la vulnerabilidad de los cultivos al estrés. Es por ello que resulta fundamental evaluar su estado de conservación en un contexto de Cambio Climático y así, poder valorar su capacidad para diversificar los paisajes cafetales. Con el presente trabajo se espera identificar los frutales neotropicales con mayor potencial ecológico para la diversificación de los paisajes cafetales. Secundariamente, y para ello, identificamos el área de distribución potencial actual y futura de las especies cultivadas y parientes de los géneros Persea y Annona en América Latina y evaluamos su estado de conservación y diversidad interespecífica. Las hipótesis que planteamos son: 1) los frutales neotropicales son ecológicamente aptos para la diversificación de los paisajes cafetales en América Central, 2) el Cambio Climático tiene un impacto negativo en las especies de frutales seleccionadas, y 3) los parientes de estos frutales pueden servir como recurso para mejoramiento de los mismos ante el Cambio Climático. 4 2. Metodología 2.1. Especies seleccionadas y datos de presencia Se seleccionaron un total de 38 especies frutales arbóreas de cultivos y parientes de los cultivos. Las especies seleccionadas se corresponden a los géneros de Persea y Annona que fueron priorizados sobre los demás según los siguientes criterios: a) plantas cultivadas con un elevado valor socioeconómico (frutales), b) nativas de Centroamérica, c) de sistemas agroforestales, e c) incluidas en la Estrategia Hemisférica racional de Conservación de los recursos fitogenéticos para la alimentación y la agricultura en las Américas (Anexo 1). Para la selección de las especies de cultivos y sus parientes se consultó el Inventario de Parientes Silvestres de los Cultivos (Crop Wild Relatives and Climate Change, 2013), el Atlas de los Parientes Silvestres de los Cultivos de Guatemala (Azurdia et al., 2011) como principal centro de domesticación de plantas de América Central, el Atlas de los Parientes Silvestres de Bolivia (Hunter & Heywood, 2011) como centro de diversidad en América del Sur, así como Germplasm Resources Information Network (USDA et al., 2015) y la opinión de diversos expertos (Anexo 2). Los datos de presencia de las especies se obtuvieron a partir de Global Biodiversity Information Facility Portal (GBIF, 2014), diversas bases de datos (Collecting Missions Database; Naturalista-CONABIO, 2014; speciesLink, 2014; Tropicos, 2014), herbarios nacionales (Herbario Forestal MOL; Herbarium WU, 2014; Museo Nacional de Costa Rica, 2014), jardines botánicos (The NY Botanical Garden, 2014), tesis de Maestrías de CATIE, artículos publicados en revistas especializadas (Quesada, 2004; Tacán, 2007; van Zonneveld et al., 2012), etc. Tras eliminar los puntos duplicados y una vez georreferenciados los datos de pasaporte descriptivos de las especies con menos observaciones en base a la descripción de la localidad mediante Google Earth, GeoNames y otros, se recopilaron un total de 9220 citas históricas a diferente resolución y extensión América Latina (30 países). Sin embargo, pese a que se han creado modelos predictivos con sólo 5 puntos de presencia utilizando MaxEnt (Phillips et al., 2006), para que las predicciones sean robustas sólo se incluyeron las especies con ≥20 puntos de presencia (Hernandez et al., 2007; MAPFORGEN, 2013), resultando en 7 especies (de 10 iniciales) para el género Persea y 27 especies (de 28 iniciales) para el género Annona. Control de calidad Las fronteras políticas se representaron a tres niveles de unidades administrativas. Las coordenadas de los respectivos puntos de presencia de las especies se revisaron a nivel administrativo 1 con un área de amortiguación de 10arcmin. Según la denominación de cada país, el nivel administrativo 1 incluyó: Departamento (Bolivia, Colombia, El Salvador, Haití, Honduras, Paraguay, Perú, Guatemala, Nicaragua), Estado (Brasil, México, Venezuela), Provincia (Argentina, 5 Costa Rica, Cuba, Ecuador, Panamá, República Dominicana), Distrito (Bahamas, Belice, Guayana Francesa, Islas Vírgenes de los EEUU, Surinam) y Municipalidad (Puerto Rico). Además, se identificaron los puntos con valores atípicos con base a variables bioclimáticas derivadas de Worldclim (Hijmans et al., 2005) al percentil 0.025, previo análisis de correlación, con el método Reverse Jacknife (para conjuntos de datos grandes) y con el método 1.5IQR (para pocas observaciones) (Chapman, 2005). La elección de un método u otro se realizó en función del número de puntos atípicos obtenidos, seleccionando siempre aquel método menos estricto con el fin de mantener el mayor número de puntos. Para ambos géneros establecimos un valor umbral del 25% de variables ambientales para considerar un punto atípico. Como resultado de estos análisis, 97 puntos fueron descartados y Annona macroprophyllata Donn. Sm. fue eliminada de los análisis posteriores por no alcanzar ≥20 observaciones. Por tanto, tras el control de calidad, se obtuvieron un total de 9091 puntos georreferenciados (Tabla 1). 6 Tabla 1. Especies seleccionadas y número de observaciones. Especie 1 Persea americana Mill. 2 P. caerulea (Ruiz & Pav.) Mez nº obs. 1546 717 3 P. donnell-smithii Mez 67 4 P. liebmannii Mez 160 5 P. rigens C.K.Allen 102 6 P. schiedeana Nees 7 P. vesticula Standl. & Steyerm. 198 66 1 Annona amazonica R.E.Fr 99 2 A. ambotay Aubl. A. cordifolia (Szyszył.) Poepp. ex Maas & 3 Westra 205 4 A. coriacea Mart. 583 5 A. cornifolia A.St.-Hil. 137 6 A. cherimola Mill. 1122 7 A. dioica A.St.-Hil. 283 8 A. excellens R.E.Fr. 9 A. foetida Mart. 10 A. glabra L. 59 112 315 11 A. liebmanniana Baill. 27 12 A. longiflora S.Watson 32 13 A. hypoglauca Mart. 337 14 A. macrocalyx R.E.Fr. 25 16 A. montana Macfad. 377 17 A. monticola Mart. 18 A. muricata L. 19 A. nutans (R.E.Fr.) R.E.Fr. 165 445 141 20 A. paludosa Aubl. 21 A.purpurea Moc. & Sessé ex Dunal 22 A. reticulata L. 84 178 635 23 24 25 26 27 29 29 192 304 284 A. scandens Diels ex Pilg. A. scleroderma Saff. A. sericea Dunal A. squamosa L. A. tomentosa R.E.Fr. Total: 36 9091 7 2.2. Datos ambientales Para evitar un sobreajuste del modelo por multicolinearidad (Dormann et al., 2013), se realizó un test de correlación (Coeficiente de correlación de Pearson, r>0,70) para cada género. Se contabilizó el número de correlaciones de cada variable ambiental y se seleccionó aquella con un mayor valor y número de correlaciones, descartando las variables correlacionadas a ésta y continuando del mismo modo con la siguiente variable ambiental con un mayor valor y número de correlaciones. En total, fueron seleccionadas 6 variables ambientales para el género Persea (Tabla 1) y 8 para el género Annona (Tabla 2a, 2b). Tabla 2a. Variables ambientales empleadas para la modelización de las especies del género Persea. Código o nombre de la variable, descripción, unidades, resolución y tipo (continua o categórica). Código Descripción Unidad Resolución Tipo BIO 1 Tª media anual ºC x 10 2,5arc-min Continua BIO 2 Rango medio diurno ºC x 10 2,5arc-min Continua BIO 4 Estacionalidad térmica (SD x100) 2,5arc-min Continua BIO 12 Precipitación anual mm 2,5arc-min Continua BIO 17 Precipitación del cuatrimestre más seco mm 2,5arc-min Continua BIO 18 Precipitación del cuatrimestre más cálido mm 2,5arc-min Continua Tabla 2b. Variables ambientales empleadas para la modelización de las especies del género Annona. Código o nombre de la variable, descripción, unidades, resolución y tipo (continua o categórica). Código Descripción Unidad Resolución Tipo BIO 1 Tª media anual ºC x 10 2,5arc-min Continua BIO 2 Rango medio diurno ºC x 10 2,5arc-min Continua BIO 4 Estacionalidad térmica (SD x100) 2,5arc-min Continua BIO 12 Precipitación anual mm 2,5arc-min Continua BIO 14 Precipitación del mes más seco mm 2,5arc-min Continua BIO 15 Estacionalidad precipitaciones (C.V.) mm 2,5arc-min Continua BIO 18 Precipitación del cuatrimestre más cálido mm 2,5arc-min Continua BIO 19 Precipitación del cuatrimestre más frío mm 2,5arc-min Continua Las capas climáticas, tanto actuales como futuras, fueron obtenidas a partir de Worldclim (Hijmans et al., 2005) a una resolución de 2,5arc-min (≈4,5km en el Ecuador). En condiciones futuras, se utilizaron todos los modelos climáticos globales (GCMs) disponibles con los escenarios RCP4.5 ([CO2] intermedia) y RCP8.5 ([CO2] alta) 8 de emisión de gases de efecto invernadero (Representative Concentration Pathways) (IPPC, 2014) para el año 2050. Con el software R se georreferenciaron las capas ambientales a la misma proyección y se transformaron al formato .ascii, requerido por MaxEnt. 2.3. Análisis de distribución de especies El área de distribución de las especies seleccionadas se modelizó con la técnica de Máxima Entropía (MaxEnt; Phillips et al., 2006) ya que se ha demostrado que posee un poder de predicción mayor en comparación con otros métodos (Elith et al., 2006) y cuando los tamaños de muestra son pequeños (Hernández et al., 2006; Wisz, 2008; Elith et al., 2011). Este método estima la probabilidad de presencia de la especie buscando la distribución de probabilidad lo más uniforme posible (Máxima Entropía), bajo la restricción de que el valor esperado de cada capa de información deba acercarse a su media empírica (Entropía) (Phillips et al., 2004). Ajuste y evaluación de los modelos Las especies de los géneros Annona y Persea fueron modelizadas a extensión América Latina. Se mantuvo la configuración y formato de salida (logístico) por defecto del programa, validado en una amplia gama de especies, condiciones ambientales, nº de muestras y sesgos (Phillips y Dudík, 2008). El 80% de las presencias se utilizó para su calibración y el 20% restante se reservó para su validación. En caso de existir más de una presencia en una cuadrícula obtenida por distintos colectores, se mantuvo el número total de puntos con el propósito de tener en cuenta la abundancia de las especies. La evaluación de los modelos se realizó mediante la curva ROC (Receiver Operating Characteristic), que describe la relación entre la proporción de presencias correctamente predichas (sensibilidad; positivos reales) y la proporción de ausencias predichas como presencias (1-especificidad; falsos positivos o error de comisión). La curva ROC proporciona el valor del área bajo la curva AUC, el cual mide la potencia del modelo. El rango de valores AUC obtenido se interpretó según Swets (1988) como: excelente (AUC>0,90), bueno (0,80>AUC<0,90), aceptable (0,70>AUC<0,80), pobre (0,60>AUC<0,70) y malo (AUC<0,60). Riqueza de especies observada y modelada La riqueza observada de las especies para ambos géneros se calculó mediante el Análisis Point to Grid en DIVA-GIS (Scheldeman & van Zonneveld, 2010). Por su parte, para poder calcular la riqueza modelada de las especies, los modelos continuos generados en MaxEnt fueron convertidos a modelos binarios (presencia/ausencia), de forma que se identificaron las áreas con máxima probabilidad de presencia de las especies. Para ello existen diversos métodos que establecen un valor límite o umbral de corte (Valverde y Lobo, 2007; Lobo et al., 2008); en nuestro caso, seleccionamos 9 el valor de probabilidad con el máximo valor de sensibilidad y especificidad (Liu et al., 2005) y sumamos los mapas obtenidos. Impacto del Cambio Climático El efecto del Cambio Climático en las especies seleccionadas se evaluó a través de la superposición de las proyecciones obtenidas del promedio de los modelos climáticos globales (GCMs) en los escenarios RCP4.5 y RCP8.5 de emisión de gases de efecto invernadero y los modelos binarios de máxima probabilidad de presencia de las especies. Solapamiento de las zonas cafetales Las capas de las celdas con máxima probabilidad de presencia de las especies más relevantes y las áreas con ≥1% de superficie de café (Monfreda et al., 2008) se superpusieron entre si y se calculó el porcentaje de celdas con máxima probabilidad de presencia que se encuentran en zonas cafetaleras en América Central. Requerimientos climáticos y adaptabilidad Se extrajeron los datos climáticos de los puntos de presencia de cada especie y se evaluó el nicho climático del género Persea y de aquellas especies más relevantes del género Annona en función de las variables climáticas más representativas ecológicamente: temperatura y precipitación media anual. 10 3. Resultados y discusión Ajuste y evaluación de los modelos Según la clasificación de Swets (1988), consideramos como buenos o excelentes todos los modelos actuales obtenidos. Riqueza de especies observada y modelada Tanto para el género Persea como para el género Annona, la mayor riqueza de especies observada se concentra en América Central y sureste de Brasil (Fig.1), si bien se puede decir que la distribución de las especies del género Annona es más homogénea y engloba todo el norte de América del Sur y América Central (Fig. 1). En cambio, si se tienen en cuenta los mapas de riqueza de especies modelada obtenidos a partir de las preferencias climáticas, se observa que además de las zonas mencionadas, en el caso del género Persea, los países de Chile, Bolivia y Perú también presentan una alta diversidad potencial, mientras que en el caso del género Annona, destacan Guyana Francesa, Suriname y Guayana (Fig. 2). Impacto del Cambio Climático El análisis del impacto del Cambio Climático sobre las especies seleccionadas muestra una clara reducción en las áreas climáticamente idóneas para el género Persea, siendo P. americana Mill. y P. caerulea (Ruiz & Pav.) Mez las especies más afectadas con hasta una pérdida del 7% (Tabla 3). Igualmente, A. cherimola Mill. y A. muricata L. también presentan una reducción del 2,4% y 4% de su área potencial actual, respectivamente (Tabla 3); no obstante, las nuevas presencias de A. muricata L. son mayores que las pérdidas, observándose un aumento y desplazamiento de su nicho ecológico (Fig. 3). 11 Fig. 1. Riqueza de especies observada de los géneros de a) Persea y b) Annona en América Latina a una resolución de 1º (en oscuro las áreas con riqueza más elevada). Fig. 2. Riqueza de especies modelada de los géneros de a) Persea y b) Annona en América Latina a una resolución de 2,5ˈ (en oscuro las áreas con riqueza más elevada). 12 Tabla 3. Impacto previsto de los escenarios RCP4.5 y RCP8.5 de emisiones de Gases de Efecto Invernadero en el área de distribución potencial (%) de las especies seleccionadas en América Latina (NP=Nuevas presencias; M=Se mantiene; NA=No Apto; PD=Presencias desaparecidas). RCP 4.5 RCP 8.5 NP M NA PD NP M NA PD Persea americana Mill. P. schiedeana Nees 0.5 0.7 9.0 12.6 83.5 81.2 7.0 5.5 0.6 0.8 8.0 11.7 83.5 81.1 8.0 6.4 P. caerulea (Ruiz & Pav.) Mez 1.5 8.3 82.9 7.4 2.4 7.3 81.9 8.4 P. donnell-smithii Mez 0.1 7.1 89.2 3.6 0.2 6.4 89.2 4.3 P. liebmannii Mez 0.6 4.4 90.1 4.9 0.5 3.7 90.1 5.6 P. rigens C.K.Allen P. vesticula Standl. & Steyerm. 0.1 0.0 1.1 1.8 97.8 97.1 1.1 1.1 0.1 0.0 0.9 1.5 97.8 97.1 1.3 1.4 A. cherimola Mill. 0.4 6.1 2.8 12.3 94.4 77.6 2.4 4.0 0.5 6.7 2.5 11.7 94.3 77.0 2.7 4.6 A. muricata L. Fig. 3. Efecto previsto de los escenarios a) RCP4.5 y b) RCP8.5 de emisiones de Gases de Efecto Invernadero en el área de distribución potencial de A. muricata L. Por lo tanto, estos resultados apoyan la hipótesis de que el Cambio Climático tiene un impacto negativo en los frutales seleccionados, salvo en el caso de A. muricata L. Solapamiento de las zonas cafetales 13 La Figura 4 es el resultado de la superposición de los mapas con máxima probabilidad de presencia de las especies más relevantes de los géneros Persea y Annona con las zonas cafetaleras en América Central. En todos los casos el área coincidente entre el paisaje cafetal y los mapas de distribución potencial de las especies seleccionadas supera el 24%, destacando A. muricata L. con cerca un 60% del área superpuesta. De modo que, se puede decir que se prueba la hipótesis de que los frutales neotropicales seleccionados son adecuados para la diversificación de los paisajes cafetales en América Central. a) b) c) d) Fig. 4. Superposición de los mapas con máxima probabilidad de presencia de P. americana Mill, b) P. schiedeana Nees, c) A. cherimola Mill y d) A. muricata L. con las áreas con ≥1% de superficie de café. 14 Requerimientos climáticos y adaptabilidad El análisis de distribución climática de las especies revela que P. americana Mill es la especie que mayor nicho ecológico presenta, englobando prácticamente todas las regiones ecogeográficas de las demás especies (Fig. 5a, 5b). Por su parte, se observa que A. cherimola Mill tolera peor las precipitaciones y se distribuye en áreas con temperaturas más suaves, mientras que A. muricata L. presenta una tendencia a zonas claramente más calurosas (Fig. 6). Fig. 5a. Nicho ecológico de P.americana L. y tres parientes silvestres en función de la envoltura climática. 15 Fig. 5b. . Nicho ecológico P. americana L. y tres parientes silvestres en función de la envoltura climática. Fig. 6. Nichos ecológicos de A. cherimola L. y A. muricata L. en función de la envoltura climática. 16 4. Discusión Los modelos de distribución de especies son una herramienta sencilla, especialmente útil en aquellos casos donde el conocimiento relativo a la distribución de una especie es escaso. Sin embargo, hay que tener en cuenta que la predicción de los modelos varía en función de la metodología empleada, como puntos mínimos necesarios (Hernandez et al., 2006; Pearson et al., 2007) y calidad de los mismos (Lobo, 2008), variables ambientales seleccionadas (Luoto et al., 2007; Peters et al., 2014), escala espacial (resolución y extensión) (Guisan y Thuiller, 2005; Song et al., 2013), etc. Es por ello que la predicción de los modelos generados puede ser inexacta y resulta por tanto necesario comprobar la presencia de las especies en estos lugares, añadiendo nuevas variables ambientales, como interacciones bióticas y capacidad de dispersión, y/o realizar muestreos en las zonas identificadas como propicias. No obstante, si bien son muchas las limitaciones, los mapas obtenidos muestran aquellos lugares con condiciones ambientales idóneas para las especies, permitiendo realizar una primera aproximación de las especies arbóreas con potencial para la diversificación de los paisajes cafetales. 5. Conclusiones Contrariamente a la hipótesis que planteábamos, los resultados obtenidos sugieren que los parientes de los cultivos del género Persea no son tan prioritarios como lo es el propio frutal P. americana Mill. Asimismo, y en base a los análisis realizados, se puede decir que entre las cuatros especies evaluadas, la especie con mayor potencial ecológico para la diversificación de los paisajes cafetales es A. muricata L. 17 6. Bibliografía Azurdia, C., Williams, K.A., Williams, D.E., Van Damme, V., Jarvis, A., Castaño, S.E. 2011. Atlas of Guatemalan Crop Wild Relatives. Disponible en: www.ars.usda.gov/ba/atlascwrguatemala United States Department of Agriculture/Agricultural Research Service (USDA/ARS), Bioversity International, International Center for Tropical Agriculture (CIAT) and the University of San Carlos in Guatemala (FAUSAC). Bacon, C.M. et al. 2014. Explaining the “hungry farmer paradox”: Smallholders and fair trade cooperatives navigate seasonality and change in Nicaragua’s corn and coffee markets. Global Environmental Change 25, 133-149. Bandara, J.S. & Cai, Y. 2014. The impact of climate change on food crop productivity, food prices and food security in South Asia. Economic Analysis and Policy 44 (4), 451-465. Barbalho, M. et al. 2012. Annona sp: Plants with Multiple Applications as Alternative Medicine - A Review. Current Bioactive Compounds 8 (3), 277-286. Borkhataria, R., Collazo, J.A., Groom, M.J., Jordan-Garcia, A. 2012. Shade-grown coffee in Puerto Rico: Opportunities to preserve biodiversity while reinvigorating a struggling agricultural commodity. Agriculture, Ecosystems & Environment 149, 164-170. Bost, J. B. 2009. Edible plants of the Chinantla, Oaxaca, Mexico with an emphasis on the participatory domestication prospects of Persea schiedeana. pp. 113. (Doctoral dissertation, University of Florida). Calzadilla, A., Zhu, T., Rehdanz, K., Tol, R.S.J., Ringler, C. 2014. Climate change and agriculture: Impacts and adaptation options in South Africa. Water Resources and Economics 5, 24-48. Chapman, A.D. 2005. Principles and methods of data cleaning – primary species and speciesoccurrence data, version 1.0. Report for the Global Biodiversity Information Facility, Copenhagen. Collecting Missions Database. www.bioversity.github.io/geosite/ Bioversity International. Disponible en: Cordero, J. & Boshier, D. (eds.). 2003. Árboles de Centroamérica: un manual para extensionistas. Bib. Orton IICA / CATIE. pp. 1079. Crop Wild Relatives and Climate www.cwrdiversity.org/checklist/ Change. 2014. Online resource. Disponible en: Dormann, C.F., Elith, J., Bacher, S., Buchmann, C., Carl, G., Carré, G., Marquéz, J.R.G., Gruber, B., Lafourcade, B., Leitão, P.J., Münkemüller, T., McClean, C., Osborne, P.E., Reineking, B., Schröder, B., Skidmore, A.K., Zurell, D., Lautenbach, S. 2013. Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36, 27-46. EEA, European Environment Agency. 2012. Climate Change Impacts and Vulnerability in Europe 2012. 304 pp. Elith, J., Graham, C.H., Anderson, R.P., Dudík, M., Ferrier, S., Guisan, A., Hijmans, R.J., Huettmann, F., Leathwick, J.R., Lehmann, A., Li, J., Lohmann, L.G., Loiselle, B.A., Manion, G., Moritz, C., Nakamura, M., Nakazawa, Y., McC.Overton, J., Peterson, A.T., Phillips, S.J., Richardson, K., Scachetti-Pereira, R., Schapire, R.E., Soberón, J., Williams, S., Wisz, M.S., Zimmermann, N.E. 2006. Novel methods improve prediction of species´distributions from ocurrence data. Ecography 29, 129-151. 18 Elith, J., Phillips, S.J., Hastie, T., Dudík, M., Chee, Y., Yates, C. J. 2011. A statistical explanation of Max.Ent for ecologist. Diversity and Distributions 17, 43-57. FAO. 2003. Medium-term prospects for Agricultural Commodities. Projections to the Year 2010. FAO Commodities and Trade Technical Paper 1. Food and Agriculture Organization of the United Nations, Rome. GBIF, 2014. Global Biodiversity Information Facility. Disponible en: www.gbif.org (revisado en diciembre 2014). Glenn, M., Kim, S.Y., Ramirez-Villegas, J., Laderach, P. 2013. Response of perennial horticultural crops to climate change. Horticultural Reviews 41, 47-130. Guisan, A. y Thuiller, W. 2005. Predicting species distribution: offering more than simple habitat models. Ecology Letters 8, 993-1009. Hajjar, R. & Hodgkin, T. 2007. The use of wild relatives in crop improvement: a survey of developments over the last 20 years. Euphytica 156, 1-13. Herbario Forestal MOL. Herbario de la Facultad de Ciencias Forestales de la Universidad Nacional Agraria-La Molina. Herbarium WU. Institute of Botany, University herbarium.univie.ac.at [consultado en diciembre 2014]. of Vienna. 2014. Disponible en: Hernandez, P.A., Graham, C.H., Master, L.L., Albert, D.L. 2006. The effect of simple size and species characteristics on performance of different species distribution modeling methods. Ecography 29, 773-785. Hijmans, R.J., Cameron, S.E., Parra, J.L., Jones, P.G., Jarvis, A. 2005. Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology 25, 19651978 [datos bioclimáticos disponibles en: www.worldclim.org] Hunter, D. & Heywood, V. (eds.). 2011. Crop wild relatives. A manual of In situ conservation. Bioversity International, Rome (Italy). IPPC. 2014. Fifth Assessment Report. Climate Change 2014. Synthesis Report. pp. 132. Jones, P.G. & Thornton, P.K. 2003. The potential impacts of climate change on maize production in Africa and Latin America in 2055. Global Environmental Change 13, 51-59. Kell, S.P., Max.ted, N., Bilz, M. 2012. European crop wild relative threat assessment: knowledge gained and lessons learnt. In: Max.ted, N., Dulloo, M.E., Ford-Lloyd, B.V., Frese, L.L., Iriondo, J.M., Pinheiro de Carvalho, M.A.A. (Eds.), Agrobiodiversity Conservation: Securing the Diversity of Crop Wild Relatives and Landraces. CAB International, Wallingford, pp. 218-242. Kehlenbeck, K. et al. 2013. Diversity of indigenous fruit trees and their contribution to nutrition and livelihoods in sub-Saharan Africa: Examples from Kenya and Cameroon. Diversifying Food and Diets: Using Agricultural Biodiversity to Improve Nutrition and Health, 257-269. Laderach, P., Haggar, J., Lau, C., Eitzinger, A., Ovalle, O., Baca, M., Jarvis, A., Lundy, M. 2013. Mesoamerican Coffee: Building A Climate Change Adaptation Strategy. CIAT Policy Brief No. 2. Centro Internacional de Agricultura Tropical (CIAT), Cali, Colombia. Liu, C., Berry, P.M., Dawson, T.P., Pearson, R.G. 2005. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 28, 385-393. 19 Lobo, J.M. 2008. More complex distribution models or more representative data. Biodiversity informatics 5, 14-19. Lobo, J.M., Jiménez-Valverde, A. Real, R. 2008. AUC: a misleading measure of the performance of predictive distribution models. Global Ecology and Biogeography 17, 145-151. Luoto, M., Virkkala, R., Heikkinen, R.K. 2007. The role of land cover in bioclimatic models depends on spatial resolution. Global Ecology and Biogeography 16, 34-42. MAPFORGEN. 2013. Atlas para la conservación de los recursos genéticos forestales. Bioversity International, INIA, LAFORGEN. Disponible en: http://www.mapforgen.org Maxted, N. 2003. Conserving the genetic resources of crop wild relatives in European protected areas. Biological Conservation 113, 411-417. Mendelsohn, R. 2014. The Impact of Climate Change on Agriculture in Asia. Journal of Integrative Agriculture 13 (4), 660-665. Monfreda, C., Ramankutty, N., & Foley, J.A. 2008. Farming the planet: 2. Geographic distribution of crop areas, yields, physiological types, and net primary production in the year 2000. Global biogeochemical cycles 22 (1). Museo Nacional de Costa Rica. 2014. Disponible en: www.museocostarica.go.cr [consultado en diciembre 2014]. NATIFRUT, Red Temática sobre frutales nativos americanos. Naturalista-CONABIO. 2014. Disponible en: naturalista.conabio.gob.mx [consultado en diciembre 2014]. Nonato de Souza, H., de Goede, R.G.M., Brussaard, L., Cardoso, I.M., Duarte, E.M.G., Fernandes, R.B.A., Gomes, L.C., Pulleman, M.M. 2012. Protective shade, tree diversity and soil properties in coffee agroforestry systems in the Atlantic Rainforest biome. Agriculture, Ecosystems & Environment 146 (1) 179-196. Pearson, R., Raxworthy, C.J., Nakamura, M., Peterson, A.T. 2007. Predicting species distribution from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. Journal of biogeography 34, 102-117. Peters, J., Waegeman, W., Van doninck, J., Ducheyne, E., Calvete, C., Lucientes, J., Verhoest, N.E.C., De Baets, B. 2014. Predicting spatio-temporal Culicoides imicola distributions in Spain based on environmental habitat characteristics and species dispersal. Ecological Informatics 22, 69-80. Phillips, S.J. y Dudík, M. 2008. Modeling of species distribution with MaxEnt: new extensions and a comprehensive evaluation. Ecography 31, 161-175. Phillips, S.J., Andersonb, R.P., Schapire, R.E. 2006. Maximun entropy modeling of species geographic distribution. Ecological Modelling 190, 231-259. Phillips, S.J., Dudík, M., Schapire, R.E. 2004. A max.imum entropy approach to species distribution modeling. En: Proceedings of the Twenty-First International Conference on Machine Learning, ACM Press, 655-662. Quesada, P. 2004. Inventario y caracterización de algunas especies de Annona en Costa Rica. Escuela de Ciencias Agrarias. Heredia, C.R. Rev. de Agricultura Tropical 34, 61-72. Regmi, A. 2001. Changing Structure of Global Food Consumption and Trade. Economic Research Service. U.S. Department of Agriculture, Agriculture and Trade Report. WRS-01-1. pp. 111. 20 Scheldeman, X. & van Zonneveld, M. 2010. Training Manual on Spatial Analysis of Plant Diversity and Distribution. Bioversity International, Rome, Italy. 179 pp. Scheldeman, X., Willemen, L., Coppens d’Eeckenbrugge, G., Romeijn-Peeters, E., Restrepo, M. T., Romero Motoche, J., Jiménez, D., Lobo, M., Medina, C. I., Reyes, C., Rodríguez, D., Ocampo, J. A., Van Damme, P., Goetgebeur, P. 2007. Distribution, diversity and environmental adaptation of highland papayas (Vasconcellea spp.) in tropical and subtropical America. Biodiversity Conservation 16, 1867-1884. Schmidhuber, J., Tubiello, F.N. 2007. Global food security under climate change. Proc. Natl. Acad. Sci. USA 104, 19703-19708. Somarriba, E., Harvey, C., Samper, M., Anthony, F., Gonzalez, J., Staver, C., Rice, R., 2004. Biodiversity in coffee plantations. In: Schroth, G., Fonseca, G., Harvey, C., Gascon, C., Vasconcelos, H., Izac, A.M.N. (Eds.), Biodiversity in Coffee Plantations. Island Press, Washington, DC, pp. 198-226. Song, W., Kim, E., Lee, D., Lee, M., Jeon, S-W. 2013. The sensitivity of species distribution modeling to scale differences. Ecological Modelling 248, 113-118. speciesLink. 2014. Disponible en: splink.cria.org.br [consultado en diciembre 2014]. Swets, K.A. 1988. Measuring the accuracy of diagnostic systems. Science, 240, 1285-1293. En: Araújo, M.B., Pearson, R., Thuiller, W., Erhard, M. 2005. Validation of species climate impact models under climate change. Global Change Biology 11, 1-10. Tacán, M. V. 2007. Caracterización agromorfológica e identificación de zonas potenciales de conservación y producción de guanábana (Annona muricata) y chirimoya (Annona cherimola) en fincas de agricultores y condiciones ex situ en Costa Rica. CATIE. Escuela de Posgrado. pp. 97. The New York Botanical Garden. 2014. Catalog of Vascular of Eastern Brazil. Disponible en: nybg.org [consultado en diciembre 2014]. Plant Species The Plant List. 2013. Version 1.1. Publicado en Internet: www.theplantlist.org Tropicos. 2014. Missouri Botanical Garden. Disponible en: www.tropicos.org [consultado en diciembre 2014]. Valverde, A.J. y Lobo, J.M. 2007. Threshold criteria for conversion of probability of species presence to either–or presence–absence. Acta Oecologica 31 (3), 361-369. van Zonneveld, M., Scheldeman, X., Escribano, P., Viruel, M.A., Van Damme, P., García, W., Tapia, C., Romero, J., Sigueñas, M., Hormaza, J.I. 2012. Mapping genetic diversity of cherimoya (Annona cherimola Mill.): application of spatial analysis for conservation and use of plant genetic resources. PloS one 7 (1). Vargas-Amelin, E. & Pindado, P. 2014. The challenge of climate change in Spain: Water resources, agriculture and land. Journal of Hydrology 518, 243-249. 21 Anexo 1 Listado de géneros estudiados y valoración obtenida (se asigna un valor 1 a aquellos géneros presentes en los distintos criterios) (en negrita, géneros seleccionados). Género Familia Annona Persea Pouteria Psidium Spondias Juglans Anacardium Bactris Brosimum Cordia Crescentia Inga Solanum Byrsonima Carica Casimiroa Citrus Chamaedorea Chrysophyllum Dialium Genipa Hymenaea Manilkara Muntingia Passiflora Physalis Pithecellobium Senna Simarouba Theobroma Bunchosia Cassia spp Castilla Ceiba Coccoloba Cocos Crataegus Diospyros Erythrina Gliricidia Hylocereus Maclura Malpighia Annonaceae Lauraceae Sapotaceae Myrtaceae Anacardiaceae Juglandaceae Anacardiaceae Arecaceae Moraceae Boraginaceae Bignoniaceae Leguminosae Solanaceae Malpighiaceae Caricaceae Rutaceae Rutaceae Arecaceae Sapotaceae Leguminosae Rubiaceae Leguminosae Sapotaceae Muntingiaceae Passifloraceae Solanaceae Leguminosae Leguminosae Simaroubaceae Malvaceae Malpighiaceae Leguminosae Moraceae Bombaceae Polygonaceae Arecaceae Rosaceae Ebenaceae Leguminosae Leguminosae Cactaceae Moraceae Malpighiaceae GRIN1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 Zeven & De Wet, 19822 1 1 1 1 1 1 1 1 1 1 1 1 1 Cordero & WAC & Boshier, 20033 CATIE, 20144 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 22 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 CAPGERNet et al., 20085 1 1 1 1 1 1 1 Total 5 5 5 5 5 5 4 4 4 4 4 4 4 3 3 3 3 3 3 3 3 3 3 3 3 3 3 3 3 3 2 2 2 2 2 2 2 2 2 2 2 2 2 Género Familia Mangifera Minquartia Monstera Parmentiera Pereskia Prunus Roseodendron Rubus Terminalia Acnistus Alibertia Artrocarpus Bellucia Bixa Borojoa Brahea Caryocar Clidemia Conostegia Cyrtocarpa Chrysobalanus Entada Euterpe Ficus Garcinia Guazuma Jaltomata Macadamia Macleania Matisia Morus Myrciaria Nectandra Nephelium Omphalea Rollinia Sambucus Schinus Solandra Tamarindus Vasconcellea Ximenia Zanthoxylum Anacardiaceae Coulaceae Araceae Bignoniaceae Cactaceae Rosaceae Bignoniaceae Rosaceae Combretaceae Solanaceae Rubiaceae Moraceae Melastomataceae Bixaceae Rubiaceae Arecaceae Caryocaraceae Melastomataceae Melastomataceae Anacardiaceae Chrysobalanaceae Leguminosae Arecaceae Moraceae Clusiaceae Malvaceae Solanaceae Proteaceae Ericaceae Malvaceae Moraceae Myrtaceae Lauraceae Sapindaceae Euphorbiaceae Annonaceae Adoxaceae Anacardiaceae Solanaceae Leguminosae Caricaceae Ximeniaceae Rutaceae GRIN1 1 1 1 1 Zeven & De Wet, 19822 Cordero & WAC & Boshier, 20033 CATIE, 20144 1 1 1 CAPGERNet et al., 20085 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 USDA, Total 2 2 2 2 2 2 2 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 ARS, National Genetic Resources Program. 2015. Germplasm Resources Information Network (GRIN). Available in: www.arsgrin.gov 2 Zeven, A.C. & De Wet, J.M.J. 1982. Dictionary of Cultivated Plants and Their Regions of Diversity: Excluding Most Ornamentals, Forest Trees and Lower Plants. Centre for Agricultural Publishing and Documentation, Wageningen. 23 3 Cordero, J. & Boshier, D. (eds.). 2003. Árboles de Centroamérica: un manual para extensionistas. Bib. Orton IICA / CATIE. pp. 1079. 4 World Agroforesty Centre & CATIE. 2014. Central America Agroforestry Tree Functional Traits. de Recursos Fitogenéticos del Caribe (CAPGERNet), Red Norteamericana de Recursos Fitogenéticos (NORGEN), Red Andina de Recursos Fitogenéticos (REDARFIT), Sub-Programa para Recursos Fitogenéticos de PROCISUR (REGENSUR), Red Mesoamericana de Recursos Fitogenéticos (REMERFI) y Red Amazónica de Recursos Fitogenéticos (TROPIGEN). 2008. Hacia una Estrategia Hemisférica racional de Conservación de los recursos fitogenéticos para la alimentación y la agricultura en las Américas. 58 p. 5 Red Anexo 2 Priorización de especies cultivadas y Parientes de Cultivos del género Persea Género Especies cultivadas Parientes del cultivo Persea Persea americana Mill. P. americana Mill. P. schiedeana Nees P. schiedeana Nees P. donnell-smithii Mez P. liebmannii Mez P. rigens C.K.Allen P. sessilis Standl. & Steyerm. P. standleyi C.K.Allen P. vesticula Standl. & Steyerm. P. pallescens (Mez) LoreaHern * La nomenclatura de las especies seleccionadas se estandariza con The Plant List (2013). Priorización de especies cultivadas y Parientes de Cultivos del género Annona. Género Especies cultivadas Parientes silvestres del cultivo Annona Annona cherimola Mill. A. glabra L. A. amazonica R.E.Fr A. ambotay Aubl. A. cordifolia (Szyszył.) Poepp. ex Maas & Westra A. cornifolia A. St.-Hil. A. coriacea Mart. A. macroprophyllata Donn. Sm. A. montana Macfad. A. muricata L. A. purpurea Moc. & Sessé ex Dunal A. reticulata L. A. scleroderma Saff. A. squamosa L. A. A. A. A. A. A. dioica A. St.-Hil. excellens R.E.Fr. foetida Mart. hypoglauca Mart. macrocalyx R.E.Fr. montana Macfad. A. A. A. A. A. monticola Mart. nutans (R.E.Fr.) R.E.Fr. paludosa Aubl. scandens Diels ex Pilg. sericea Dunal * La nomenclatura de las especies seleccionadas se estandariza con The Plant List (2013). 24