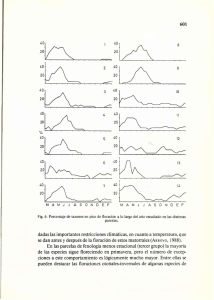
Naufraga balearica - Herbari Virtual del Mediterrani Occidental
Anuncio

MEMORIA DE INVESTIGACIÓN Naufraga balearica (Apiaceae) especie endémica y vulnerable de las Islas Baleares. Biología reproductiva y efecto de la herbivoría Por: Eva Moragues Botey Dirección: Dr. Juan Rita Larrucea Departamento de Biología. Lab. Botánica Universidad de las Islas Baleares Enero 2005 Naufraga balearica Constance et Cannon ÍNDICE pág. 1. Introducción . . . . . . . . . 4 . . . . . . . . . . . . . . . . . . . . . 6 9 11 3.1 Morfología del aparato vegetativo . 3.2 Reproducción asexual . . 3.3 Inflorescencias, Flores y Frutos . 3.4 Cariología y variabilidad genética . . . . . . . . . . . . . . . . . . . . . 12 16 16 18 . . 19 . . . . 21 22 . . 23 2. Medio Físico 2.1 Descripción del hábitat 2.2 Climatología . . 2.3 Edafología . . 3. Descripción de Naufraga balearica 4. Biología de Naufraga balearica 4.1 Reclutamiento. Estudios in situ 4.1.1 Material y métodos . . . . . 4.1.2 Resultados 4.1.2.1 Marcaje de plántulas. Éxito reproductivo 4.1.2.2 Estima del reclutamiento . . . 4.1.2.3 Supervivencia de plántulas . . . 4.1.3 Discusión y Conclusiones 4.1.3.1 Reclutamiento, longevidad y éxito reproductivo 4.2 Desarrollo del aparato vegetativo y fenología de la reproducción. Estudios ex situ 4.2.1 Material y Métodos . . . . . . . 25 4.2.2 Resultados 4.2.2.1 Desarrollo del aparato vegetativo y reproductor. Floración, fructificación y ciclo de floración a) Sistema foliar . . . . . . 27 b) Propagación vegetativa por estolones . . . 28 c) Desarrollo de las Inflorescencias . . . 32 d) Formación de las Umbelas . . . . 33 e) Fructificación . . . . . . 34 f) Ciclo de floración . . . . . . 35 4.2.3 Discusión y Conclusiones 4.2.3.1 Reproducción asexual . . . . . 37 4.2.3.2 Reproducción sexual . . . . . 39 2 Naufraga balearica Constance et Cannon 4.3 Polinización 4.3.1 Material y Métodos . . . . . . 41 4.3.2 Resultados 4.3.2.1 Comportamiento de los polizadores . . 42 4.3.2.2 Actividad circadiaria y esfuerzos de polinización de las hormigas . . . . . . 44 4.3.3 Discusión y Conclusiones . . . . . 45 5. Efecto del pisoteo de herbívoros sobre las poblaciones de Naufraga balearica 5.1 Introducción . . . . . . . . . 47 . . . . . . . . 48 48 5.2 Metodología 5.2.1 Área de estudio. Parcelas permanentes 5.2.2 Medida de la frecuencia . . 5.3 Resultados 5.3.1 Evolución poblacional de Naufraga balearica en las parcelas expuestas y No expuestas al pisoteo de herbívoros . . . . 51 5.3.2 Análisis individual de las parcelas 5.3.2.1 Parcelas No expuestas al pisoteo de herbívoros . . 52 5.3.2.2 Parcelas expuestas al pisoteo de herbívoros . . 54 5.3.3 Relación de Naufraga balearica con Erodium reichardii y Bellium bellidioides . . . . . . . . 56 5.4 Discusión 5.4.1 Análisis de las parcelas expuestas y No expuestas al pisoteo de herbívoros . . . . . . . . 59 5.4.2 Relaciones de Naufraga balearica con Erodium reichardii y Bellium bellidioides . . . . . . . . 60 6. Bibliografía . . . . . . . . . 62 3 Naufraga balearica Constance et Cannon 1. Introducción Naufraga balearica es una pequeña umbelífera endémica de la isla de Mallorca (Islas Baleares), considerada como un paleoendemismo1 y que forma parte de un género monoespecífico de las Islas Baleares. Fue descubierta en 1962 por el botánico belga J. Duvigneaud (Duvigneaud, 1970) y descrita posteriormente en 1967 por Constance & Cannon. Esta especie está legalmente protegida por el Convenio de Berna 19862, el Real decreto 439/903 y la Directiva Hábitat 19924 (Mus,1993). En 1997 fue catalogada por la IUCN como Especie en Peligro de Extinción, pero actualmente ha sido recalificada a la baja como especie Vulnerable (Sáez & Rosselló, 2001) debido al reciente descubrimiento de una nueva población en 1995 por G. Bibiloni y J. Soler (Bibiloni & Soler, 2002). Inicialmente se la relacionó con la subfamilia de las Hidrocotyloideae por presentar estrechas relaciones con algunos géneros sudamericanos (Constance & Cannon, 1967; Froebe, 1979; Heywood, 1985). Los caracteres típicos de las Hidrocotyloideae son: frutos con endocarpo leñoso, sin carpóforo libre, sin canales secretores (vitas) o sólo en las costillas primarias, y umbelas simples con estípulas. Algunos autores consideran que las Hidrocotyloideae están originadas a partir de las Araliaceae independientemente de las otras Apiaceae (Cronquist, A. 1981). Sin embargo, Downie et al.(2000) estudió en profundidad la filogenia de las Apiaceae a nivel molecular concluyendo que Naufraga balearica está muy relacionada con Apium graveolens; por lo tanto pertenece sin duda a la familia de las Apiaceae. En el presente trabajo, se ha llevado a cabo una revisión bibliográfica de N. balearica y se ha realizado un estudio basado en el conocimiento de la biología y morfología de la especie así como también un seguimiento detallado de las posibles variables bióticas (herbivoría y competencia entre especies) y abióticas (déficit hídrico) que inicialmente pensábamos que podían afectar negativamente a la estabilidad y desarrollo del taxon. Los principales Objetivos son: 1) Estudio del desarrollo morfológico del aparato vegetativo y de la biología reproductiva (ahondar en el conocimiento de la fenología de la floración y fructificación así como también en el éxito reproductivo de los nuevos reclutas en condiciones naturales). 2) Esclarecer si la herbivoría tiene un efecto negativo en el desarrollo y crecimiento de las poblaciones naturales de N. balearica. 1 Endemismo cuya presencia se remonta a épocas geológicas antiguas (Font Quer, 1975). Convenio relativo a la Conservación de la Vida Silvestre y del Medio Natural en Europa. En el cual se recoge una lista de las especies estrictamente protegidas, siendo la última revisión en el año 1998. 3 Real Decreto 439/90 que regula el Catálogo Nacional de Especies Amenazadas creado por la ley 4/1989 de Conservación de los Espacios Naturales y de la Flora y Fauna Silvestres. Cada Comunidad autónoma realiza sus propios catálogos. 4 La Directiva 92/43/ UE del Consejo (Directiva de Hábitats), recoge una lista de las especies vegetales de interés comunitario por las cuales los estados miembros de la UE designarán Zonas Especiales de Conservación. 4 2 Naufraga balearica Constance et Cannon 3) Conocer las relaciones interespecíficas entre N. balearica y otras dos especies vegetales de similar forma vital y que comparten hábitat y recursos naturales con la especie foco: Erodium reichardii y Bellium bellidioides. ¿Son relaciones de competencia? ¿Se desplazan unas a otras ó más bien viven independientemente las unas de las otras?. La finalidad de este estudio es tener un conocimiento amplio del funcionamiento de la especie y de los factores bióticos/abióticos que influyen en su desarrollo, en favor de la elaboración de futuros planes de conservación y protección de la especie y de su hábitat que aseguren su permanencia y un óptimo desarrollo de las poblaciones naturales del taxon. 5 Naufraga balearica Constance et Cannon 2. Medio Físico 2.1 Descripción del hábitat N. balearica se encuentran en la sierra de Tramuntana, una alineación montañosa de orientación NE-SO paralela a la línea de costa norte-occidental de la isla de Mallorca. Las dos únicas localidades donde ha sido encontrada son ses Coves Blanques en Pollensa con un área aproximada de 150 m2 (descubierta por Duvigneaud, J. en 1962 (Duvigneaud, 1970)) y Cap de Catalunya en el cabo de Formentor (descubierta por Bibiloni & Soler en 1995 (Bibiloni et al., 2002)) (Fig 1). Ambas localidades muy próximas entre ellas (11km aproximadamente) están localizadas en la cara norte de la sierra en taludes que se asientan a la sombra de acantilados escarpados y de pendientes elevadas. Son zonas húmedas donde el recalado del agua de los acantilados calcáreos y el rocío matinal parecen ser aportes hídricos necesarios para la supervivencia del taxon. Se encuentran aproximadamente a 100 y 200 m sobre el nivel del mar en zonas ventosas y afectadas por los aerosoles marinos (Foto 1). Fig.1. Localidades de N. balearica: Ses Coves Blanques en Pollensa, Cap de Catalunya y Mont Fumat en el cabo de Formentor. 6 Naufraga balearica Constance et Cannon a) b) Foto 1. Hábitat de N. balearica. a) Cap de Catalunya (Peninsula de Formentor) b) Coves Blanques (Pollensa) 7 Naufraga balearica Constance et Cannon Este paisaje cárstico y abrupto soporta un fuerte dinamismo del substrato ocasionado por los fuertes desniveles y el continuo tránsito de herbívoros. Son zonas poco accesibles para la circulación de personas, pero al mismo tiempo es un hábitat muy vulnerable a cualquier perturbación a/biótica que pueda alterar el ritmo ecológico natural de la zona. La vegetación del área está caracterizada por un matorral esclerófilo bajo y abierto, donde abunda Ampelodesmos mauritanica (Poiret) T. Durand et Schinz junto con caméfitos espinescentes ( Smilax aspera L. subsp. balearica Willk.) que recubren gran parte de la localidad de la península de Formentor, y comunidades de pequeños caméfitos y hemicriptófitos ricas en taxones endémicos como (Paeonia cambessedesii (Willk.)Willk., Cymbalaria aequitriloba (Viv.) A. Cheval., Sibthorpia africana L., Solenopsis minuta (L.) C. Presl subsp. balearica (E. Wimm.) Meikle, Erodium reichardii (Murray) DC., y Bellium bellidioides L. entre otros). En zonas más abiertas y expuestas a los aerosoles marinos y a una radiación solar mayor, hay prados de gramíneas perennates con abundancia de Dactylis glomerata. Duvigneaud (1970) incluye a N. balearica en la asociación Sibthorpio-Arenarietum balearicae de la Alianza Arenarion balearicae Bolòs & Molinier (1958)1969 (Bolòs, 1996). La población del Penyal Fumat, también localizada en el Cap de Formentor, es una población muy pequeña formada por pocos individuos (con una superficie aproximadamente 9m2) y situada a nivel del mar (Fig 1). Esta localidad no se encuentra en acantilados costeros, pero sí a la sombra de roquedos muy húmedos y protegidos de la exposición directa del sol (a 500-800 m de la línea de costa). En 1981 se descubrió una pequeña población de N. balearica en Córcega la cual se extinguió dos años después de su descubrimiento (Gamisans et al., 1996). La veracidad de esta población se puso en duda mediante análisis genéticos en el 2000 por Fridlender and Boisselier-Dubayte. La mayoría de trabajos realizados in situ con N. balearica que se presentan en esta memoria se han ejecutado en la localidad del Cap de Catalunya (excepto los censos de polinización realizados en ambas localidades) debido a que se trata de la población menos escarpada, de mejor acceso y con mayor número de individuos. La población de Ses Coves Blanques (Pollensa) actualmente se encuentra algo depauperada y alterada debido a la posible recolección de ejemplares por parte de botánicos durante los primeros años tras su descubrimiento, y por una fuerte erosión del suelo en un área de pendientes muy elevadas y abundante escorrentia superficial (Mus, 1993). 8 Naufraga balearica Constance et Cannon 2.2 Climatología El clima mediterráneo se caracteriza por un período seco estival más o menos prolongado y por la presencia de precipitaciones el resto de las estaciones, principalmente en invierno. Rodó y Comín (2001) afirman que dicha variabilidad interanual es uno de los principales factores controladores en el funcionamiento ecológico de los ecosistemas terrestres mediterráneos, en los cuáles la disponibilidad de agua se erige temporalmente como uno de sus principales requerimientos limitantes. Las dos estaciones climatológicas más cercanas a las poblaciones de N. balearica son las localizadas en la bahía de Pollensa (X=508.6 km, Y=4417.9 km, Z=2 m) y en la península de Formentor (X = 518,2 km, Y = 4423,7 km) en el extremo NO de la sierra de Tramuntana. La estación de la península de Formentor está situada en una zona muy similar al hábitat de N. balearica; está localizada a 150 m sobre el nivel del mar en zonas con aportaciones de aerosoles marinos. Esta última estación climatológica está caracterizada por una menor precipitación respecto al resto de las estaciones climatológicas de la sierra de Tramuntana (Fig.2). Precipitación media de 1999 a 2002 P(mm) 200 180 160 140 120 100 80 60 40 20 0 POLLENSA FORMENTOR E F M A M J J A S O N D Fig 2. Precipitación media durante el período 1999-2002 de las estaciones climatológicas de la Bahía de Pollensa y Cap de Formentor. Debido a que la estación climatológica de la península de Formentor solo registra datos pluviométricos, también se ha seleccionado la estación de la bahía de Pollensa para tener una estimación de la temperatura media mensual. El diagrama ombrotérmico, que más se aproxima al clima del área natural de la especie (Cap de Formentor), se ha creado a partir de los datos climatológicos de ambas estaciones; los valores de temperatura pertenecen a la estación de Pollensa y los valores de precipitación a la estación del Cap de Formentor (Fig 3.). 9 Naufraga balearica Constance et Cannon 50 100,0 40 80,0 30 60,0 20 40,0 10 20,0 0 P(mm) Formentor TºC (Pollensa) Diagrama ombrotérmico 0,0 E F M A M J J A S O N D Fig 3. Diagrama ombrotérmico de la estación de Pollensa Dicho diagrama ombrotérmico sólo representa las variaciones climáticas de los últimos cuatro años (1999-2002), con lo que las curvas de temperatura y precipitación no se ajustan a las típicas del clima mediterráneo de veranos secos con elevadas temperaturas y épocas lluviosas de temperaturas suaves durante el resto del año. Hemos ajustado el diagrama ombrotérmico a estos cuatro años de estudio para tener una aproximación más real de las variaciones climáticas anuales que intervienen en la dinámica global de la especie. A este ritmo anual se superpone un ciclo de años secos y húmedos que tienen gran trascendencia para la estabilidad de las comunidades (Ver apartado 5.3.3). En el período de estudio (1999-2002) las poblaciones de N. balearica sufrieron prolongados períodos de déficit hídrico (sequía) a lo largo de todo el primer año, desde el mes de enero hasta finales de la época estival en el mes de septiembre. Las precipitaciones fueron abundantes con la llegada del otoño durante tan solo tres meses, desde octubre a diciembre. La figura 3 refleja un fuerte aumento de las precipitaciones en el mes de noviembre consecuencia del fuerte temporal que se sufrió en las islas durante el año 2001. La precipitación mínima media es de 1,7 mm en el mes de junio y la máxima media de 92,1 mm en el mes de noviembre. Los períodos de temperaturas más extremos corresponden a los meses invernales, con una temperatura mínima media mensual de 11,1 ºC en el mes de enero, y a los meses estivales con una mínima media mensual de 25,2 ºC en agosto. No obstante, pensamos que este intervalo tan largo de sequía quedó amortiguado por el microclima particular de la zona donde vive: la exposición norte de las poblaciones y su cercanía al mar. Las poblaciones están expuestas a corrientes de aire húmedo procedente del mar quedando también protegidas de largos e intensos períodos de radiación solar gracias a la sombra y protección que facilitan los acantilados. Como consecuencia de estas peculiares características de su hábitat parece que la especie se encuentra en un ambiente de temperatura más baja y valores de humedad mayores que los que están representados en el diagrama ombrotérmico. 10 Naufraga balearica Constance et Cannon 2.3 Edafología La sierra de Tramuntana, de unos 15 Km de ancho y 89 Km de longitud, constituye una alineación montañosa, orientada de SW a NE y que posee los picos más altos de la Baleares. La cara norte es extraordinariamente abrupta, con barrancos que alcanzan 400 m sobre el nivel del mar. Al alternarse niveles de rocas duras y blandas, la erosión, extraordinariamente activa, sobre estas laderas tan abruptas, ha cortado acantilados y barrancos sobre las rocas calcáreas y dolomíticas del Jurásico, y laderas más suaves o valles en el Trias superior margoso y en el mioceno (Riba et al., 1979). Los suelos mediterráneos vienen determinados en gran parte por variables como el clima, la geología, la orografía y la vegetación, por lo que dependiendo de las características físico-químicas del suelo podremos conocer la potencialidad y la fertilidad del mismo; aspectos que son determinantes en el desarrollo de la cubierta vegetal. Se han efectuado análisis de muestras de suelo procedentes de la localidad del Cap de Catalunya (tabla 1) que revelan la existencia de suelos muy calcáreos de textura franca y con un nivel satisfactorio en materia orgánica. Respecto a la salinidad del suelo, los análisis indican la presencia de suelos sódicos (porcentajes de saturación del sodio (PSI) superiores al 15%) resultado de la continua llegada de los aerosoles marinos que disminuyen la fertilidad física del suelo (ya que las arcillas corren peligro de dispersarse). Una característica usual de los suelos desestructurados por el exceso de Na+ es su casi nula permeabilidad, resultado de la dispersión de las arcillas (Saña et al. 1995), por lo que tanto la pendiente pronunciada de la zona como la desestructuración del suelo dificultan la infiltración del agua de lluvia y favorecen la erosión por pérdida de suelo. PARÁMETROS ANALIZADOS RESULTADOS INTERPRETACIÓN Arenas (%) 48,00% Limos (%) 44,00% Arcillas (%) 8,00% Textura Franca Materia Orgánica 2,92% Satisfactoria CIC (meq/100g) 13,98 Normal Carbonatos (%) 74,77% Suelo muy calcáreo Calcaria activa (%) 4,12% Baja PH (en pasta saturada) 7,86 Básico Conductividad (dS/m, 25oC) 3,76 Suelo ligeramente salino Nitrogeno total (%) 0,17% Medio Relación N/C 9,96 Liberación de N elevada Fósforo asimilable (P) (ppm) 9,00 Correcto Potasio intercambiable (K) (ppm) 98,00 Correcto Ca intercambiable (meq/100g) 8,94 Alto Sodio intercambiable (Na) (ppm) 549,00 Muy alto % de Na intercambiable (PSI) 17,07% Suelo sódico Tabla 1. Análisis físico-químicos de muestras de suelo de la población del Cap de Catalunya. [Análisis efectuados por la Conselleria d’Agricultura i Pesca del Govern Balear (2002)] 11 Naufraga balearica Constance et Cannon 3. Descripción de Naufraga balearica 3.1 Morfología del aparato vegetativo. N. balearica es un pequeño caméfito herbáceo de carácter cespitoso, que ocasionalmente puede comportarse como hemicriptófito. Su sistema radical es axonomorfo en los individuos vigorosos y fasciculado en los ejemplares jóvenes de pequeño porte. Los individuos adultos de raíz pivotante, consiguen superar las épocas estivales de estrés hídrico gracias a las sustancias de reserva almacenadas en la raíz (Friedlender, 1999). En el medio natural se observó el rebrote de nuevas hojas en los ejemplares que las habían perdido durante los meses de sequía. Estos órganos subterráneos desempeñan una función de reserva y proporcionan a la planta capacidad de supervivencia durante las épocas desfavorables (Foto 2), en estos casos N. balearica actúa como hemicriptófito con una diapausa estival. Foto 2. Engrosamientos tuberosos (Rizoma) de N. balearica. Posee hojas caulinares pecioladas y dispuestas en roseta basal aplicadas al sustrato que caracterizan la morfología de la especie. En condiciones de cultivo, las hojas pueden alcanzar longitudes de hasta 3,5 cm. (de 2 a 6 cm. según Fridlender, 1999) y presentan polimorfismo foliar (Fig. 4), la mayoría son pinnatisectas con 3-5(7) folíolos, pero algunas de ellas pueden ser enteras (Foto 3) o bipinnatisectas. Son imparipinnadas, con folíolos de margen entero (1)2-5 x 1-3 mm, glabros, ovalado– oblongos y mucronados, con una mancha blanca en el ápice del folíolo. El pecíolo está dilatado en la base y está acompañado de un par de estípulas escariosas y mucronadas (Fig. 5). En condiciones de cultivo, el tamaño de los folíolos y la longitud de los pedicelos foliares varía con la cantidad de luz incidente. Parece que los individuos que reciben una radiación suficiente poseen un sistema foliar de menor longitud que los ejemplares que crecen con escasez de recursos lumínicos. En este caso, los folíolos son de color verde claro y de consistencia poco carnosa. En la naturaleza, por su parte, las plantas son de porte más pequeño y compacto, con el limbo de las hojas brillante y de 12 Naufraga balearica Constance et Cannon consistencia carnosa, de color más oscuro y con los nervios más marcados en el haz; aunque también se han encontrado de forma esporádica algunos individuos con el sistema foliar laxo y de consistencia poco carnosa. Según Fridlender (1999) los ejemplares de N. balearica de Córcega presentaban un porte bien diferenciado a los ejemplares de Mallorca. En su caso las plantas desarrollaban un “tronco” vertical vivaz de color pardo amarillento de 5-6 mm. de diámetro y de 4 a 6 cm. de longitud, de superficie rugosa debido a las cicatrices foliares. En consecuencia, los individuos tenían forma de cojinete erguido con pocas inflorescencias axilares. Estos tallos erectos pero casi completamente soterrados también han sido observados esporádicamente en las poblaciones mallorquinas (Foto 4). Fig. 4. Siluetas de las hojas de N. balearica. La mayoría son pinnatisectas con 3-5(7) folíolos, pero algunas de ellas pueden ser enteras (Fridlender 1999). 13 Naufraga balearica Constance et Cannon Los cotiledones son de oval - oblongos a oval – lanceolados, glabros y de color verde claro. El limbo es de 5 a 6 mm. de longitud y de 1 a 1,5 mm. de anchura soportado por un pecíolo de unos 5 mm. (Foto 5). Foto 3. Polimorfismo foliar. Foto 4. “Tronco” vertical vivaz que les confiere forma de cojinete erguido. Individuos de ses Coves Blanques (Pollensa) 14 Naufraga balearica Constance et Cannon Fig.5. N. balearica. a) Hábito, b) hoja basal, c) fascículo de hojas caulinares, d) pétalo, e) Fruto inmaduro, f) sección del fruto. Imágenes obtenidas de Constance, 1967. Foto 5. Cotiledones de N. balearica. 15 Naufraga balearica Constance et Cannon 3.2 Reproducción asexual Los estolones son el mecanismo de reproducción asexual de N. balearica. Existen dos tipos de estolones, los superficiales y los subterráneos. Ambos son delgados brotes axilares con una longitud que oscila entre un mínimo de 0,6 cm y un máximo de 3,4 cm (Media ± ES: 1.85 ± 0.08). Estos estolones desarrollan una roseta de hojas en su parte apical que puede enraizar y formar una plántula nueva que con el tiempo se independizará. El número de estolones originados por la planta varía en función del tamaño de los ejemplares: plantas jóvenes de pequeño tamaño y pocas hojas forman aproximadamente de 5 a 7 estolones superficiales dispuestos radialmente alrededor de la planta madre. Las plantas de mayor porte y elevado número de hojas, generan abundantes estolones (tanto subterráneos como superficiales) pudiendo llegar a un número superior de 30 por planta. En este caso, la disposición de los estolones no es uniforme, pudiendo ser radial, bilateral o sin disposición regular (Foto 6). Los estolones que une la planta madre con la nueva plántula acabará secándose y muriendo. No todos los estolones generados por una planta son capaces de producir nuevos individuos; solo sobrevivirán las nuevas plántulas capaces de enraizar, que con el tiempo se independizarán y podrán desarrollar a su vez nuevos individuos por reproducción vegetativa. 3.3 Inflorescencia, flores y frutos Del eje central de la roseta de hojas crecen pedúnculos florales de (0,8)1,53 ± 0,03(2,3) cm. de longitud con umbelas simples de (2)11 ± 0,18(14) flores. Las flores son hermafroditas, pentámeras (aunque se han citado flores de 6 pétalos, Fridlender, 1999), de pequeño tamaño (2 mm. de diámetro) y pétalos de color blanquecino-rosado. Se han observado frecuentes variaciones en el número de flores por umbela tanto en la naturaleza como en condiciones de cultivo. Los pétalos son enteros, ovados, planos o ligeramente cóncavos con una nerviación central bien marcada. Miden menos de 1 mm. de longitud y normalmente terminan en un mucrón que se dobla hacia el interior. Sus limbos, en células globosas epidérmicas, presentan reflejos cristalinos. El corto filamento de los estambres soporta las anteras dorsifijas de color rosa vivo que producen poco polen (de color blanco puro o amarillo pálido). La morfología del polen y el desarrollo de la flor ha sido detalladamente estudiado por Fridlendler, 1999. Sus observaciones fueron las siguientes: los granos de polen son muy pequeños (p = 11-12 µm e = 7 µm) trilobados en vista polar que presentan aperturas largas con endoaperturas ovales. Elípticos, y regulares, se presentan todos viables. La exina es de gran espesor (de 1 a 2 µm). Antes de que se marchiten los estambres, el néctar empieza a gotear del estilopodio, plano y de color verdoso. Los estilos se desarrollan tardíamente; primero están envueltos en el estilopodio, se enderezan y se alargan terminando en plena madurez en un estigma globoso translúcido. La fructificación es abundante tanto en condiciones de cultivo como en la naturaleza. Como en todas las umbelíferas, los frutos (0.7-1 mm) están formados por dos mericarpos que en la madurez se separan y caen por acción de la gravedad. La 16 Naufraga balearica Constance et Cannon ausencia de carpóforo también se considera un carácter arcaico que en su día motivó la adscripción de esta especie a las Hidrocotyloideae. Fridlender (1999) encontró que algunas plantas poseían frutos formados por 3 carpelos, este carácter no ha sido observado por nosotros. Las semillas tienen crestas poco marcadas, con los lados primarios materializados por un haz fibroso que se separara fácilmente del tegumento. En sección los mericarpos son redondeados y ligeramente comprimidos lateralmente. El tamaño y número de canales secretores varía según los frutos, en cualquier caso su presencia se considera un carácter evolucionado y que confirma la adscripción de la especie a las Apioideae. 17 Naufraga balearica Constance et Cannon Foto 6. Disposición de los estolones subterráneos y aéreos de Naufraga balearica. 3.4 Cariología y variabilidad genética (resumen de Fridlender, A. & BoisserlierDubayle, M-C, 2000) Con la recolección de individuos de N. balearica procedentes de la población de Ses Coves Blanques (Mallorca) se realizaron dos recuentos cromosómicos que dieron resultados divergentes. Sobre la meiosis, Constance et al. 1976 confirman el recuento de Marchant (n = 10) publicado por Moore (1971) y sobre la mitosis, Cardona en 1977 encontró 2n = 22 cromosomas, número confirmado con posterioridad por Verlaque (1999). En otro estudio realizado en el 2000 por Fridlender, se apuntó que el origen autóctono de la población de N. balearica descubierta en 1981 en Córcega, y extinguida dos años después, era dudoso. Comparó la diversidad genética de plantas procedentes del Jardín Botánico de Lyon, Brest y Porquerolles (ejemplares recolectados en Córcega en 1981) y plantas procedentes de las localidades naturales de Mallorca. La diversidad genética fue evaluada mediante marcadores RAPD que mostraron que las plantas procedentes de los jardines botánicos tienen un origen clonal (el 81% de los individuos mostraban una “pauta dominante”). Las cinco poblaciones Mallorquinas fueron genéticamente diferentes (individuos procedentes de fecundación cruzada). Las plantas cultivadas originarias del Jardín Botánico de Lyon, Brest y Porquerolles están genéticamente muy relacionadas con los individuos muestreados procedentes de Cala San Vicente en Mallorca. 18 Naufraga balearica Constance et Cannon 4. Biología de Naufraga balearica 4.1. Reclutamiento. Estudios in situ. 4.1.1 Material y Métodos Durante cuatro años consecutivos (1999-2002) se marcaron plántulas de N. balearica en cinco parcelas naturales de un metro cuadrado aproximadamente mediante la colocación de alfileres contiguos a cada planta. Se contabilizaron y siguieron un total de 256 plántulas marcadas en seis momentos correspondientes a 3 años diferentes. Salvo en el mes de febrero y parcialmente en el mes de octubre de 1999, cuando se marcaron inicialmente un máximo de 20 plántulas en distintas parcelas de la localidad de Cap de Catalunya, en el resto de momentos de muestreo se marcaron todas las plántulas que se encontraron dado que no se alcanzó la cifra de 20 individuos por parcela. Para diferenciar el marcaje de diversas épocas de germinación, se atribuyó un color de alfiler a cada época de germinación. Con el fin de diferenciar las plántulas nuevas de las pequeñas plántulas ya existentes o de rebrote, se seleccionaron solo aquellas plántulas que aún presentaban cotiledones (solo se marcaron aquellas plántulas de las cuales se podía hacer un seguimiento claro de las mismas). El hecho de que N. balearica sea una planta cespitosa, supuso un problema a la hora de encontrar plantas con cotiledones en áreas de alta densidad de individuos. El seguimiento de las germinaciones se realizó cada tres meses. (Foto 7) 19 Naufraga balearica Constance et Cannon Foto 7. Ejemplares N. balearica de la misma generación. Plántulas germinadas en el 2000. 20 Naufraga balearica Constance et Cannon 4.1.2 Resultados 4.1.2.1 Marcaje de plántulas. Éxito reproductivo N. balearica germina después de las primeras lluvias otoñales (en condiciones de cultivo con riegos regulares se ha constatado la germinación a partir del mes de agosto). En el medio natural se observó un segundo momento de germinación en el mes de febrero, confirmando la hipótesis de dos momentos de germinación al año planteada por Fridlender (1999). 4.1.2.2 Estima del reclutamiento. En febrero de 1999, cuando se inició el trabajo de campo, se pretendía conocer la longevidad de N. balearica y la supervivencia anual de las plántulas marcadas. Ese mes la germinación fue masiva y no hubo problemas para marcar 20 individuos en cada una de las parcelas. Sin embargo, en octubre de ese año y en los años sucesivos la germinación fue menos intensa por lo que se marcaron todas las plántulas que se encontraron en cada parcela. En consecuencia, la tabla 2 permite una estima muy aproximada de la intensidad de la germinación que se produjo en esos años. Parcela 1 Parcela 2 Parcela 3 Parcela 4 Parcela 5 20 20 20 20 20 Feb. 99 20 12 20 0 0 Oct. 99 15 1 0 0 0 Feb.00 20 6 19 5 11 Oct.00 0 0 0 0 0 Feb.01 Oct. 01 15 0 12 0 0 Feb. 02 90 39 71 25 31 TOTAL Tabla 2. Número de plántulas germinadas y marcadas en cinco parcelas de localidad del Cap de Catalunya (Formentor). Total 100 52 16 61 0 27 256 1 m2 de la En esta tabla se pueden observar diferencias notables entre parcelas y épocas de germinación. Así, por ejemplo, en octubre de 1999 en las parcelas 4 y 5 no se encontró ninguna plántula, mientras que en otras dos (1 y 3) se pudieron marcar las veinte plántulas previstas en el protocolo. Los valores acumulados de plántulas marcadas durante los tres años muestran diferencias entre parcelas de hasta el 70 % (90% en la parcela 1 y 25% en la parcela 4). Las diferencias entre épocas de muestreo fueron aún más notables, con un máximo de 100 plántulas marcadas en febrero de 1999 y un mínimo de 0 en febrero de 2001. Los meses de febrero de 2000 y 2001 fueron especialmente desfavorables para la germinación. En este periodo se habían acumulado más de 24 meses consecutivos con 21 Naufraga balearica Constance et Cannon índices de sequía5 positivos (precipitaciones por debajo de la media) que tuvieron su máximo en febrero de 2000 con valores próximos al 50 % de desviación sobre la media de la precipitación acumulada a lo largo de 12 meses. En febrero de 2002, cuando los valores de precipitación se habían normalizado, se volvieron a encontrar nuevas germinaciones, pero con valores muy discretos: 27 en total entre las cinco parcelas. A estos valores tan bajos, en comparación con los observados al inicio del experimento, puede haber contribuido a la disminución del banco de semillas del suelo en las parcelas estudiadas, posiblemente ocasionada por dos factores que se han reforzado mutuamente: el largo periodo de sequía y la exclusión de los herbívoros en las parcelas objeto de estudio, que dio lugar a una disminución de la densidad de N. balearica (tal como se analiza en el apartado 5 de esta memoria). 4.1.2.3 Supervivencia de las plántulas Analizando la evolución de las germinaciones durante los dos primeros años, se observó una mortalidad muy importante después del primer verano de vida de las plántulas. Sin embargo esta mortalidad ha sido sustancialmente diferente entre las germinaciones marcadas en 1999 y las de 2000 (Fig.6). Las plántulas marcadas en octubre de 1999 y febrero de 2000 (primera época de germinación) sufrieron una mortalidad del 70,37% después del primer verano, y únicamente el 14,81% sobrevivieron al segundo verano (2001). Estos datos corresponden a dos años consecutivos muy secos, con valores del índice de sequía del 48 % en agosto de 2000 y 24% en agosto de 2001, cifras que probablemente reflejen los valores máximos dentro de la horquilla de la variabilidad de la mortalidad de las plántulas. Las plántulas que germinaron en octubre de 2000 y febrero de 2001 (segunda época de germinación), sufrieron una mortalidad en el primer verano del 60,66 % (9,71 puntos inferior que las germinadas entre octubre de 1999 y febrero de 2000). El número de plántulas marcadas en la segunda época de germinación es inferior al número de plántulas marcadas en la primera época de germinación debido a la baja germinación encontrada durante ese periodo. La supervivencia después del segundo verano (2002) fue del 21,31% (6,5 puntos superior que las germinadas entre octubre de 1999 y febrero de 2000). 5 Anomalía Interanual Relativa de la Precipitación. Es la diferencia entre la precipitación de cada mes y la correspondiente precipitación anual normal, todo dividido por la precipitación normal y expresado el resultado en porcentaje. Instituto nacional de Meteorología. 22 Naufraga balearica Constance et Cannon Plántulas germinadas % 100.00 50.00 0.00 1999 Oc t.99Feb.00 100.00 2000 1999 Oct.00Feb.01 Oc t.01Feb.02 29.63 14.81 5.56 100.00 39.34 21.31 2000 2001 Oct.02 2002 Fig. 6. Porcentaje de supervivencia de plántulas respecto al inicio en dos períodos de germinación: entre octubre de 1999 y febrero de 2000 y entre octubre de 2000 y febrero de 2001. Ambos períodos de germinación sufren las sequías del primer verano con bruscos descensos en el número de individuos. 4.1 3 Conclusiones y Discusión 4.1.3.1 Reclutamiento, longevidad y éxito reproductivo Las épocas de germinación de N. balearica tanto en condiciones de cultivo como en el medio natural están fuertemente ligadas a las condiciones ambientales, principalmente a la temperatura y precipitación. En el medio natural N. balearica tiene dos períodos de germinación; uno en el mes de octubre tras las primeras lluvias de otoño y otro en el mes de febrero pasadas las temperaturas bajas del invierno y con todavía humedad en el sustrato. No obstante, al margen de esta constante en las épocas de germinación, se ha observado que, tanto ex situ como in situ, N. balearica es capaz de germinar en otras épocas del año, p.ej. el verano de 2002 fue una estación anormal con lluvias en el mes de julio y agosto que favorecieron una germinación masiva en el mes de agosto con una repercusión negativa en el reclutamiento de ese mismo otoño. En condiciones de cultivo (con riego diario) también se ha observado la germinación en épocas estivales. Existen marcadas diferencias en el número de germinaciones encontradas en el medio natural, estas diferencias son tanto espaciales (entre parcelas) como temporales (entre años diferentes). La cantidad de germinaciones varía de un año a otro, de manera que los otoños después de un verano cálido y árido se traducen en un número muy bajo de reclutas, y años más lluviosos se manifiestan con índices de germinación más abundantes. El año 2000 y 2001 tuvo índices de sequía positivos lo que perjudicó drásticamente el número de germinaciones en esos años; el 2002 fue un año muy favorable en cuanto a la precipitación y la cantidad de germinaciones aumentó considerablemente llegando incluso a germinar en los meses de verano. Estos resultados 23 Naufraga balearica Constance et Cannon sugieren que el reclutamiento de nuevas plántulas en el medio natural sufre variaciones muy notables entre diferentes ambientes y años. Y que situaciones ambientalmente adversas, como una sequía prolongada, pueden reducir drásticamente el reclutamiento de nuevos individuos lo que, a su vez, contribuye a depauperar esas poblaciones. Este ciclo, que se refuerza a si mismo, pone de manifiesto la vulnerabilidad de las poblaciones situadas en los ambientes menos adecuados. Según Grimm and Uchmański (2001) si la dinámica de población está influenciada por pequeños y continuos cambios en los niveles de los recursos naturales entonces, el tamaño de la población queda influenciado por amplios cambios que a menudo pueden llevar incluso a la extinción. Respecto a la supervivencia de los reclutas en el medio natural se observa una mortandad importante pasado el primer verano de vida de las plántulas (70,37% en el 2000 y 60,66% en el 2001). Los meses estivales son la época más crítica y de mayor vulnerabilidad en el establecimiento de las plántulas. Pasada esta época clave en la supervivencia de los reclutas, los años consecutivos presentan índices de mortalidad también altos pero menos drásticos que los ocurridos tras el primer verano de vida. Del total de las plántulas germinadas el primer año (entre los meses de octubre de 1999 y febrero de 2000) y transcurridos cuatro años, sólo ha sobrevivido un 5,5%, y un 21’3% para las germinadas el segundo año (entre los meses de octubre de 2000 y febrero de 2001) y transcurridos tres años de estudio. 24 Naufraga balearica Constance et Cannon 4.2 Desarrollo del aparato vegetativo y fenología de la reproducción. Estudios ex situ. 4.2.1 Material y Métodos Se estudió el desarrollo del aparato vegetativo y reproductor en condiciones de cultivo de 73 ejemplares, de diferentes edades, recolectados en el medio natural (Población de Cap de Catalunya. Formentor). Las plantas se sembraron en bandejas de 0,70 x 0,4 x 0,08 m. y se mantuvieron en el exterior del campo experimental de la Universidad. Durante las épocas estivales estas bandejas fueron resguardadas bajo una cubierta traslúcida y con riego diario por aspersión de aproximadamente 3 minutos/día. (Foto 8). Foto 8. Estudios ex situ. Bandejas de experimentación con N. balearica. De cada ejemplar se contabilizó, con una periodicidad mensual, el número total de hojas, estolones e inflorescencias, así como el inicio y la duración de los períodos de formación de cada etapa de la floración. Para conseguir un estudio más detallado de la fenología floral se marcaron 5 individuos adultos y con una periodicidad diaria se recogieron las siguientes variables: - Número y longitud de hojas, estolones e inflorescencias. - Tiempos de inicio y crecimiento de hojas, estolones e inflorescencias. - Número de flores y frutos por umbela. - Tiempos de desarrollo de los botones florales, de las flores y su maduración. - Tiempo de maduración y caída de los frutos. Para el estudio de las variables relacionadas con la floración se han seguido 66 pedúnculos florales pertenecientes a 39 estolones diferentes. A su vez se estudiaron 124 flores de 17 umbelas para obtener las secuencias y períodos de la floración. 25 Naufraga balearica Constance et Cannon Se realizaron pruebas de viabilidad de semillas sembrando166 semillas en placas de petri, sobre gasas humedecidas y a temperatura ambiente. Para conocer cualquier tipo de dispersor biótico de las semillas en el medio natural, se colocaron 10 trampas de semillas dispuestas principalmente en las zonas de paso de las hormigas. El experimento se llevó acabo durante tres días enteros en la estación de Cap de Formentor y durante el pico de fructificación. Para conocer la biología de esta especie y su desarrollo en el medio natural, también se hicieron observaciones, mensuales durante dos años y trimestrales durante el último año, de las épocas de germinación, floración, fructificación y dispersión de los frutos. 26 Naufraga balearica Constance et Cannon 4.2.2 Resultados 4.2.2.1. Desarrollo de del aparato vegetativo y reproductor. Floración, fructificación y ciclo de floración. a) Sistema Foliar En el medio natural N. balearica crece formando un “césped” continuo de individuos de tal manera que es difícil de diferenciar unos individuos de otros. La propagación por estolones a distancias muy cercanas de la planta madre y el pequeño tamaño del taxon hace que el sistema foliar, debido a la escasez de espacio, crezca de forma reducida y compacta. Bajo condiciones de cultivo donde los individuos crecen aislados y sin estrés hídrico el sistema foliar es más laxo y tanto las hojas como los pedúnculos foliares son de mayor tamaño. (Foto nº 9). a) b) Foto 9. Peculiaridades del sistema foliar. a) en el medio natural y b) en condiciones de cultivo. 27 Naufraga balearica Constance et Cannon Durante los meses precedentes a la floración (de febrero a abril-mayo), la inversión energética de la planta está dirigida exclusivamente a la formación y crecimiento de nuevas hojas y probablemente raíces. Bajo condiciones óptimas de humedad los ejemplares alcanzan una media de 17,63 hojas/indv. pudiendo llegar a cifras de hasta 65 hojas/indiv. La longitud máxima media de la hoja en condiciones de cultivo es de 1,84 cm. (Tabla 3) N Rango Mín. Máx. Media ± ES 84 54,00 6,00 65,00 17,63 ± 0,93 Nº Hojas 96 4,50 0,5 5,00 1,84 ± 0,08 Long. Hojas (cm) 66 25,00 5,00 30,00 9,24 ± 0,74 Nº Estolones 68 3,20 0,60 3,40 1,85 ± 0,08 Long. Estol. (cm) 49 6,00 2,00 8 3,90 ± 0,11 Nº Inflor./estl. 42 4,00 1,00 5,00 3,10 ± 0,19 Nº Infloresc. centrales 95 1,50 0,80 2,30 1,53 ± 0,03 Long. Inflor. (cm) 96 12,00 2 14 11,02 ± 0,18 Nº Flor/Umb Tabla 3: Datos morfológicos de partes vegetativas de N. balearica desde el comienzo de su desarrollo hasta la floración. b) Propagación vegetativa por Estolones A mediados del mes de marzo las plantas comienzan a emitir del eje central de la roseta basal inicial6 estolones tanto superficiales como subterráneos. El número de estolones que genera un individuo posiblemente está relacionado con el vigor y tamaño de la planta más que con la edad del taxon en si; en condiciones naturales, se observó que individuos de tan solo una generación podían formar estolones y reproducirse vegetativamente, mientras que individuos de mayor edad pero de pequeño tamaño y reducido número de hojas no eran capaces de reproducirse ni sexual ni asexualmente. Solo generaron estolones los individuos que pudieron crecer formando un número suficiente de hojas (± min. 5). Se observó que los individuos de hojas enteras no divididas no forman estolones a lo largo del primer año de vida, mientras que los individuos de igual número de hojas pero pinnatisectas sí pueden hacerlo. Como se explica en el apartado 3 (Descripción de Naufraga balearica), individuos de la misma generación pueden variar mucho en tamaño (número de hojas) y en esfuerzo reproductivo (formación o no es estolones) (Foto 7). Se correlacionaron las variables “tamaño del sistema foliar (número de hojas)” y “número de estolones” para conocer si existe una conexión entre ambos caracteres morfológicos. Los resultados obtenidos demostraron que efectivamente hay una relación positiva muy significativa entre el número de hojas y el número de estolones que desarrollan los individuos (r: 0,79; P < 0,000; n = 82); de manera que cuanto mayor sea un individuo (en función del número de hojas), mayor es el número de estolones que forma y viceversa. 6 Al hablar de “roseta basal inicial” nos referimos a la roseta madre de la cual mediante reproducción asexual brotan estolones que crecen y pueden dar lugar a nuevos individuos también en forma de roseta de hojas. 28 Naufraga balearica Constance et Cannon Los estolones comienzan a formarse a finales de febrero y principios de marzo hasta su completo desarrollo en el mes de abril. Los estolones superficiales inician y finalizan su desarrollo antes que los estolones subterráneos. El número medio de estolones que forma un individuo es de 9,24 est./indv. con una longitud media de 1,85 cm/est. (Tabla 3). Cuando los primeros estolones llegan a su máximo crecimiento, comienza desde el ápice del mismo el desarrollo de una roseta de hojas que da lugar a un nuevo individuo (Foto 10). Algunas de estas nuevas rosetas de hojas tras florecer y fructificar consiguen enraizar e independizarse de la planta madre por degradación y separación del estolón que los unía a la planta madre. Finalmente hemos comprobado, mediante cultivos en placas de Petri, que únicamente la parte apical del estolón tiene capacidad meristemática de crecer y formar un nuevo individuo, incluso separado de la planta madre (Foto 11). 29 Naufraga balearica Constance et Cannon a) Primordio umbela Estolón b) Foto 10. Desarrollo de nuevas plántulas a partir de estolones que brotan del eje central de la planta madre. a) Inicio del desarrollo de la roseta de hojas. El sistema foliar se forma al mismo tiempo que la formación de inflorescencias. b) La parte central muerta corresponde a la planta madre, las rosetas de hojas que rodean el centro son el resultado de su reproducción asexual mediante estolones que dieron lugar a esos nuevos individuos. 30 Naufraga balearica Constance et Cannon Foto nº 11 Crecimiento en placas de petri de trozos de estolón. Solo la parte apical de los mismos tiene capacidad de crecer y formar nuevas hojas. Siembra y crecimiento posterior en macetas con tierra. Foto 11. Crecimiento en placas de petri de trozos de estolón. Solo la parte apical de los mismos tiene capacidad de crecer y formar nuevas hojas. No es necesaria su unión a la planta madre para desarrollar la roseta de hojas. 31 Naufraga balearica Constance et Cannon c) Desarrollo de las Inflorescencias Los pedúnculos de las inflorescencias nacen de la parte central de las rosetas de hojas y en su extremo superior soportan una umbela simple de flores diminutas y poco vistosas. Estas inflorescencias surgen tanto de la roseta principal de hojas (inflorescencias centrales) como de las pequeñas rosetas de los estolones (inflorescencias del estolón). Ambas inflorescencias son morfológicamente idénticas pero se diferencian en su tiempo de formación. El tamaño medio de ambas inflorescencias es de 1,53cm (Tabla 3). Su crecimiento y desarrollo responde a una secuencia característica: inicialmente la planta desarrolla las inflorescencias centrales (3,10/indv.) al mismo tiempo que empieza la formación y desarrollo de los estolones. Una vez que los estolones han alcanzado su tamaño máximo comienza, desde el ápice de cada uno de ellos, la formación de una hoja. Cuando ésta se ha desarrollado, se inicia el crecimiento de una segunda hoja acompañada de una inflorescencia en su axila (Foto 10). A medida que ambas (segunda hoja y primer pedúnculo floral o inflorescencia) se van formando, comienza a despuntar una tercera hoja acompañada también de una inflorescencia. Este proceso puede continuar hasta formar un máximo de ocho umbelas en un mismo estolón. Cuando los botones florales de la primera inflorescencia del estolón están maduros a punto de abrirse, las flores de las inflorescencias centrales ya han tenido tiempo de abrirse y fructificar. El tiempo de formación de los estolones es generalmente el mismo que necesita la planta madre para formar y desarrollar las umbelas centrales hasta su fructificación. El número medio de inflorescencias producidas por los estolones es de 3,90, aunque se han observado individuos que pueden desarrollar hasta ocho. El número medio de umbelas centrales es ligeramente inferior (3,10) y el número máximo nunca supera las cinco. Tras la formación del primer pedúnculo floral del estolón pasan 9,24 días para el comienzo de la formación del segundo, 23,91 días para el tercero y 34,47 días para el cuarto (Tabla 4). El período mínimo de floración de un individuo es de aproximadamente un mes sin tener en cuenta las inflorescencias centrales de la roseta madre. En el medio natural la época de floración abarca desde el mes de abril hasta el mes de agosto (Tabla 5). En condiciones de cultivo controlado los períodos de floración y fructificación pueden prolongarse hasta el mes de noviembre (Fridlender 1999). Pedúnculo Días transcurridos a partir Máximo de días Mínimo de días floral de la formación del primer transcurridos transcurridos (Inflorescencias) pedúnculo (Media ± ES) segundo 9,2 ± 3.58 14 8 tercero 23,9 ± 5.14 27 16 cuarto 34,9 ± 2.41 35 29 Tabla 4: Evolución temporal de las inflorescencias de los estolones de N. balearica. 32 Naufraga balearica Constance et Cannon No tenemos datos precisos del desarrollo de la primera inflorescencia de estolón pero nuestras observaciones nos indican que tarda en madurar un tiempo similar al de la segunda. Esta segunda inflorescencia tiene un período de formación más corto y rápido, alrededor de nueve días, que las inflorescencias tercera y cuarta que tardan 14,67 y 11,01 días respectivamente. Esta diferencia tan notable podría ser atribuida a una infestación de pulgones que tuvo lugar en esa época en las plantas cultivadas de N. balearica. Periodo de crecimiento Nº medio / Individuo (Media ± ES) Septiembre – Abril 17,6 ± 0,9 Sistema foliar Marzo - Abril 9,2 ± 0,7 Estolones Abril - Junio 3,9 ± 0,1 Inflorescencias Abril - Julio 11,0 ± 0,18 Botones florales Abril - Agosto Flores abiertas Abril - Septiembre Inicio Frutos Tabla 5: Ciclo vital de N. balearica desde el brote de las primeras hojas hasta la maduración de los frutos. d) Formación de las Umbelas Fridlender (1999) describe las inflorescencias de N. balearica como “umbelas cimosas” con una apertura floral centrífuga (del interior hacia el exterior) cuyo eje remata en una flor. Este aspecto de “cima” no lo hemos observado en nuestros estudios de fenología en condiciones cultivo. Por el contrario, hemos observado que la apertura floral es centrípeta (del exterior hacia el interior) propia de las umbelíferas. Se trata de una inflorescencia racemosa simple donde los radios de la umbela nacen del eje central. A lo largo del desarrollo de la umbela, ésta va sufriendo una serie de estadíos fenológicos hasta la maduración y escisión de los mericarpos que caen por barocoria. Estos estadíos florares son, progresivamente: - Formación y maduración de los botones florales. Apertura floral: Tiempo de maduración de los órganos sexuales y momento en que las flores están receptivas a ser fecundadas por polinizadores. Flor fecundada: Engrosamiento del ovario y caída de pétalos. Formación del fruto y maduración de los mismos. Las umbelas presentan una media de 11,02 flores (Tabla 3). El tiempo medio de formación de los botones florales desde el su inicio hasta la apertura de la primera flor es de 17,25 ± 0,29 días (Tabla 6). Los botones florales van madurando, floreciendo y fructificando progresivamente, desde la periferia hacia el interior de la umbela, a medida que se van formando. Desde la apertura de la primera flor hasta la formación (no maduración) del último fruto de la umbela transcurren 8,4 ± 0,35 días. Aproximadamente una semana es el tiempo necesario para la total apertura de todas las flores de una umbela. 33 Naufraga balearica Constance et Cannon Dado que cada flor permanece abierta alrededor de 1,56 ± 0,04 días, el tiempo que tienen los polinizadores para fecundar las flores es escaso (Ver apartado 4.3 de polinización), aspecto que posiblemente puede dificultar la fecundación y viabilidad de los frutos de N. balearica. Entre la apertura de las flores y la caída de los pétalos transcurren 1,3 ± 0,04 días hasta la formación del fruto, momento en el cual comienza la maduración de los mismos. Una flor tarda desde su apertura hasta el inicio de la formación del fruto una media de 2,6 días. Tiempos de formación n Media días ± ES Min. Max. Botones florales/Umb. 78 17,3 ± 0,29 10,00 21,00 Antesis floral 148 1,6 ± 0,04 1 2 Maduración Flor 127 1,3 ± 0,04 1 3 De Fl a Fr/ Umb. 20 8,4 ± 0,35 6 11 Tabla 6. Duración de los diferentes estadíos de la floración. Botones florales/Umb.: Tiempo que tardan todos los botones florales de un umbela en formarse; Antesis Floral: Momento de apertura del botón floral; Maduración Flor: Tiempo transcurrido desde la antesis hasta la maduración de la flor (cuando los estambres se han abierto); De Fl a Fr/ Umb.: Tiempo transcurrido desde la maduración floral hasta la formación del fruto (no maduración del mismo). Los datos presentados en este apartado son los resultados procedentes de las dos primeras inflorescencias desarrolladas por las plantas de estudio. Los resultados producto de la tercera y cuarta inflorescencia no se han utilizado debido a la invasión de pulgones que sufrieron las plantas sembradas durante la época de floración e) Fructificación Tanto en condiciones naturales como en condiciones de cultivo la fructificación es abundante. La maduración y caída de los frutos transcurre durante un mínimo de dos semanas y al igual que la apertura de las flores es secuencial (los frutos exteriores maduran antes). Cuando los frutos han madurado, éstos se van desprendiendo de la umbela por acción de la gravedad (barocoria) y caen a una distancia muy próxima a la inflorescencia. La mayoría de los mericarpos viables se separan y desprenden por separado. Las semillas, de muy pequeño tamaño, son fáciles de transportar por la escorrentia superficial del agua de lluvia y posiblemente por las hormigas (Fridlender, 1999), aunque la mirmerocoria no ha sido constatada por nosotros en las observaciones de campo. En el momento de máxima fructificación de la especie y en ambas localidades se hicieron ensayos para conocer la dispersión de los frutos. En las zonas de paso de las hormigas y durante días enteros se colocaron trampas de semillas, aunque no se observó 34 Naufraga balearica Constance et Cannon variación alguna en el número de frutos así como tampoco se observó ningún vector que pudiera ayudar en la dispersión de los mismos. Del total de frutos recolectados en condiciones de cultivo, un 62% resultó estar abortado. Mirando los mericarpos abortados bajo la lupa binocular, éstos tienen una apariencia anómala arrugada tipo “pasa” y son de menor tamaño que las semillas aparentemente viables, las cuales son de mayor tamaño, redondeadas, carenadas y más duras. Los frutos abortados no se separan en la madurez de los pedicelos, y tardan más tiempo en caer al suelo que los frutos viables. Se realizaron pruebas de viabilidad de semillas para verificar la capacidad germinativa de los frutos aparentemente viables y se observó que del total de semillas sembradas germinó un 91%; del total de semillas aparentemente inviables ninguna de ellas se llegó a desarrollar, confirmando nuestra apreciación inicial. Pese a este porcentaje tan elevado de frutos abortados en condiciones de cultivo (62%) la producción de semillas viables es globalmente elevada por la abundante floración y fructificación. Queda por comprobar si estos valores de viabilidad de los frutos se dan de igual manera en el medio natural. f) Ciclo de floración Se calculó el porcentaje de cada uno de los estadíos fenológicos (botones florales, flor abierta, flor fecundada, inicio de la formación de los frutos) para los doce días que como máximo dura la apertura y fructificación de las flores. Para los cálculos de dichos porcentajes se unificaron los tiempos de desarrollo considerando que todas las umbelas inician su desarrollo en el mismo momento. Se tomó el día 0 como el anterior a la abertura de la primera flor; de esta manera hemos podido calcular el porcentaje de cada estadío para el primer, segundo y sucesivos días a partir de dicho día 0. En un período de doce días una umbela es capaz de completar su ciclo de floración desde la apertura de los primeros botones florales hasta el inicio de la formación del último fruto (Tabla 7). Las flores permanecen abiertas 1 ó 2 días para pasar al segundo estadío de “flor fecundada”. A lo largo del ciclo de la umbela las flores se abren progresivamente de forma que normalmente sólo hay entre dos y tres flores abiertas al mismo tiempo. Por este motivo, el porcentaje de “flores abiertas” y también de “flores fecundadas” sufren muy poca variación a lo largo del período de desarrollo de la umbela. Por el contrario, los estadíos de “botones florales” y “fructificación” siguen una pauta descendente y ascendente respectivamente. 35 Naufraga balearica Constance et Cannon Días % Capullos % Flor abiertas % Flor senescentes % Flores aparentemente fecundadas 100 0 0 0 0 86,92 13,08 0 0 1 78,41 16,10 5,22 0,26 2 68,46 16,11 9,53 5,90 3 57,72 16,54 10,09 15,64 4 45,36 16,83 13,98 23,68 5 30,92 18,72 13,48 36,88 6 16,32 19,81 14,28 49,59 7 4,98 15,12 16,29 63,60 8 1,08 6,46 14,31 78,15 9 0 0,88 6,15 92,85 10 0 0,12 0,37 99,51 11 0 0 0 100 12 Tabla 7. Evolución secuencial en el ciclo de floración de N. balearica para los 12 días que como máximo dura la floración y fructificación de las flores de las umbelas. n= 124 flores pertenecientes a 17 umbelas. A lo largo del ciclo de floración de las dos primeras inflorescencias del estolón, se observa que la duración del ciclo (17-18 días) y el número de botones florales de ambas es similar; sin embargo, los ritmos de apertura floral y su comportamiento difieren en varios aspectos (Fig.7). Podemos observar como la curva de la primera inflorescencia del estolón presenta un comportamiento más o menos parabólico con un máximo de apertura floral a la mitad del período de floración; en el décimo día del ciclo floral, el desarrollo de la inflorescencia alcanza el pico o punto máximo de floración seguido de un descenso en el número de flores abiertas hasta el total cuajado de la umbela. La antesis tiene una duración de aproximadamente 3 días. Nº de flores abiertas de la muestra 120 Pedúnculo 1 Pedúnculo 2 100 80 60 40 20 0 1 3 5 7 9 11 13 15 17 19 21 23 25 27 Días de floración Fig.7: Ciclo de floración de las dos primeras inflorescencias. Cada punto corresponde al total de flores abiertas para cada día sobre un total de n =549 flores para la primera inflorescencia y n =445 para la segunda. La segunda inflorescencia del estolón tiene una duración de la antesis mayor, en este caso de 7 días. Por lo tanto el pico de la floración es más prolongado pero con un 36 Naufraga balearica Constance et Cannon menor número de flores abiertas en cada instante. Al tener ambos pedúnculos florales similar número de botones y misma duración de ciclo floral, se deduce que las flores del segundo pedúnculo están abiertas menos tiempo. Los tiempos de apertura floral son un promedio de 1,92 ± 0,13 (n = 16) días para la primera inflorescencia y de 1,57 ± 0,12 (n =16) días para la segunda. 4.2.3 Discusión y Conclusiones Con la llegada de las primeras lluvias otoñales, N. balearica inicia su crecimiento y desarrollo con el brote de nuevas hojas. Al inicio del período de floración (marzo), las rosetas de hojas dedican todo su esfuerzo energético al crecimiento de inflorescencias y estolones, disminuyendo el desarrollo de hojas nuevas hasta el año siguiente. 4.2.3.1 Reproducción asexual A mediados del mes de marzo las plantas comienzan a desarrollar dos tipos de estolones: unos aéreos y otros subterráneos o subsuperficiales. El crecimiento de los estolones puede darse desde el primer año de vida de la planta, aunque en condiciones naturales se ha observado que individuos jóvenes de pocas hojas no producen estolones hasta que no alcanzan un tamaño adecuado, normalmente en años sucesivos. El número de estolones está positivamente correlacionado con el número de hojas, de manera que cuantas más hojas tenga un individuo, mayor cantidad de estolones forma. Existen grandes diferencias en el número de hojas entre individuos de la misma generación; de ahí que el número de estolones por individuo dependa del vigor y tamaño de la planta, y no de la edad del individuo. Los estolones superficiales inician y finalizan su desarrollo antes que los estolones subterráneos, que son más tardíos que los superficiales. La formación de estolones es un proceso que dura un mes aproximadamente. Cuando los primeros estolones llegan a su máximo desarrollo, comienza la formación de una roseta de hojas en el ápice de cada estolón. La secuencia seguida por los estolones en la formación de nuevos individuos es la siguiente: 1. Llegados los estolones al punto máximo de crecimiento y desde el ápice del mismo comienza a brotar la primera hoja de la roseta. 2. Una vez formada la primera hoja, comienza el desarrollo del primer pedúnculo floral. Junto al pedúnculo floral tiene lugar y de manera simultánea el crecimiento de la segunda hoja de la roseta. Este crecimiento sincrónico de las inflorescencias junto a una hoja es una pauta común en el desarrollo de todas las inflorescencias del estolón (crecimiento simultáneo del pedúnculo floral y hoja basal) (Foto12). [En las inflorescencias centrales de la planta madre no se ha podido observar este hecho, 37 Naufraga balearica Constance et Cannon aunque lo más probable es que este crecimiento simultaneo de la inflorescencia con hoja también se de]. Foto 12. Formación de la primera inflorescencia del estolón tras la formación de la primera hoja (ya desarrollada). El pedúnculo floral crece siempre acompañado de una hoja. 3. Crecimiento secuencial de nuevas inflorescencias del estolón (siempre acompañadas con el desarrollo simultáneo de una hoja). Durante el desarrollo y floración de los botones florales de la primera inflorescencia se inicia, desde el eje central de la roseta, el crecimiento de la siguiente inflorescencia cuyas umbelas empiezan a abrirse cuando las umbelas del primer pedúnculo ya han fructificado. La floración de la segunda umbela transcurre durante la maduración de los frutos de la primera. Existe un período de solapamiento entre el final de la floración del primer pedúnculo floral y el inicio del segundo pedúnculo floral. Es un solapamiento de aproximadamente ocho días dónde solo coinciden al mismo tiempo unas pocas flores abiertas de ambos pedúnculos. Para estudios posteriores sería interesante continuar con el seguimiento constante de todas las inflorescencias del estolón y evaluar la sincronía floral, trabajo que no se ha podido afrontar por la invasión de pulgones. 4. Desde el mes de abril hasta el mes de julio tiene lugar la floración y fructificación; esta especie dedica cuatro meses al aumento del pool genético y a la perpetuación de la especie. (ver apartado 4.2.3.2 Reproducción sexual) 5. Finalizada la floración y fructificación, algunas de las rosetas son capaces de enraizar e independizarse de la planta madre. Las rosetas que no consiguen fijarse al sustrato mueren, por la imposibilidad de captar nutrientes del suelo, cuando los estolones de la planta madre se secan y se desprenden. Las nuevas rosetas que han enraizado consiguen 38 Naufraga balearica Constance et Cannon sobrevivir y crecer para en años consecutivos reproducirse y dar lugar a nuevos individuos. Se ha observado que algunas plantas madre al finalizar el ciclo reproductivo mueren dejando su descendencia clonal alrededor de la misma. (Foto 10). Una peculiaridad en el comportamiento de N. balearica, observada tanto en cultivo como en su medio natural, es la muerte súbita e inexplicable de plantas. En condiciones de cultivo, bandejas llenas de individuos quedaron vacías tras la muerte repentina de la mayoría de individuos después de su floración; durante dos años consecutivos a este suceso, tuvo lugar una germinación masiva y el crecimiento de nuevas plantas procedentes de semillas de la primera generación que florecieron y se reprodujeron cubriendo nuevamente las bandejas del cultivo. 4.2.3.2 Reproducción sexual Las plantas de pequeño tamaño (normalmente de primera generación) no producen estolones y tampoco florecen. Las plantas que forman estolones, producen también del eje central de la planta madre de 2 a 4 pedúnculos florales que crecen progresivamente al mismo tiempo en que se van desarrollando los estolones. Formados los estolones, éstos también formarán pedúnculos florales, de manera que un individuo forma dos tipos de inflorescencias: las centrales (que salen de la planta madre) y las del estolón (que brotan de los estolones formados por la planta madre). Ambas son morfológicamente iguales (longitud y número de flores) y presentan el mismo comportamiento floral (secuencia en la apertura de las flores y formación de frutos), salvo que la floración de las inflorescencias del estolón no se inicia hasta que las centrales están en los últimos estadíos de la fructificación. La floración comienza a principios de abril y se extiende hasta julio. Las umbelas tardan un máximo de 8-9 días en completar su ciclo de floración (hasta la formación de los frutos verdes). Las flores permanecen receptivas durante 1-2 días hasta la fecundación de las mismas. En condiciones de cultivo se han observado diferencias en el tiempo de apertura entre las flores de la primera inflorescencia y la segunda. Esta diferencia de tiempos puede ser atribuida al número de polinizadores presentes en las bandejas de cultivo o bien, a la aparición de pulgones entre las flores. Al comienzo de la floración el número de polinizadores es bajo; si bien, a lo largo del ciclo el número de flores de la muestra es mayor y por tanto la atracción hacia los polinizadores también aumenta. Esta conclusión es preliminar y necesitan de estudios más precisos para la total confirmación. Los resultados de fructificación en cultivo dan valores muy elevados de semillas inviables (62%). Sin embargo, la fructificación en el medio natural es muy abundante debido al elevado número de umbelas por individuo, aunque ésta es poco eficaz en la conservación, expansión y continuidad de la especie debido a los niveles tan bajos de supervivencia de los nuevos reclutas. Con un número tan elevado de flores, es de esperar que la fructificación y reclutamiento de nuevos individuos sea también elevado. Con las observaciones de campo de varios años y la reducida expansión del taxon en el medio natural nos hizo pensar en una primera hipótesis relacionada con la baja viabilidad de los frutos o bien con una elevada depredación de los mismos. Tras los datos obtenidos de viabilidad de 39 Naufraga balearica Constance et Cannon semillas y la escasa presencia de depredadores de éstas nos preguntamos: ¿Porqué siendo el número de frutos/individuo y la viabilidad de las semillas relativamente elevada, N. balearica está en peligro de extinción?. La respuesta la encontramos en los resultados obtenidos de germinación en el medio natural que revelaron una muy baja supervivencia de los nuevos reclutas; con tan solo un 5,56% de supervivencia en individuos germinados hace tres años (de 1999 a 2002) y un 21,31% de supervivencia en individuos germinados hace dos años (de 2000 a 2002). 40 Naufraga balearica Constance et Cannon 4.3 Polinización 4.3.1 Material y Métodos Se estudiaron los insectos que visitan las flores de N. balearica y que podían tener una función como vectores tanto “in situ” en el medio natural como en condiciones de cultivo. En la primavera/verano de 2001 se realizó un seguimiento sobre una muestra de 12 parcelas en la localidad del Cap de Catalunya desde las 8 hs. hasta las 16 h s., y en 8 parcelas en la localidad de Ses Coves Blanques desde las 6h s. hasta las 8 h s. Las parcelas de 16 cm2, en algún caso se amplió el área para incorporar un número suficiente de flores abiertas, abarcaban los diferentes microambientes presentes en cada población: zonas pedregosas en zonas con sombra, taludes terrosos, pastizales soleados, etc. Los censos se realizaron durante períodos de 10 minutos a las 8h, 10h 12h, 14h y 16h solares, contabilizándose el número de flores que eran visitadas por cada especie de insecto (que básicamente se trataba de diferentes especies de hormigas), su tiempo de permanencia sobre las flores y el número total de flores presentes en cada parcela. En los casos en que había varias hormigas visitando las flores en el mismo momento se midieron los tiempos de permanencia una vez finalizados los 10 minutos de censo. En total se hizo un seguimiento de 205 flores en Cap de Catalunya y de 104 en Ses Coves Blanques y se midieron 219 tiempos de permanencia de Plagiolepis pygmaea Latreille y 51 tiempos de Lasius Níger Foerster; las dos únicas especies de hormigas encontradas en ambas localidades Se calculó el número total de flores visitadas sumando las visitas realizadas en el total de parcelas; el cociente entre esta suma y el número total de flores observadas nos permitió estimar el promedio de visitas que recibe cada flor durante los diez minutos del censo. Lo mismo se hizo para los tiempos de permanencia. Estos datos se extrapolaron a un periodo de dos horas, una anterior y otra posterior del periodo de 10 minutos observado, para poder tener una estima del número de flores visitadas y los tiempos de permanencia a lo largo de un día completo. Para la franja de las 6 h solares únicamente se han tenido en cuenta los datos de Ses Coves Blanques, para las 8 h s se han considerado los datos de las dos localidades. Para el resto de horas se utilizaron únicamente los datos de Cap de Catalunya. El momento escogido para hacer estas observaciones fue desde el inicio de la antesis floral. 41 Naufraga balearica Constance et Cannon 4.3.2 Resultados N. balearica es polinizada por diferentes especies de hormigas, tanto en el medio natural como en condiciones de cultivo. Se observó la actividad sobre las flores de dos especies diferentes de hormigas en su hábitat natural (Plagiolepis mygmaea y Lasius niger). Se constató la presencia de polen de N. balearica sobre los cuerpos de ambos formícidos mientras visitaban las flores por lo que es indudable su actividad como vectores de polinización. No se observaron otros insectos con una actividad manifiesta sobre las flores que pudieran contribuir al intercambio de polen. En cultivo se produjo, durante dos años consecutivos, la presencia masiva de áfidos, sobre las inflorescencias. Es una plaga de rápida proliferación y de abundante población en una misma planta. Los áfidos succionan la savia de las hojas, causando amarillamiento y marchitez de las mismas. Cuando hay alta población de estos pulgones en la etapa de floración, no hay una adecuada polinización y las plantas muy infestadas detienen su desarrollo y disminuyen sus rendimientos. La escasa movilidad de estos animales y su efecto perturbador sobre las flores hacen muy poco probable que puedan actuar como posibles polinizadores. 4.3.2.1 Comportamiento de los polizadores. Plagiolepis pygmaea visita de forma regular las flores en busca de néctar, pasa de una flor a otra en una misma umbela y circula también de unas umbelas a otras. En ocasiones puede realizar varias vistas sucesivas a la misma flor con breves interrupciones entre ellas. El tiempo medio de permanencia sobre las flores es de (1) 7,2’’± 0,4 (35) (n= 184) en la estación de Cap de Catalunya (Formentor) y en la localidad de Ses Coves Blanques (Cala San Vicente) se anotaron valores similares de (2) 6,9’’± 0,9 (29) (n =28). Se contabilizaron algunas visitas de más de 15’’ (máximo de 35’’), en estos casos se trata de los tiempos acumulados de varias visitas sucesivas a la misma flor. A lo largo del día no se observaron diferencias notables en el tiempo de permanencia, salvo a las 12 h cuando el valor medio (2) 12,1’’± 2,0 (35) (n =21) es significativamente superior a los medidos en otros momentos del día (Tabla 8). Hora Solar n 8h 10h 12h 14h 16 16 44 21 36 35 t’’ P. pygmaea n t’’ L. niger t’’ medios totales P. pygmaea 6,93 ± 0,92* 7,14 ± 0,92** 7,44 ± 1,03 5,71 ± 0,56 15 3,07 ± 0,51 12,10 ± 1,98 2 6,50 ± 1,50 6,78 ± 0,63 10 6,70 ± 1,01 6,19 ± 0,63 24 4,17 ± 0,49 7,64 ± 0,97 5,11 ± 0,88 Tabla 8. Tiempos de permanencia de Plagiolepis pygmaea y Lasius niger sobre las flores de N. balearica. (*: Ses Coves Blanques; **: Cap de Catalunya). (Media ± ES). 42 Naufraga balearica Constance et Cannon Lasius niger tiene un comportamiento más dinámico que Plagiolepys pygmaea pero visita las flores de N. balearica de forma menos regular; pese a que manifiesta una gran actividad circulando entre las hojas de esta planta con frecuencia no llega a visitar las flores. Sobre 51 observaciones de visitas a flores se ha medido un tiempo medio de permanencia de 5,11’’, inferior al de Plagyolepis pygmaea (7,64’’) pero suficiente para cumplir con un papel como polinizador; el tiempo máximo de permanencia ha sido de 14’’a las 14h. Las 12 y 14h han sido los momento en que se han observado tiempos de permanencia más prolongados (media de 6,6’’ en comparación con una media de 3,05’’ a las 10h de la mañana). También se ha observado una cierta segregación entre los territorios de ambas especies de hormigas. Las distancias de separación entre las doce parcelas analizadas es inferior a 50 m sin embargo en solo cuatro de estas parcelas se ha observado la presencia de ambas hormigas sobre las flores en algún momento del día, pero incluso en éstas se pudo apreciar una marcada predominancia de una de las dos especies (Tabla 9). Esta separación puede deberse a una segregación espacial o ecológica de los territorios de ambas especies o a una separación etológica entre ellas. Parcela Nº de Flores Flores visitadas por Flores visitadas por Total Flores Plagiolepis pygmaea Lasius niger visitadas 1 23 40 0 40 2 17 43 0 43 3 22 4 89 7 4 13 0 58 58 5 10 35 1 36 6 21 41 0 41 7 11 0 0 0 8 8 31 19 50 9 22 52 3 55 10 19 159 0 120 11 26 185 0 185 12 13 62 0 62 Tabla 9. Total de flores visitadas durante 10h (de las 6hs. hasta 16hs.) por ambas hormigas y para cada parcela que contenía un número determinado de flores. En la localidad de Ses Coves Blanques, L. niger visita regularmente las flores de N. balearica; durante dos horas de observación esta hormiga visitó un mayor número de veces las flores de N. balearica que P. pygmaea, que aparentemente es menos abundante en esta localidad. En condiciones de cultivo se ha observado que las hormigas también visitan las flores de N. balearica en busca de néctar, actuando con toda probabilidad como vector de polinización. No se ha observado que otras especies de hormigas actúen también como polinizadores. 43 Naufraga balearica Constance et Cannon 4.3.2.2 La actividad circadiaria y esfuerzos de polinización de las hormigas Plagiolepis pygmaea presenta un ritmo circadiario en su actividad de visitas en las flores. En las primeras horas del día (6 hora s.), presenta una actividad casi nula que se incrementa notablemente sobre las 8 hora s. de la mañana y presenta un máximo de actividad a las 10 hora s. En las horas centrales del día disminuye su actividad sobre las flores, manteniéndose de esta manera hasta el comienzo de la tarde (14 h s.) para disminuir claramente durante la tarde (16 hs) (Fig. 8). Lasius niger presenta un comportamiento más uniforme a lo largo del día. Los valores a primera hora de la mañana (8h s.) son claramente similares a los que manifiesta a lo largo del resto del día, sin embargo estos datos no son concluyentes porque en esta franja horaria únicamente se dispone de los datos de un grupo de parcelas que se caracterizó por la baja presencia de esta especie a lo largo de todo el día. Durante el resto del día la actividad de esta hormiga se mantuvo más o menos constante. Nº de visitas diarias de ambas especies 250 200 Visitas P. Pygmaea Visistas L. niger 150 100 50 0 6 h* 8h 10 h 12 h 14 h 16 h Fig. 8. Actividad circadiaria de P. pygmaea y L. niger. (* Ses Coves Blanques) Extrapolando las visitas recibidas durante diez minutos a una horquilla de tiempo que abarca una hora antes y una hora después del momento de medida se pudo tener una idea aproximada del número de visitas que puede recibir cada flor durante dos horas y sumando los valores del total del periodo de observación se puede tener una estima del número probable de visitas que recibe una flor a lo largo el día (Tabla 10). El valor mínimo observado es de 1 visitas en el periodo entre las 5 y las 7 h.s. El valor máximo es de 15 visitas a las 10 h.s. En total se puede estimar que a lo largo del día una flor de N. balearica de la localidad de Cap de Catalunya (Formentor) puede recibir alrededor de 44,5 visitas de una hormiga independientemente de qué especie se trate. Sin embargo la dispersión de los datos es bastante importante en el conjunto de las parcelas estudiadas. Así se ha registrado desde un mínimo de 0 visitas a lo largo de todo el día en una parcela con 11 flores situada sobre un pequeño talud vertical con sombra durante todo el día, y donde las hormigas circulaban por debajo de las umbelas (situadas sobre largos pedúnculos); hasta un máximo de 85 visitas diarias en una parcela con 26 flores situada en una zona iluminada y relativamente horizontal con las umbelas próximas al nivel del suelo. 44 Naufraga balearica Constance et Cannon Hora solar Número total de visitas de cada flor durante 10 minutos para cada hora 0,09 0,31 1,26 0,72 0,82 0,50 Número total de visitas estimado durante dos horas 1,04 3,74 15,16 8,66 9,89 5,97 44,47 6h 8h 10h 12h 14h 16h Total visitas diarias Tabla 10. Número de visitas que recibe cada flor durante 10’/h o bien cada dos horas por cualquiera de las dos hormigas polinizadoras. En todos los casos el número de visitas total ha de ser necesariamente superior pues hay que añadir la actividad producida con posterioridad a las 16 hs. 4.3.3 Discusión y Conclusiones N. balearica es polinizada en el medio natural y en condiciones de cultivo por diferentes tipos de hormigas que visitan las flores en busca de néctar. En ambas poblaciones únicamente se ha observado la actividad polinizadora de dos hormigas: Plagiolepis pygmaea y Lasius niger. Esta función de transferencia de polen por parte de las hormigas se ha constatado por la presencia de polen en la parte frontal de la cabeza de las mismas y por su continua actividad y circulación entre y sobre las flores. En condiciones de cultivo, también se ha observado la presencia de hormigas sobre las flores y además la aparición de áfidos (pulgones) que tuvieron un efecto perturbador en el correcto desarrollo de la planta. Se les descarta la posibilidad de tener una cierta actividad polinizadora ya que causaron el amarillamiento y marchitez de las hojas e inflorescencias de la especie. Una flor de N. balearica tiene un valor medio diario de 44,47 visitas, independientemente del tipo de hormiga que actúe como vector. Este número de visitas diarias por flor presenta una dispersión espacial bastante marcada; en algunas parcelas situadas en zonas de sombra, de elevada pendiente y con umbelas de largos pedúnculos florales el número de visitas es cero (en parcela de 11flores), y en las parcelas soleadas de zonas más horizontales se ha contabilizado un máximo de 85 visitas en una parcela con 26 flores donde las umbelas están a la altura del sistema foliar por donde mayormente circulan las hormigas. Analizando el comportamiento de cada especie, P. pygmaea presenta un ritmo circadiario con un máximo de actividad a las 10h.s. Desde las 6h.s ya se observa la actividad de esta hormiga con el mayor tiempo de permanencia a las 14h.s. (7,2’’ y 6,9’’). Es más abundante en la localidad del Cap de Catalunya. L. niger tiene una actividad más dinámica pero menos regular que P. pygmaea. Se mueve rápidamente entre las hojas de N. balearica pero con menos frecuencia visita las flores. Su tiempo medio de permanencia es de 5,11’’ y tiene un máximo entre las 12 y las 14h.s. Su comportamiento es más uniforme y su ritmo de actividad es muy similar a lo largo de todo el día sin un máximo bien definido. En la localidad de Cap de Catalunya la 45 Naufraga balearica Constance et Cannon encontramos en menos parcelas que a P.pygmaea que se encuentra mayormente en todas ellas; en ses Coves Blanques parece ser más abundante. Entre ambas hormigas se ha observado una cierta segregación de territorios, en parcelas donde abunda una de las dos especies, normalmente la otra hormiga es muy poco abundante. 46 Naufraga balearica Constance et Cannon 5. Efecto del pisoteo de herbívoros sobre las poblaciones de Naufraga balearica. 5.1 Introducción Nos planteamos la hipótesis de la herbivoría como una de las posibles causas en la alteración y degradación de las poblaciones de N. balearica. La cabra silvestre (Capra hircus L.) es un mamífero introducido muy abundante y cuyas poblaciones han experimentado un aumento en los últimos años en la sierra de Tramuntana. El tránsito y pastoreo de este mamífero de grandes dimensiones podría provocar un impacto negativo en el desarrollo, establecimiento y estabilidad de las poblaciones de N. balearica. Debido al carácter cespitoso y grácil de la planta, hace que ésta sea muy vulnerable a las modificaciones y alteraciones del medio natural como consecuencia de procesos bióticos (pisoteo herbívoros, competencia entre especies...) y también abióticos (déficit hídrico, erosión del suelo). Dado que los procesos de sucesión van ligados a una perturbación del medio, la presencia de herbívoros en el hábitat de N. balearica nos hizo pensar que la alteración del medio podría ser un problema para la supervivencia de la especie. Estudios relacionados con la herbivoría (pastoreo intensivo) establecen que ésta es la principal causa en el aumento de la heterogeneidad del medio, la degradación de los hábitats y el dominio de las especies dominantes (Ellenberg, 1996; Wang et al., 2002; Blomqvist et al., 2003), con consecuencias perjudiciales sobre la biodiversidad del lugar. Por otro lado, también se ha demostrado que un pastoreo moderado, no intensivo, está vinculado con el aumento de la diversidad de especies (Grime, 1979; McIntyre & Lavorel, 1994; Wang et al., 2002) e incluso con la conservación de especies protegidas (Miller et al., 1999). Según Markus & Sonja (2002) las especies de montaña poco abundantes sobreviven con dificultad bajo condiciones de pastoreo, mientras que las especies más generalistas y dominantes responden con un mayor número de individuos, mediada posiblemente por la reproducción asexual. Debido a la reducida área de dispersión de la especie y a su calificación como especie vulnerable por la IUCN (Sáez & Rosselló, 2001), consideramos que es imprescindible el estudio de la posible influencia de los herbívoros sobre las poblaciones de N. balearica. El riesgo de extinción de la especie es elevado debido a la existencia de tan solo dos poblaciones naturales y a la aparente recesión observada desde el descubrimiento de la misma. Esta baja expectativa de vida, está soportada en la literatura por la idea fundamental de “minimum viable populations concept” (Shaffer, 1981; Soulé, 1987) que indica que las poblaciones de pequeño tamaño tienen mayor probabilidad de extinguirse que las poblaciones de mayor tamaño (Endels et al., 2002). Otro aspecto importante en la estabilidad de los ecosistemas, son las perturbaciones ambientales provocadas por los episodios periódicos de sequía; que pueden eliminar de la comunidad a aquellas especies incapaces de resistir un determinado umbral de escasez de agua, favoreciendo por tanto a las especies más resistentes que se mantienen y expanden colonizando nuevos espacios (Pugnaire et al., 2001). Al cerrar las parcelas de N. balearica al paso de herbívoros, se observó que la presencia de dos taxones que conviven con ella (Bellium bellidioides L. y Erodium 47 Naufraga balearica Constance et Cannon reichardii (Murria) DC.) también podría determinar el crecimiento y desarrollo de la especie. Estas tres especies herbáceas tienen una gran demanda de agua y un sistema radical muy superficial que favorece la competencia hídrica entre ellas. Las relaciones de competencia (espacio, nutrientes y disponibilidades de agua) entre estos tres taxones de similar forma vital y los períodos de estrés hídrico podrían determinar la supervivencia de N. balearica. La competencia tiene efectos significativos en los patrones de distribución de las plantas, en su abundancia relativa y en la diversidad, con importantes efectos sobre la estructura de la comunidad (Goldberg & Barton 1992). En esta memoria se evalúa el papel del pisoteo de los herbívoros sobre la evolución y dinámica de crecimiento de N. balearica así como también la relación existente entre sus dos especies competidoras más cercanas (B. bellidioides y E. reichardii). Las preguntas planteadas son: (1) ¿la actividad de los herbívoros altera el medio natural de N. balearica provocando algún cambio en el crecimiento, regresión o estacionalidad de las poblaciones?, (2) ¿tienen alguna repercusión los períodos prolongados de sequía sobre las poblaciones de N. balearica? y (3) ¿puede ser desplazada N. balearica por las dos especies que principalmente coexisten con ella? 5.2 Metodología 5.2.1 Área de estudio. Parcelas permanentes En la localidad del Cap de Catalunya (Formentor) se seleccionaron quince parcelas permanentes de 25 cm2, unas sin ningún tipo de protección frente a la actividad de los vertebrados herbívoros (n =7) y otras protegidas con una red metálica (luz de 0,5 cm) que las protegía de la perturbación de estos animales (n =8). De cada una de las parcelas se midió la cobertura vegetal (frecuencia) de N. balearica, Bellium bellidioides y Erodium reichardii, especies que comparten el mismo hábitat y que “a priori” podrían actuar como posibles especies competidoras por tener formas vitales semejantes. Durante cuatro años (1999-2002) y con una periodicidad trimestral, se analizaron y compararon las diferencias de cobertura entre ambos tratamientos (parcelas expuestas vs. parcelas no expuestas a herbívoros) para cada una de las tres especies. Al ser las poblaciones muy pequeñas y tratarse de una especie amenazada, se actuó con gran precaución en la elección del número y tamaño de las parcelas, para alterar lo mínimo posible el desarrollo normal de la especie, el cual podía ser afectado por la colocación de las mallas permanentes y la circulación de personas. 5.2.2 Medida de la Frecuencia (= Cobertura) Para evaluar el crecimiento o retroceso temporal y espacial de las tres especies a estudiar, se realizaron reproducciones gráficas (fotografías) de cada una de las parcelas con la previa colocación de una rejilla de aluminio que dividía cada parcela en 100 subcuadrículas. Una vez en el laboratorio se les otorgó un valor cuantitativo de frecuencia o cobertura a cada una de estas subcuadrículas (software Adobe Photoshop 4.0). El criterio seguido fue el siguiente: valores de 1: Cobertura del 1 al 25%; 2: del 26 al 50%; 3: del 51 al 75%; 4: del 76 al 99%; 5: 100% y 0: 0% de cobertura. Como 48 Naufraga balearica Constance et Cannon resultado final se obtuvieron 15 matrices de datos para cada trimestre (60 matrices anuales/taxon) y durante cuatro años. Se trabajó, por lo tanto, con un total de 180 matrices de 100 valores de cobertura (Fig. 9). La suma de los valores de cobertura de las subcuadrículas de una parcela se utilizó como un valor frecuencia para cada especie y en cada cuadrícula. Además, la suma de los valores de las matrices de las quince parcelas, y para cada especie, se ha utilizado como un valor indicador de la frecuencia para el conjunto de la población de la localidad estudiada. Para comparar las parcelas sujetas a los dos tratamiento (expuestas y no expuestas a herbívoros) se ha realizado un análisis de la varianza (ANOVA) de las frecuencias anuales de N. balearica. Para analizar los cambios anuales que sufre N. balearica en las parcelas con o sin E. reichardii y B. bellidioides, se ha aplicado una prueba T para muestras relacionadas, la cual compara las medias de dos variables de un solo grupo (software: SPSS). 49 Naufraga balearica Constance et Cannon a) b) c) =a1 =a2 Fig.9. Secuenciación de la metodología utilizada en la obtención de los valores de frecuencia. a) Colocación de las mallas que evitan el paso de herbívoros y bajo las cuales se situaron las parcelas no expuestas al pastoreo. b) Colocación de una rejilla de aluminio de 25 cm2 sobre las parcelas abiertas y cerradas al pastoreo. Fotografía de las mismas. c) Valor de cobertura de las tres especies estudiadas (N. balearica, E. reichardii y B. bellidioides) para cada subcuadrículas. d) Obtención de las matrices de números para cada trimestre y durante cuatro años. d) Frecuencia o Cobertura = Σ an 50 Naufraga balearica Constance et Cannon 5.3 Resultados 5.3.1 Evolución poblacional de N. balearica en las parcelas expuestas y No expuestas a herbívoros Al contrario de lo inicialmente esperado, y durante los cuatro años de estudio, la cobertura de N. balearica sufrió una disminución importante en las parcelas aisladas al pisoteo de herbívoros. Esta bajada fue brusca el primer año (1999) y menos acentuada en los siguientes. Por el contrario, en las parcelas control (no protegidas a la herbivoría), la cobertura vegetal se mantuvo constante o únicamente sufrió pequeñas variaciones. En las parcelas cerradas a herbívoros, la cobertura de N. balearica descendió un 52,59% el primer año de estudio, más de la mitad del valor inicial de 1999. Esta bajada de cobertura fue muy brusca el primer año y menos acentuada en los siguientes (4,76% en el 2000 y 6,69% en el 2001) (Tabla 11). Por el contrario, en las parcelas control, el descenso de cobertura en 1999 fue mínimo con un valor del 0,48%, en tanto que los valores del año 2000 y 2001 tuvieron un incremento de un 2,07% y un descenso de un 21,23% respectivamente (Tabla 11). Año % Variación anual de la % Frecuencia anual Frecuencia respecto a 1999 Parcelas NO EXPUESTAS a Herbívoros 1999 100 2000 - 52,59 47,41 2001 - 4,76 42,65 2002 - 6,69 35,96 Parcelas EXPUESTAS a Herbívoros 1999 100 2000 - 0,48 99,52 2001 + 2,07 101.60 2002 - 21,23 80,37 Tabla 11. Variación de la frecuencia de N. balearica en las parcelas expuestas y no expuestas a herbívoros (datos obtenidos a partir de los valores del mes de febrero de 1999). La cobertura de N. balearica en las parcelas protegidas representaba, a final de 2002, el 35,96 % de cobertura respecto al valor inicial de 1999. En las parcelas control (no protegidas) la cobertura final fue del 80,37 %, la cual permaneció estable hasta el año anterior (2001). Por tanto, después de los cuatro años de estudio, la reducción de la cobertura resultó ser casi el doble (en un 44,41%) en las parcelas no expuestas a herbívoros que en las parcelas si expuestas. Los cambios en el tamaño de las poblaciones de N. balearica reflejan diferencias interanuales significativas entre ambos tratamientos, observándose un descenso de la frecuencia en las parcelas no expuestas a herbívoros y una oscilación más estacional en las parcelas abiertas a herbívoros (de 1999 a 2000: F = 6,48 n = 14 p < 0,026; de 2000 a 2001 F = 7,26 n = 14 p < 0,020 y de 2001 a 2002 F = 9,98 n = 14 p < 0.08) (Fig. 10). 51 Naufraga balearica Constance et Cannon Frecuencia de N. balearica en parcelas cerradas a herbívoros 1000 1999 2000 800 2001 2002 600 400 200 0 Febrero Ma yo Agosto Novi embre Frecuencia d e N . balearica en p arcelas ab iertas a herb ívo ros 1000 800 600 400 200 0 Fe brero Ma yo Agosto Novie m bre Fig. 10 Evolución mensual y anual de N. balearica en parcelas expuestas y no expuestas a la perturbación por herbívoros. Cada barra corresponde a la suma de los valores de frecuencia de todas las parcelas de cada categoría. 5.3.2 Análisis individual de las parcelas 5.3.2.1 Parcelas No expuestas a herbívoros La tendencia general de N. balearica en el conjunto de parcelas cerradas a herbívoros fue una disminución de su cobertura, como se ha explicado en el apartado anterior, sin embargo hemos observado diferentes comportamientos, en cuanto a la frecuencia, al analizar individualmente las parcelas que pueden justificar este descenso de la cobertura vegetal. Se ha observado que en tres de las parcelas cerradas a herbívoros (Fig.11, parcela 5 y 13) se produjo, durante la primera primavera, un fuerte descenso en la cobertura de N. balearica. Por el contrario, en estas parcelas E. reichardii no manifestó este comportamiento, sino que mantuvo una tendencia estacional siempre estable con ligeros incrementos de población. Este comportamiento nos hizo pensar que esta especie podía desarrollarse y crecer independiente de la presencia o no de herbívoros. A su vez, B. bellidioides incrementó considerablemente su número de individuos tras la bajada de N. balearica; en este caso la especie parece verse fuertemente favorecida por el aislamiento de los herbívoros. 52 Naufraga balearica Constance et Cannon Otro comportamiento más constante (sin cambios anuales relevantes) o bien con ligeros descensos progresivos en la cobertura de N. balearica es el observado en otras tres parcelas excluidas al pisoteo de herbívoros (Fig.11 parcela 9 y 10). N. balearica E. reichardii B. bellidioides Parcela 5(Cerrada a Herbívoros) 300 300 250 N. b alearica Parcela 10(Cerrada a Herbívoros) B. b ellidioides 450 300 400 250 350 200 300 Frecuencia 350 150 100 50 Feb.02 Nov.01 Ag.01 My.01 Feb.01 Ag.00 Nov.00 My.00 Feb.00 Oct.98 Ag.02 My.02 Feb.02 Ag.01 Nov.01 My.01 Feb.01 Ag.00 Nov.00 My.00 Feb.00 Ag.99 Nov.99 My.99 0 Feb.99 50 0 Ag.99 100 50 Parcela 9 (Cerrada aHerbívoros) N. balearica E. reichardii 250 200 150 100 Parcela 1 (Cerrada a Herbívoros) 50 My. 02 feb-02 Nov.01 Ag.01 My.01 Feb.01 Nov.00 Ag.00 My.00 Feb.00 Nov.99 Ag.99 My.99 Feb.99 Nov.98 0 0 Oct.98 250 My.99 Nov.99 My.00 Nov.00 My.01 Nov.01 Parcela 2 (Cerrada a herbívoros) N. balearica E. reichardii My.02 N. balearica 250 200 Frecuencia 200 150 100 150 100 50 50 Fig. 11. Análisis individual de diferentes parcelas de N. balearica No expuestas al pisoteo de herbívoros. En estos casos, las especies acompañantes se comportaron de igual manera que en el caso anterior (B. bellidioides sufrió subidas importantes en la frecuencia de individuos y E. reichardii respondió con una evolución más estable pero progresivamente ascendente), sin embargo, N. balearica no sufrió un descenso brusco inicial en el número de individuos sino que se produjo una bajada de densidad de manera más suave y paulatina a lo largo de los cuatro años de estudio. Estas diferencias en el comportamiento de N. balearica pueden atribuirse a que en estas parcelas su 53 Nov.02 My.02 Nov.01 My.01 Nov.00 My.00 Nov.99 Nov.02 My.02 Nov.01 My.01 Nov.00 My.00 Nov.99 My.99 Nov.98 My.99 0 0 Nov.98 Frecuencia 150 Nov.99 100 200 My.99 150 Feb.99 Frecuencia 200 Nov.98 Frecuencia 250 Frecuencia N. balearica E. reichardii B. bellidioides Parcela 13(Cerrada a Herbívoros) Naufraga balearica Constance et Cannon densidad inicial era menor que en el caso anterior, con lo que la disminución de cobertura es menos notoria. Los bajos niveles de N. balearica y las tendencias ascendentes de B. bellidioides y E. reichardii nos hizo pensar en una futura desaparición progresiva de N. balearica reemplazada por estas especies competidoras. Por último, dos de las ocho parcelas restantes colapsaron en noviembre de 2002, último trimestre del estudio. En ellas desde 1999 la cobertura de N. balearica fue descendiendo gradualmente hasta su repentina desaparición en el año 2002. Este proceso se produjo como consecuencia de un rápido y denso crecimiento de Dactylis glomerata (gramínea habitualmente pastada por las cabras), que cubrió completamente los individuos de N. balearica que aún vivían en ellas, provocando su muerte, así como también la de sus dos especies competidoras (Fig.11, parcela 1 y 2). 5.3.2.2 Parcelas expuestas a herbívoros (parcelas control) Un patrón común observado en las parcelas de N. balearica abiertas a herbívoros y que lo diferencia de las parcelas cerradas es la ausencia de colapsos y bajadas bruscas en la abundancia de la especie. Todas las parcelas o bien se mantuvieron estables durante los cuatro años de estudio o bien sufrieron suaves descensos en el número de individuos que puede explicarse por la variabilidad propia de la especie en función de las condiciones ambientales (Fig.12 parcelas 4,7,12 y 14). El comportamiento de E. reichardii fue muy similar al encontrado parcelas cerradas a herbívoros, presentó un ciclo interanual regular con oscilaciones estacionales. En las parcelas con B. bellidioides se observaron incrementos de esta especie que aparentemente no afectaron al desarrollo balearica. en las fuertes fuertes de N. La parcela 11 de N. balearica fue un caso particular de comportamiento, que rompió la tendencia general seguida por las parcelas abiertas a herbívoros, debido a un particular evento climático extremo que provocó modificaciones drásticas sobre los valores de cobertura de las especies presentes en la parcela (Fig.12 parcela 11). En esta parcela el crecimiento de N. balearica fue muy positivo a lo largo de los años de estudio hasta noviembre de 2001 cuando fue truncado por una intensa granizada que destrozó parte de la población. Este momento supuso un punto de inflexión que provocó un cambio brusco en la tendencia de ambas especies: N. balearica disminuyó drásticamente y E. reichardii aumentó hasta prácticamente cubrir el total de la parcela de estudio. Con el deshielo E. reichardii, favorecido por su capacidad de proliferación, ocupó los espacios vacíos y creció colonizando gran parte de la parcela, disminuyendo los espacios susceptibles a ser recolonizados por N. balearica. Este caso particular muestra la sensibilidad de esta especie amenazada a eventos puntuales, que pueden resultar catastróficos debido a su menor capacidad de respuesta respecto a las especies con las que compite. 54 Naufraga balearica Constance et Cannon Parcela 4 (Abierta a Herbívoros) 250 200 200 Frecuencia 100 50 100 50 N. balearica E. reichardii B. bellidioides Parcela 14 (Abierta a Herbívoros) 160 300 140 250 120 80 60 N. balearica E. reichardii 250 200 150 100 50 Nov.02 My.02 Nov.01 My.01 Nov.00 My.00 Nov.99 My.99 Nov.98 0 Fig. 12. Análisis individual de diferentes parcelas de N. balearica expuestas al pisoteo de herbívoros. Otro comportamiento interesante en la evolución de N. balearica fue el observado en las parcelas 3 y 8 situadas en el borde de un camino transitado por herbívoros. La existencia de caminos permanentes (Fig. 13, parcela 8) o la apertura de nuevos caminos (Fig. 13, parcela 3) produce un aumento del pisoteo así como una mayor erosión del suelo por la circulación del agua de lluvia que provocó la inestabilidad y pérdida del suelo. En ambas parcelas N. balearica no soportó las perturbaciones intensivas del medio lo que causó una reducción importante de su cobertura. Por el contrario, E. reichardii en la parcela 8 no se vió alterada por el hecho de encontrarse en una zona de paso; parece ser una especie mucho más resistente. B. bellidioides tuvo un comportamiento similar al de N. balearica con un descenso brusco en sus niveles de cobertura. 55 Nov.02 My.02 Nov.01 My.01 Nov.02 My.02 Nov.01 My.01 Nov.00 My.00 Nov.99 My.99 0 Nov.98 20 0 Nov.00 40 50 My.00 100 100 Nov.99 150 My.99 Frecuencia 200 Parcela 11 (Abierta a Herbívoros) Frecuencia Nov.02 My.02 Nov.01 My.01 Nov.00 My.00 Nov.99 Nov.98 N. balearica E. reichardii B. bellidioides Parcela 12 (Abierta a Herbívoros) My.99 0 Nov.02 My.02 Nov.01 My.01 Nov.00 My.00 Nov.99 My.99 Nov.98 0 Frecuencia 150 Nov.98 Frecuencia 250 150 N. balearica E. reichardii B. bellidioides Parcela 7 (Abierta a Herbívoros) N. balearica Naufraga balearica Constance et Cannon 250 150 Fig. 13. Análisis individual de diferentes parcelas de N. balearica expuestas al pisoteo de herbívoros. 5.3.3 Relación de Naufraga balearica con Erodium reichardii y Bellium bellidioides Como hemos indicado parece ser que existe una relación directa entre N. balearica y sus dos taxones competidores más cercanos, B. bellidioides y E. reichardii. En este apartado se evalúan los cambios de cobertura a lo largo del tiempo en las parcelas donde N. balearica está sola o en compañía de estas dos especies competidoras. En la Tabla 12 se pueden observar los cambios de cobertura de N. balearica sufridos en cuatro situaciones diferentes en función de las especies acompañantes. La evolución de N. balearica, en las parcelas donde no coexistía con otras especies y después de cuatro años, tuvo valores de crecimiento en su mayoría positivos (ver columna 1 de la Tabla 12). Por el contrario, cuando N. balearica coexistía con los otros dos táxones, su crecimiento respecto a los años anteriores fue mayormente negativo. N. balearica N. balearica N.balearica N. balearica + + + B. bellidioides E. reichardii B. bellidioides y E. reichardii + Feb.01 – Feb99 -∗ + My.01 – My99 -∗ -∗ + + Ag.01 - Ag.99 Nov.01 – Nov99 -∗ -∗ -∗ Feb.02 – Feb99 -∗ -∗ -(∗) -∗ ∗ ∗ My.02 – My99 + + Ag.02 - Ag.99 -∗ ∗ ∗ ∗ Nov.02 – Nov99 + + -∗ Tabla 12. Evolución de las poblaciones de N. balearica entre las fechas indicadas, en parcelas con y sin las especies competidoras. ∗ : Valores estadísticamente significativos para una prueba T de muestras relacionadas. + : Crecimiento positivo de N. balearica entre las fechas indicadas - : Crecimiento negativo de N. balearica entre las fechas indicadas. En Noviembre de 2001 un gran temporal de granizo afectó seriamente a las poblaciones de las tres especies modificando sustancialmente los resultados hasta agosto de 2002. 56 Nov. 02 My.02 My.01 Nov.00 My.00 Nov.02 My.02 Nov.01 My.01 Nov.00 My.00 0 Nov.99 0 My.99 50 Nov.99 100 50 My.99 100 200 Nov.98 Frecuencia 300 200 Nov.98 Frecuencia 250 150 N. balearica E. reichardii B. bellidioides Parcela 8 (Abierta a Herbívoros) Nov.01 N. balearica B. bellidioides Parcela 3 (Abierta a Herbívoros) Naufraga balearica Constance et Cannon Centrando nuestra atención en el crecimiento interanual de la población se observa que desde el año 1999 hasta el año 2001 o 2002 se produjo un declive de N. balearica en las parcelas donde coexistía con las otras dos especies competidoras y en las parcelas donde únicamente coexistía con E. reichardii. No está tan claro el papel que juega la presencia de B. bellidioides, aunque también se produjo un paulatino descenso en la cobertura de N . balearica. En las parcelas dónde N. balearica no coexiste con ninguna de sus especies competidoras se observan valores de crecimiento positivos. El descenso de cobertura que experimentó esta especie desde noviembre de 1999 a noviembre de 2001 se puede explicar por las particulares condiciones meteorológicas que padeció en noviembre de 2001 (fuertes granizadas), con un declive en el número de individuos; aspecto que quedó reflejado en los valores de cobertura de ciertas parcelas. A lo largo del año 2002 y gracias a las buenas condiciones meteorológicas de ese año, de elevadas precipitaciones, las poblaciones se recuperaron y se observó un aumento de la cobertura a partir de agosto de 2002. En la Fig.13 se ha representado la evolución de la cobertura (frecuencia) de las tres especies integrando los valores de las quince parcelas estudiadas, de forma que la gráfica muestra el comportamiento global de las poblaciones de las tres especies. En la misma gráfica se ha representado el valor del índice de sequía. Al incorporar la variable clima (Índice de sequía7) junto a los valores de cobertura de N. balearica, B. bellidioides y E. reichardii, hemos observado que, de las tres especies, E. reichardii fue el taxon más estable y resistente a los cambios climáticos; esta especie siguió pautas muy constantes en el tiempo con bruscos descensos de cobertura en los meses estivales y una rápida recuperación en los meses de lluvias. Bellium bellidioides también presenta un patrón de oscilación estacional. Sin embargo, los descensos de cobertura a lo largo de los meses estivales no fueron tan severos como los que experimentó E. reichardii, aunque por el contrario, su recuperación posterior siguió una pauta positiva si bien con ritmos de crecimiento mucho más lentos. La tendencia general de esta especie fue siempre positiva. 7 Anomalía (%) Interanual Relativa de Precipitación. Es la diferencia entre la precipitación de cada mes y la correspondiente precipitación anual normal, todo dividido por la precipitación normal y expresado el resultado en porcentajes. Instituto nacional de Meteorología (http://www.inm.es/cmt/palm/index.html.). 57 Naufraga balearica Constance et Cannon N. balearica E. reichardii B. bellidioides Drought Index (% x 10) 2000 1500 500 0 -500 Nov.02 Ag.02 My.02 Feb.02 Nov.01 Ag. 01 My.01 Feb.01 Nov.00 Ag.00 My.00 Feb.00 Nov.99 Ag.99 My.99 -1000 Feb.99 Frecuencia 1000 Fig. 13. Evolución temporal de la frecuencia poblacional de N. balearica, E. reichardii y B. bellidioides. Comparación con el Índice de Sequía (Anomalía Interanual Relativa de la Precipitación). Sin embargo, N. balearica, presentó un comportamiento irregular frente a los cambios climáticos. Bajo efectos climáticos extremos (granizada, sequías, etc.) las frecuencias de cobertura experimentaron descensos importantes y difíciles de recuperar; su tendencia de crecimiento a lo largo de los años de estudio fue siempre negativa. Entre el año 1999 y 2000 se padeció una sequía muy persistente que llevó a un déficit pluviométrico que registró valores históricos en las Baleares. La cobertura de N. balearica obtenida entre estos años bajó con el inicio de la sequía, hecho que nos hizo asociar la depauperación de las poblaciones a esta condición meteorológica. Para comprobar el grado de relación existente entre la variable índice de sequía y el descenso mensual de la cobertura de N. balearica se calculó el coeficiente de correlación de Pearson con sus niveles de significación. Los resultados obtenidos demostraron una correlación negativa aceptable entre el índice de sequía y la cobertura de N. balearica (r = -0,69, P = 0,05, n = 8). La misma operación se realizó para E. reichardii y B. bellidioides cuyos resultados son respectivamente: r = 0,02, P = 0,96, n = 8 y r = 0,71, P = 0,05, n = 8. Estas correlaciones apoyan lo expuesto anteriormente: el desarrollo de E. reichardii no parece estar afectado por la falta de precipitaciones, mientras que B. bellidioides aprovecha los episodios de estrés hídrico para crecer y establecerse a expensas de sus especies acompañantes. Desde finales del año 2001 y principios del 2002 N. balearica sufrió un aumento considerable de individuos en la población coincidiendo con índices de sequía negativos. Esta asociación, N. balearica - Índice de sequía, refleja una dependencia de la especie con las oscilaciones climáticas, principalmente la precipitación 58 Naufraga balearica Constance et Cannon 5.4 Discusión 5.4.1 Análisis de las parcelas expuestas y No expuestas al pisoteo de herbívoros El análisis de la cobertura de N. balearica revela tendencias diferentes según el tratamiento: parcelas cerradas y parcelas abiertas a herbívoros. En las parcelas no expuestas a herbívoros se produce siempre un descenso de N. balearica que puede incluso, en algunos casos, llegar a desaparecer. Por otro lado, la tendencia de las parcelas control, expuestas a herbívoros, responde a cambios asociados a la estacionalidad del clima y a eventos meteorológicos puntuales, por lo que sólo se producen ligeros cambios positivos y/o negativos de la frecuencia. Nuestros datos sugieren que la depauperación de las poblaciones de N. balearica, en las parcelas cerradas a herbívoros, puede estar relacionada con el crecimiento y proliferación de otras especies de similar forma vital (B. bellidioides y E. reichardii), estas especies rivalizan por los recursos hídricos y por el espacio. Hemos observado que ambas especies competidoras aumentan su recubrimiento, colonizando nuevos espacios o bien asentándose en los ya ocupados por N. balearica, superponiéndose a ésta y dominando gran parte de la superficie de las parcelas. Por otro lado, la gramínea Dactylis glomerata es otra especie competidora de mayor tamaño con un efecto también importante sobre la estabilidad de N. balearica. En las parcelas donde esta especie estaba presente, la disminución del pastoreo produjo un boom de crecimiento que provocó el desplazamiento de todas las especies vegetales presentes en la parcela e inferiores a ella en tamaño. En todas las situaciones observadas es siempre N. balearica la especie más vulnerable y más afectada cuando se elimina el pastoreo del medio natural. Con lo que la exclusión de los herbívoros provoca consecuencias negativas para la conservación de esta especie amenazada. Un patrón común observado en las parcelas de N. balearica abiertas a herbívoros, y que lo diferencia de las parcelas cerradas, es la ausencia de colapsos y bajadas bruscas en sus poblaciones. En todas las parcelas estudiadas, las poblaciones o bien se mantenían estables durante los cuatro años de estudio o bien sólo sufrían suaves descensos en el recubrimiento. Estos resultados son compatibles con la hipótesis de que la ausencia de herbívoros es el factor ambiental que diferencia ambos comportamientos. De las especies acompañantes se ha observado que E. reichardii presenta una oscilación estacional de sus poblaciones muy marcada, pero al mismo tiempo estas poblaciones son muy estables de un año a otro, con un suave crecimiento al final de los cuatro años de estudio. Por otro lado, B. bellidioides mantiene también una dirección ascendente aunque más paulatina. El aumento de cobertura de esta última especie no afecta de manera notable al desarrollo de N. balearica como lo hace E. reichardii. Sin embargo, se ha observado que la combinación de estas dos especies juntas provoca importantes descensos en la cobertura de N. balearica, causando drásticas consecuencias para la conservación y supervivencia del taxon, especialmente bajo condiciones de exclusión de herbívoros. 59 Naufraga balearica Constance et Cannon Las parcelas expuestas a herbívoros sufren ligeras modificaciones sujetas a cambios naturales tales como los rigores estacionales y las perturbaciones físicas del hábitat, mientras que en las parcelas no expuestas a herbívoros parece que son los factores bióticos la principal causa en la bajada de la frecuencia de N. balearica. Son cambios atribuidos al rápido crecimiento de las especies que conviven con N. balearica (E. reichardii, B. bellidioides y D. glomerata principalmente) que perjudican a la especie más vulnerable y de menor tamaño. Podemos afirmar pues, que la actividad moderada de herbívoros (herbivoría y/o pisoteo) favorece la heterogeneidad del medio y la apertura de nuevos espacios vacíos disponibles a ser colonizados. Este aumento del espacio como recurso es aprovechado por aquellas especies vegetales capaces de soportar una alteración moderada del medio. Por tanto, N. balearica es la especie más favorecida por la actividad moderada de los herbívoros, ya que disminuyen la proliferación y crecimiento de las especies vegetales más cercanas y competidoras (B. bellidioides y E. reichardii por efecto del pisoteo y D. glomerata por la herbivoria directa). Una eliminación total de los herbívoros provocaría un rápido crecimiento de las especies competidoras y de mayor proliferación, pudiendo provocar el ahogamiento y desaparición de la especie. Este comportamiento ha sido observado también por otros autores, por ejemplo Miller et al. (1999) estudiaron la respuesta de Gentiana nivalis frente al pisoteo de herbívoros y observaron que la presencia de ovejas ayudó en la estabilidad de sus poblaciones. Por otro lado, el pastoreo excesivo puede ocasionar una alteración intensa del medio que repercute negativamente sobre las poblaciones de N. balearica, hecho que se ha constatado por la total desaparición de la especie en zonas de paso de los herbívoros (caminos y zonas de descanso bajo los acantilados) donde prácticamente la presencia de vegetación es nula, si exceptuamos las especies nitrófilas. En consecuencia una supresión total de las cabras en el medio natural implicaría una drástica disminución de las poblaciones de N. balearica, por lo que en los futuros planes conservación se ha de contemplar la presión moderada de las cabras en la zona. 5.4.2 Relaciones de Naufraga balearica con Erodium reichardii y Bellium bellidioides La interacción de N. balearica con las especies acompañantes es una relación de competencia por el espacio y los recursos naturales (ej. agua, luz, espacio). Todas las parcelas con la presencia de las tres especies simultáneamente han manifestado una disminución de los valores iniciales de N. balearica después de los cuatro años de observación. Este comportamiento se ve exacerbado si se excluye la herbivoría, como se ha comentado anteriormente. No obstante, N. balearica tiene un comportamiento más estable cuando se encuentra aislada. Se ha observado también, que las plántulas de N. balearica que llegan a germinar en zonas colonizadas por otras especies mueren como consecuencia de la competencia por el espacio, pues son cubiertas completamente por las otras especies; el mayor número de plántulas observadas siempre ha sido en zonas desocupadas o de poca 60 Naufraga balearica Constance et Cannon cobertura vegetal (el estudio del reclutamiento ha sido abordado en la primera parte de la memória). Respuesta de las especies al déficit hídrico El desarrollo y crecimiento de N. balearica está fuertemente influenciado por los episodios de estrés hídrico. Esta especie, muy vulnerable, sufre la falta de precipitaciones con fuertes descensos en su cobertura. La importancia de las condiciones meteorológicas hace que las influencias bióticas, producto de la competencia entre especies, pasen a un segundo plano. Nuestros datos confirman la opinión de que “la interacción entre dos especies de plantas se puede considerar como algo dinámico cuyo resultado, en términos de productividad o reproducción, dependen de las condiciones abióticas” (Pugnaire et al., 2001). En los años de estudio la evolución y desarrollo de las poblaciones de E. reichardii es independiente a la falta de precipitación, ya que las alteraciones provocadas por las incidencias meteorológicas fueron fácilmente restablecidas mediante un rápido y efectivo crecimiento que le permitió recuperar con facilidad los niveles de iniciales de cobertura. B. bellidioides es más sensible que la especie anterior a la variabilidad de las condiciones meteorológicas y especialmente a los periodos de sequía. Aunque los cambios en el tamaño de sus poblaciones son graduales, cada año alcanza niveles de cobertura superiores a los anteriores. Por tanto, ambas especies presentan un marcado crecimiento poblacional a lo largo de los cuatro años de estudio con futuras tendencias de crecimiento positivas. N. balearica se ha mostrado como el taxon más vulnerable frente a las perturbaciones meteorológicas desfavorables (principalmente la sequía), las cuales provocan elevados índices de mortalidad. Su recuperación se ha mostrado difícil y lenta una vez restablecidas las condiciones favorables para su crecimiento. Todo ello hace que la evolución futura de la especie muestre una tendencia negativa con valores cada vez más bajos de cobertura. El único aspecto favorable que se ha observado en el desarrollo de N. balearica, en contra de toda expectativa esperada, es la actividad moderada de los herbívoros sobre el desarrollo de la especie. Este papel de los herbívoros en el medio natural, evita que se origine un crecimiento desorbitado de las especies competidoras, haciendo que conserve estables sus poblaciones. Salvo este punto, son las perturbaciones climáticas extremas el principal problema en el desarrollo óptimo de la especie: períodos prolongados de sequía y episodios puntuales de granizo o nieve afectan drásticamente las poblaciones de la especie. Si a ello añadimos el efecto negativo de las especies competidoras sobre el taxon, hace que la tendencia general de la especie y sus expectativas de futuro sean muy poco favorables, dirigiendo a la especie hacia una reducción importante en el número de individuos de las poblaciones, específicamente bajo un escenario de cambio climático. 61 Naufraga balearica Constance et Cannon 62 Naufraga balearica Constance et Cannon 6. Bibliografía 63 Naufraga balearica Constance et Cannon Bibiloni, G. & Soler, J. 2002. Bolletí de la Societat d’història Natural. Notes florístiques de les Illes Balears (XIV). Aportació al coneixement de la flora de Mallorca. Palma de Mallorca. Blomqvist, M.M., Vos, P., Klinkhamer, P.G.L. & Keurs, W. J. 2003. Declining plant species richness of grassland ditch banks – a problem of colonisation or extinction? 109, 391-406 Bolòs de, O. 1996. La Vegetació de les Illes Balears. Comunitats de Plantes. Institut d’estudis Catalans. Barcelona. Cardona MA, Contandriopoulos J. 1977. L’endemisme dans les flores insulaires méditerranéennes. Mediterranea 2: 49–77. Constance, L. & Cannon, J.M.F. 1967. Naufraga, a new genus of Umbelliferae from Mallorca. In V.H. Heywood (eds): Flora Europaea. Notulate systematicae ad Floram Europaeam spectantes, 6. Feddes Reppert. Spec. Nov. Regni Veg. 74, 1-4 Constance, L., T.I. Chuang, and C.R. Bell. 1976. Chromosome numbers in Umbelliferae V. Amer. J. Bot. 63: 608-625. Constance*, L. and Cannon, J.F.M. 1967. Naufraga—a New Genus of Umbelliferae from Mallorca. Heywood ed. Flora Europaea. Notulae Systematicae ad Floram Europaeam spectantes nº6. Feddes Repert. Spec. Nov. Regni. Veg. 74: 1-4. Cronquist, A. 1981. An integrated system of classification of flowering plants. Columbia University Press, New York. Downie, S.R.; Katz-Downie, D. and Spalik, K. 2000. A phylogeny os Apiaceae tribe Scandiceae : evidence from nuclear ribosomal DNA internal transcribed spacer sequences. American Journal of Botany 87(1) : 76-95. Duvigneaud, J. 1970. Ecologie de Naufraga balearica Constance & Cannon (Umbelliferae). Bull. Soc. Roy. Bot. Belgique 103, 31-38. Ellenberg, H. 1996. Vegetation Mitteleuropas mit den Alpen in oekologischer, dynamischer und historischer Sicht, 5th edn. Ulmer Stuttgart. Endels, P., Jacquemyn, H., Brys, R., Hermy, M. & De Blust, G. 2002. Temporal changes (1986-1999) in populations of primrose (Primula vulgaris Huds.) in an agricultural landscape and implications for conservation. Biological Conservation 105, 11-25. Font Quer, P. 1975. Diccionario de Botánica. Ed. Labor, S.A. Barcelona. Fridlender, A. 1999. Naufraga balearica Constance & Cannon (Apiaceae).I. Description, écologie, répartition et projets de conservations en Corse. Article à paraître au Journal de Botanique. Laboratorie de Phanérogamie, MNHN. Paris. Fridlender, A. & Boisselier-Dubayte, M.C. 2000. Comparaison de la diversité génétique (RAPD) de collections ex situ et de populations naturelles de Naufraga balearica Constance & Cannon. Comptes Rendus de l’Academie des Sciences Serie III Sciences de la Vie 323 (4), 399-406. Froebe, H.A. 1979. Die infloreszenzen der Hydrocotyloideen (Apiaceae). Trop. Subtrop. Pflanzenwelt 29 : 1-181. Gamisans, J., Moret, J., Fridlender, A. Deschâtres, R. & Dutartre, G. 1996. Le Naufraga balearica est-il en Corse? Etude du site originel, recherque de stations comparables, possibilités de réintroduction. In Jeanmonod, D. & Burdet, H.M. Notes et Contributions à la Flore de Corse, XII. Candollea 51, 552-557. Goldberg, D.E. & Barton, A.M. 1992. Patterns and consequences of interspecific competition in natural communities: a review of field experiments with plants. American Naturalist 139:771-801 Grime, J.P. 1979. Plant strategies and vegetation processes. Wiley, Chichester. 64 Naufraga balearica Constance et Cannon Grimm, V. & Uchmański, J. 2001 Individual variability and population regulation: a model of the significance of within-generation density dependence. Oecologia 131, 196-202. Heywood, V.H. 1985. Las Plantas con flores. Editorial Reverté, S.A. Barcelona. Markus, F and Sonja W. 2002. Effect of low-intensity grazing on the species-rich vegetation of traditionally mown subalpine meadows. Biololgical Conservation 104(1): 1-11. Grazing on Swiss meadows can gradually lead to the loss of species diversity. McIntyre, S. & Lavorel, S. 1994. Predicting richness of native, rare and exotic plants in response to habitat and disturbance variables across a variegated landscape. Conservation Biology 8, 521-531 Miller, G.R., Geddes, C. & Mardon, D.K. 1999. Response of the alpine gentian Gentiana nivalis L. to protection from grazing by sheep. Biological Conservation 87(3): 311-318. Moore, D.M. 1971. Chromosome studies in the Umbelliferae. The Biology and Chemistry of the Umbelliferae (ed. V.H. Heywood). Botanical Journal of the Linnean Society, 64 (Suppl. 1), 233 256. Mus, M. 1993. Plans de Conservació dels Vegetals Amenaçats de Balears. I. Mallorca. Documents tècnics de Conservació de la Consellería d’Agricultura i Pesca. Govern Balear. Pugnaire, F., Armas, C. & Tirado, R. 2001. Balance de las interacciones entre plantas en ambientes mediterráneos. Ecosistemas mediterráneos (Análisis Funcional). Simposio de la Sociedad Española de Ecología Terrestre. Editado por Zamora, R y Pugnaire, F.I Riba, O.; de Bolòs, O.; Panareda, J.M.; Nuet, J. y Gosàlbez, J. 1979. Geografia física dels països Catalans. Editora Cutres. Barcelona Rodó, X & Comín, F. 2001. Fluctuaciones del clima mediterráneo: conexiones globales y consecuencias regionales. Ecosistemas mediterráneos (Análisis Funcional). Simposio de la Sociedad Española de Ecología Terrestre. Editado por Zamora, R y Pugnaire, F.I Sáez, LL. & Rosselló, J.A. 2001. Llibre vermell de la flora vascular de les Illes Balears. Documents tècnics de conservació II època, num.9. Govern de les Illes Balears. Conselleria de Medi Ambient. Palma de Mallorca Saña, J, Moré, J.C. & Cohí, A. 1995. La gestió de la fertilidad de los suelos. Fundamentos para la interpretación de los análisis de suelo y la recomendación de abonado. Barcelona Shaffer, M.L. 1981. Minimum population sizes for species conservation. Bioscience 31, 131-134. Soulé, M.E. 1987. Viable populations for Conservation. Cambridge University Press, Cambridge. Wang, Y., Shiyomi, M., Tsuiki, M., Tsutsumi, M., Yu, X., & Yi, R., 2002. Spatial heterogeneity of vegetation under different grazing intensities in the Northwest Heilongjiang Steppe of China. Agriculture, Ecosystems and Environment 90(3): 217229. 65
![póster Api y Nau [Modo de compatibilidad]](http://s2.studylib.es/store/data/006112261_1-7d8d12080e35345733fd4cb0f38518b4-300x300.png)