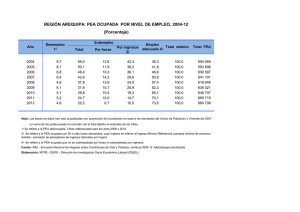
Polímeros bioestables para fabricación de implantes protésicos
Anuncio

Polímeros bioestables para fabricación de implantes protésicos: caracterización físico-química y respuesta biológica in vitro Tesis presentada por Alberto J. Campillo Fernández para obtener el título de Dr. Ing. Sci. en la Universitat Politècnica de València, octubre 2014 Directores de tesis: Dr. J.L. Gómez Ribelles Dr. J.M. Meseguer Dueñas Miembros del Tribunal: Prof. Dr. Manuel Monleón Pradas Departamento de Temodinámica Aplicada y Centro de Biomateriales e Ingeniería Tisular, Universitat Politècnica de València, España. Dr. Senentxu Lanceros-Méndez Departamento de Física. Universidade do Minho, Braga, Portugal, e INL (Iberian International Nanotechnology Laboratory), Braga, Portugal Dr. Manuel Mata Roig Departamento de Patología. Universitat de València, España La investigación descrita en esta tesis se llevó a cabo en el Centro de Biomateriales e Ingeniería Tisular de la Universitat Politècnica de València y en la Universidad Johannes Gutenberg, Mainz (Alemania). El trabajo fue financiado parcialmente por el Ministerio de Ciencia y Educación español mediante el proyecto MAT2003-5391-C03, la Generalitat Valenciana bajo la ayuda Grupos 03/018 y el programa Marie Curie del FP6 mediante el proyecto MES-CT-2004-008104 A mi familia, por su constante apoyo AGRADECIMIENTOS Al Prof. C. James Kirkpatrick, director del instituto de patología de la Universidad Johannes Gutenberg, Mainz, Alemania por acogerme en mi estancia predoctoral bajo la acción Marie Curie del FP6, Early Stage Researcher Training y a todo su equipo. Al Prof. Manuel Salmerón Sánchez, quien ha contribuido con su talento en buena parte de este trabajo. A la Dra. Marta Abad Collado, por la realización de los cultivos de células epiteliales sobre los substratos planos, realizados en Vissum, Alicante, bajo la dirección del Prof. Alió. Finalmente, quiero agradecer a mis directores José Luis Gómez Ribelles y Pepe Meseguer su ayuda y buenos consejos para que este trabajo se haya hecho, por fin, realidad. 5 INDICE Resumen ................................................................................ 8 Resum.................................................................................. 10 Abstract............................................................................... 12 Capítulo 1 . Introducción ..................................................... 14 1.1.- Ingeniería tisular ........................................................................... 15 1.1.1.- Métodos de fabricación de scaffolds ....................................... 16 1.1.2.- El método de disolución de plantilla ...................................... 18 1.1.3.- Scaffolds de acrilatos y metacrilatos ...................................... 18 1.2.- Estructura y propiedades físicas de los soportes empleados ......... 19 1.2.1.- Sobre los materiales empleados en estado seco ...................... 19 1.2.1.1.- Estructura química .................................................................... 19 1.2.1.2.- Propiedades físicas ..................................................................... 24 1.2.1.3.- Elasticidad de redes poliméricas ................................................ 31 1.2.2.- Interacción con el agua de los materiales empleados ............. 35 1.2.2.1.- Mezclas polímero-solvente ......................................................... 35 1.2.2.2.- Hinchamiento de redes poliméricas ............................................ 37 1.2.2.3.- La teoría de Flory-Huggins ........................................................ 38 1.2.2.4.- Hidrogeles .................................................................................. 39 1.2.2.5.- La transición vítrea en mezclas homogéneas .............................. 39 1.2.2.6.- La interacción hidrófoba ............................................................ 41 1.3.- Interacción material-célula ........................................................... 44 1.4.- La vascularización en los scaffolds ................................................ 45 1.5.- La epitelización en implantes protésicos de córnea ...................... 49 Hipótesis y objetivos ............................................................ 52 6 Capítulo 2 . Materiales y métodos ....................................... 53 2.1.- Materiales ..................................................................................... 54 2.1.1.- Polímeros en bloque. Sistemas P(EA-co-HEA) y P(EA-coMAAc) ................................................................................................... 54 2.1.2.- Polímeros porosos. Scaffolds de P(EA-co-HEA). ................... 55 2.2.- Métodos ........................................................................................ 58 2.2.1.- Ensayos de los materiales empleados en estado seco ............. 58 2.2.1.1.- Espectroscopía dinámico-mecánica (DMS) ................................ 58 2.2.1.2. Calorimetría diferencial de barrido (DSC) .................................. 58 2.2.1.3.- Microscopía de fuerza atómica (AFM) ...................................... 59 2.2.2.- Ensayos de los materiales empleados hidratados ................... 59 2.2.2.1.- Energía superficial (SE) ............................................................. 59 2.2.2.2.- Espectroscopía dinámico-mecánica (DMS) ................................ 59 2.2.2.3.- Calorimetría diferencial de barrido (DSC) ................................. 60 2.2.2.4.- Espectroscopía de relajación dieléctrica (DRS) .......................... 61 2.2.2.5.- Microscopía de fuerza atómica (AFM) ...................................... 61 2.2.2.6.- Contenido de agua en equilibrio (EWC) .................................... 62 2.2.3.- Adsorción de fibronectina sobre los substratos 2D ................ 62 2.2.3.1.- Preparación de polímeros en bloque para adsorción de fibronectina ................................................................................................... 62 2.2.4.- Cultivos celulares sobre los substratos 2D y 3D .................... 63 2.2.4.1.- Tipos celulares y condiciones de crecimiento ............................. 63 2.2.4.2.- Preparación de los polímeros en bloque y scaffolds para cultivo celular ........................................................................................................... 64 2.2.4.3.- Adhesión y viabilidad celular de epiteliales NHC sobre los substratos poliméricos en bloque................................................................... 65 2.2.4.4.- Tinción con hematoxilina-eosina de epiteliales NHC sobre los substratos poliméricos en bloque................................................................... 66 2.2.4.5.- Análisis de la proliferación celular de epiteliales NHC sobre los substratos en bloque ..................................................................................... 66 2.2.4.6.- Visualización y caracterización del crecimiento de las endoteliales HUVEC y los fibroblastos MRC-5 en los scaffolds........................................ 67 7 2.2.4.7.- Análisis estadístico .................................................................... 68 2.2.4.8.- Análisis molecular de la expresión génica de las endoteliales HUVEC sobre los scaffolds ........................................................................... 68 2.2.4.9.- HUVEC: análisis inmunofluorescente de Selectina E (CD62E) y PECAM-1 (CD31) ........................................................................................ 69 Capítulo 3 . Resultados y discusión ..................................... 71 3.1.- Propiedades físicas de los polímeros soporte................................. 73 3.1.1.- Caracterización de los materiales en estado seco ................... 73 3.1.2.- Propiedades de los soportes inmersos en medio acuoso. Separación de fases por interacción hidrófoba ....................................... 86 3.2.- Cultivos celulares en soportes 2D y 3D ...................................... 108 3.2.1.- Cultivo de células epiteliales conjuntivales, NHC................ 109 3.2.2.- Cultivo de fibroblastos, MRC-5 ........................................... 116 3.2.3.- Cultivo de células endoteliales, HUVEC ............................. 120 3.3.- Conformación de la fibronectina en los materiales ..................... 128 Capítulo 4 . Conclusiones ...................................................133 Glosario ..............................................................................136 Referencias..........................................................................138 8 Resumen La necesidad de polímeros bioestables para fabricación de implantes protésicos queda patente, entre otros indicadores, por la proliferación de dispositivos actualmente comercializados. La caracterización físico-química así como la respuesta biológica de un conjunto de materiales poliméricos bioestables es el objetivo último de esta tesis. En este trabajo se han sintetizado diferentes materiales poliméricos de la familia de los acrilatos y metacrilatos variando sutilmente sus características superficiales, como el grado de hidrofilia o la distribución de cargas eléctricas. El procedimiento consistió en la copolimerización via radical de acrilato de etilo, EA, acrilato de 2-hidroxietilo, HEA, y ácido metacrílico, MAAc. Se ha caracterizado los materiales en estado seco y en presencia de diferentes contenidos de agua mediante calorimetría diferencial de barrido, DSC, análisis dinámico-mecánico, DMA, microscopía de fuerza atómica, AFM, análisis dieléctrico, DRS, contenido de agua en equilibrio, EWC, y energía superficial, SE, persiguiendo el objetivo de dilucidar si el agua es capaz de inducir cambios conformacionales en las cadenas poliméricas que den lugar a una separación de fases. Sobre los materiales en forma de scaffold poroso con poros esféricos interconectados se ha cultivado fibroblastos y endoteliales. La compatibilidad de las células endoteliales se midió en términos de viabilidad celular y la adecuada diferenciación endotelial y su funcionamiento. Se han realizado cultivos de células endoteliales humanas primarias, HUVEC, y se ha determinado si su morfología y función se vio afectada por el material. Se examinó la adhesión y proliferación de las mismas, así como un marcador importante de activación endotelial, la E-selectina. Se evaluó si se mantuvieron los fenotipos endoteliales normales y sus funciones observadas 9 in vivo mediante análisis de los contactos célula-célula y la regulación de la expresión génica del marcador de activación E-selectina cuando se añadió un estímulo (LPS). Además, como posible aplicación de estos materiales en una prótesis de córnea artificial, y dado que los fibroblastos del estroma de la córnea (es decir, los queratocitos) son de relevancia en la cicatrización de la córnea se determinó cómo afectaba la hidrofilicidad del substrato a la adhesión celular de la línea de fibroblastos humanos MRC-5, como modelo celular para estudiar la disposición del citoesqueleto tras la adhesión a los diferentes soportes mediante la detección de F-actina. Asimismo, se ha sembrado células epiteliales evaluando su comportamiento/funcionamiento celular ya que uno de los requisitos esenciales para que un implante de queratoprótesis tenga éxito es que se cree y mantenga una capa de células epiteliales que impidan entrar a las bacterias al interior del ojo y permita la difusión de la capa lagrimal de manera estable en el tiempo. Así, se han analizado parámetros celulares como adhesión, proliferación y viabilidad de una línea de células epiteliales de conjuntiva humana, NHC, cultivada sobre substratos poliméricos con diferentes grados de hidrofilia y cargas eléctricas superficiales buscando qué grado de hidrofilicidad permite la epitelización del substrato y podría darle al material la flexibilidad e hidrofilicidad necesaria para un mejor contacto con los párpados y lágrima. Los resultados obtenidos se han correlacionado con la adsorción y conformación de una proteína de la matriz extracelular, la fibronectina. 10 Resum La necessitat de polímers bioestables per a fabricació d’implants protèsics queda patent, entre altres indicadors, per la proliferació de dispositius actualment comercialitzats. La caracterització fisicoquímica i la resposta biològica d’un conjunt de materials polimèrics bioestables és l’objectiu últim d’aquesta tesi. En aquest treball s’han sintetitzat diversos materials polimèrics de la família dels acrilats i metacrilats variant-ne subtilment les característiques superficials, com ara el grau d’hidrofilicitat o la distribució de càrregues elèctriques. El procediment ha consistit en la copolimerització via radical d’acrilat d’etil, EA, acrilat de 2-hidroxietil, HEA, i àcid metacrílic, MAAc. S’ha caracteritzat els materials en estat sec i en presència de diversos continguts d’aigua per mitjà de calorimetria diferencial d’escombratge, DSC, anàlisi dinamicomecànica, DMA, microscòpia de força atòmica, AFM, anàlisi dielèctrica, DRS, contingut d’aigua en equilibri, EWC, i energia superficial, SE, amb l’objectiu de dilucidar si l’aigua és capaç d’induir canvis conformacionals en les cadenes polimèriques que donen lloc a una separació de fases. Sobre els materials en forma de scaffold porós amb porus esfèrics interconnectats s’han cultivat fibroblastos i endotelials. La compatibilitat de les cèl·lules endotelials s’ha mesurat en termes de viabilitat cel·lular, diferenciació endotelial adequada i funcionament. S’han dut a terme cultius de cèl·lules endotelials humanes primàries, HUVEC, i s’ha determinat si la seua morfologia i funció es veu afectada pel material. S’ha examinat l’adhesió i la proliferació d’aquestes, i també un marcador important d’activació endotelial, la E-selectina. S’ha avaluat si es mantenen els fenotips endotelials normals i les seues funcions observades in vivo per mitjà d’anàlisi dels 11 contactes cèl·lula-cèl·lula i la regulació de l’expressió gènica del marcador d’activació E-selectina quan s’afegeix un estímul (LPS). A més, com a possible aplicació d’aquests materials en una pròtesi de còrnia artificial, i atès que els fibroblastos de l’estroma de la còrnia (és a dir, els queratòcits) són de rellevància en la cicatrització de la còrnia, s’ha determinat com afecta l’hidrofilicitat a l’adhesió cel·lular de la línia de fibroblastos humans MRC-5, com a model cel·lular per a estudiar la disposició del citosquelet després de l’adhesió als diversos suports per mitjà de la detecció de F-actina. Així mateix, s’ha sembrat cèl·lules epitelials i se n’ha avaluat el comportament/funcionament cel·lular, ja que un dels requisits essencials perquè un implant de queratopròtesi tinga èxit és que es cree i mantinga una capa de cèl·lules epitelials que impedisquen entrar els bacteris a l’interior de l’ull i permeta la difusió la capa llagrimall de manera estable en el temps. Així, s’han analitzat paràmetres cel·lulars com l’adhesió, la proliferació i la viabilitat d’una línia de cèl·lules epitelials de conjuntiva humana, NHC, cultivada sobre substrats polimèrics amb diferents graus d’hidrofilicitat i càrregues elèctriques superficials, tot buscant quin grau d’hidrofilicitat permet l’epitelització del substrat i podria donar al material la flexibilitat i la hidrofilicitat necessària per a un millor contacte amb parpelles i llàgrima. Els resultats obtinguts s’han correlacionat amb l’adsorció i conformació d’una proteïna de la matriu extracel·lular, la fibronectina. 12 Abstract The need for bio-stable polymers for fabrication of prosthetic implants is evidenced, among other indicators, by the proliferation of currently marketed devices. The physico-chemical characterization and biological response of a set of bio-stable polymeric materials is the ultimate goal of this thesis. In this work we have synthesized various polymeric materials of the family of acrylates and methacrylates subtly varying surface characteristics, such as degree of hydrophilicity or distribution of electric charges. The procedure consisted of radical copolymerization of ethyl acrylate, EA, and 2hydroxyethyl acrylate HEA, and methacrylic acid, MAAc. We have characterized the materials in its dry state and in the presence of different water contents by differential scanning calorimetry, DSC, dynamic mechanical analysis, DMA, atomic force microscopy, AFM, dielectric analysis, DRS, equilibrium water content, EWC and surface energy, SE, pursuing the objective of ascertaining whether the water is able to induce conformational changes in the polymer chains leading to a phase separation. On materials in the form of spherical porous scaffold with interconnected pores fibroblasts and endothelial cells were cultivated. The compatibility of the endothelial cells was measured in terms of cell viability and suitable endothelial differentiation and function. Cultures were made from primary human endothelial cells, HUVEC, and it was determined whether their morphology and function were affected by the material. Adhesion and proliferation of HUVEC were examined, as well as an important marker of endothelial activation, E-selectin. We assessed the normal endothelial function phenotypes observed and maintained in vivo by analysis of the cellcell contacts and the regulation of gene expression of the activation marker E-selectin when a stimulus (LPS) was added. 13 Further, as potential application of these materials in a prosthetic artificial cornea, and since stromal fibroblasts of the cornea (i.e., keratinocytes) are relevant in the healing of the cornea the effect of hydrophilicity of substrate to adhesion of a human fibroblasts cell line, MRC-5, was determined. MRC-5 is a cell model to study the arrangement of the cytoskeleton after joining the different media by detecting F-actin. Epithelial cells have been seeded onto these substrates as well, evaluating their behaviour/cell function as one of the essential requirements for a successful keratoprosthesis implantation is that creating and maintaining a layer of epithelial cells that prevent bacteria enter into the eye and allow diffusion tear layer stably over time. Thus, cell parameters have been assessed such as adhesion, proliferation and viability of a line of human conjunctiva epithelial cells, NHC, grown on polymer substrates with different degrees of hydrophilicity and surface electric charges seeking what degree of hydrophilicity of the substrate allows epithelialisation and could give the material flexibility and hydrophilicity required for better contact with eyelids and tear. The results have been correlated with the adsorption and conformation of a protein of the extracellular matrix, fibronectin. 14 Capítulo 1. Introducción 15 1.1.- Ingeniería tisular La medicina ha tratado de resolver los problemas derivados de una falta o disfunción de un órgano mediante trasplantes, autoinjertos, prótesis artificiales y, recientemente, mediante la ingeniería tisular. La ingeniería tisular es una metodología multidisciplinar que persigue el objetivo de regenerar estructural y funcionalmente tejidos dañados mediante el uso de materiales apropiados. Estos materiales deben presentar características adecuadas para la adhesión y proliferación celular permitiendo a las células desarrollar sus funciones presentando un beneficio-riesgo adecuado para el receptor. Dentro de los materiales empleados en ingeniería tisular podemos encontrar materiales cerámicos, metálicos, poliméricos y/o compuestos a los que se les puede anclar/mezclar diferentes moléculas que promuevan una determinada respuesta celular. Los materiales poliméricos presentan algunas ventajas debido, fundamentalmente, a su gran variedad tanto de combinación de monómeros como de métodos de fabricación. Se suelen dividir, según su origen, en materiales poliméricos de origen natural o sintético. También en materiales biodegradables y no biodegradables en función de si el implante será reabsorbido por el huésped o permanecerá sin degradarse (prótesis) y, en cuanto a su arquitectura, en substratos o estructuras tridimensionales porosas (scaffolds). Mientras que una gran rama de la ingeniería de tejidos se centra en el desarrollo de materiales biodegradables, los materiales no biodegradables son necesarios para la sustitución/reemplazo de algunos tejidos. Un ejemplo de producto comercializado podemos encontrarlo en la prótesis de cornea artificial Alphacor®, donde la prótesis consiste en una lente transparente no porosa rodeada por un anillo macroporoso. Este material de anclaje es un polímero de origen sintético, no biodegradable y poroso [1]. El objetivo del 16 anillo poroso (scaffold) es promover que las células del estroma corneal del huésped crezcan en el seno de la estructura porosa y el conjunto quede fijado evitando la extrusión de la prótesis [1],[2]. Otro ejemplo de producto comercial disponible son los cementos óseos acrílicos empleados para fijación de prótesis de cadera, rodilla, etc., cuya función es la de actuar de interfase entre la prótesis y el hueso huésped, fijando la prótesis y transmitiendo las cargas externas de manera adecuada. Se trata en este caso de un polímero sintético, no degradable y no poroso. El objetivo de esta tesis es la caracterización físico-química así como la respuesta biológica de un conjunto de materiales poliméricos bioestables destinados a aplicaciones protésicas. 1.1.1.- Métodos de fabricación de scaffolds Para conseguir la regeneración tisular o la fijación protésica es necesario en algunas aplicaciones una estructura porosa tridimensional (scaffold), que debe simular la matriz extracelular en la cual las células puedan proliferar y funcionar adecuadamente. Existen diferentes métodos de fabricación de scaffods poliméricos que dan lugar a diferentes estructuras porosas. Por ejemplo, se pueden obtener estructuras porosas utilizando el método de disolución de partículas y/o la separación de fases inducida térmicamente; también se han fabricado scaffolds constituidos por fibras a escala nanométrica (electrohilado y autoemsamblado de nanofibras) y scaffolds fabricados mediante prototipado rápido (SFF techniques) [3]. El método de plantillas consiste en crear un negativo del soporte que se pretende fabricar con un material soluble en un disolvente que no dañe el material del soporte. La plantilla puede estar formada por partículas esféricas o cilíndricas sinterizadas, o producida por prototipado rápido, en cualquier 17 caso interconectadas entre sí, porque las superficies de conexión en la plantilla darán lugar a la interconexión entre poros en el scaffold. Esa plantilla se embeberá en un monómero que se polimeriza, o bien en una disolución de polímero o bien en un polímero fundido. En nuestro caso, será un monómero el que se introducirá en la plantilla y se polimerizará in situ, como se explicará en detalle en las secciones siguientes. Cuando la plantilla se disuelve, se genera una estructura porosa negativo de la plantilla confeccionada [4]-[17]. También se emplean partículas dispersas de porógenos en una disolución polimérica que, aunque no están inicialmente adheridas unas con otras, se unen durante el proceso de eliminación del disolvente dando lugar igualmente a poros interconectados. En este método, se han utilizado partículas de materiales muy diversos: sales, azúcar, partículas de agua congelada, microesferas poliméricas o de parafinas y otros [18]-[38]. La separación de fases inducida térmicamente explota el hecho de que una mezcla polímero-solvente sea completamente miscible a una determinada temperatura pero se separe en fases a otra distinta. Al separarse en fases (una rica en solvente, otra rica en polímero) se extrae el solvente mediante algún tratamiento térmico (sublimación, evaporación, etc.) [39]. En el electrohilado (electrospinning) se disuelve un polímero dipolar y se le hace pasar por una boquilla muy fina. Entre la boquilla y un plato colector separado una cierta distancia de la boquilla se establece un fuerte campo eléctrico externo lo que induce a cambiar la posición de los dipolos y al estiramiento de la solución en el proceso, creando hilos de tamaño nanométrico o micrométrico en función de las condiciones de operación, que se depositan sobre la placa colectora [40]. El autoensamblaje molecular de nanofibras consiste en utilizar las propiedades químicas de moléculas individuales para hacer que dichos componentes se auto-organicen o auto-ensamblen en cierta conformación útil [41]. 18 La creación de materiales poliméricos porosos mediante prototipado rápido (SFF fabrication techniques) consiste en la generación de estructuras tridimensionales mediante sofisticada maquinaria que deposita el material capa a capa, proporcionando una enorme versatilidad y control de la geometría de la pieza diseñada [42]. 1.1.2.- El método de disolución de plantilla En este trabajo se ha utilizado el método de la disolución de plantilla hecha por partículas esféricas. El método consistió en preparar unas plantillas formadas por esferas poliméricas sin entrecruzar que fueron sometidas a una sinterización mediante presión y temperatura para unir dichas partículas. Por otro lado, se prepararon una serie de soluciones monoméricas con un agente entrecruzador. La plantilla de porógeno se embebió en la solución monomérica rellenado los huecos de la plantilla. Se procedió a polimerizar la solución monomérica generándose un polímero entrecruzado. Para eliminar la plantilla de esferas se buscó un buen solvente que hinchara la red polimérica y que disolviera simultáneamente al porógeno. Una vez disuelta la plantilla se procedió al intercambio progresivo del solvente. Este paso es crítico para evitar el colapso de los poros ya que si el intercambio no se produce a una velocidad lenta, donde las cadenas de polímero puedan progresivamente adaptarse a las nuevas condiciones, la red colapsa [15]. 1.1.3.- Scaffolds de acrilatos y metacrilatos Los polímeros usados en este trabajo pertenecen a la familia de los acrilatos y metacrilatos cuyas propiedades se describen en los siguientes apartados. Estos materiales se han sintetizado en forma de scaffolds bioestables (no degradables) por diversos grupos para algunas aplicaciones de 19 ingeniería tisular. Por ejemplo, en [43] se sintetizó un copolímero entrecruzado y se evaluó su potencial pro-angiogénico in vivo. En [44], los autores describen la fabricación de un soporte poroso de PMMA con aplicaciones en el campo de la ingeniería tisular ósea modificando parámetros para controlar el módulo elástico y la permeabilidad. También, se han fabricado soportes de dichas familias para cultivo celular de fibroblastos y endoteliales [11], osteoblastos [45]. Asimismo, se han utilizado como anillo de anclaje de prótesis de córnea [1] y como soportes para la regeneración de dentina [8]-[10], [17]. 1.2.- Estructura y propiedades físicas de los soportes empleados 1.2.1.- Sobre los materiales empleados en estado seco 1.2.1.1.- Estructura química Los polímeros de este trabajo se encuadran en la familia de los acrilatos y metacrilatos cuya estructura química de la unidad monomérica se muestra en la Figura 1.1. H R1 · C C· H C O O R2 Figura 1.1.- Unidad monomérica de la familia de los acrilatos y metacrilatos Se puede observar que de cada unidad estructural cuelga un grupo lateral éster (COO) lo que les confiere a estos polímeros una asimetría. Además, 20 estos polímeros son atácticos –los grupos laterales se sitúan al azar- lo que impide que los polímeros resultantes de esta familia puedan cristalizar, siendo todos ellos, por tanto, materiales amorfos. Si el grupo R1 es un hidrógeno, H, el proceso de polimerización dará lugar a la serie de acrilatos mientras que si R1 es un grupo metilo, CH3, se obtendrá la familia de los metacrilatos. La estructura química del grupo R2 influye notablemente en las propiedades del polímero, en particular, en su carácter hidrófilo. Así, por ejemplo, si R1 es el grupo H y R2 es el grupo etilo, CH2CH3, el polímero obtenido será de poli(acrilato de etilo), PEA, un polímero hidrófobo. Por el contrario, si R1 es el grupo H y R2 es el grupo hidroxietilo, CH2CH2OH, se obtendrá el poli(acrilato de hidroxietilo), PHEA. El PHEA exhibe un carácter hidrófilo debido al grupo hidroxilo, OH, en la cadena lateral que interactúa con las moléculas de agua a través de enlaces de hidrógeno. En la Tabla 1.1 se muestran algunos polímeros de la familia de los acrilatos y metacrilatos. Concretamente, el PEA, PHEA y copolímeros P(EA-co-HEA) y P(EA-co-MAAc) que han sido usados en este trabajo. 21 Grupo R1 Polímero Grupo R2 H H ( C H CH2CH3 C ) H n C O O CH2CH3 poli(acrilato de etilo), PEA H H ( C C ) H H CH2CH2OH n C O O CH2CH2OH poli(acrilato de hidroxietilo), PHEA CH3 H H CH3 ( C C ) H n C O O H poli(ácido metacrílico), PMAc Tabla 1.1.- Polímeros de la familia de los acrilatos y metacrilatos 22 Los monómeros de las series de los acrilatos y metacrilatos polimerizan fácilmente por vía radical utilizando iniciadores térmicos, fotosensibles, pares redox y otros que pueden generar radicales libres capaces de abrir el doble enlace del monómero e iniciar una reacción en cadena. Si uno de los co-monómeros tiene una funcionalidad mayor de dos, es decir, si tiene más de un doble enlace en la cadena principal, se forma una red polimérica en la que todas las cadenas están enlazadas. El material así obtenido es una red polimérica insoluble. Los polímeros utilizados en este trabajo fueron obtenidos por polimerización por via radical utilizando benzoína, que es un fotoiniciador que se descompone bajo la luz ultravioleta, produciendo dos radicales libres capaces de romper el doble enlace de una molécula de monómero e iniciar la polimerización. Los polímeros y copolímeros utilizados en este trabajo se han obtenido usando etilenglicol dimetacrilato, EGDMA, como agente de reticulación, cuya estructura química se muestra en la Figura 1.2. CH2 O CH3 CH3 C C C C OCH2CH2O CH2 O Figura 1.2.- Estructura química del entrecruzador, EGDMA La presencia de dos unidades de metacrilato en la molécula del EGDMA permite formar enlaces con funcionalidad cuatro (hasta cuatro cadenas de polímero pueden empezar o terminar en la unidad de EGDMA en el polímero). Por lo tanto, el material obtenido por la co-polimerización de una mezcla de EGDMA y otros monómeros de acrilato o metacrilato es una red 23 de polímero que puede hincharse en varios disolventes, pero es insoluble en cualquiera de ellos. En la Figura 1.3 se muestra esquemáticamente una red polimérica de PEA y el detalle de un punto de entrecruzamiento. Figura 1.3.- Representación de una red polimérica de PEA entrecruzada con EGDMA Los polímeros y copolímeros producidos a partir de estos monómeros por polimerización radical son atácticos y, consecuencia de ello, no son capaces de cristalizar debido a la falta de regularidad en la cadena principal; los grupos laterales están situados al azar a un lado u otro de la cadena principal y, por tanto, estos polímeros y copolímeros son amorfos ya que el orden molecular de largo alcance no existe y existe sólo en el corto alcance. Por tanto, debe considerarse a estos materiales como líquidos. En este trabajo se ha sintetizado una serie de redes de homopolímero o copolímero con diferentes grados de hidrofilia y cargas eléctricas superficiales tratando de variar mínimamente su estructura química. Los copolímeros se producen mediante polimerización radical a partir de una mezcla líquida de dos monómeros, incluyendo un agente de reticulación (de hecho, un tercer comonómero con funcionalidad mayor que dos) y el iniciador. Cuando la 24 molécula de iniciador se descompone para formar dos radicales libres e iniciar el crecimiento de una cadena de polímero la probabilidad de que uno o los otros co-monómeros se una a la cadena en crecimiento depende de la reactividad de los monómeros. El índice de reactividad de uno de los comonómeros, A, mide la probabilidad relativa de que un tipo de monómero A se una a una cadena en crecimiento que termina en una unidad monomérica A. En el caso de la copolimerización de EA e HEA el índice de reactividad del EA es 0.5 [46], lo que significa que es igualmente probable que el siguiente segmento de una cadena en crecimiento cuyo último elemento era una unidad monomérica de EA sea un EA o una unidad monomérica de HEA. Por el contrario, el índice de reactividad para el HEA es 0.97 [46], es decir, es muy probable que una cadena en crecimiento cuyo último elemento era el HEA se le una otra unidad de HEA. El resultado de esta situación es que cuando la polimerización progresa, el consumo de monómero HEA en la mezcla monomérica sea más rápido que el de las moléculas de EA. Si la reacción se detiene antes de alcanzar la conversión completa de los monómeros, la composición de las cadenas de copolímero serán más ricas en HEA de lo esperado a partir de la composición de la mezcla monomérica. Sin embargo, si la reacción alcanza la conversión completa, el resultado es una cadena de polímero que contiene dos bloques, uno de ellos que es más rico en HEA y el otro que se ha formado al final de la polimerización -cuando el HEA en la mezcla líquida está casi agotado-, que es más rico en EA o incluso puede consistir en un bloque de poli(acrilato de etilo) [47],[48]. 1.2.1.2.- Propiedades físicas Probablemente, la característica principal en cuanto a propiedades físicas que diferencia un polímero de otros materiales, tales como metales o materiales cerámicos, es la movilidad conformacional. Las posibilidades 25 rotacionales del enlace covalente C-C de la cadena principal permite que ésta pueda adoptar un número enorme de diferentes conformaciones espaciales. Esta capacidad explica por qué el polímero puede llegar a grandes deformaciones elásticas o plásticas cuando se aplica una fuerza externa. Además, la movilidad conformacional también permite que las redes de polímero se hinchen en presencia de un buen solvente aumentando su volumen hasta varias veces su volumen inicial. La alta expansión térmica y los altos coeficientes de compresibilidad en materiales poliméricos también se deben a la movilidad conformacional [49],[50]. La respuesta del material a una acción externa tal como la aplicación de una fuerza externa o un cambio repentino de la temperatura implica reordenamientos cooperativos de los segmentos de polímero. El movimiento de un segmento de polímero necesita el precedente o el movimiento simultáneo de otros grupos situados en su vecindad. Estos movimientos no son instantáneos y, además, son disipativos debido a la fricción interna. La respuesta del material polimérico se retarda con respecto a la acción externa, una característica que se ha llamado comportamiento viscoelástico en el caso de la respuesta mecánica del material [49]. La velocidad de respuesta (por lo general, se caracteriza por medio de los tiempos de relajación) depende exponencialmente de la temperatura. Cuando la temperatura disminuye, la contracción necesaria para mantener el sistema en equilibrio termodinámico se puede lograr por medio de la reorganización correspondiente de las cadenas de polímero que, a altas temperaturas tiene lugar lo suficientemente rápido. Por el contrario, cuando disminuye la temperatura, la velocidad de los reordenamientos conformacionales disminuye y las cadenas de polímero no son capaces de adoptar las nuevas condiciones externas; el material se encuentra en esas condiciones en un estado de no-equilibrio. El volumen y otras variables termodinámicas como la entalpía y la entropía, superan los valores de equilibrio. El material se dice que está en el estado vítreo, en el que el aspecto externo es el de un sólido ya que la movilidad conformacional 26 se inhibe y los segmentos de polímero casi no pueden cambiar su posición en respuesta a cualquier acción externa. Aunque la respuesta mecánica (y otras propiedades físicas) del material se parecen a las de un sólido, el orden molecular de largo alcance sigue ausente y, como consecuencia, el material en estado vítreo debe ser considerado un líquido de muy alta viscosidad. La transición vítrea es un fenómeno cinético que se revela por un cambio de pendiente en el volumen específico, la entalpía o entropía cuando se representa frente a la temperatura durante un ensayo de enfriamiento como se muestra en la Figura 1.4 [51]. La temperatura de transición vítrea, Tg, se puede definir como la temperatura a la que la extrapolación de la entalpía específica, o volumen específico correspondiente al líquido (a temperaturas por encima de la de transición vítrea) y el vidrio (a temperaturas inferiores a la transición) se v, h, s cruzan. Tg T Figura 1.4.- Representación del fenómeno de la transición vítrea 27 Dado que los movimientos conformacionales cooperativos alcanzan distancias de unos pocos nanómetros o pocas decenas de nanómetros en la temperatura de transición vítrea [52], cuando un sistema es heterogéneo, es decir, separado en fases de diferente composición, y el tamaño de los diferentes dominios de cada fase son más grandes que estos pocos nanómetros o decenas de nanómetros, aparecerán dos temperaturas de transición vítrea en el sistema, correspondientes a las temperaturas de los componentes puros. Por contra, en el caso de una mezcla homogénea de dos componentes poliméricos aparecerá una sola temperatura de transición vítrea en el intervalo de temperatura entre las temperaturas de transición vítrea de los componentes puros. Se utilizará este criterio para detectar la separación de fases en nuestros copolímeros. Como se ha comentado anteriormente la diferencia entre las familias de acrilatos y metacrilatos es el hidrógeno o el grupo -metilo, respectivamente, en la posición R1. Este grupo tiene una gran influencia en la temperatura de transición vítrea debido al impedimento estérico que impone el grupo metilo a la rotación alrededor de los enlaces covalentes C-C de la cadena principal. Por tanto, la movilidad conformacional es mucho más rápida en acrilatos que en metacrilatos. En términos generales, un polímero de metacrilato tiene una temperatura de transición vítrea alrededor de 100 K por encima de la del acrilato con la misma cadena lateral. La posibilidad de tener movimientos conformacionales cooperativos puede ser también detectada en la respuesta mecánica. Cuando se aplica una fuerza a un material polimérico (por ejemplo, una fuerza aplicada longitudinalmente a una barra de polímero con pequeña sección transversal) la respuesta consiste en una deformación instantánea seguida de un aumento continuo de la longitud de la barra que se aproxima en el tiempo (si no hay flujo irreversible) a un valor de equilibrio de acuerdo con el nuevo estado de tensión. A temperaturas por encima de Tg la movilidad conformacional 28 permite grandes deformaciones de polímero cuando se aplica una fuerza externa, es decir, el material posee un módulo elástico bajo. El proceso de relajación se llama relajación principal o . Cuando la temperatura del sistema está por debajo de la temperatura de transición vítrea la deformación del polímero es de hasta tres órdenes de magnitud más pequeña que cuando el polímero está por encima de la Tg y, por lo tanto, el módulo elástico aumenta hasta tres órdenes de magnitud. Este proceso se debe a que la velocidad de respuesta del material cuando se aplica una fuerza externa disminuye drásticamente a temperaturas inferiores a la de transición vítrea [49]. Merece la pena señalar que la relajación mecánica principal y la transición vítrea no son el mismo proceso aunque ambos se originan por el mismo fenómeno, es decir, la desaceleración de los movimientos conformacionales en un intervalo de temperaturas estrecho. Del mismo modo que en un sistema separado en fases se observan dos Tg, también se observan dos relajaciones dinámico-mecánicas. A temperaturas por debajo de la transición vítrea, en el estado vítreo, todavía pueden producirse movimientos locales, tales como rotaciones de los grupos laterales. La diferencia entre estos movimientos respecto a los movimientos conformacionales radica en que los movimientos locales no son cooperativos, es decir, el movimiento de un grupo no necesita el movimiento de otros grupos adyacentes. Cuando la temperatura desciende estos movimientos son impedidos en los sucesivos procesos de relajación, a los que se denomina relajaciones secundarias. El comportamiento de relajación viscoelástico de los materiales poliméricos se caracteriza a menudo mediante ensayos dinámico-mecánicos. Una descripción completa de la técnica se puede encontrar en [49], [50], [53]: una muestra de polímero se somete a una deformación sinusoidal a una frecuencia fija mientras se mide la fuerza para mantener esta deformación, 29 que también es sinusoidal. La onda de fuerza medida se desplaza con respecto a la deformación inducida debido a la respuesta retardada del material. La relación entre la fuerza y la deformación permite determinar el módulo elástico complejo, cuya parte real, E', relaciona las amplitudes de onda de tensión y deformación. La parte real del módulo elástico tiene que ver con el almacenamiento de energía por ciclo de deformación. La componente imaginaria del módulo elástico complejo, E'', es proporcional a la energía disipada por ciclo de deformación debida a la fricción interna. La tangente del ángulo de desfase entre la fuerza y la deformación, tan δ, denominada tangente de pérdidas, es el cociente E''/E'. Un proceso de relajación puede ser identificado por una caída en E' en un cierto intervalo de temperaturas acompañado de un pico en tan δ o E'', en los gráficos de E', E'' y tan δ vs T a una frecuencia constante. El comportamiento de la relajación de acrilatos y metacrilatos se ha descrito en detalle en varias referencias clásicas [49],[50],[53]. Ambos, poliacrilatos y polimetacrilatos presentan además de la relajación principal, una relajación secundaria, denominada , atribuida a las rotaciones del grupo carbonilo -COOR2 alrededor de su unión con la cadena principal. La temperatura a la que aparece esta relajación secundaria es independiente de la longitud del grupo R2 [50]. Si R2 contiene más de cuatro enlaces covalentes, los movimientos locales dentro de esta cadena lateral producen otra relajación secundaria, de baja temperatura, denominada . Por ejemplo, la relajación es muy evidente en el PHEA, mientras que está ausente en el PEA (Figura 1.5). 30 H H ( C C ) H n C O O CH2CH2OH Figura 1.5.- Relajaciones secundarias del PHEA Como ocurre con la relajación , la relajación se muestra a temperaturas más altas en polimetacrilatos que en poliacrilatos debido a la presencia del grupo -metilo. La relajación γ tiene lugar a las mismas temperaturas en ambas familias, ya que los movimientos internos en la cadena lateral no están influenciados por la rigidez de la cadena principal. El efecto de una longitud cada vez mayor del grupo lateral alquilo en los espectros de relajación de acrilatos y metacrilatos implica un descenso en la temperatura de la relajación principal [54] y disminuye la intensidad de la relajación secundaria β, pero no altera su temperatura. Por lo tanto, la magnitud de la relajación β del poli(acrilato de metilo), PMA, es mayor que la del poli(acrilato de etilo), PEA, y esta última mayor que la de poli(acrilato de propilo), PPA; acrilatos con cadenas laterales más largas que los PPA no exhiben la relajación β debido al impedimento estérico. La espectroscopía dieléctrica es análoga a la técnica dinámico-mecánica [55]. El papel de la fuerza es interpretado por la aplicación de un campo eléctrico oscilante a la muestra. La técnica permite determinar la evolución con la frecuencia y la temperatura de la permitividad dieléctrica compleja, *= '+i '', donde i es - 1 . Los procesos de relajación aparecen debido a la orientación de los dipolos eléctricos permanentes presentes en las cadenas de polímero en la dirección del campo aplicado y, en consecuencia, la intensidad relativa de los diferentes procesos de relajación puede ser diferente a la que 31 aparece en los experimentos dinámico-mecánicos debido a la posición de los dipolos en la cadena. En el PHEA, de manera diferente que en el PEA, el movimiento del grupo alquilo es dieléctricamente activo, ya que orienta el dipolo-CH2CH2OH, lo que explica la alta intensidad de la relajación [56]. 1.2.1.3.- Elasticidad de redes poliméricas Un hidrogel es una red polimérica hinchada de agua en estado de goma. Su análisis tiene en cuenta la teoría de la elasticidad de gomas así como de la teoría de las disoluciones poliméricas. La primera permite correlacionar algunas cantidades medibles experimentalmente con parámetros estructurales de la red. Efectivamente, a través de la energía libre de Gibbs se puede obtener datos del número de cadenas, peso molecular promedio entre puntos de entrecruzamiento, etc. El cálculo de la energía libre se realiza a través de la entropía de la red, que depende de la estadística configuracional de las cadenas. Parámetros dimensionales de redes poliméricas El cálculo de los parámetros dimensionales de redes poliméricas, la teoría de la elasticidad de caucho así como la teoría de las disoluciones poliméricas puede encontrarse en [51]. La media cuadrática de la distancia entre el inicio y el final de cadena de cadenas libremente enlazadas se puede obtener mediante: r2 random = nl 2 , siendo n el número de unidades monoméricas de una cadena y l la longitud de la cadena si n>10 y r<<r máx Para configuraciones en las que las cadenas estén perturbadas se puede 32 efectuar la corrección a través de r2 0 = C r2 random = Cnl 2 donde C varía con n para cadenas cortas y tiende a C∞ para cadenas largas. El parámetro C depende de la interacción red-solvente. Un buen solvente de una red tendrá valores de C elevados y la red se expandirá mientras que un mal solvente hará que la red se contraiga y el parámetro C será bajo. Imaginemos la red polimérica como una malla en la que las cadenas están unidas a través de los puntos de entrecruzamiento. Si Nx es el número de puntos de entrecruzamiento, Nx = NA mx Mx siendo NA el número de Avogadro, mx la masa de entrecruzador y Mx su peso molecular. Ya que la funcionalidad del entrecruzador empleado en este trabajo es 4 y la del monómero 2 el número de cadenas, N c , es el doble del número de puntos de entrecruzamiento, N x . El número medio de unidades monoméricas en una cadena, ν= , será: NA m u N c Mu Ecuación 1.1 siendo mu la masa de monómero y Mu el peso molecular de una unidad monomérica. Así, el peso molecular promedio entre uniones (una cadena): M c = νM u Ecuación 1.2 33 Y también podemos obtener el tamaño de malla o distancia entre nudos, ζ = r2 1 0 2 1 1 = C 2 ν 2l Ecuación 1.3 el parámetro dimensional más característico de la red polimérica. Teoría de la elasticidad de gomas Imaginemos un material polimérico en estado de goma que es sometido a una deformación. Entonces, la energía libre de Gibbs puede escribirse como ΔG = ΔH - TΔS = Δ(U + pV) - TΔS ≈ -TΔS , donde la deformación se considera isoterma e isócora ( ΔU = 0 e ΔV = 0 ). Si se asume que el cambio de entropía del sistema es el cambio de entropía de una cadena debido al proceso de deformación, Δs(λ x , λ y , λ z ) multiplicado por el número de cadenas, Nc, y teniendo en cuenta la teoría de Maxwell-Boltzmann, en la que el cambio de entropía está relacionado con la probabilidad de encontrar un final de cadena en (x,y,z) cuando el otro está en (0,0,0), se deduce que: ΔS ΔG ΔG 1 kN c (λ 2x λ 2y λ 2z - 3) 2 1 -TΔS kTN c (λ 2x λ 2y λ 2z 2 1 RTn c (λ 2x λ 2y λ 2z - 3) 2 3) ó Si, además, la deformación es isótropa: 1 kTN c (3 λ 2 - 3) = 2 3 3 = kTN c (λ 2 - 1) = RTn c (λ 2 - 1) 2 2 ΔG = -TΔS = 34 donde se puede comprobar que el incremento de energía libre debido a un proceso de deformación de una red polimérica está relacionado con el número de cadenas que conforman la red, que es un parámetro estructural. Relaciones tensión-deformación Supongamos un material sometido a una carga unidireccional, f, cuya longitud inicial es L0. Aplicando la primera ley de la termodinámica y suponiendo un proceso isentrópico (adiabático y reversible): dU δQ - δW dG dU TdS - pdV dH - d(TS) pdV TdS - pdV → dG pV) - d(TS) Vdp - TdS - SdT fdL fdL d(U fdL → pdV Vdp - TdS - SdT → Vdp - SdT A T,p constantes dG fdL → G(L) - G(L 0 ) L fdL L0 Bajo condiciones de carga constante G(L) = f(L - L 0 ) con G(L 0 ) 0. El límite elástico E se define como la variación de la tensión soportada por el sistema respecto de deformación sufrida por el mismo cuando ésta tiende a 0. Por tanto, 35 f dσ dσ A E lim lim lim ε → 0 dε λ → 1 dλ λ→ 1 dλ d 1 G d 1 G lim lim λ → 1 dλ A λ → 1 dλ A L L d 3 nc RT V2 3 ρ RT Mu Ecuación 1.4 lo que permite relacionar el límite elástico E, medible experimentalmente, con el número de cadenas de la red polimérica. Además, ΔG = 3 1 RTn c (λ 2 - 1) = EV(λ 2 - 1) 2 2 Ecuación 1.5 1.2.2.- Interacción con el agua de los materiales empleados 1.2.2.1.- Mezclas polímero-solvente Sea una mezcla de dos componentes en la que designaremos con el subíndice 1 al agua y 2 a la red polimérica. La mezcla puede formar una única fase si el agua se encuentra homogéneamente distribuida en la red polimérica mientras que el sistema estará separado en dos fases si el agua es capaz de formar una fase independiente de la mezcla agua-red polimérica. Esto se puede comprobar en calorimetría diferencial de barrido, DSC, ya que cuando se compara el comportamiento de la red polimérica en estado seco (xerogel) con estados en los que se ha hinchado la red en agua con diferentes contenidos de la misma se observa una plastificación, es decir, una disminución progresiva de la temperatura de transición vítrea. Si la red 36 polimérica se satura debido a las propiedades elásticas de la misma, entonces el agua es capaz de permanecer en su seno en dos fases diferentes, una de ellas como componente de la mezcla red polimérica-agua y otra como agua pura. En una mezcla red-agua (2 componentes, 1 fase) el número de variables intensivas independientes para que el sistema quede determinado viene dado por la regla de las fases de Gibbs, que en este caso conduce a tres grados de libertad (T, p y fracción molar ó másica de uno de los componentes). Si el sistema es monofásico (red-agua) las propiedades del mismo deben calcularse mediante las propiedades parciales, ya que el sistema es una mezcla homogénea. Las propiedades parciales están relacionadas mediante la ecuación de Gibbs-Duhem-Margules (GDM) a T, p constantes y permite obtener una de ellas mediante el conocimiento de la otra propiedad parcial. Podemos definir el incremento de mezcla de una propiedad como el valor que toma la propiedad en la mezcla (G, H, U, S, V) menos el valor que tendría si consideráramos aditividad en las mismas condiciones de T,p en equilibrio, lo que conduce en el caso de la energía libre de Gibbs al criterio de miscibilidad de componentes. Efectivamente, si el incremento de mezcla de la energía libre de Gibbs es negativo el sistema será miscible mientras que si es positivo el sistema estará separado en fases. Como dijimos antes, puede ocurrir que el sistema, para cantidades elevadas de agua, forme dos fases, una mezcla de red polimérica-agua y otra de agua pura. Este fenómeno ocurrirá siempre que el polímero alcance su capacidad de absorción máxima, momento en el cual aparecerá una nueva fase de agua en bloque, que puede ser observada mediante calorimetría, por ejemplo, ya que cristalizará y fundirá como el agua pura. En este caso, los potenciales químicos del agua en la fase red-agua y del agua pura serán iguales y, por tanto, la actividad del agua en ambas fases igual a la unidad. 37 1.2.2.2.- Hinchamiento de redes poliméricas Imaginemos una red polimérica que debido al efecto del hinchado en presencia de un solvente se expande de manera isótropa hasta alcanzar un nuevo estado. El cambio de energía libre de Gibbs debido a la expansión de la red polimérica exclusivamente será según la Ecuación 1.5, ΔG 2w = 1 EV2 (λ 2 - 1) 2 Si definimos el grado de hinchamiento como V2 V2 V V2 (λ 3 ) G2 w - G 2 1 EV2 ( 2 2 ΔG2 w ΔS 2w 1 → λ λ3 2 2 3 3 - n c R( 2 2 3 3 3 n c RT( 2 - 1) 2 1 2 2 2 3 - 1) - 1) Cuando una red polimérica se hincha por la interacción con un solvente el incremento de energía libre se puede ver como la superposición de dos contribuciones; de una parte el incremento de energía libre de Gibbs debido a la expansión de la red y de otro lado el incremento de energía libre como consecuencia del proceso de mezcla en una red expandida. Así, Δ m G = G - G1 - G 2 = [G(T, p, w 1 ) - G1 (T, p) + G 2 w (T, p)] - [G 2 w (T, p) - G 2 (T, p)] = = Δ m G sol + ΔG 2 w El cálculo de Δ mGsol se realiza mediante la teoría de las disoluciones de Flory-Huggins. 38 1.2.2.3.- La teoría de Flory-Huggins En la teoría de Flory-Huggins el incremento de mezcla de energía libre consecuencia de un proceso de mezcla entre un polímero y un solvente viene determinado por la siguiente expresión: Δ mGsol donde i RT(n1 ln n2 ln 1 χ 12 n1 2 ) 2 representa la fracción entre el volumen del componente i antes de ser hinchado y después y χ es el parámetro de interacción polímero12 solvente. Así, Δ mG G - G1 - G 2 RT (n 1 ln Como μ ˆ1(T, p, w 1 ) 1 n 2 ln μ1(T, p) χ 12 n 1 2 2 ΔG 2 w 3 nc( 2 ) 2 2 3 - 1) RT ln â1(T, p, w 1 ) y, por definición, μˆ1(T, p, w 1 ) - μ1(T, p) → ln â 1 Δ m G sol m G w1 ln(1 - 2 ) 2 → RT ln â 1 χ 12 22 ) v1 m nc V2 G w1 1 2 → 3 La ecuación anterior es la isoterma de absorción de Flory-Huggins. Cuando la actividad del agua (componente 1) es 1 (la red inmersa en agua líquida o en vapor saturado) se obtiene la ecuación de Flory-Rhener, ln â 1 0 ln(1 - ** 2 ) ** 2 χ 12 ** 2 2 ) v1 nc V2 1 2 3 Ecuación 1.6 expresión que permite relacionar nc/V2 (previamente obtenido a través de E) con el parámetro de interacción 12 . 39 1.2.2.4.- Hidrogeles A bajos contenidos de agua (actividad del agua <1) toda el agua está homogéneamente mezclada con las cadenas poliméricas formando así una mezcla de los componentes cuyas propiedades se pueden determinar fijando p, T y w1. La presencia de agua en bloque separada dentro del hidrogel tiene lugar cuando se dan dos condiciones: (i) la actividad del agua en la mezcla se iguala a 1 y (ii) la presencia de cavidades en la red polimérica que permitan alojar la fase agua pura. La capacidad de sorción de un gel depende principalmente del tamaño de malla, ya que éste indica la distancia media entre puntos de entrecruzamiento. Si uno imagina dos redes poliméricas con la misma cantidad de unidades monoméricas, pero una de ellas está poco entrecruzada, mientras que la otra está muy entrecruzada puede concluirse que la energía elástica en la expansión de la red más entrecruzada es mayor y por eso se absorbe menos agua, tal y como se muestra en la ecuación de Flory-Rehner, Ecuación 1.6. Por tanto, la manera de controlar el tamaño de malla es, principalmente, a través del entrecruzador. 1.2.2.5.- La transición vítrea en mezclas homogéneas Ya hemos indicado anteriormente que, cuando la temperatura de un polímero líquido amorfo disminuye, la movilidad molecular se reduce y, en cierto intervalo, tiene lugar la transición vítrea, que da lugar a estados donde la movilidad de las moléculas es prácticamente nula, como en el estado sólido, pero sin orden cristalino. Se dice que el material está en estado vítreo. Desde el punto de vista termodinámico el estado vítreo es un estado de no equilibrio (es un líquido subenfriado). La temperatura de transición vítrea, Tg, es una característica importante de los materiales amorfos, como 40 los utilizados en este trabajo, debido al cambio que se produce en sus propiedades, tales como el módulo mecánico, propiedades dieléctricas, conductividad, índice de refracción, etc., al atravesar la transición. De hecho, la transición vítrea refleja la existencia de un proceso de relajación estructural. En efecto, cuando el material sufre un cambio de temperatura y está en estado de líquido los cambios conformacionales son prácticamente instantáneos mientras que en estado vítreo éstos están inhibidos y el material se comporta como un sólido. En el intervalo de temperaturas de la transición vítrea nos encontramos ante el cambio de comportamiento ya que por debajo de ella el material necesita un tiempo para encontrar el equilibrio, mayor cuanto más baja es la temperatura. En el caso de sistemas multicomponente como son las mezclas se puede obtener una expresión que relacione la Tg de la mezcla en función de la cantidad de uno de los componentes en la mezcla (en nuestro caso, el agua). Partiendo de la igualdad de entropías en los estados vítreo y líquido del sistema en la temperatura de transición vítrea y asumiendo que: ΔCp,solv Clp,solv - Cgp,solv e ΔCp, xerogel = Clp, xerogel - Cgp, xerogel son proporcionales a la temperatura se llega a la ecuación de CouchmanKarasz [57]: Tg = w xerogel ΔC p, xerogel Tg,xerogel + w solvente ΔC p,solvente Tg.solvente w xerogel ΔC p, xerogel + w solvente ΔC p,solvente Ecuación 1.7.- Ecuación de Couchman-Karasz que predice un descenso monótono de Tg de la mezcla desde la Tg del polímero seco conforme wxerogel crece. Esta ecuación ha sido empleada para predecir la transición vítrea de mezclas de P(EA-co-HEA) con diferentes contenidos de agua. 41 1.2.2.6.- La interacción hidrófoba La ecuación de Couchman-Karasz no tiene en cuenta los efectos de las interacciones específicas –en caso de existir- que se producen entre el solvente y segmentos de la cadena polimérica, que induce cambios conformacionales que modifican la temperatura de transición vítrea predicha por la ecuación. La interacción hidrófoba es una de esas interacciones específicas y hace referencia a los cambios conformacionales en un polímero que son inducidos por la interacción con un solvente; tiene como resultado la agregación de grupos hidrófobos de la cadena polimérica en presencia de agua para minimizar su exposición a la misma lo que da lugar a débiles interacciones inter e intramoleculares que producen la reorganización del conjunto. La existencia de este tipo de interacción es una consecuencia de las dificultades de mantener la red de enlaces de hidrógeno del agua alrededor de los conjuntos hidrófobos. El mantenimiento de la red de enlaces de hidrógeno tiene que ver con el tamaño del soluto. Si el volumen de exclusión de la molécula hidrófoba es suficientemente pequeño, su presencia en agua no implica la rotura de la red de enlaces de hidrógeno. Pero la situación es diferente cuando se trata de grandes solutos hidrófobos, donde es imposible que exista una red de enlaces de hidrógeno completa con el fluido que lo rodea y una parte de las opciones de enlaces de hidrógeno se pierden cerca de la superficie [58]. La tendencia de las partículas a agregarse en presencia de agua se explica mediante el balance entre dos fuerzas opuestas. Una de ellas viene determinada por la energía libre de mezcla del sistema; si la energía libre del sistema homogéneo es mayor que la que tendría el sistema separado en dos fases, dicho sistema se separará. Ahora bien, para obtener el sistema separado en fases se debe crear una nueva superficie y, tal y como predice la teoría de la nucleación, es necesaria la formación de un núcleo crítico [59]. 42 Las interacciones hidrofóbas se han utilizado para explicar los cambios conformacionales de cadenas individuales en disoluciones poliméricas, que no podían ser explicadas por la interacción convencional entre moléculas [60]. Los principios que aplican a solutos puramente hidrofóbos también se aplican a las moléculas que contienen algunas unidades hidrófilas, pero tienen lugar efectos entrópicos adicionales ya que las unidades hidrófobas e hidrófilas se tienen que acomodar en presencia de agua [61]. Cuando hay grupos hidrófilos e hidrófobos presentes en una molécula, la asociación de moléculas de agua alrededor de las partes hidrófobas cambia la conformación de las cadenas de polímero pasando de una estructura al azar a una colapsada [61]-[68]. Algunos copolímeros con grupos hidrófobos e hidrófilos cuando se disuelven en agua por encima de una concentración crítica tienden a asociarse por interacción hidrófoba intermolecular, generando una red transitoria a través de asociaciones de los grupos hidrófobos [69]-[73]. Las distribuciones de unidades monoméricas a lo largo de la cadena es un factor importante a tener en cuenta cuando se trata de las transiciones de fase en geles ya que podría conducir a la separación de fases como consecuencia de la organización molecular de la cadena de copolímero. Sin embargo, incluso en el caso de un copolímero al azar, la segregación en nanofases podría tener lugar como consecuencia de las asociaciones intracadenas. Cuando el sistema investigado consistió en fases bien definidas de grupos hidrófilos e hidrófobos (tales como un copolímero de bloque o una red polimérica interpenetrada de fases separadas) [74], cada una de las fases se comportó en presencia del agua como si el otro componente no estuviera presente, es decir, las moléculas de agua se asocian alrededor de la fase hidrófila ignorando la presencia de la segunda fase. No existe mucha información acerca de los cambios conformacionales inducidos por el solvente en este tipo de sistemas topológicamente restringidos. 43 Se ha investigado redes de poli(acrilato de etilo-co-acrilato de hidroxietilo), P(EA-co-HEA), hinchadas en benceno mediante DSC; en ese trabajo se mostró la existencia de los cambios conformacionales inducidos por la presencia del disolvente [74]. En el presente trabajo, los cambios conformacionales en una red P(EA-coHEA) inducida por la presencia de agua se han analizado mediante diferentes técnicas experimentales, y se han comparado con los xerogeles correspondientes. La interacción entre los grupos hidrófobos y un disolvente polar es capaz de modificar la Tg con respecto a la esperada para una mezcla homogénea según la teoría de las mezclas. Partiendo de la ecuación de Couchman-Karasz se ha añadido términos de carácter empírico para relacionar la fuerza de estas interacciones. Una de ellas es la ecuación de Kwei: Tg = w solvente Tg,solvente + w xerogel kTg,xerogel w solvente + kw xerogel + qw solvente w xerogel Ecuación 1.8.- Ecuación de Kwei En esta ecuación el término de la izquierda de la segunda parte de la igualdad representa la ecuación de Couchman-Karasz cuando k= ΔC p,xerogel ΔC p,solvente El término de la derecha de la segunda parte de la igualdad representa el efecto de las interacciones específicas. 44 1.3.- Interacción material-célula La interacción inicial célula-material es un proceso multietapa que comienza con la adsorción y conformación de proteínas sobre el substrato. Si la célula es capaz de comunicarse correctamente con las proteínas adsorbidas sobre el substrato, tiene lugar la adhesión celular y expansión de las mismas. Posteriormente tienen lugar los procesos relacionados con el crecimiento, la diferenciación, la deposición de matriz y el funcionamiento celular. Así, en un primer momento, la respuesta celular ante un material de origen externo viene determinada por la adsorción y conformación de las proteínas sobre el substrato, actuando éstas como una interfaz entre las propias células y el material. Por tanto, las células no “ven” al substrato directamente sino a las proteínas adsorbidas sobre la superficie del mismo [75]-[92]. En este trabajo, hemos estudiado una de las proteínas de la matriz extracelular, ECM, la fibronectina, FN, interactuando con diferentes substratos en los que se ha modificado monótonamente el grado de hidrofilia. Para ello, se han elegido copolímeros de la familia de los acrilatos -acrilato de etilo/acrilato de hidroxietilo-, ya que estos sistemas poseen una estructura química muy similar y se puede variar el grado de hidrofilia a voluntad. El proceso de anclaje celular al substrato comienza con el reconocimiento por los receptores de las integrinas (proteínas transmembrana celular) de secuencias específicas de la FN, como la RGD, por ejemplo. Las integrinas proporcionan los vínculos entre la matriz extracelular y el citoesqueleto de actina [93]. Una vez que estos receptores se unen a la fibronectina, las integrinas activan una cascada de rutas de señalización intercelular y se desarrollan las adhesiones focales, FAK, un agregado de diferentes proteínas, que anclan efectivamente la célula al substrato, desencadenando la subsiguiente respuesta celular [94]-[95]. 45 Las características físico-químicas del substrato tienen una influencia directa sobre la adsorción y conformación de las proteínas y éstas responden de modo general a tres tipos de propiedades superficiales: químicas, topográficas y mecánicas [96]. Un parámetro clásico que afecta a la adsorción de proteínas y la adhesión celular es el balance hidrófilo/hidrófobo y hay numerosos trabajos que estudian la adsorción y conformación de diferentes proteínas sobre soportes de la familia de los acrilatos y metacrilatos con variaciones en el grado de hidrofilia [97]-[102]. 1.4.- La vascularización en los scaffolds Cuando un scaffold con células en su seno se introduce en el tejido biológico se origina una respuesta inflamatoria inducida en parte por el procedimiento quirúrgico. Además, las células crean un estado de hipoxia en el implante que provoca la liberación de factores de crecimiento angiogénicos [103]. Este crecimiento de vasos sanguíneos es, en general, lento e insuficiente para proporcionar los nutrientes a las células del constructo trasplantado, y es uno de los problemas centrales de la ingeniería tisular cuando un implante poroso que alberga en su seno células entra en contacto con el tejido receptor. En las condiciones in vivo las células de los tejidos reciben oxígeno y nutrientes a través de los vasos sanguíneos y, por tanto, la formación de nuevos vasos sanguíneos para alimentar a las células en el seno del scaffold, tiene que producirse de manera rápida. Existen diferentes estrategias que persiguen mejorar la vascularización de constructos para ingeniería tisular como: (i) el diseño del scaffold, (ii) la inclusión de factores angiogénicos y (iii) la prevascularización in vivo e in vivo [104]. 46 Las dos primeras estrategias, el diseño del scaffold y la liberación de factores angiogénicos se basan en el crecimiento de nuevos vasos sanguíneos en el implante en su conjunto; tiene como desventaja que el tiempo de crecimiento de los nuevos vasos es demasiado largo hasta alcanzar el centro del constructo. La prevascularización in vivo implica preimplantar el constructo en un lugar del organismo que no es definitivo y, en principio, podría servir para obtener un constructo totalmente vascularizado al implantarlo en el lugar definitivo; sin embargo, durante el periodo de preimplantación deben formarse nuevos vasos sanguíneos espontáneamente, por lo que la falta de nutrientes podría ocurrir de igual manera. La prevascularización in vitro no da lugar a una perfusión instantánea del constructo ya que los vasos del huésped deben conectar con los vasos preformados in vitro. Este último método disminuye notablemente el tiempo necesario para vascularizar el implante comparado con la liberación de factores y al diseño del scaffold [104]. En el diseño del scaffold, los factores que más influyen en la vascularización son el tamaño de poro y la interconectividad de los mismos [105]-[107]. Dado que las técnicas de prototipado rápido (SFF) pueden ofrecer un mayor control de la arquitectura del scaffold éstas están recibiendo una atención creciente [108],[109]. La adición de factores angiogénicos en los constructos puede mejorar la vascularización tras el implante [110], mejorando la vascularización en sus diferentes etapas. Los factores vascular endotelial growth factor, VEGF, y basic fibroblast growth factor, bFGF, se han utilizado para la formación de nuevos vasos sanguíneos [111]; es importante que los nuevos vasos sean estables (no desorganizados, con fugas o hemorrágicos) y factores como el platelet-derived growth factor, PDGF, el transforming growth factor , TGF- , y la angiopoietin-1, Ang1, se han utilizado en este sentido. La 47 liberación simultánea de VEGF y PDGF ha dado lugar a la formación de vasos sanguíneos maduros en los scaffolds implantados [110]-[114]. Otro tipo de estrategia consiste en la adición de factores como el sonic hedgehog homolog, SHH, y el hypoxia-inducible factor 1, HIF-1 o el bone morphogenetic protein, BMP-2, -4 o -6, que estimulan a las células cercanas al implante a la producción de factores angiogénicos [115]-[117]. La prevascularización in vivo se realiza en dos epatas. En la primera el constructo es implantado en una región con una irrigación sanguínea adecuada como, por ejemplo, un músculo; o bien, una arteria es implantada en el injerto [118]. Con el paso del tiempo se crea una red microvascular en el seno del constructo, el cual se extrae junto con la arteria y se reimplanta en el lugar definitivo. La prevascularización in vitro se basa en el hecho de que las células endoteliales son capaces de formar un red microvascular in vitro cuando son cultivadas en condiciones adecuadas. Los estudios de prevascularización in vitro incluyen piel [119]-[121], músculo esquelético [122], hueso [123]-[126], y músculo cardiaco [127],[128]. Con esta aproximación los vasos sanguíneos del huésped no necesitan crecer en todo el constructo sino sólo en la parte exterior hasta conectar con el sistema prevascular creado in vitro. Las células endoteliales juegan un papel esencial en el éxito de un implante y cómo estas células reaccionan ante biomateriales es de suma importancia para la supervivencia de las células que crecen en el material. Por ejemplo, en [6] el homopolímero, PEA, y sus copolímeros p(EA-co-HEA) y p(EA-co-MAAc) en forma de scaffold se utilizaron como soportes de cultivo in vitro con neural stem cells (NSC). En el caso del reemplazo de córnea, que es un tejido no vascularizado, mediante una queratoprótesis del tipo “core and skirt”, el oxígeno y otros nutrientes deben llegar a las células epiteliales que recubren la lente por 48 difusión. El proceso de difusión está obstaculizado por la presencia del scaffold, y esto puede comprometer la viabilidad a largo plazo del constructo. Una posible estrategia en el diseño e implantación de una prótesis de córnea es inducir la angiogénesis en el anillo de anclaje ( scaffold) y, por tanto, proporcionar a las células los nutrientes necesarios para crecer y anclar efectivamente el constructo. Al mismo tiempo, el anillo de anclaje debe preservar el núcleo de la prótesis (la lente) de la vascularización. Una vez que el scaffold sembrado con las células del estroma corneal del paciente se implanta, los nuevos vasos sanguíneos deben invadir sólo el anillo de anclaje del scaffold. Esto implica que el sistema vascular de la conjuntiva debe generar nuevos capilares sanguíneos, que a continuación deben crecer en el material poroso y proporcionar un suministro de sangre a las células en el implante. La vascularización del scaffold puede beneficiarse de la prevascularización in vitro de células endoteliales autólogas que se unen al sistema vascular del huésped tras el implante. En este sentido, en este trabajo se han utilizado células endoteliales humanas primarias como un tipo de célula modelo para probar la compatibilidad de las células endoteliales con diferentes scaffolds poliméricos con diferentes características de hidrofilia. La compatibilidad de las células endoteliales se midió en términos de viabilidad celular y de la adecuada diferenciación endotelial/funcionamiento. En concreto, se han realizado cultivos de células endoteliales humanas primarias, HUVEC, y se ha determinado si su morfología y función se vio afectada por el material. Se examinó la adhesión y proliferación de las mismas, así como un marcador importante de activación endotelial, la E-selectina. Se evaluó si se mantuvieron los fenotipos endoteliales normales y sus funciones observadas in vivo mediante análisis de los contactos célula-célula y la regulación de la expresión génica del marcador de activación E-selectina cuando se añadió un estímulo (LPS). 49 Además, como posible aplicación de estos materiales en una prótesis de córnea artificial, y dado que los fibroblastos del estroma de la córnea (es decir, los queratocitos) son de relevancia en la cicatrización de la córnea [129] se determinó cómo afectaba la hidrofilicidad a la adhesión celular de la línea de fibroblastos humanos MRC-5, que sirven como modelo celular [130],[131] para estudiar la disposición del citoesqueleto tras la adhesión a los diferentes soportes mediante la detección de F-actina. 1.5.- La epitelización en implantes protésicos de córnea El epitelio es el tejido formado por una o varias capas de células unidas entre sí, distinguiéndose en el cuerpo humano el epitelio de revestimiento (el tejido que tapiza todas las superficies del organismo) y el epitelio glandular, cuya función es la secreción de moléculas al exterior de la célula. Existen diferentes tipos de epitelio, a saber, plano simple, cúbico simple, cilíndrico simple, cilíndrico pseudoestratificado, de transición, plano estratificado y cilíndrico estratificado [132],[133]. La parte anterior del ojo humano, la córnea, es una estructura transparente que permite el paso de los rayos de luz. La córnea tiene en torno a 0.8 mm de espesor en su centro y 1,1 mm en la región periférica. Es avascular y consiste en un grupo altamente organizado de células y matriz extracelular. La córnea está compuesta de cinco capas: el epitelio, la membrana de Bowman, el estroma, la membrana de Descemet y el endotelio. El epitelio de la córnea tiene en torno a 50 micras de espesor y está formado por 5-7 capas de células epiteliales con alta capacidad de regeneración. El epitelio corneal descansa sobre la membrana de Bowman, que tiene de 6 a 9 micras de espesor y separa el epitelio del estroma. Este último representa aproximadamente el 90% del espesor de la córnea y consta 50 de un tejido conjuntivo transparente formado por fibras de colágeno que tienen una disposición muy regular. Las células del estroma de la córnea son fibroblastos largos y estrechos, llamados queratocitos, que permanecen entre las fibras de colágeno. La membrana de Descemet tiene un espesor de 5 a 10 micras y separa el estroma del endotelio. Se trata de una lámina basal producida por el endotelio corneal. En la actualidad, el tratamiento más extendido y aceptado para la ceguera corneal es el trasplante de córnea procedente de un donante (queratoplastia penetrante). La escasez de donantes ha motivado que diferentes grupos de investigación estén trabajado en el reemplazo de córnea utilizando diferentes estrategias: la construcción de prótesis artificiales (KPros) y el uso de terapias regenerativas basadas en células (construcciones celulares auto-ensambladas). En el caso de las prótesis artificiales el modelo “core and skirt” ha sido el más empleado. Estos dispositivos –algunos de ellos aprobados para uso médico- poseen una parte central transparente que permite el paso de la luz y un anillo soporte poroso que proporciona el anclaje al tejido del huésped [134]. Uno de los requisitos esenciales para que un implante de queratoprótesis tenga éxito es que se cree y mantenga una capa de células epiteliales que impidan entrar a las bacterias al interior del ojo y permita la difusión la capa lagrimal de manera estable en el tiempo [135]. Por ello, diferentes trabajos han perseguido ajustar las propiedades del material para conseguir que las células epiteliales se adhirieran al substrato. Una de las estrategias ha sido anclar a los substratos péptidos como el IKVAV, YIGRS y RDG [136]-[140]. En el presente trabajo, se han analizado parámetros celulares como adhesión, proliferación y viabilidad de una línea de células epiteliales de 51 conjuntiva humana, NHC [141], cultivada sobre substratos poliméricos con diferentes grados de hidrofilia y cargas eléctricas superficiales buscando qué grado de hidrofilicidad permite la epitelización del substrato y podría darle al material flexibilidad y la hidrofilicidad necesaria para un mejor contacto con los párpados y lágrima. 52 Hipótesis y objetivos Se plantea la hipótesis de que “en los copolímeros producidos con monómeros acrílicos por vía radical con un componente hidrófilo y otro hidrófobo, la composición del copolímero controla la conformación de las proteínas adsorbidas sobre ellas y, a través de ello, la adhesión y proliferación celular”. Objetivo General: Desarrollar substratos planos y tridimensionales con copolímeros de acrilato de etilo, acrilato de hidroxietilo y ácido metacrílico. Evaluar la correlación existente entre sus propiedades morfológicas y físicas por un lado, y la respuesta biológica en cultivos in vitro por otro. Objetivos concretos: 1. Investigar los reordenamientos conformacionales inducidos por el agua en estos materiales. 2. Estudiar el efecto del balance hidrófobo/hidrófilo en la superficie sobre la adhesión de células epiteliales, fibroblastos y células endoteliales. 3. Estudiar el efecto del balance hidrófobo/hidrófilo en la superficie sobre la adsorción de fibronectina y correlacionar los resultados con el cultivo celular. 53 Capítulo 2. Materiales y métodos 54 2.1.- Materiales 2.1.1.- Polímeros en bloque. Sistemas P(EA-co-HEA) y P(EA-coMAAc) Los monómeros de acrilato de etilo, EA (Aldrich, 96%), acrilato de hidroxietilo, HEA (Aldrich, 99%) y MAAc (Aldrich, 99%) se usaron sin purificación adicional. La redes de copolímeros se polimerizaron en luz ultravioleta, a temperatura ambiente, usando 2% en peso de dimetacrilato de etilenglicol, EGDMA (Aldrich, 98% puro), como agente reticulante y 1% en peso de benzoína (Scharlau 98% puro) como fotoiniciador. La polimerización se llevó a cabo en un molde que consistía en dos placas de vidrio con un espaciador de goma lo que permitió preparar una lámina de polímero de aproximadamente 1 mm de espesor. Las sustancias residuales de bajo peso molecular que permanecían tras la polimerización se extrajeron con etanol hirviendo durante 24 h; a continuación las muestras se secaron a 60ºC a vacío hasta peso constante. Las composiciones de las diferentes redes de homopolímero y copolímeros en bloque así como la nomenclatura utilizada en este trabajo se muestran en la Tabla 2.1. 55 Fracción en peso de EA en Nomenclatura Composición BH000 PEA 100 BH010 P(EA-co-HEA) 90/10 90 BH020 P(EA-co-HEA) 80/20 80 BH030 P(EA-co-HEA) 70/30 70 BH050 P(EA-co-HEA) 50/50 50 BH070 P(EA-co-HEA) 30/70 30 BH100 PHEA 0 BM010 P(EA-co-MAAc) 90/10 90 BM020 P(EA-co-MAAc) 80/20 80 % Tabla 2.1.- Composición de las redes de homopolímero y copolímeros empleadas 2.1.2.- Polímeros porosos. Scaffolds de P(EA-co-HEA) Se utilizaron microesferas de poli(metacrilato de metilo), de 80-110 micras de diámetro (PMMA Colacryl DP300, Lucite International) como porógeno. La sinterización de las partículas para preparar una plantilla se realizó como sigue: las microesferas se introdujeron en un molde que consistía en dos placas de vidrio y un anillo de goma. El molde se introdujo en un horno a 160ºC para superar la Tg de las microesferas de PMMA y llevarlas al estado de goma; se aplicó una fuerza de compresión sobre la placa de porógeno y se disminuyó el espesor en la relación deseada. Por otro lado, las soluciones de monómeros de acrilato de etilo, EA (Aldrich, 96%), y acrilato de hidroxietilo, HEA (Aldrich, 99%) se prepararon usando un 2% de etilenglicol dimetacrilato, EGDMA (Aldrich) como agente de reticulación y un 1 % de benzoína (Scharlau) como fotoiniciador. Se obtuvieron cuatro composiciones diferentes, siendo la fracción másica de EA: 1, 0.9, 0.8 y 0.5, 56 respectivamente. Las plantillas de las microesferas sinterizadas se sumergieron en las soluciones de monómeros y se polimerizaron con luz ultravioleta a temperatura ambiente. Después de la polimerización se llevó a cabo la disolución de la plantilla de PMMA con acetato de etilo durante 48 horas en extractor Soxhlet. Entonces, el acetato de etilo se intercambió progresivamente con etanol, etanol/agua y agua para permitir que los scaffolds se contrajeran lentamente y evitar su colapso. Finalmente, los scaffolds se secaron a 60ºC en vacío hasta peso constante. Se cortaron varias réplicas de cada scaffold para los diferentes ensayos y se obtuvieron micrografías electrónicas de barrido en un microscopio ISIDS-130 a un voltaje de aceleración de 10 kV para mantener la integridad de las muestras. La figura 2.1 muestra la estructura de los diferentes scaffolds mientras que su composición se muestra en la Tabla 2.2. Muestra Composición(% en peso) SH000 PEA SH010 P(EA-co-HEA) 90/10 SH020 PEA-co-PHEA 80/20 SH050 PEA-co-PHEA 50/50 Tabla 2.2.- Composición de los diferentes scaffolds de P(EA-co-HEA) 57 P(EA-co-HEA) 100/0 P(EA-co-HEA) 90/10 P(EA-co-HEA) 80/20 P(EA-co-HEA) 50/50 Figura 2.1.- Microscopía electrónica de barrido de los scaffolds de la serie poli(acrilato de etilo-co-acrilato de hidroxietilo), P(EA-co-HEA). Los números indican la cantidad en porcentaje de cada componente respecto al peso total de la mezcla monomérica. Barrita: 100 micras. 58 2.2.- Métodos 2.2.1.- Ensayos de los materiales empleados en estado seco 2.2.1.1.- Espectroscopía dinámico-mecánica (DMS) Se prepararon muestras secas (xerogeles) de los copolímeros de la serie P(EA-co-HEA) con dimensiones aproximadas de 10 x 4,5 x 1 mm3. Se utilizó el analizador Seiko DMS 210 para llevar a cabo la espectroscopia dinámicomecánica (DMS) a la frecuencia de 1 Hz. La dependencia con la temperatura del módulo de almacenamiento (E'), del módulo de pérdidas (E'') y de la tangente de pérdidas (tan δ) se midieron desde -50ºC hasta 25ºC a una velocidad de calentamiento de 1 K/min en el caso de BH050, y de -60ºC a 140ºC en los homopolímeros (BH000 y BH100). 2.2.1.2. Calorimetría diferencial de barrido (DSC) Las muestras secas de la serie P(EA-co-HEA) se analizaron en un calorímetro diferencial de barrido Pyris 1 (PerkinElmer). Se hizo fluir nitrógeno gas través del horno de medida con un caudal másico de 20 ml/minuto. La temperatura del equipo se calibró con agua e indio en el primer caso y con zinc e indio en el segundo. El calor de fusión del indio se usó para calibrar el flujo de calor. Las muestras secas se sometieron a un barrido de enfriamiento desde la temperatura ambiente hasta -30ºC, seguido de un barrido de calentamiento desde -30ºC hasta la temperatura de 60°C, ambos barridos a una velocidad de 10ºC/minuto. Las temperaturas de transición características se calcularon a partir de los termogramas de DSC. Como temperatura de transición vítrea, 59 Tg, se tomó la temperatura a la que el incremento de Cp en la transición es la mitad del incremento total en el termograma. 2.2.1.3.- Microscopía de fuerza atómica (AFM) Una muestra en forma de disco, seca a vacío, del polímero BH5050 se utilizó para un ensayo de microscopía de fuerza atómica, AFM, en un NanoScope III de Digital Instruments que operó en modo tapping al aire; se utilizó un soporte voladizo de Si con una constante de fuerza de 42 N/m y una frecuencia de resonancia de 290 kHz. La señal de fase se ajustó a cero en la frecuencia de resonancia de la punta. La frecuencia de tapping fue ligeramente inferior a la de resonancia. 2.2.2.- Ensayos de los materiales empleados hidratados 2.2.2.1.- Energía superficial (SE) Las mediciones del ángulo de contacto se realizaron usando un Dataphysics OCA 10. Se utilizó el método sessile-drop para ángulo de contacto estático. Las muestras de los polímeros en bloque se hidrataron previamente en agua a temperatura ambiente. Para los cálculos de tensión superficial γs de las muestras hidratadas se utilizaron glicerol, diyodometano y formamida. Las componentes polar y dispersiva (γp y γd) se calcularon siguiendo el método de Owens y Wendt [142]. 2.2.2.2.- Espectroscopía dinámico-mecánica (DMS) Para espectroscopía dinámico-mecánica, DMS, se cortaron tres réplicas de BH050de la placa de polímero en bloque, con dimensiones aproximadas de 10 x 4,5 x 1 mm3. La primera de ellas se utilizó en estado seco. La segunda 60 se hinchó en un desecador cerrado que contenía un vaso con una solución de cloruro de litio saturado a 20ºC, de tal manera que la atmósfera de aire húmedo que rodeó a la muestra tenía una humedad una humedad relativa del 11,30% [143], lo que condujo a obtener un contenido de agua en la muestra muy bajo. La tercera réplica se hinchó hasta equilibrio por inmersión en agua líquida. Se utilizó el analizador Seiko DMS 210 para llevar a cabo la espectroscopia dinámico-mecánica (DMS) a 1 Hz. Se midió la dependencia con la temperatura del módulo de almacenamiento (E'), del módulo de pérdidas (E'') y de la tangente de pérdidas (tan ), desde -50ºC hasta 25ºC a una velocidad de calentamiento de 1 K/minuto. 2.2.2.3.- Calorimetría diferencial de barrido (DSC) Diferentes réplicas de BH050 se sumergieron en agua durante periodos tiempos crecientes para llegar a diferentes contenidos de agua, desde muy bajos hasta los valores de contenido en equilibrio. El barrido se realizó en un aparato Pyris 1 (PerkinElmer). Se hizo fluir nitrógeno gas través del horno de DSC con un caudal másico de 20 ml/minuto. La temperatura del equipo se calibró igual que se describe en el apartado 2.2.1.2. Tres réplicas de P(EA-co-HEA) 50/50 (BH050) se sometieron a un barrido de enfriamiento desde la temperatura ambiente hasta -100ºC, seguido de un barrido de calentamiento desde esa temperatura hasta los 20°C, ambos barridos a una velocidad de 10ºC/minuto. Las temperaturas de transición características se calcularon a partir de los termogramas de DSC. Como temperatura de transición vítrea, Tg, se tomó la temperatura a la que el incremento de Cp en la transición es la mitad del incremento total en el termograma, mientras que para la transición de fase del agua se tomó la temperatura máxima del pico exotérmico o endotérmico mayor. La masa de agua en el gel hinchado se determinó gravimétricamente. El contenido de 61 disolvente (agua) en el gel hinchado se determinó cuantitativamente mediante la fracción en peso de agua respecto del peso total del gel, w. w= m agua m agua + m xerogel 2.2.2.4.- Espectroscopía de relajación dieléctrica (DRS) Se cortaron cuatro muestras de la placa de polímero BH050 en forma de discos para realizar la espectroscopía de relajación dieléctrica, DRS. Tres de ellas se hincharon en atmósferas de aire húmedo, cuya humedad relativa se controló con diferentes soluciones salinas saturadas, para obtener diferentes contenidos de agua, a saber, cloruro de litio (11,30% de humedad relativa), nitrato de magnesio (54,38% de humedad relativa), nitrato de magnesio (54,38% de humedad relativa) y sulfato de potasio (97,59% de humedad relativa) [143]. La cuarta réplica se utilizó en estado seco. Los experimentos de espectroscopía de relajación dieléctrica (DRS) se realizaron con un analizador de impedancia Alfa-S. El sistema de control de temperatura fue el Quatro Cryosystem de Novocontrol GmbH. Las muestras se colocaron entre dos electrodos de oro chapado (diámetro 20 mm) de un condensador de placas paralelas. La celda, con convertidor dieléctrico de cabeza activa, se montó en un criostato (BDS 1100) y se expuso a una corriente de gas calentado que se evaporó desde un Dewar de nitrógeno líquido. En estos experimentos, la permitividad dieléctrica compleja se determinó en función de la frecuencia (10-1-107 Hz) en un intervalo de temperatura entre -50ºC y 50ºC. 2.2.2.5.- Microscopía de fuerza atómica (AFM) Se cortó una muestra de la placa de polímero BH050 en forma de disco para microscopía de fuerza atómica, AFM, y se hinchó hasta equilibrio. La 62 muestra se valoró en un NanoScope III de Digital Instruments en modo tapping cubierta por una gota de agua líquida. Un soporte voladizo de Si con constante de fuerza de 0.58 N/m fue utilizado para el AFM en modo líquido. 2.2.2.6.- Contenido de agua en equilibrio (EWC) El contenido de agua de equilibrio de toda la serie de muestras de los polímeros en bloque en inmersión se determinó dejando que las muestras secas se equilibraran en agua destilada a 25ºC hasta que el cambio de peso de las muestras fue de menor de 10-4 g. 2.2.3.- Adsorción de fibronectina (FN) sobre los substratos 2D 2.2.3.1.- Preparación de polímeros en bloque para adsorción de fibronectina Los polímeros en bloque de la serie P(EA-co-HEA) se pegaron a un portaobjetos de AFM y se recubrieron con fibronectina (1 mg/ml, Roche, Mannheim, Alemania) en suero oftalmológico (1:200) durante 15 minutos. Las muestras se lavaron suavemente con suero para eliminar las proteínas que no se adsorbieron. Por último, los polímeros en bloque se secaron con nitrógeno gas y se colocaron en el AFM. El estudio de la adsorción de fibronectina se realizó en los polímeros en bloque BH000, BH010, BH020 y BH050 del mismo modo que el descrito anteriormente para las muestras en estado seco operando en modo tapping al aire. Se utilizó un soporte voladizo de Si con constante de fuerza de 2,8 N/m y frecuencia de resonancia de 75 kHz. La señal de fase se ajustó a cero en la frecuencia de resonancia de la punta. La frecuencia de tapping fue ligeramente inferior a la de resonancia. 63 2.2.4.- Cultivos celulares sobre los substratos 2D y 3D 2.2.4.1.- Tipos celulares y condiciones de crecimiento Células epiteliales Se cultivaron células conjuntivales humanas normales (IOBA-NHC) [141], inmortalizadas espontáneamente en medio DMEM/F12 + 10% de suero bovino fetal inactivado térmicamente + 2,5μg/ml Fungizone + 5.000 U/ml/5.000μg/ml Penicilina/Estreptomicina + 2ng/ml de factor de crecimiento epitelial (EGF) de ratón + 1μg/ml de insulina de páncreas bobino + 0.1 μg/ml de toxina del cólera + 5μg/ml de hidrocortisona. Células endoteliales Células endoteliales humanas fueron aisladas de cordón umbilical (células endoteliales de cordón umbilical humano, HUVEC) de acuerdo con un método publicado previamente [144]. Las células se cultivaron en medio M199 (Sigma-Aldrich, Steinbach, Alemania) + 20% de suero bobino fetal (Life Technologies, Karlsruhe, Alemania) + 2 mM Glutamax I (Life Technologies) + 100 U/100 mg/ml de penicilina/estreptomicina + 25 mg/ml de heparina sódica (Sigma-Aldrich) + 25 mg/ml de suplemento de factor de crecimiento endotelial (ECG, Becton Dickinson, Bedford, MA). Fibroblastos La línea celular normal de fibroblastos de pulmón humano, MRC- 5, se cultivó en medio EMEM (Sigma-Aldrich, Steinbach, Alemania ) + 10% de suero bovino fetal (Life Technologies, Karlsruhe, Alemania) + 2 mM Glutamax I (Life Technologies) + 100 U/100 mg/ml Pen/Strep. 64 2.2.4.2.- Preparación de los polímeros en bloque y scaffolds para cultivo celular Los polímeros en bloque y los scaffolds se cortaron con forma de discos de diámetro aproximado de 3-4 mm y 1 mm de espesor, se lavaron con etanol en un baño de ultrasonidos durante 3 minutos y se envasaron para su esterilización con radiación gamma, 25 kGy (Aragogamma, SA). La Dra. Marta Abad-Collado (en Vissum, Alicante) estudió la adhesión, viabilidad, morfologías y proliferación de las células normales de la conjuntiva humana sobre sustratos en bloque. Los polímeros en bloque se sembraron en placas de 96 pocillos y se lavaron durante la noche con 1xDPBS (Sigma) estéril. Al día siguiente, el PBS se eliminó de los pocillos y los polímeros se equilibraron con el medio de cultivo completo. Después de este tiempo se retiró el medio y las células normales de conjuntiva humana, NHC, se sembraron con una densidad de 5x103 células/réplica en 10 l de DMEM/F12 completo que contenía lo descrito anteriormente. Las células se decantaron durante al menos 2 horas, tras lo cual se añadió suavemente 200 l/pocillo de medio de cultivo. La incubación se realizó a 37ºC en un incubador de CO2 al 5%. La siembra de las células de cordón umbilical, HUVEC y los fibroblastos, MRC-5, sobre los scaffolds se realizó por el doctorando en el REPAIR Lab, de la Universidad de Mainz, en una estancia predoctoral bajo el programa Marie-Curie. En una placa de 96 pocillos diferentes réplicas de scaffolds se pegaron con silicona médica (Sinelastic ®) a cada pocillo. Después de 15 minutos a 37ºC se añadieron células endoteliales humanas de cordón umbilical, HUVEC, o una línea celular normal de fibroblastos de pulmón humano, MRC- 5, a cada pocillo (3x104 células por pocillo) y se incubaron a 37ºC, 5% de CO 2. Se reemplazó el medio cada 2 ó 3 días. Para la siembra de HUVEC, los scaffolds 65 se recubrieron previamente con fibronectina (1 mg/ml, Roche, Mannheim, Alemania) en PBS (1:200) durante 1 hora a 37ºC previamente a la siembra de células para permitir la adhesión celular. 2.2.4.3.- Adhesión y viabilidad celular de epiteliales NHC sobre los substratos poliméricos en bloque Para el análisis de la adhesión celular de las NHC a determinados tiempos, 3, 5, y 7 días, las células fueron lavadas dos veces suavemente con 200 l de 1xDPBS para eliminar medio de cultivo. Los substratos poliméricos en bloque se trasladaron a una nueva placa de 96 pocillos, se tripsinizaron usando 150 l de tripsina al 0.25% con EDTA 1 mM por pocillo y se incubaron durante 90 segundos a 37ºC en un incubador de CO2 al 5%. La tripsina se bloqueó mediante la adición de 150 l de medio de bloqueo en frío que contenía medio DMEM/F12 enriquecido con 10% de FBS y 5000 U/ml de los antibióticos penicilina/estreptomicina. Las NHC se resuspendieron y combinaron en tubos eppendorf estériles que contenían 450 l más de medio de bloqueo. Los tubos se centrifugaron a 800 r.p.m. durante 5 minutos, se lavaron se lavaron una vez con 1xDPBS y finalmente se resuspendieron con 500 l de DPBS. Se llevó a cabo una doble dilución con azul tripán y se cuantificaron las células. Para la viabilidad celular se realizaron los mismos ensayos de cultivo, pero, a determinados tiempos, las células NHC se lavaron dos veces con DPBS y 150 l de 0.2% de azul tripán diluido en PBS se añadió a los pocillos; se incubaron a temperatura ambiente durante 10 minutos. Después de este período, el colorante de viabilidad se retiró y las células se lavaron dos veces con DPBS. Las células se observaron bajo un microscopio óptico invertido y se obtuvieron fotografías. 66 2.2.4.4.- Tinción con hematoxilina-eosina de epiteliales NHC sobre los substratos poliméricos en bloque Para la evaluación de la morfología de las células NHC, se llevó a cabo una tinción hematoxilina-eosina sobre las muestras fijadas de polímero en bloque y cultivadas como antes. Los biomateriales colonizados fueron fijados con 4% de paraformaldehido (PFA) durante 20 minutos a temperatura ambiente, se lavaron dos veces con DPBS y dos veces con agua destilada (dH2O), 5 minutos por lavado. Después de eso, los materiales se sumergieron en hematoxilina de Mayer (Merk) durante 3 minutos y el lavaron con dH2O durante 5 minutos. Las muestras se lavaron luego 2 minutos en etanol al 80% y 5 minutos con eosina Y (Electron Microscopy Sciences). Por último, las muestras se deshidrataron en 4 pasos: 5 minutos en 96% de etanol, 5 minutos en 100% de etanol y 2 últimos pasos en Xilol (Panreac), 5 minutos cada uno. Los materiales fueron montados en un portaobjetos de microscopio con un medio de montaje especial (Electron Microscopy Sciences) y se observaron a través de un microscopio de luz. 2.2.4.5.- Análisis de la proliferación celular de epiteliales NHC sobre los substratos en bloque Para la proliferación celular, las células NHC fueron cultivadas sobre los biomateriales a diferentes tiempos: a 3, 5 y 7 días como se explicó anteriormente. Para cada tiempo las células se lavaron dos veces con 1xDPBS y se fijaron con 4% de formaldehído (Panreac) durante 15 minutos a temperatura ambiente. Las células se lavaron dos veces con DPBS ultra frío, permeabilizadas durante 10 minutos con 0.5% de saponina (Sigma), lavadas 3 veces con DPBS y bloqueadas con 1% de BSA (Sigma) en PBS durante 30 minutos para bloquear uniones inespecíficas. Las células fueron incubadas con un anticuerpo primario monoclonal de ratón anti-humano 67 contra el antígeno nuclear Ki67 (Dako) durante 1 hora a temperatura ambiente y se lavaron 3 veces con DPBS, 5 minutos por lavado. Los biomateriales colonizados fueron incubados con anticuerpo secundario de cabra anti-ratón marcado con Alexa 488 (Molecular Probes) durante 1 hora a temperatura ambiente en oscuridad. Después de esta incubación, las muestras se lavaron 3 veces con DPBS en la oscuridad y, finalmente, se montaron con Prolong (Molecular Probes) en un portaobjetos de microscopio, cubiertas con cubreobjetos, curadas durante al menos 24 horas y selladas con esmalte de uñas. Se observaron las muestras y se tomaron imágenes utilizando un microscopio de contraste de fases equipado con fluorescencia. 2.2.4.6.- Visualización y caracterización del crecimiento de las endoteliales HUVEC y los fibroblastos MRC-5 en los scaffolds Tras varios intervalos diferentes de tiempo después de la adición de las células, los scaffolds sembrados con HUVEC se retiraron del medio de cultivo y se colocaron en medio que contenía 0.1 mM de calceína-AM (Mobitec, Alemania) y se incubaron durante 10-20 min a 37ºC [145],[146]. La CalceínaAM se vuelve fluorescente cuando se metabolizada por las células viables y la fluorescencia se extiende por toda la célula. Los scaffolds teñidos se pusieron en un portaobjetos de microscopio y se examinaron por microscopía confocal de barrido láser (CLSM: Leica TCS NT). En el caso de MRC-5 la inmunofluorescencia se utilizó para evaluar el crecimiento de los fibroblastos en los scaffolds. Tras varios intervalos diferentes de tiempo los scaffolds sembrados con MRC-5 se retiraron del medio de cultivo y se enjuagaron brevemente con PBS; después, se fijaron con paraformaldehído al 3,7% en PBS durante 15 min a temperatura ambiente, se lavaron 4 veces con PBS y se permeabilizaron con 0.2% de Triton X-100 durante 10 minutos. Después de lavar 4 veces con PBS, se 68 añadió faloidina 1:80 y se incubó a temperatura ambiente durante 30 minutos. Los scaffolds se lavaron después 4 veces con PBS y se incubaron con Hoescht 1:1000 durante 3 minutos Las réplicas de los scaffolds se lavaron a continuación con PBS, se colocaron en Gel/Mount (Natutec, Alemania) y se examinaron por CLSM. 2.2.4.7.- Análisis estadístico Con el fin de evaluar las diferencias estadísticas en el crecimiento de las HUVEC en los cuatro scaffolds, SH000, SH010, SH020 y SH050, se calculó la superficie cubierta por las HUVEC en los mismos por medio de Adobe Photoshop. Se utilizó el ratio de píxeles verdes proporcionadas por calceínaAM vs píxeles totales en una imagen de CLSM para establecer la superficie cubierta. Los datos se expresaron como media +/- desviación estándar (SD) y las diferencias estadísticas se determinaron mediante el test de KruskalWallis para el material y los donantes. Cada experimento se realizó por duplicado y se repitieron al menos tres veces utilizando diferentes donantes de HUVEC. 2.2.4.8.- Análisis molecular de la expresión génica de las endoteliales HUVEC sobre los scaffolds Se aisló el ARN de las células endoteliales cultivadas en scaffolds recubiertos de fibronectina, con presencia o ausencia de 100 ng/ml de LPS (Sigma) durante 4 horas, utilizando el RNeasy MICROKIT (Qiagen, Hilden, Alemania). Se colocaron tres réplicas con células endoteliales directamente en tampón de lisis. Las células se retiraron de los flascones de cultivo con tripsina, se centrifugaron y los sedimentos celulares se resuspendieron en tampón de lisis. La concentración del ARN extraído se determinó utilizando el microespectrofotómetro NanoDrop (Nanodrop Technologies, Rockland, 69 Delaware) y 20 ng de ARN se transcribió en cDNA (kit Omniscript RT, Qiagen); después, se almacenó a 4ºC. La amplificación de genes endoteliales específicos se llevó a cabo mediante PCR en 200 pg de cDNA con los reactivos suministrados en el kit de polimerasa Taq de DNA (Qiagen) con los cebadores descritos en la referencia [147]. Resumidamente, se llevó a cabo una desnaturalización inicial a 94ºC durante 2 minutos, seguida de 35 ciclos a 94ºC durante 30 s, temperatura de apareamiento (variada para diferentes pares de cebadores, ver [147]) durante 30 s y 72ºC durante 30 s. La rtPCR se terminó con un paso final a 72ºC durante 10 minutos. Los productos de la PCR se resolvieron por electroforesis en gel de agarosa (0.8%), electroforesis en tampón TBE (Sigma) y se visualizaron por tinción con bromuro de etidio. Además, para garantizar que los productos amplificados resultaron de cDNA, en lugar de contaminación de DNA genómico, todas las muestras de ARN fueron tratadas con ADNasa I (DNasa libre de RNasa, Qiagen). 2.2.4.9.- HUVEC: análisis inmunofluorescente de Selectina E (CD62E) y PECAM-1 (CD31) Las HUVEC que crecieron en los scaffolds recubiertos de fibronectina se enjuagaron brevemente con PBS y después se fijaron con paraformaldehído al 3,7% en PBS durante 15 minutos a temperatura ambiente; se lavaron 4 veces con PBS y se permeabilizaron con 0.2% de Triton X-100 durante 10 minutos. Después de lavar 4 veces con PBS, se añadió el anticuerpo (1:100, E-selectina, Monosan, Países Bajos o 1:100, Santa Cruz Biotechnology, Inc., Alemania PECAM-1) y se incubó durante la noche a 4ºC. Los scaffolds se lavaron después cuatro veces con PBS y se añadió el anticuerpo secundario (1:1000, anti-ratón de Alexa Fluor 546 para E-selectina y 1:1000, anti-ratón de Alexa Fluor 488 para PECAM-1, Molecular Probes); finalmente, se incubaron durante 1 hora a temperatura ambiente. Se prosiguió con un breve enjuague en PBS y después se incubaron con 1:1000 de Hoescht y yoduro de 70 propidio durante 3 minutos respectivamente. Las réplicas de los scaffolds se lavaron a continuación con PBS, se colocaron en Gel/Mount (Natutec, Alemania) y se examinaron por CLSM. 71 Capítulo 3. Resultados y discusión 72 Uno de los factores que gobiernan la interacción de la célula con el material soporte sobre el que se cultiva, es la tensión superficial del material que determina su interacción con el medio acuoso en el que se realiza el cultivo. Pero como se ha descrito en la introducción y veremos más adelante en nuestros resultados, la interacción entre la célula y el soporte mediada por proteínas se ve afectada simultáneamente por un gran número de factores, lo que hace difícil desentrañar el papel de cada uno de ellos. Por ejemplo, no es sólo importante el valor medio de la energía de superficie, sino que la distribución de grupos hidrófilos e hidrófobos a escala nanométrica será importante en la conformación que las proteínas de adhesión adopten al ser adsorbidas sobre la superficie; esa conformación será a su vez determinante en la adhesión y morfología celular. De cara a poder extraer conclusiones sobre la forma en que la estructura química y las propiedades físicas del soporte influyen en la respuesta biológica, hemos sintetizado una serie de copolímeros que combinan unidades monoméricas que contienen grupos hidroxilo en la superficie con otras hidrófobas. En la sección 3.2 veremos la respuesta celular sobre estos substratos. Para analizar esos resultados, se requiere una información detallada sobre la distribución sobre la superficie del material de los grupos químicos presentes, lo que viene condicionado por la homogeneidad o separación de fases de las unidades monoméricas que componen las cadenas del copolímero. Esta información se va a extraer tanto de análisis teórico de la reacción de copolimerización como de medidas experimentales de propiedades físicas. 73 3.1.- Propiedades físicas de los polímeros soporte Esta sección está basada en los artículos publicados “Water-induced (nano)organization in poly(ethyl acrylate-co-hydroxyethyl acrylate) networks. Campillo-Fernández, A. J. et al. Eur Polym J 44; 1996-2004 (2008)” y “Future design of a new keratoprosthesis: Physical and biological analysis of polymeric substrates for epithelial cell growth. CampilloFernandez, A. J. et al. Biomacromolecules 8, 2429–36 (2007)”. 3.1.1.- Caracterización de los materiales en estado seco Los materiales empleados en este trabajo son redes poliméricas en las que el número de puntos de entrecruzamiento resulta el doble que el número de cadenas debido a que la funcionalidad del entrecruzador utilizado (EGDMA) es cuatro mientras que la de los monómeros es dos. En estas condiciones y utilizando la teoría de la elasticidad de gomas podemos obtener los parámetros dimensionales teóricos de los materiales empleados (número de unidades monoméricas entre cadenas, peso molecular promedio, módulo mecánico esperado y en definitiva, el tamaño de red) y compararlos con los reales, medidos experimentalmente a través de sus propiedades mecánicas. En concreto, a partir de la estequiometría, el módulo mecánico de los materiales empleados puede relacionarse con el número de unidades monoméricas promedio de una cadena según la Ecuación 1.1. A partir de TEO utilizando la ecuación Ecuación 1.3 con l=0.158 nm y C=6 [148] se determina la distancia entre nudos en el espacio, TEO (nm), para la serie P(EA-co-HEA). Para obtener los valores experimentales ensayo dinámico-mecánico, DMTA, EXP para y EXP (nm) nos servimos del obtener módulo elástico 74 experimental en la zona de goma. A partir de la Ecuación 1.4 se determinan los valores de las unidades monoméricas experimentales promedio entre cadenas y la distancia entre nudos, EXP y EXP (nm). La Figura 3.1 muestra el espectro dinámico-mecánico de los copolímeros de la serie P(EA-co-HEA) y la Tabla 3.1 los módulos mecánicos experimentales de cada material obtenido del ensayo de DMTA. Material Eexp’(Pa) PEA 1,14E+06 P(EA-co-HEA) 90/10 1,55E+06 P(EA-co-HEA) 80/20 1,53E+6 P(EA-co-HEA) 70/30 1,51E+06 P(EA-co-HEA) 50/50 1,67E+06 P(EA-co-HEA) 30/70 1,74E+06 PHEA 1,99E+06 Tabla 3.1.- Módulo mecánico experimental en estado de goma de los diferentes materiales 75 2 10 9 1 8 7 5 -1 4 3 -2 2 1 0 -100 -50 0 50 100 -3 150 Temperature (ºC) Figura 3.1.- Módulo de almacenamiento (símbolos sin relleno) y tangente de pérdidas (símbolos con relleno) del copolímero P(EA–co–HEA) 50/50, () y las redes de homopolímero de PEA () y de PHEA () log (tan ) log (E'/ Pa) 0 6 76 Como se puede observar en la Tabla 3.2 y Figura 3.2 los datos experimentales caen por debajo de la predicción teórica y esto puede ser atribuido a la existencia de enlaces físicos entre cadenas; estos puntos en los que las cadenas se enmarañan actúan como puntos de entrecruzamiento que se añaden a los entrecruzamientos químicos, incrementándose el módulo mecánico respecto de la predicción teórica. TEO (nm) EXP (nm) TEO EXP PEA 98 91 3,83 3,69 P(EA-co-HEA) 90/10 97 70 3,80 3,24 P(EA-co-HEA) 80/20 95 70 3,77 3,23 P(EA-co-HEA) 70/30 94 72 3,74 3,28 P(EA-co-HEA) 50/50 91 65 3,69 3,12 P(EA-co-HEA) 30/70 88 61 3,64 3,03 PHEA 85 53 3,56 2,83 Tabla 3.2.- Número de unidades monoméricas y tamaños de malla teóricos y experimentales en función de la composición 77 100 4,00 90 80 3,50 70 60 3,25 50 ditsancia entre nudos, unidades mooméricas promedio, 3,75 3,00 40 30 2,75 0 10 20 30 40 50 60 70 80 90 100 % HEA, en el copolímero Figura 3.2.- Representación gráfica de las unidades monoméricas promedio (: teórico; : experimental) y distancia entre nudos (: teórico; : experimental) La teoría de la copolimerización permite estimar, a través del modelo terminal, la composición de la cadena de copolímero una vez finalizado el proceso de polimerización [47],[48]. La fracción molar de monómero Fi en el copolímero en términos de la fracción molar de monómero en la mezcla de reacción fi y los índices de reactividad ri resulta ser: F1 F2 = f1 (f1 r1 + f2 ) f2 (f2 r2 + f1 ) Ecuación 3.1.- Fracciones molares esperadas (F1 y F2) en un copolímero en función de los índices de reactividad (r1 y r2) y de las fracciones molares (f1 y f2) en la mezcla inicial 78 La composición predicha por el modelo de terminal se muestra en la Figura 3.3. 1 0,9 0,8 0,7 xEA cop 0,6 0,5 0,4 0,3 0,2 0,1 0 0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1 xEAo Figura 3.3.- Fracción molar de unidades de EA en el copolímero predicha por el modelo terminal (xEAcop) en función de la fracción molar de monómero de EA en la mezcla reactiva (xEA0). La línea recta sería la composición de un copolímero al azar en el cual la fracción de monómeros en la mezcla original se mantiene también en el material polimerizado La predicción teórica se encuentra por debajo de la línea recta que da la composición aleatoria de la cadena de copolímero (es decir, el copolímero final manteniendo la misma proporción de los dos componentes presentes en la mezcla de reacción original). Esto significa que el monómero más reactivo, el HEA, va a consumirse en mayor proporción durante la formación del copolímero y al final quedará una cantidad de EA que polimerizará formando un bloque de PEA. Por tanto, 79 tenemos que esperar que nuestras redes de copolímeros sean en alguna medida copolímeros de bloque, un bloque de copolímero P(EA-co HEA) y otro de homopolímero de PEA, lo que puede inducir la separación de fases. La separación de fases se observa con frecuencia en copolímeros de bloque y es una propiedad que se ha utilizado frecuentemente aplicaciones biomédicas, tal como la liberación controlada de fármacos, así como en otras aplicaciones industriales. La tendencia a la separación de fases se rige por el criterio de equilibrio termodinámico. A una temperatura y presión dadas el estado de equilibrio es el del mínimo de energía libre (G = H-TS, siendo H la entalpía y S la entropía). La mezcla polimérica de dos tipos diferentes de cadenas de polímero será homogénea si la energía libre de la mezcla es menor que la de los componentes separados. En sistemas poliméricos el incremento de la entropía de mezcla es mucho menor que en los líquidos de bajo peso molecular debido a que el número de posibilidades de organizar las cadenas de polímero en el volumen que ocupan también es pequeño. Por lo tanto, la contribución de la entropía a la disminución de la energía libre en la mezcla es modesta y rara vez puede compensar el incremento positivo de entalpía. Esto es lo que se puede esperar en nuestros copolímeros ya que una de las fases consistiría en una cadena de copolímero rico en HEA y, por lo tanto, hidrófila, mientras que el otro es principalmente EA y, por lo tanto, hidrófoba. La repulsión entre secuencias hidrofílicas e hidrofóbicas produce una contribución positiva de la entalpía de la mezcla a la energía libre y, por lo tanto, contribuye a separar los dos bloques de copolímero. Sin embargo, la separación de fases necesita la capacidad de los bloques de copolímeros de difundir y agregarse para formar dominios de composición uniforme, y eso no es fácil debido a los bloques relativamente cortos EA que se pueden esperar de los cálculos que se muestran en la Figura 3.3 y del hecho de que las cadenas de copolímero estén reticuladas. Por tanto, un primer paso en la caracterización de los copolímeros será determinar si la 80 separación de fases tiene lugar en nuestras redes de copolímero seco, utilizando DSC, DMTA y ensayos de AFM. La técnica dinámico-mecánica, DMTA, es apropiada para investigar la separación en nano-fases ya que incluso dominios muy pequeños pueden mostrar su propio proceso de relajación principal asociado a su transición vítrea; ésta se manifiesta como un pico en el gráfico isócrono de la tangente de pérdidas frente a la temperatura, con una caída simultánea del módulo elástico. La Figura 3.1 muestra la dependencia con la temperatura de la parte real del módulo de elasticidad, E', y la tangente de pérdidas, tan , del PEA, PHEA, y de la red del copolímero BH050. La caída de alrededor tres órdenes de magnitud en el módulo de elasticidad es característica de la relajación principal de las cadenas de polímero que separa el estado vítreo a bajas temperaturas -en las que estos polímeros tienen la apariencia de un sólido rígido con un valor característico del módulo elástico alrededor de 9 5x10 Pa- y el estado de goma a temperaturas por encima de la transición, en el que los materiales presentan un comportamiento elastomérico y la deformación elástica bajo una fuerza aplicada dada es, en esta región de temperatura, alrededor de mil veces más grande que en el estado vítreo para nuestras muestras. La posición de la relajación principal, dinámico-mecánica o , se puede caracterizar por el máximo del pico que se muestra en la tangente de pérdidas, T . Como se puede observar en la Figura 3.1, la relajación aparece a temperaturas más bajas en el PEA que en la red de PHEA y a temperaturas intermedias en los copolímeros. Debido a que la relajación es producida por un movimiento molecular cooperativo en la que una serie de segmentos de cadena pertenecientes a la misma cadena o a una vecina, con un volumen de material de algunas decenas de nanómetros cúbicos, la aparición de un pico único en la tangente de pérdidas en la relajación principal del copolímero indica que los segmentos de ambos componentes 81 están participando en los movimientos cooperativos, por lo que se puede decir que la mezcla es homogénea en la escala nanométrica. Merece la pena señalar que las mediciones se muestran en la Figura 3.1 corresponden a las muestras secas. Como se muestra en dicha figura todo indica que en el caso del copolímero P(EA-co-HEA) 50/50 no existe separación de fases en estado seco, ya que la forma de la tangente de pérdidas y del módulo elástico es similar en el copolímero a las redes de homopolímeros puros. Para demostrar la validez de los resultados se sintetizó un sistema polimérico similar, empleando el EA y el ácido metacrílico en proporciones 90/10 y 80/20. El sistema P(EA-co-MAAc) 80/20 es un copolímero que pierde su transparencia lo que indica que, posiblemente, esté separado en fases. Efectivamente, en el caso de los copolímeros de P(EA-co-MAAc) la situación cambia. En la Figura 3.4 la forma del pico de la tangente de pérdidas del copolímero que contiene un 10% MAAc indica la posible superposición de dos picos. Para aclarar aún más este hecho, un copolímero que contenía un 20 % MAAc se incluyó en el estudio. Como se muestra en dicha figura aparecen claramente dos máximos en el gráfico de la tangente de pérdidas. La baja temperatura del que aparece en torno a los 0ºC se puede asociar a una fase rica en PEA ya que el pico aparece a la misma temperatura que en el homopolímero PEA (Figura 3.4). La relajación de alta temperatura corresponde a un copolímero aleatorio de EA y MAAc. La relajación principal dinámico-mecánica de la red de homopolímero PMAAc no se muestra ya tiene lugar a temperaturas muy altas, después del inicio de la degradación del polímero. 82 2 10 9 1 8 7 5 -1 4 log (tan ) log (E'/ Pa) 0 6 3 -2 2 1 0 -100 -50 0 50 100 -3 150 Temperature (ºC) Figura 3.4.- Módulo de almacenamiento (símbolos sin relleno) y tangente de pérdidas (símbolos con relleno) del homopolímero de PEA () y de los copolímeros P(EA–co–MAAc) que contienen 10% () y 20% () de MAAc La Figura 3.5 representa los termogramas DSC para la serie P(EA-coHEA) en estado seco. Los termogramas muestran claramente un salto único en la capacidad calorífica en los copolímeros situados a temperaturas 83 intermedias entre las temperaturas de transición vítrea de los dos homopolímeros, PEA y PHEA. Por lo tanto, las mediciones calorimétricas no muestran ningún signo de separación de fases en las muestras secas, en consonancia con los resultados del ensayo dinámico-mecánico. PEA 2 mW endo P(EA-co-HEA) 90-10 P(EA-co-HEA) 80-20 P(EA-co-HEA) 70-30 P(EA-co-HEA) 50-50 P(EA-co-HEA) 30-70 PHEA -60 -50 -40 -30 -20 -10 0 10 20 30 40 50 60 70 80 90 100 Temperature (ºC) Figura 3.5.- Medidas de DSC de la serie P(EA-co-HEA) en estado seco Se midió la Tg en cada termograma de la Figura 3.5 a partir del punto medio de la transición y se ha representado en la Figura 3.6 junto con la ecuación de Couchman-Karasz, Ecuación 1.7. La dependencia de la temperatura de transición vítrea en una mezcla es no-lineal y precisamente el hecho de que Tg cambie con la composición como lo hace indica que se trata de un sistema homogéneo. 84 290 285 280 T(K) 275 270 265 260 255 0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1 wEA Figura 3.6.- Temperaturas de transición vítrea experimentales de los diferentes copolímeros de la serie en estudio () y comparación con la Tg predicha por la ecuación de Couchman-Karasz () La presencia de grupos hidrófilos afecta a la respuesta biológica no sólo a través de su influencia sobre el contenido de agua de equilibrio o la tensión superficial, sino que la distribución de estos grupos en la superficie podría ser importante también. Dominios alternos hidrofílicos e hidrofóbicos afectan la absorción de proteínas [149],[97] y, posiblemente, a través de éstas a la adhesión celular y la proliferación [45]. Aquí reside la razón por la que la nanomorfología de los copolímeros debe caracterizarse. Para corroborar la hipótesis de que la serie P(EA-co-HEA) no presentaba separación de fases en estado seco se decidió realizar sobre la muestra P(EA- co-HEA) 50/50 –una de las que podría presentar una estructura bifásica 85 según el modelo terminal de la teoría de la copolimerización- un ensayo de microscopía de fuerza atómica para revelar la posible existencia de nanodominios. Figura 3.7.- Imagen de AFM de fase (izquierda) y topografía (derecha) tomada en modo tapping de la muestra seca 50/50; área barrida 1 m x 1 m Como se puede observar se ve una superficie homogénea, tanto en la imagen de fase como en la de altura en una escala de 1x1 m. No existe evidencia de una separación de fases en este sistema, por lo que, en estado seco, ninguna de las técnicas que han servido para caracterizar los xerogeles de la serie P(EA-co-HEA) ha mostrado evidencia de una posible separación de fases. Como conclusión de este apartado podemos decir que a pesar de la diferente reactividad de los dos monómeros que forman el copolímero en la red de P(EA-co-HEA), que hace que se formen bloques de homopolímero de PEA en la red, éstos no llegan a agruparse para formar nanodominios sino que quedan disueltos en la parte de la red formada por las cadenas de copolímero. Al no formar agregados de dimensiones nanométricas, no hay movimientos conformacionales en los que sólo intervengan segmentos de EA 86 y no se produce una transición vítrea independiente, producida por ellos. 3.1.2.- Propiedades de los soportes inmersos en medio acuoso. Separación de fases por interacción hidrófoba El objeto de esta sección es el estudio de las propiedades superficiales de los materiales inmersos en medio acuoso, como en los cultivos celulares. Se caracterizará en primer lugar la tensión superficial de los substratos. Luego se analizará la interacción del agua con el copolímero P(EA-co-HEA) para comprobar si la presencia de agua induce cambios conformacionales en el copolímero reorganizando sus cadenas para minimizar la exposición de sus partes hidrófobas al agua, lo que induce la separación de fases por interacción hidrófoba. Es bien sabido que las propiedades físicas de la superficie afectan a la adhesión celular debido a su influencia sobre la conformación de las proteínas que se adsorben sobre ella y, en particular, tiene especial relevancia la hidrofilicidad del substrato así como su tensión superficial. El contenido de agua en equilibrio, EWC, (masa de agua absorbida por unidad de masa de polímero) de los copolímeros de este estudio se muestra en la Tabla 3.3. La red de PEA absorbe menos del 2% de agua; por lo tanto, el PEA puede considerarse un material hidrófobo. La absorción de agua aumenta de manera uniforme con el contenido de acrilato de hidroxietilo, HEA, en los copolímeros de la serie P(EA-co-HEA). 87 Composición EWC (x100) PEA 1.7 ± 0.4 0.1 ± 2.3 31.0 ± 1.6 31±4 P(EA-co-HEA) 90/10 2.3 ± 0.3 1±3 26 ± 3 28±6 P(EA-co-HEA) 80/20 2.8 ± 0. 3 1 ± 11 29 ± 5 30±15 P(EA-co-HEA) 70/30 7.6 ± 0.9 1±4 31 ± 4 32±8 P(EA-co-HEA) 50/50 18.2 ± 1.7 3±9 33 ± 5 36±14 P(EA-co-HEA) 30/70 40.6 ± 1.3 3±6 35 ± 5 37±11 PHEA 134 ± 5 19 ± 7 27 ± 5 46±12 p (mN/m) d (mN/m) s (mN/m) Tabla 3.3.- Composición, EWC, componente polar, dispersiva y tensión superficial medida tras inmersión en agua líquida a 37ºC durante 48 horas También se muestra las medidas del ángulo de contacto de los diferentes substratos en presencia de formamida. El mismo estudio no podría llevarse a cabo con el agua, ya que se absorbe completamente por el PHEA y los copolímeros que contienen PHEA. La alta polaridad de la formamida explica el alto ángulo de contacto entre la gota de formamida y la superficie de PEA, cerca de 90 grados. También se observa que el aumento en el contenido de HEA de los copolímeros produce una disminución continua del ángulo de contacto que es sólo 46 grados en la superficie PHEA, como se muestra en la Figura 3.8. 88 angulo de ocntacto con formamida (º) 100 90 80 70 60 50 100 90 80 70 60 50 40 30 20 10 0 40 %HEA Figura 3.8.- Medida del ángulo de contacto de una gota de formamida sobre los copolímeros de P(EA–co–HEA) Los valores del ángulo de contacto permiten la determinación de la tensión superficial del polímero siguiendo el método de Owens y Wendt [142]. La teoría de Owens y Wendt divide la energía superficial en dos componentes: la energía superficial debida a las interacciones dispersivas y la energía superficial debida a las interacciones polares. Esta teoría se deriva de la combinación de relación de Young, que relaciona el ángulo de contacto con las energías superficiales del sólido y líquido con la tensión interfacial, y la ecuación de Good, que relaciona la tensión interfacial con la componente polar y dispersiva de la energía superficial resultando la Ecuación 3.2: σ L (cos θ + 1) 2 σ D L = σ SP σ PL σ Ecuación 3.2 D L + σ SD 89 Si se dibuja un gráfico de σ L (cos θ + 1) vs. 2 σD L σ PL , las componentes σ DL polar y dispersiva se pueden obtener a partir de la pendiente y la ordenada en el origen. En este trabajo se midió el ángulo de contacto con diyodometano, formamida y glicerol, cuyos resultados en el PEA fueron 58, 87 y 99 grados, respectivamente. El gráfico de la Figura 3.9 representa la componente polar y dispersiva de la tensión superficial a partir de la pendiente y la ordenada en el origen de una línea recta ajustada a los resultados experimentales para el PEA, PHEA, y BH050. 12,0 11,0 2 σ DL σ L (cos θ + 1) 10,0 9,0 8,0 7,0 6,0 5,0 0,20 0,40 0,60 0,80 σ 1,00 1,20 1,40 P L σ DL Figura 3.9.- Representación gráfica para la determinación de la componente polar y dispersiva de la tensión superficial siguiendo el método de Owens y Wendt del PEA (), BH050 () y PHEA () La energía superficial de la serie P(EA-co-HEA), mostrada en la Tabla 3.3, disminuye de manera uniforme con el incremento del componente 90 hidrofóbico, PEA. La componente polar de la tensión superficial, con un alto valor para el PHEA, hidrófilo puro, disminuye hasta llegar a ser casi nula en el componente hidrófobo. La situación conformacional de un copolímero en estado seco puede ser modificada por la absorción de agua, por lo que se estudió distintas propiedades físicas en muestras hidratadas con diferentes contenidos de agua. Este trabajo se llevó a cabo en la muestra BH050. Con el fin de entender la organización de la red de copolímero en presencia de agua diferentes réplicas de P(EA-co-HEA) 50/50 se hincharon en agua líquida durante diferentes tiempos para alcanzar contenidos de agua crecientes, hasta el EWC. En un sistema homogéneo en el que no existe separación de fases cabe esperar un descenso monótono de la temperatura de transición vítrea conforme aumenta el contenido de agua en el copolímero. Existen diferentes modelos que tratan de correlacionar la Tg del sistema copolímero-solvente, como es el caso de la ecuación de Couchman-Karatz, Ecuación 1.7 Tg = w cop ΔC p,cop Tg,cop + w solv ΔC p,solv Tg.solv w cop ΔC p,cop + w solv ΔC p,solv La transición vítrea de las muestras hidratadas con diferentes contenidos de agua se muestra claramente en las curvas de enfriamiento de DSC (Figura 3.10 y Figura 3.11. La temperatura de transición vítrea Tg se desplaza con el aumento de contenido de agua primero hacia temperaturas más bajas hasta un contenido de agua de w = 0.016, como cabe esperar según lo descrito anteriormente. Pero a continuación, la Tg del sistema aumenta, y la temperatura de transición vítrea de la muestra con w = 0.031 es casi la misma que la del xerogel. 91 endo > w=0 w=0.004 w=0.016 w=0.022 w=0.031 1 J/gºC -100 -80 -60 -40 -20 0 20 40 Temperature (ºC) Figura 3.10.- Curvas de enfriamiento de DSC a 10 C/min del sistema P(EA-coHEA) 50/50 con diferentes contenidos de agua (el contenido se indica en cada curva) endo > 92 w=0.050 w=0.061 w=0.109 w=0.159 1 J/gºC -100 -80 -60 -40 -20 0 20 40 Temperature (ºC) Figura 3.11.- Curvas de enfriamiento de DSC a 10 C/min del sistema P(EA-coHEA) 50/50 con diferentes contenidos de agua (el contenido se indica en cada curva) Para contenidos de agua más altos la Tg disminuye con el aumento de contenido de agua muy ligeramente, la diferencia entre la Tg de la muestra hinchada en inmersión en agua líquida (el punto experimental con el contenido de agua más alto) y la de la muestra seca es sólo 7 C (Figura 3.12). El mismo comportamiento se muestra en las curvas de calentamiento para contenidos bajos de agua (por debajo de 0.031). 93 10 I II III 0 Tg (ºC) -10 -20 -30 -40 0,00 0,05 0,10 0,15 0,20 0,25 0,30 w' Figura 3.12.- Temperatura de transición vítrea del sistema P(EA-co-HEA) 50/50 en función del contenido de agua referido al componente hidrófilo, w’ (cuadrados). La línea sólida representa la predicción de la ecuación de Couchman-Karasz para mezclas homogéneas del polímero-agua. Los círculos representan la Tg en función del contenido de agua, w, para las muestras con w=0.004 y 0.016. Se describen tres áreas en el gráfico cualitativamente diferentes: (I) plastificación de los dominios hidrófilos en el copolímero, (II) cambio abrupto de la Tg como consecuencia de la interacción hidrófoba, (III) transición vítrea de los dominios hidrófobos Las curvas de enfriamiento realizadas con muestras que contienen contenidos de agua más altos son análogas a la mostrada en la Figura 3.10 para w = 0.031. La Figura 3.13 muestra las curvas de DSC correspondientes a la muestra hinchada hasta equilibrio sumergida en agua líquida, w = 0.159. Vale la pena señalar que no se observa cristalización del agua durante el 94 enfriamiento. Sin embargo, el posterior calentamiento de las muestras con contenidos de agua por encima de w = 0.031 muestra algunos indicios de cristalización del agua. No se manifiesta claramente un pico exotérmico debido a la superposición de la transición vítrea del polímero con la cristalización. Sin embargo un pequeño pico de fusión por debajo de 0ºC, cuya fenomenología concuerda con agua cristalizada en equilibrio con el gel, indica que una pequeña cantidad de agua absorbida cristaliza durante el endo > calentamiento [150]. w=0.159 1 J/gºC -100 -80 -60 -40 -20 0 20 40 Temperature (ºC) Figura 3.13.- Curvas de DSC de enfriamiento (abajo) y calentamiento (arriba) de la muestra P(EA-co-HEA) 50/50 inmersa en agua líquida, w=0.159 La plastificación de la red de polímero para bajos contenidos de agua también se representa claramente en los resultados dieléctricos. Se muestran 95 a continuación sólo algunos resultados característicos de DRS. La Figura 3.14 muestra la dependencia con la temperatura de la componente imaginaria de la permitividad dieléctrica medida a 100 Hz. El pico en ε'' es característico de la relajación dieléctrica principal o α, relacionado con los reordenamientos conformacionales cooperativos de las cadenas que constituyen la red de polímero. Para contenidos bajos de agua, hasta w = 0.013, aparece claramente un máximo en ε'', aunque en el lado de alta temperatura se ve afectado por la contribución de la corriente continua (cc), que conduce a un aumento continuo de ε'' al aumentar la temperatura. Conforme el contenido de agua aumenta la contribución de la corriente continua es más importante y, de hecho, en las muestras con w = 0.107 en adelante el pico dipolar no se puede distinguir. La plastificación de la red de polímero por las moléculas de agua produce que la relajación cambie hacia temperaturas más bajas. Vale la pena señalar que en la relajación dieléctrica el intervalo de contenidos de agua que se puede analizar sólo cubre contenidos de agua bajos y el aumento de la temperatura de transición vítrea observado en DSC para contenidos superiores a w = 0.016 no se puede observar en DRS. 96 3 log '' 2 1 0 -1 -2 -60 -40 -20 0 20 40 60 Temperature (ºC) Figura 3.14.- Componente imaginaria de la permitividad dieléctrica del copolímero P(EA-co-HEA) 50/50 seco () y con diferentes contenidos de agua: w = 0.007 (), w = 0.013 () y w = 0.107 () El analizador dinámico-mecánico utilizado en este trabajo no dispone de control de humedad. Las muestras analizadas estaban en condiciones ambiente y el control del contenido de agua de la muestra no pudo ser tan preciso como en los experimentos de DSC, donde la muestra se encierra en cápsulas selladas, o en los experimentos dieléctricos, donde la muestra está entre los electrodos de la celda. En estos experimentos se utilizó una muestra de P(EA-co-HEA) 50/50 con el contenido máximo de agua, una de composición intermedia y una muestra de seca. Como se muestra en la Figura 3.15 la temperatura del máximo de la tangente de pérdidas es menor en las redes hinchadas que en el xerogel, pero la diferencia máxima es de sólo 97 alrededor de diez grados, similar a lo encontrado en experimentos de DSC. La característica principal que se encuentra en los experimentos dinámicomecánicos es la presencia de un segundo pico que aparece en el gráfico de la tangente de pérdidas de la muestra con w = 0.159 alrededor de -17ºC. Este pico se analiza en la siguiente sección, como consecuencia del proceso de separación de fases en la muestra debido a la presencia de agua. 98 10 2,0 9 1,5 8 1,0 0,5 6 5 0,0 4 log tan log (E' / Pa) 7 -0,5 3 -1,0 2 -1,5 1 0 -50 -2,0 -40 -30 -20 -10 0 10 20 30 Temperature (ºC) Figura 3.15.- Dependencia del modulo elástico con la temperatura (símbolos sin relleno) y tangente de pérdidas (símbolos rellenos) de la muestra P(EA-co-HEA) 50/50 en estado seco (), una muestra con contenido de agua, w = 0.007 (), y una muestra hinchada en inmersión en agua líquida, w= 0.159 () 99 Se realizó un experimento de AFM con una muestra del copolímero P(EA-coHEA) 50/50 en estado seco (que ya analizamos en el apartado 3.3.1 pero repetimos aquí para facilitar la comparación), y con una muestra hinchada previamente equilibrada en agua líquida. En el primer caso se realizó con las condiciones indicadas en el apartado 2.2.1.3 y, en el segundo caso, con la punta inmersa en una gota de agua depositada sobre la muestra. La imagen de la topografía y ángulo de fase indica claramente que la superficie del xerogel es completamente homogénea (Figura 3.12a, c). Sin embargo, cuando la muestra se hinchó previamente en agua líquida se revela claramente un patrón con dominios alternos de cientos de nanómetros con diferentes propiedades viscoelásticas (Figura 3.12b, d). 100 Figura 3.16.- Imágenes de fase de AFM del sistema P(EA-co-HEA) 50/50, (a) y (b), y topografía, (c) y (d), obtenidas en modo tapping del copolímero xerogel, (a) y (c), y de la red hinchada en agua del mismo copolímero, (b) y (d); área barrida 1 mx1 m En la red de copolímero P(EA-co-HEA) 50/50, según predice la teoría de la copolimerización, la cantidad fracción de EA en la cadena de copolímero es ligeramente inferior al de la mezcla original, lo que resulta en una red formada por cadenas de copolímero donde no se distribuyen al azar el EA y el HEA, sino que pequeños bloques de HEA están presentes preferencialmente. Sin embargo, la transición vítrea y las relajaciones principales dieléctricas y dinámico-mecánicas del xerogel así como el ensayo de AFM no muestran indicios de separación de fases en estado seco, es decir, 101 fluctuaciones de composición espacial con distribución unimodal. La longitud de cooperatividad en la transición vítrea se ha evaluado en polímeros amorfos que debe rondar unos pocos nanómetros. Esto determina un orden de magnitud para el tamaño de los agregados más pequeños que un sistema separado en fases debe tener para presentar transiciones vítreas de las fases individualmente. Aunque los grupos polares (hidrofílicos) a lo largo de las cadenas no están distribuidos al azar, no hay ninguna señal de la agregación de grupos polares o no polares con unas dimensiones suficientes para producir transiciones vítreas independientes en el xerogel. Cuando el sistema BH050 se hincha con diferentes contenidos de agua, en las primeras etapas de la absorción, hasta que w = 0.016, la transición vítrea se desplaza a temperaturas más bajas como corresponde a la mezcla homogénea de cadenas de copolímero y agua. Para enfatizar esta característica la predicción de la ecuación Couchman-Karasz Tg = (1 - w' )ΔCp,cop Tg,cop + w' ΔCp,agua Tg.agua (1 - w' )ΔCp,cop + w' ΔCp,agua de la temperatura de transición vítrea de la red de copolímero en función del contenido de agua está representada en la Figura 3.12. En esta ecuación w' es la fracción de masa de agua por unidad de masa de HEA en el sistema, Tg,agua = 134 K y -1 -1 Cp,agua = 1,94 J·g K se han tomado de [151], mientras que los valores para el red de copolímero de Tg,cop = 275,1 K e -1 0.374 J·g K -1 Cp,cop = se han obtenido de forma experimental en este trabajo a partir de la curva de DSC correspondiente al xerogel (Figura 3.10). Es importante remarcar que la línea en la Figura 3.12 no es un ajuste de los puntos experimentales, sino una predicción. La buena concordancia entre los puntos experimentales obtenidos a partir de mediciones de DSC para contenidos de agua bajos y la curva teórica se puede interpretar en el sentido de que la cantidad inicial de agua que entra en el sistema se mantiene 102 homogéneamente mezclada con las unidades hidrófilas de las cadenas de copolímero en la red (región I en la Figura 3.12). La cantidad máxima de agua en este régimen es bastante pequeña, w = 0.016, y corresponde sólo a una molécula de agua por cinco grupos OH en la cadena polimérica. Al sobrepasar esta modesta cantidad de agua (w = 0.016) un rápido aumento de la temperatura de transición vítrea se observa en la red de polímero con respecto a la esperada para la mezcla homogénea de moléculas de agua y cadenas poliméricas (región II en la Figura 3.12). La influencia de la interacción hidrófoba sobre la temperatura de transición vítrea se ha tenido en cuenta cuantitativamente para redes de polímero con un componente hidrófilo y uno hidrófobo usando la ecuación Kwei [74], [150], [152]-[154]. Sin embargo, el fuerte aumento de la transición vítrea en este caso en una estrecha banda de contenidos de agua no puede ser explicado por la mera contribución de la interacción entre las unidades monoméricas hidrofóbicas en un sistema homogéneo. Más bien, el comportamiento se explica por la separación de fases entre una fase no plastificada hidrófoba y la fase hidrófila que contiene agua, que conduce a la formación de (nano)agregados de homopolímero PEA en las cadenas de copolímero. Las temperaturas de transición vítreas experimentales representadas en la Figura 3.12 se han encuadrado en tres regiones cualitativamente diferentes: la región I describe la plastificación inicial del copolímero como consecuencia de las primeras moléculas de agua que entran en el sistema. El alojamiento inicial de agua en el sistema ya induce algunos reordenamientos conformacionales que conducen a la incorporación de agua preferentemente junto a los segmentos hidrófilos, como se confirma por el hecho de que los puntos experimentales se ajusten a la ecuación de Couchman-Karasz cuando se representan frente a w' en lugar de frente a w. La región II se considera que es consecuencia de un régimen de transición; la incorporación adicional de agua en el sistema da lugar a la llamada interacción hidrofóbica que lleva a un proceso de separación en nanofases que aumenta la temperatura de 103 transición vítrea del copolímero. Por último, la región III describe el sistema para altos contenidos de agua y la Tg medida experimentalmente se atribuye a la vitrificación de los (nano) agregados hidrófobos. La organización de los grupos hidrofóbicos e hidrofílicos en estos agregados (región III en la Figura 3.12) debe ser bastante imperfecta debido a la falta de regularidad a lo largo de las cadenas de copolímero (como se obtiene de la interpretación del modelo de terminal, Figura 3.3) y la presencia de puntos de entrecruzamiento. Sin embargo, el número de grupos hidrófilos incluido en los dominios hidrófobos es lo suficientemente bajo como para obligar a que el núcleo de los agregados no contengan moléculas de agua, o al menos, menos agua que la media. El aumento adicional en la absorción de agua (de 0.05 en adelante) reduce la temperatura de transición vítrea observada ligeramente (Figura 3.12), lo que significa que primero, la transición vítrea representada en los termogramas experimentales para estos contenidos de agua debe ser atribuida a los dominios ricos en unidades de componente hidrófobo y, en segundo lugar, que las moléculas de agua se acumulan fuera de ellos, es decir, estos dominios organizados no se plastifican por la presencia de agua en la red de copolímero. Vale la pena señalar que el hecho de que la transición vítrea de estos dominios hidrófobos se pueda detectar por DSC demuestra que su tamaño debe estar al menos en el orden de magnitud de la longitud de la cooperatividad en la temperatura de transición vítrea (aprox. 10 nm) [155]. Por otro lado, no se detecta transición vítrea de los dominios hidratados mediante DSC, como se muestra en la curva de enfriamiento de la Figura 3.13, para la muestra con el mayor contenido de agua. En este ejemplo, el contenido de agua es de alrededor de dos moléculas de agua por grupo hidroxilo. En el caso de una red de PHEA puro reticulado con 2% en peso de EGDMA la cantidad de agua absorbida en equilibrio por la muestra sumergida en el agua líquida es de alrededor de diez moléculas por grupo-OH [45]. En el caso de la red de copolímero de este trabajo, la presencia de los agregados hidrófobos limita la capacidad de 104 expansión de la red y, por otra parte, un número de grupos hidroxilo debe quedar atrapado dentro de la fase hidrófoba y, por lo tanto, inaccesibles por las moléculas de agua. Se puede argumentar que la movilidad en la fase hidrófila no permite los movimientos cooperativos necesarios para producir la transición vítrea. La transición vítrea de la fase hidrófoba se lleva a cabo a temperaturas de alrededor de la de la red seca (aprox. -15ºC) [156]. Esto significa que a temperaturas más bajas (en el intervalo de temperaturas en el que se espera que la transición vítrea de la red hidrófila plastificada tenga lugar) los segmentos de cadena que pertenecen a la fase hidrófila - que está hinchada con agua - se unen a la parte vitrificada hidrófoba de la red de polímero. La situación es de alguna manera similar a la de la fase amorfa intraesferulítica de un polímero semicristalino en el que la movilidad de las cadenas amorfas está fuertemente limitada por la presencia de los cristales [157], [158]. Esta falta de movilidad puede explicar la dificultad de la cristalización del agua en el enfriamiento también. Sin embargo, alguna indicación de los reordenamientos moleculares en esta fase se puede ver a partir de los experimentos dinámico-mecánicos (Figura 3.15), por el pico que aparece en la tangente de pérdidas alrededor de -17ºC y la disminución gradual del módulo de elasticidad en el intervalo de temperatura entre -30 y 20ºC. Hay que tener en cuenta que la relajación secundaria no aparece en este intervalo de temperaturas en hidrogeles de PHEA puro [159]. 105 Figura 3.17.- Representación de la situación molecular. (a) de una red ideal compatible con los resultados experimentales (ausencia de cualquier tipo de separación de fases) y con la predicción del modelo de terminal. La cadena de copolímero consta de unidades hidrofílicas en la que alternan algunos segmentos hidrófobos. Hay que tener en cuenta que en una red de polímero las cadenas no están estiradas, sino de forma aleatoria y enredadas entre ellas. Además, cuando existen puntos de entrecruzamiento al azar no son de longitud uniforme. (b) organización inducida por el agua en agregados hidrófilos e hidrófobos. El dominio I representa un agregado hidrofóbico. El dominio II representa un agregado hidrofóbico con algunos segmentos hidrófilos atrapadas en su interior. El dominio III representa un agregado hidrófilo vinculado a uno hidrófobo que no puede participar en movimientos conformacionales como un dominio puro La separación de fases inducida por la presencia del disolvente se observa claramente en las imágenes AFM. La Figura 3.16 muestra el área de escaneado en la misma muestra antes (a y c) y después (b y d) de su inmersión en agua líquida. La homogeneidad del copolímero en el estado de xerogel se verifica en la imagen de fase (se debe tener en cuenta la baja escala para el ángulo de fase). Los agregados brillantes en las imágenes de fase se identifican con los dominios hidrófobos cuyo tamaño es lo 106 suficientemente grande como para dar cuenta de una relajación adicional y de una transición vítrea en DSC. La formación de los agregados tiene un efecto también sobre la señal de la topografía en el AFM. Del mismo modo, la imagen de topografía para el xerogel es bastante uniforme hasta aproximadamente 100 nm y la presencia de agua induce la formación de agregados que sobresalen de la superficie aproximadamente 80-100 nm. El peso molecular entre reticulaciones se puede estimar por el modelo afín de la teoría de la elasticidad de caucho (Ecuación 1.4) [160] Mc 3ρRT E donde ρ es la densidad de la red, R la constante de los gases, T es la temperatura en el estado de goma y E el módulo mecánico. De acuerdo con los datos en la Figura 3.15, el peso molecular entre nudos en la red seca da como resultado aprox. 1500 g/mol. Una segregación más allá del tamaño de la red requiere una distorsión mayor de la red, y la longitud de la cadena correspondiente a Mc ≈ 1500 g/mol es del orden de diez nanómetros, es decir, mucho más pequeña que el tamaño de los dominios en la superficie detectados por AFM (unos cientos de nanómetros). Por lo tanto, la red debe ser bastante heterogénea como consecuencia de la cinética de polimerización que se traduce en una red mucho más escasa en HEA cerca de la superficie. Además, la teoría de la elasticidad del caucho permite calcular el módulo de la red hinchada Ew a partir del conocimiento exclusivamente del módulo del xerogel E y la fracción de volumen de disolvente en el gel de Ew (1 - 1 [161] 1 1 )3 E Teniendo en cuenta el contenido de agua de equilibrio del gel hinchado en agua líquida (w = 0.159), la ecuación anterior predice el módulo de la red 107 hinchada en Ew = 5.12 MPa, mucho más alto que el valor experimental obtenido a partir de la Figura 3.15, Eexp = 1.38 MPa. La diferencia debe ser atribuida a la distribución no uniforme de agua dentro de la muestra, es decir, a la formación de agregados con más agua que la media que conducen a un comportamiento mecánico más pobre. Conclusiones: Como conclusión de cara a los estudios de la respuesta biológica, cabe decir que la superficie del soporte plano o la pared de los poros en un soporte macroporoso va a presentar una alternancia de agregados hidrófilos e hidrófobos con dimensiones del orden de las decenas de nanómetros, es decir mucho más pequeñas que las dimensiones celulares. Esta nanoestructura sin embargo es susceptible de afectar a la respuesta celular porque puede determinar la conformación de las proteínas de adhesión que se distribuyen en la superficie [45], [97]-[102]. 108 3.2.- Cultivos celulares en soportes 2D y 3D Esta sección está basada en los artículos “Future design of a new keratoprosthesis: Physical and biological analysis of polymeric substrates for epithelial cell growth. Campillo-Fernandez, A. J. et al. Biomacromolecules 8, 2429–36 (2007)” y “Analysis of the biological response of endothelial and fibroblast cells cultured on synthetic scaffolds with various hydrophilic/hydrophobic ratios: influence of fibronectin adsorption and conformation. Campillo-Fernandez, A. J. et al. Tissue Engineering Part A, 1331-41 (2009)”. La respuesta biológica a nuestros materiales soporte se ha probado con tres tipos celulares diferentes en los que la adhesión al soporte en cultivo es determinante para su viabilidad, proliferación y mantenimiento del fenotipo. Por una parte, células epiteliales conjuntivales se cultivaron sobre los substratos planos sin pre-impregnación de proteínas adicional a las que llevó el medio de cultivo. Se pretendía analizar la respuesta de las células epiteliales a los diferentes substratos poliméricos, variando el grado de hidrofilia y las cargas eléctricas superficiales. Las muestras se equilibraron en medio de cultivo previamente a la siembra celular para evitar dilataciones del substrato (especialmente en los más hidrófilos) una vez sembradas las células. Por otro lado, se sembraron sobre scaffolds una línea celular de fibroblastos con y sin pre-impregnación de fibronectina, una proteína que favorece la adhesión celular. Los fibroblastos no mostraron diferencias de adhesión entre los scaffolds pre-impregnados y los impregnados. Por el contrario, las células endoteliales humanas de cordón umbilical sí necesitaron de tal pre-impregnación de fibronectina. No es de extrañar ya que incluso los cultivos de este tipo celular en los flascones de cultivo estándar requieren de pre-recubrimiento de gelatina. 109 3.2.1.- Cultivo de células epiteliales conjuntivales, NHC Como se muestra en la Figura 3.18 el número de células adheridas a los diferentes substratos se correlaciona con los grupos-OH presentes en la superficie, es decir, la cantidad de componente hidrófilo. Por lo tanto, para la serie P(EA-co-HEA), cuanto más hidrófobo es el substrato mejor es la adhesión de las células NHC. Todos los substratos con hasta 20% de componente hidrófilo mostraron adherencia mientras que el cultivo de células NHC sobre substratos con más de 20% de componente hidrófilo no mostró adherencia. Después de una semana, las células alcanzaron 90% de confluencia en el PEA, P(EA-co-HEA) 90/10, P(EA-co-HEA) 80/20 y en el control de poliestireno, TCPS, como se muestra en la Figura 3.19. Sin embargo, no se encontraron diferencias significativas entre los diferentes biomateriales analizados para cada punto temporal (test de Kruskal-Wallis, p = 0.393 para el día 3, p = 0.124 para el día 5, p = 0.175 para el día 7). Nº células / soporte (x102) 1600 1400 1200 1000 800 600 400 200 0 0 3 5 7 Días de cultivo Figura 3.18.- Número de células adheridas a los diferentes substratos en función del tiempo. TCPS (), PEA (), P(EA-co-HEA) 90/10 (),P(EA-co-HEA) 80/20 () y P(EA-co-MAAc) 90/10 () Conteo realizado en cámara de Neubauer. Los substratos P(EA-co-HEA) 70/30, P(EA-co-HEA) 50/50, P(EA- co-HEA) 30/70 y PHEA no presentaron adhesión celular 110 a 20x b 20x c 20x Figura 3.19.- Células NHC tras 7 días de cultivo sobre a) PEA, b) P(EA-coMAAc) 90/10 y c) TCPS. Las imágenes están tomadas con una magnificación 20x 111 La viabilidad observada por tinción con azul tripán mostró una gran mayoría de células vivas y sólo algunas de ellas se tiñeron tras 7 días en cultivo sobre el PEA, P(EA-co-HEA) 90/10, P(EA-co-HEA) 80/20, P(EA- co-MAAc) 90/10 y TCPS (Figura 3.20). a b c 10x 20x 10x d e 20x 10x Figura 3.20.- Tinción con azul tripán de las células sobre diferentes substratos: (a) PEA, (b) P(EA-co-HEA) 90/10, (c) P(EA-co-HEA) 80/20, (d) P(EA-coMAAc) 90/10 y (e) control TCPS. Las células fueron viables hasta que alcanzaron la confluencia en una semana. Imágenes tomadas a 10x ó 20x (indicado en cada figura) La morfología celular se analizó mediante tinción con hematoxilina-eosina, que reveló la morfología característica epitelial y poligonal de las células NHC, con un gran núcleo y numerosos nucleolos intensamente teñidos (Figura 3.21). 112 a 40x c 40x b 20x d 40x Figura 3.21.- Morfología de las células NHC cultivadas en diferentes substratos. (a) PEA, (b) P(EA-co-HEA) 90/10, (c) P(EA-co-MAAc) 90/10 y (d) TCPS tras 7 días de cultivo. La morfología de las NHC en estos substratos permaneció inalterada al igual que para el control. Imágenes tomadas a 20x ó 40x (indicado en cada figura) Algunos cultivos se mantuvieron durante más de una semana y las células mantuvieron la morfología típica de las células epiteliales con una tasa de mortalidad esperada debido a la confluencia. Ninguno de los materiales resultó tóxico para el crecimiento celular. Con el fin de analizar la proliferación de las células NHC en los diferentes substratos el kit de ensayo de proliferación celular CyQUANT NF que contenía un colorante de unión al ADN fluorescente nos permitió obtener un registro fotográfico a través de un microscopio de fluorescencia. El 113 seguimiento reveló un alto nivel de proliferación cada vez mayor desde el día 0 al 7 (Figura 3.22). a0d a3d a5d a7d b0d b3d b5d b7d c0d c3d c5d c7d d0d d3d d5d d7d Figura 3.22.- Análisis de proliferación de las células NHC cultivadas en diferentes substratos después de 0, 3, 5 y 7 días. (a, b) las células NHC que crecen en PEA y P(EA-co-HEA) 90/10, respectivamente. Las células proliferaron de manera óptima a lo largo de los días y cierta estratificación se observó el día 7. (c) las células NHC creciendo en P(EA-co-MAAc) 90/10 mostraron un comportamiento similar al del PEA y P(EA-co-HEA) 90/10 con buena adherencia y proliferación (d) células NHC creciendo en PHEA. Muy pocas células se adhirieron y, por tanto, se observó baja proliferación en este substrato 114 La literatura ha demostrado que el impacto sobre la adhesión celular a la superficie de un substrato con grupos hidrófilos depende en gran medida de la distribución en la superficie de estos grupos y del tipo de célula. Mientras que la adhesión celular no es posible en polímeros hidrófilos tales como poli(metacrilato de hidroxietilo) (PHEMA), con fibroblastos y otras líneas celulares, el anclaje celular es factible con polímeros hidrófobos tales como poli(metacrilato de metilo) (PMMA) o poli(metacrilato de etilo) (PEMA) [162]-[166]. En este estudio con células epiteliales conjuntivales (IOBA-NHC), hemos demostrado que cuanto mayor es el contenido de componente hidrófilo en la serie P(EA-co-HEA), menor es la adhesión celular. Los substratos P(EA-coHEA) 90/10 y P(EA-co-MAAc) 90/10 mostraron las mejores propiedades de adhesión; sin embargo, no se observaron diferencias significativas entre todos los materiales hidrófobos analizados. Esto es probablemente debido a la alta desviación estándar obtenida ya que el protocolo de siembra puede hacer variar ligeramente el número final de células en cada soporte para cada experimento. Estos resultados pueden ser comparados con cultivos de condrocitos en copolímeros como el poli(metacrilato de etilo-co-acrilato de hidroxietilo), P(EMA-co-HEA), substratos en los que la separación de fases no está presente; cuanto mayor es el contenido de HEA, menor es la adhesión y proliferación celular. Sin embargo, para el poli(acrilato de etilo-cometacrilato de hidroxietilo), P(EA-co-HEMA), serie en la que hay una separación de fases en forma de alternancia de dominios hidrófobos e hidrófilos, los parámetros del cultivo de condrocitos fueron mejores para la relación 50/50 [45]. En nuestro estudio, las células epiteliales de la conjuntiva no muestran proliferación después de 24 h en PHEA, lo que no es tan evidente en otros tipos de células, tales como condrocitos [45]. Por el contrario, se observa una 115 buena adherencia en el PEA, aunque no tan buena como en el TCPS. Como se observa en la Figura 3.18, hasta un 10-20% de HEA presente en el copolímero permite la adhesión celular, aunque cantidades mayores no muestran el mismo comportamiento. Una posible explicación para esto podría ser que las proteínas de suero que no promueven la adhesión celular se adsorbe preferencialmente sobre el biomaterial o que las proteínas adsorbidas por el polímero adquieren una conformación con sus motivos de unión menos expuestos conforme el contenido hidrófilo aumenta. En este sentido, una vez que se alcanza el umbral de 20% de HEA, las redes poliméricas no permiten la adhesión (o conformación adecuada) de las proteínas y, por lo tanto, la adhesión celular falla. Cuando se añadieron unidades de MAAc al EA, se observaron diferentes resultados. Aunque el contenido de agua de equilibrio del P(EA-co-MAAc) 90/10 es un 50% mayor que en el P(EA-co-HEA) 90/10, es todavía muy pequeño; al mismo tiempo, la componente polar de la tensión superficial aumenta drásticamente debido a la presencia de grupos carboxilo. Por otra parte, los copolímeros con 10% de MAAc muestran separación de fases. Y debido a la naturaleza ácida de este copolímero éste se disocia cuando está en contacto con agua de modo que la superficie se carga negativamente, lo que favorece la adhesión celular [167]. Por un lado, la separación de los dominios hidrófobos puede aumentar la adhesión celular en sustratos con mayor cantidad de agua absorbida [168] en el caso de los condrocitos y, por otra parte, la distribución de cargas negativas afecta a la selección de proteínas absorbidas y su conformación sobre el sustrato. Probablemente, ambos efectos contribuyen a la mejora de la adhesión celular y la proliferación, pero no se puede ser concluyente sobre qué es más importante. No se encontró correlación entre la adhesión celular y el contenido de agua en equilibrio y la energía superficial. De acuerdo con nuestros datos, los copolímeros de P(EA-co-MAAc) 116 muestran mejor adhesión y proliferación celular que el grupo de control, TCPS. Aunque hay muchos estudios sobre la adhesión celular en PMAAc, la combinación con PEA, como en nuestro estudio, es novedosa. También hemos demostrado que tanto la separación de fases como las propiedades de polielectrolito del copolímero P(EA-co-MAAc) ayuda a la adhesión y proliferación de las células epiteliales conjuntivales. 3.2.2.- Cultivo de fibroblastos, MRC-5 Efecto del balance hidrófobo/hidrófilo. La línea celular de fibroblastos humanos MRC-5 fue sembrada en scaffolds de P(EA-co-HEA) con diferentes proporciones de EA y HEA para determinar la respuesta biológica de este tipo de células en función de la hidrofobicidad. Las MRC-5 no se infiltraron en los scaffolds durante la duración del cultivo. La adhesión de las MRC-5 a los diferentes scaffolds se evaluó con y sin recubrimiento de fibronectina. En ambos casos, las MRC-5 se adhirieron, propagaron, y proliferaron en los scaffolds de PEA y de P(EA- co-HEA) 90/10 hasta que las células llegaron a confluencia. El PEA indujo una tasa de crecimiento celular ligeramente superior al P(EA-co-HEA) 90/10, mientras que las células cultivadas sobre P(EA-co-HEA) 80/20 no alcanzaron la confluencia (Figura 3.23). La principal diferencia entre los substratos recubiertos con fibronectina y no recubiertos fue la tasa de proliferación celular, que pareció ligeramente mayor en el caso de los scaffolds recubiertos de fibronectina. La Figura 3.23 muestra varias imágenes de microscopía confocal de barrido donde se puede apreciar los microfilamentos de actina y los núcleos de las células MRC-5 en los scaffolds de PEA, P (EA-co-HEA) 90/10, y P(EA-co-HEA) 80/20 tras 8 días de cultivo sin recubrimiento con FN. 117 Los fibroblastos MRC-5 alcanzaron confluencia completa en los scaffolds y el recubrimiento de los substratos con fibronectina no fue necesario para la adhesión. Las MRC-5 se adhirieron a scaffolds con un contenido de componente hidrófilo de hasta un 20% (Figura 3.24). Claramente, cuanto más hidrófobo el scaffold, mejor la adherencia, difusión y proliferación celular. Este resultado está en consonancia con el que se encontró cuando la línea epitelial humana normal de las células, NHC, se sembró en polímeros en bloque con composición química idéntica [175]. Por encima de este umbral de hidrofilicidad del 20%, como en el caso de P(EA-co-HEA) 50/50 se observó que no existía adhesión celular. 118 Figura 3.23.- Imágenes de CLSM de F-actina y núcleo celular de MRC-5 teñidos con faloidina y Hoechst, respectivamente, tras 8 días de cultivo sin recubrimiento con FN. (a), (c), y (e) son PEA, P (EA-co-HEA) 90/10, y P(EA- co-HEA) 80/20, respectivamente, y (b) and (d) son los mismos materiales que (a) y (c), respectivamente, a mayores aumentos. Barrita para (a), (c), y (e): 150 micras. Barrita para (b) y(d): ): 37.5 micras x 100000 119 2 1,8 1,6 1,4 MRC-5/cm2 1,2 1 0,8 0,6 0,4 0,2 0 -10 0 10 20 30 40 50 60 %HEA Figura 3.24.- Núcleos por cm2 de MRC-5 en diferentes scaffolds sin recubrimiento de FN. Los valores se presentan como desviación media estadística a partir de dos réplicas independientes a los 8 días de la siembra 120 3.2.3.- Cultivo de células endoteliales, HUVEC Como se dijo anteriormente, las células endoteliales humanas de cordón umbilical necesitaron de pre-impregnación de fibronectina para la adhesión celular, por lo que es un sistema ideal para tratar de correlacionar la conformación de la proteína adsorbida con la adhesión y proliferación celular, como se muestra en la sección 3.3. Efecto del balance hidrófobo/hidrófilo Las células endoteliales primarias de cordón umbilical, HUVEC, fueron sembradas en scaffolds de P(EA-co-HEA) con diferentes proporciones de EA y HEA para determinar la respuesta biológica de estos tipos de células en función de la hidrofobicidad. En la ausencia de recubrimiento de fibronectina, pocas HUVEC permanecieron 4 h después de la adición a los scaffolds de P(EA-co-HEA) (datos no mostrados). Sin embargo, cuando los scaffolds de P(EA-co-HEA) se recubrieron primero con fibronectina, un número significativo de HUVEC se adhirieron (Figura 3.25). La excepción fueron los scaffolds de P(EA-coHEA) 50/50, en los que la adhesión celular no tuvo lugar, incluso después del recubrimiento con fibronectina (datos no mostrados). Los resultados que se muestran en la Figura 3.25 son representativos de al menos tres donantes de HUVEC diferentes. Con el aumento del tiempo de cultivo, las células sembradas en los scaffolds de P(EA-co-HEA) recubiertos con fibronectina comenzaron a proliferar, pero no alcanzaron confluencia en toda la superficie disponible. Dado que las células endoteliales sólo crecieron en los scaffolds de P(EA-co-HEA) recubiertos con fibronectina, todos los experimentos posteriores se llevaron a cabo con dicho recubrimiento. Las HUVEC no se infiltraron en los scaffolds durante la duración del cultivo. El test de Kruskal-Wallis de poblaciones celulares en diferentes tipos 121 de superficies reveló diferencias significativas en el grado de proliferación de HUVEC (k=10.018; p=0.007). Para dilucidar las diferencias en el crecimiento celular entre pares de grupos, se realizó el test U de MannWhitney. Los resultados mostraron diferencias entre el PEA y el P(EA -coHEA) 90/10 (U=2; p=0.017), y entre el PEA y el P(EA-co-HEA)80/20 (U=0; p=0.004), pero no se encontraron diferencias significativas entre P(EA-co-HEA) 90/10 y P(EA-co-HEA) 80/20 (U=9, p=0.180). 122 Figura 3.25.- Imágenes de CLSM de HUVEC adherentes teñidas con calceínaAM tras 5 días de siembra sobre los scaffolds, previamente recubiertos con FN. (a), (c), y (e) son PEA, P (EA-co-HEA) 90/10, y P(EA-co-HEA) 80/20, respectivamente, a 50x aumentos. (b), (d), y (f) son los mismos materiales que (a), (c), y (e), respectivamente, a 100x aumentos 123 Expresión de proteínas PECAM-1 y E-selectina por las HUVEC sobre los scaffolds A diferencia de los fibroblastos MRC-5, las HUVEC no alcanzaron confluencia completa en los scaffolds (Figura 3.25). Para probar la activación proinflamatoria de las células endoteliales, las HUVEC se cultivaron en los scaffolds de P(EA-co-HEA) recubiertos de FN y se realizó la inmunotinción para la E-selectina (rojo) y los núcleos celulares (azul) con Hoechst 33342 tras 4 horas de estimulación con LPS. Casi todas las HUVEC fueron Eselectina positivas después de la exposición a LPS, como se representa en la Figura 3.26a para el scaffold de PEA. Además, después de la inmunotinción contra moléculas de adhesión celular endotelial (PECAM-1, también conocida como CD31), las células cultivadas en los scaffolds de P(EA-coHEA) mostraron la característica típica de los contactos célula-célula de las HUVEC in vitro e in vivo (Figura 3.26b-c). 124 Figura 3.26.- Imágenes de inmunofluorescencia de (a) E-selectina y (b) PECAM1 de células endoteliales cultivadas sobre los scaffolds de PEA. (a) Las células endoteliales fueron fijadas y teñidas con un anticuerpo dirigido contra la Eselectina (rojo) y núcleos sobreteñidos con Hoechst (azul) tras de 4 h de estimulación con LPS. Prácticamente todas las HUVEC se tiñeron positivamente. Ampliación, 200x. (b, c) HUVEC teñidas con anticuerpo dirigido contra la PECAM-1 (verde) y núcleos sobreteñidos con yoduro de propidio (rojo). Las flechas blancas indican la localización de la PECAM-1 teñida en los contactos célula-célula (verde). (c) vista ampliada del rectángulo de (b). Ampliación de (b) 100x y (c) 200x Expresión génica de las HUVEC La regulación de la expresión génica de las células endoteliales a nivel de mRNA fue examinada por rtPCR para las HUVEC cultivadas en los scaffolds de P(EA-co-HEA). Como se muestra en la Figura 3.27, las células que crecen en los scaffolds de P(EA-co-HEA) recubiertos de fibronectina en ausencia de LPS (-) mostraron poca o ninguna expresión del gen para la molécula de adhesión celular E-selectina, un marcador característico de la inflamación. En presencia de LPS (+), se observó una fuerte inducción de la expresión de E-selectina en comparación con las células en ausencia de LPS (-). Por tanto, no sólo las células que crecen en los scaffolds de P(EA-coHEA) exhiben la expresión de genes endoteliales normales en ausencia de LPS, sino que estas células también mantienen las capacidades funcionales como se refleja en la fuerte inducción de un marcador inflamatorio en 125 presencia de LPS. Este comportamiento celular concuerda con la expresión de E-selectina a nivel proteico, ya que la mayoría de las HUVEC que crecieron sobre los scaffolds de P(EA-co-HEA) en presencia de LPS fueron positivas para E-selectina mediante inmunotinción (ver sección “Expresión de proteínas PECAM-1 y E-selectina por las HUVEC sobre los scaffolds”, Figura 3.26a). Por lo tanto, las células que crecen sobre los scaffolds de P(EA-co-HEA) exhibieron un fenotipo y un comportamiento celular muy similar a las células que crecen en las placas de cultivo de tejidos estándar [169]. Figura 3.27.- Análisis de PCR de la expresión génica de células endoteliales cultivadas en scaffolds precubiertos con fibronectina en presencia o ausencia de LPS. Se aisló el ARN de las células cultivadas durante 4 h con (+) y sin (-) LPS, se convirtió a cDNA, y se sometió a PCR como se describe en Materiales y Métodos. '' m'' es el marcador de tamaño de base molecular (bp) ,'' p'' es el control positivo, y'' n'' es el control negativo de agua sin cDNA Los substratos poliméricos usados en este estudio son materiales macroporosos con poros esféricos interconectados. Teniendo en cuenta las características físicas de estos scaffolds, esperábamos que las células sembradas en ellos encontraran nanodominios hidrófilos e hidrófobos alternos en la superficie del scaffold. La estructura química de las cadenas de 126 copolímero formado por EA y HEA se compone de una columna vertebral con grupos laterales -COOCH2CH2OH. La distribuidos modificación aleatoriamente de la -COOCH2CH3 relación EA/HEA en y los copolímeros usados para crear nuestros scaffolds, provoca que la densidad superficial de grupos-OH varíe y, en consecuencia, la hidrofobicidad de la superficie. La metodología empleada en este trabajo nos permitió caracterizar la interacción entre esta estructura macroporosa y las células endoteliales primarias humanas de cordón umbilical, HUVEC. El recubrimiento de los substratos con fibronectina fue imperativo para la fijación de las HUVEC. Al igual que en otros materiales sintéticos tales como poliestireno para cultivo tisular, el pre-recubrimiento con gelatina, laminina, fibronectina aumenta significativamente la adhesión de células endoteliales así como la proliferación [170]-[172]. El recubrimiento con fibronectina es necesario con frecuencia para la adhesión de las HUVEC en diferente materiales [169]. Las HUVEC se adhirieron a scaffolds con un contenido de componente hidrófilo de hasta un 20%. Pero, claramente, cuanto más hidrófobo el scaffold, mejor la adherencia, difusión y proliferación de celular. Por encima de este umbral de hidrofilicidad del 20%, como en el caso de P(EA-co-HEA) 50/50 se observó que no existía adhesión celular incluso con un prerecubrimiento de proteínas de adhesión sobre la superficies (Figura 3.28). Curiosamente, otros tipos de células tales como condrocitos, osteoblastos y células neuronales también se han cultivado en los substratos de polímero descritos aquí, pero no se observó la correlación de hidrofobicidad con la adhesión celular. En particular, las células neuronales se adhirieron mejor al P(EA-co-HEA) 50/50 [174]. 127 60% %Covered Surface 50% 40% 30% 20% 10% 0% -10 0 10 20 30 40 50 60 %HEA Figura 3.28.- Superficie cubierta por HUVEC en diferentes scaffolds. Los valores se presentan como desviación media estadística a partir de tres experimentos independientes a los 5 días de siembra. El test de Kruskal-Wallis muestra diferencias significativas entre la superficie cubierta por HUVEC en cada material (k=10.018; p = 0.007). El test de la U de Mann-Whitney mostró diferencias significativas entre PEA y P(EA-co-HEA) 90/10 (U=2; p=0.017), y entre PEA y P(EA-co-HEA) 80/20 (U=0; p=0.004), mientras que no se encontraron diferencias significativas entre P(EA-co-HEA) 90/10 y P(EA-co-HEA) 80/20 (U=9; p=0.180) Conclusiones: Como conclusión podemos decir que los substratos poliméricos empleados no presentan citotoxicidad para ninguno de los tipos celulares empleados. De cara al estudio de la conformación de proteínas, cabe decir que cuanto más hidrófobo es el material mejor es la adhesión celular. 128 3.3.- Conformación de la fibronectina en los materiales Esta sección está basada en el artículo publicado “Analysis of the biological response of endothelial and fibroblast cells cultured on synthetic scaffolds with various hydrophilic/hydrophobic ratios: influence of fibronectin adsorption and conformation. Campillo-Fernandez, A. J. et al. Tissue Engineering Part A, 1331-41 (2009)”. La Figura 3.29 muestra la magnitud de la fase de los diferentes sistemas de copolímero/fibronectina. La conformación de la fibronectina en los diferentes copolímeros en bloque está en función de la química del substrato: en particular, la densidad de grupos -OH en la superficie del copolímero. La fibronectina asume una conformación extendida en el homopolímero de PEA, el substrato más hidrófobo de la serie. La morfología de la fibronectina cambió a una morfología menos extendida con el aumento de hidrofilia de la superficie, generada por una mayor concentración de grupos-OH en la superficie del polímero. Es de destacar que la formación de una red de fibronectina tuvo lugar en el homopolímero de PEA, mientras que no hay tal red en los otros substratos de copolímero. Imágenes de la altura de la fibronectina en la superficie del homopolímero de PEA se muestran en la Figura 3.29a1, b1 y c1. Vale la pena mencionar que aunque la rugosidad del substrato es lo suficientemente pequeña para permitir la detección de las proteínas en la imagen de altura, el parámetro más adecuado para observar la fibronectina es la señal de fase [97]. 129 Figura 3.29.- Imágenes de fase de AFM de la fibronectina sobre los diferentes copolímeros a diferentes aumentos. (a-c) homopolímero PEA; (d-f) P (EA-coHEA) 90/10; (g-i) P(EA-co-HEA) 80/20; (j-l) P(EA-co-HEA) 50/50; a1-c1 representa imágenes de altura de la fibronectina en el homopolímero PEA. La primera columna muestra 5x5 µm, la segunda columna muestra 2x2 µm, y el tercero de 1x1 µm 130 Para obtener una mejor comprensión de la influencia de las proteínas sobre la adhesión celular, láminas planas de substratos de polímero con una composición química idéntica como los correspondientes scaffolds recubiertos con fibronectina se visualizaron mediante AFM. Sólo en el homopolímero de PEA se produjo la formación de una red de fibronectina, pero cada vez menos y menos conforme el aumento de grupos hidroxilo en la superficie. Esto no es lo que sucede con los copolímeros de P(EA-co-HEA) cuando están recubiertos con otras proteínas de la ECM, tales como la laminina, donde se producen la conformación más extendida de la proteína y el inicio de la formación de la red sólo más allá de una cierta concentración superficial de grupos –OH [97]. Sin embargo, el resultado biológico se correlacionó con medidas cualitativas de la conformación de la proteína en el substrato [174]. La adsorción de proteínas sobre una superficie está gobernada por la energía libre de Gibbs del sistema. La adsorción de proteínas tendrá lugar si la energía libre de Gibbs en el estado adsorbido es menor que la energía libre de Gibbs de las proteínas rodeado de moléculas de agua, es decir, cuando las proteínas no interaccionan con el substrato. Tanto la entropía y la entalpía del sistema determinan la adsorción y la conformación de las proteínas. Por lo tanto, materiales hidrófobos deben ser mejores substratos para la adhesión de proteínas debido a la liberación de moléculas de agua unidas a las proteínas, lo que conduce a un aumento en la entropía [89]. Por otro lado, los factores entálpicos tales como interacciones de van der Waals, enlaces iónicos y enlaces de hidrógeno entre las proteínas y la superficie del substrato juegan un papel importante en la energía libre de Gibbs. Ambos componentes de la energía libre de Gibbs definirán conjuntamente el estado de equilibrio, es decir, la cantidad de proteínas absorbidas y su conformación. Especialmente, la conformación de proteínas es un factor determinante en la adhesión celular. El tripéptido RGD es uno de los principales dominios de adhesión de la fibronectina [175] y parece que cuanto 131 más extendida se encuentra la fibronectina sobre una superficie, mejor la secuencia RGD se encuentra disponible para interactuar con receptores de la membrana celular. Los resultados de AFM de los scaffolds recubiertos de fibronectina son indicativos de la correlación entre la adherencia de HUVEC y la conformación de las moléculas de fibronectina previamente depositada sobre el substrato. La influencia de la estructura química del substrato sobre la adhesión celular se transmite a través de las interacciones proteínapolímero. Probablemente, los mismos principios aplican al caso de los fibroblastos, aunque el pre-recubrimiento de la superficie con la fibronectina no fue un factor decisivo como en el caso de HUVEC. Inicialmente, la adsorción de proteínas del medio que contiene suero y, más tarde, las proteínas de la ECM producidas por las células proporcionan sitios de adhesión a las células en un biomaterial sintético [89]. La vascularización del biomaterial es requisito para las células para obtener nutrientes y oxígeno para su supervivencia. Por lo tanto, en un eventual proceso de implantación del biomaterial, los vasos sanguíneos del huésped tienen que crecer hacia el biomaterial. Si un material se siembra con las propias células endoteliales de un paciente, y se proporcionan antes de la implantación condiciones adecuadas para preformar un sistema microvascular dentro del biomaterial, estos microvasos potencialmente podrían conectarse con el sistema vascular existente que rodea el implante después de la intervención. El objetivo principal de nuestro estudio fue evaluar si se mantuvieron fenotipos endoteliales normales y las funciones observadas in vivo también in vitro. Las HUVEC exhibieron contactos célula-célula normales y la expresión de la activación proinflamatoria del marcador de E -selectina a la estimulación específica. En resumen, los scaffolds de P(EA-co-HEA) no presentaron efectos negativos sobre la función de las células endoteliales y su fenotipo, y por lo tanto, podrían servir como matrices pre-vascularizadas para aplicaciones en la medicina regenerativa, tales como implantes de córnea. 132 En particular, hemos desarrollado estos biomateriales teniendo en mente un nuevo anillo de anclaje para una queratoprótesis. Por lo tanto, los fibroblastos estromales activados (es decir, los queratocitos) y las células endoteliales formando el sistema microvascular para proporcionar los nutrientes necesarios deben ser capaces de penetrar en el scaffold después de la implantación. Ya que los fibroblastos no necesitan revestimiento de fibronectina, una posibilidad sería sembrar fibroblastos autólogos en los scaffolds, y con el tiempo los factores de crecimiento y de la matriz extracelular producidos por estas células podrían reclutar la invasión de las células endoteliales en el material. En consecuencia, existirían condiciones en las que el sistema vascular del destinatario pudiera conectar rápidamente y proporcionar al implante los nutrientes necesarios para la supervivencia y la función adecuada. Conclusiones: Se puede observar que cuanto mayor es la hidrofobicidad del substrato más extendida se encuentra la proteína fibronectina, llegando a formarse una red en el caso del PEA, el substrato más hidrófobo. 133 Capítulo 4. Conclusiones 134 En esta Tesis se han alcanzado las siguientes conclusiones generales: 1.- Free radical copolymerization of ethyl acrylate, EA, hydroxyethylacrylate, HEA, and ethylene glycol dimethacrylate, EGDMA, (as crosslinker) produces a polymer network whose chains are composed of two types of blocks, on the one hand a copolymer P(EA-co-HEA) with higher HEA content than average and on the other hand blocks of PEA homopolymer. Something similar happens in the copolymerization of EA with methacrylic acid, MAAc. 2.- This particular structure is formed by the different reactivity ratios of the two monomers. 3.- In P(EA-co-MAAc) copolymers this block structure produces a phase separation with PEA aggregates at least of the order of tens of nanometers, as demonstrated by the presence of PEA main relaxation in the copolymer. By contrast in the dry copolymer P(EA-co-HEA) this phase separation does not occur, neither the main relaxation of a PEA blocks phase is observed, nor his presence by atomic force microscopy is detected. PEA homopolymer blocks exist but they are dissolved in the copolymer matrix. 4.- It is interesting to check how the immersion of copolymer P(EA-co-HEA) in water induces phase separation by hydrophobic interaction. The presence of water provokes PEA blocks clustering, that show a dynamic - mechanical main relaxation process and the presence of nanoscale aggregates is seen by atomic force microscopy. 5.- The three cell types studied adhere better to hydrophobic substrates. Epithelial cells, NHC, and fibroblasts do that using ligands which are present in the layer of proteins that adheres to the material from the culture medium containing fetal bovine serum. On the contrary endothelial cells, HUVEC, 135 need specifically a layer of fibronectin adsorbed on the substrate and that this protein forms a lattice of extended chains on the surface, as in the PEA substrate. 6.- The presence of hydrophilic groups disassemble the organization of the protein on the substrate, prevents the spontaneous fibronectin fibrillation and HUVEC adhesion drops rapidly when HEA content increases in the copolymer. Something similar happens with epithelial cells and fibroblasts. 7.- It is interesting to note the positive effect of a small amount of methacrylic acid in the culture of epithelial cells, which adhere and proliferate in monolayer even better than in polystyrene wells. 8.- The interaction between these cell types and the materials promises good in vivo performance in terms of cell proliferation, differentiation, and function 9.- HUVECs exhibited normal cell–cell contacts and expression of the proinflammatory activation marker E-selectin upon specific stimulation. Thus, P(EA-co-HEA) scaffolds had no negative effects on endothelial cell function and phenotype, and might thus serve as prevascularized matrices for applications in regenerative medicine. 136 Glosario AFM, Microscopía de fuerza atómica DMA, Análisis mecánico-dinámico DMEM, Dulbecco´s Modified Eagle Medium ECM, Matriz extracelular FN, Fibronectina PEA/EA, Poli(acrilato de etilo)/ acrilato de etilo PHEA/HEA, Poli(acrilato de hidroxietilo)/ acrilato de hidroxietilo PMMA, Poli(metacrilato de metilo PMAAc/MAAc, Poli(ácido metacrílico)/ ácido metacrílico PS, Poliestireno , ratio entre las longitudes antes y después de la deformación en el eje j j Tg, Temperatura de transición vítrea DSC, Calorimetría diferencial de barrido DRS, Espectroscopía de relajación dieléctrica G/ mG, Energía libre de Gibbs/incremento de mezcla de energía libre de Gibbs μ̂ i , potencial químico del componente i en la mezcla μi , potencial químico del componente i puro E’, Módulo de almacenamiento EGDMA, dimetacrilato de etilenglicol EWC, contenido de agua en equilibrio SEM, microscopía electrónica de barrido 137 Tα, temperatura del máximo en tan δ tan δ, tangente de pérdidas w, fracción másica Δcp, incremento de la capacidad calorífica γ , tensión superficial s γ d, γp ,componentes dispersiva y polar de la tensión superficial s , 138 Referencias [1] Chirila TV. An overview of the development of artificial corneas with porous skirts and the use of PHEMA for such an application. Biomaterials 2001; 22: 3311-3317 [2] Hicks C, Crawford G, Chirila TV, Wiffen S, Vijayasekaran S, Lou X, Fitton J, Maley M, Clayton A, Dalton P, Platten S, Ziegelaar B, Hong Y, Russo A, Constable I. Development and clinical assessment of an artificial cornea. Prog Retin Eye Res 2000; 19: 149-170 [3] Dalton PD, Woodfield T, Hutmacher DW. SnapShot: Polymer Scaffolds for Tissue Engineering. Biomaterials 2009; 30: 701-702 [4] Vallés-Lluch A, Arnal-Pastor M, Martínez-Ramos C, Vilariño-Feltrer G, Vikingsson L, Castells-Sala C, Semino CE, Monleón Pradas M. Combining self-assembling peptide gels with three-dimensional elastomer scaffolds. Acta Biomater 2013; 9: 9451-9460 [5] Martínez-Ramos C, Vallés-Lluch A, Verdugo JM, Ribelles JL, Barcia Albacar JA, Orts AB, Soria López JM, Pradas MM. Channeled scaffolds implanted in adult rat brain. J Biomed Mater Res A 2012; 100: 3276-3286 [6] Olmedilla MP, Lebourg M, Ivirico JE, Nebot I, Giralt NG, Ferrer GG, Soria J, Ribelles JG. In vitro 3D culture of human chondrocytes using modified epsilon-caprolactone scaffolds with varying hydrophilicity and porosity. J Biomater Appl 2012; 27:299-309 139 [7] Dias Veiga D, Costa Antunes J, García Gómez R, Mano JF, Gómez Ribelles JL, Soria JM. Three-Dimensional Scaffolds as a Model System for Neural and Endothelial ‘In Vitro’ Culture. J Biomater Appl 2011; 26: 293-310 [8] Vallés-Lluch A, Novella-Maestre E, Sancho-Tello M, Pradas MM, Ferrer GG, Batalla CC. Mimicking Natural Dentin Using Bioactive Nanohybrid Scaffolds for Dentinal Tissue Engineering. Tissue Eng Part A 2010; 16: 2783-2793 [9] Valles A, Novella E Sancho, M, Gallego G, Monleon M, Carda C. Bioactive nanohybrid scaffolds mimicking natural dentin xenotransplanted in immunodeficient mice. Biodental Engineering 2010: 213-217 [10] Lluch AV, Fernández AC, Ferrer GG, Pradas MM. Bioactive Scaffolds Mimicking Natural Dentin Structure. J Biomed Mater Res B Appl Biomater 2009; 90: 182-194 [11] Campillo-Fernández AJ, Unger RE, Peters K, Halstenberg S, Santos M, Salmerón Sánchez M, Meseguer Dueñas JM, Monleón Pradas M, Gómez Ribelles JL, Kirkpatrick CJ. Analysis of the Biological Response of Endothelial and Fibroblast Cells Cultured on Synthetic Scaffolds with Various Hydrophilic/Hydrophobic Ratios: Influence of Fibronectin Adsorption and Conformation. Tissue Eng Part A. 2009; 15:1331-1341 [12] Rodríguez Hernández JC, Serrano Aroca A, Gómez Ribelles JL, Pradas MM. Three-dimensional nanocomposite scaffolds with ordered cylindrical orthogonal pores. J Biomed Mater Res B Appl Biomater 2008; 84: 541-549 140 [13] Diego RB, Estellés JM, Sanz JA, García-Aznar JM, Sánchez MS. Polymer scaffolds with interconnected spherical pores and controlled architecture for tissue engineering: Fabrication, mechanical properties, and finite element modeling. J Biomed Mater Res B Appl Biomater 2007; 81: 448-55 [14] Vidaurre A, Cortazar IC, Ribelles JLG. Polymeric scaffolds with a double pore structure. J Non-Cryst Solids 2007; 353: 1095-1100 [15] Diego RB, Gómez Ribelles JL, Salmerón-Sánchez M. Pore collapse during the fabrication process of rubber-like polymer scaffolds. J Appl Polym Sci 2007; 104: 1475-1481 [16] Diego RB, Olmedilla MP, Aroca AS, Ribelles JL, Pradas MM, Ferrer GG, Sánchez MS. Acrylic scaffolds with interconnected spherical pores hydrophilicity for tissue engineering and controlled J Mater Sci Mater Med 2005; 16: 693-698 [17] Campillo Fernández AJ, Arredondo Zamudio A, Gallego Ferrer G, Monleón Pradas M, Carda Batalla MC. Polymeric scaffolds for guided dentine regeneration. Int J Artif Organs 2005; 28: 362 [18] Deplaine H, Lebourg M, Ripalda P, Vidaurre A, SanzRamos P, Mora G, Prósper F, Ochoa I, Doblaré M, Gómez Ribelles JL, Izal-Azcárate I, Gallego Ferrer G.. Biomimetic hydroxyapatite coating on pore walls improves osteointegration of poly(L-lactic acid) scaffolds. J Biomed Mater Res B Appl Biomater 2013;101:173-186 141 [19] Gamboa-Martínez TC, Ródenas Róchina J, Rico P, Lebourg M, Gómez-Ribelles JL, Salmerón-Sánchez M, Gallego G. Chondrocytes Cultured in an Adhesive Macroporous Scaffold Subjected to Stirred Flow Bioreactor Behave Like in Static Culture J. Biomater. Tissue Eng 2013; 3: 312-319 [20] Izal I, Aranda P, Sanz-Ramos P, Ripalda P, Mora G, Granero-Moltó F, Deplaine H, Gómez-Ribelles JL, Ferrer GG, Acosta V, Ochoa I, García-Aznar JM, Andreu EJ, Monleón-Pradas M, Doblaré M, Prósper F. Culture of human bone marrow-derived mesenchymal stem cells on of poly(L-lactic acid) scaffolds: potential application for the tissue engineering of cartilage. Knee Surg Sports Traumatol Arthrosc 2013;21: 1737-1750 [21] Lebourg M, Rochina JR, Sousa T, Mano J, Ribelles JL. Different hyaluronic acid morphology modulates primary articular chondrocyte behavior in hyaluronic acid-coated polycaprolactone scaffolds. J Biomed Mater Res A. 2013; 101: 518-27 [22] Panadero JA, Vikingsson L, Gomez Ribelles JL, Sencadas V, Lanceros-Mendez S. Fatigue prediction in fibrin polyepsilon-caprolactone macroporous scaffolds J Mech Behav Biomed Mater. 2013; 28: 55-61 [23] Ródenas-Rochina J, Ribelles JL, Lebourg M. Comparative study of PCL-HAp and PCL-bioglass composite scaffolds for bone tissue engineering. J Mater Sci Mater Med 2013; 24: 1293-308 142 [24] Acosta Santamaría V, Deplaine H, Mariggió D, VillanuevaMolines AR, García-Aznar JM, Gómez Ribelles JL, Doblaré M, Gallego Ferrer G, Ochoa I. Influence of the macro and micro-porous structure on the mechanical behavior of poly(L-lactic acid) scaffolds. J. Non-Cryst. Solids 2012; 23: 3141-3149 [25] Sencadas V, Correia DM, Ribeiro C, Moreira S, Botelho G, Gómez Ribelles JL, Lanceros-Mendez S. Physical-chemical properties of cross-linked chitosan electrospun fiber mats. Polym. Test 2012; 31: 1062-1069 [26] Arnal-Pastor M, Vallés-Lluch A, Keicher M, Pradas MM. Coating typologies and constrained swelling of hyaluronic acid gels within scaffold pores. J Colloid Interface Sci. 2011; 361: 361-369 [27] Gamboa-Martínez TC, Gómez Ribelles JL, Gallego Ferrer G. Fibrin coating on poly(L-lactide) scaffolds for tissue engineering. J Bioact Compat Pol 2011; 26: 464–477 [28] Cruz DM, Gomes M, Reis RL, Moratal D, SalmerónSánchez M, Ribelles JL, Mano JF. Differentiation of mesenchymal stem cells in chitosan scaffolds with double micro and macroporosity. J Biomed Mater Res A. 2010; 95: 1182-1193 [29] Antunes JC, Oliveira JM, Reis RL, Soria JM, GómezRibelles JL, Mano JF. Novel poly(L-lactic acid)/hyaluronic acid macroporous hybrid scaffolds: Characterization and assessment of cytotoxicity J Biomed Mater Res A. 2010; 94: 856-869 143 [30] Martinez-Diaz S, Garcia-Giralt N, Lebourg M, GómezTejedor JA, Vila G, Caceres E, Benito P, Pradas MM, Nogues X, Ribelles JL, Monllau JC. In Vivo Evaluation of 3-Dimensional Polycaprolactone Scaffolds for Cartilage Repair in Rabbits. Am J Sports Med 2010; 38: 509-519 [31] Lebourg M, Suay Antón J, Gomez Ribelles JL. Hybrid structure in PCL-HAp scaffold resulting from biomimetic apatite growth. J Mater Sci Mater Med 2010; 21: 33-44 [32] Ivirico JL, Salmerón-Sánchez M, Ribelles JL, Pradas MM, Soria JM, Gomes ME, Reis RL, Mano JF. Proliferation and Differentiation of Goat Bone Marrow Stromal Cells in 3D Scaffolds With Tunable Hydrophilicity. J Biomed Mater Res B Appl Biomater 2009; 91: 277-86 [33] García Cruz DM, Coutinho DF, Mano JF, Gómez Ribelles JL, Salmerón-Sánchez macroporous M. scaffolds caprolactone)/chitosan Physical based interactions on semi-interpenetrating in poly(3polymer networks. Polymer 2009; 50: 2058-2064 [34] Mano JF, Hungerford G, Ribelles JLG. Bioactive poly(Llactic acid)-chitosan hybrid scaffolds Mater Sci Eng C 2008; 28: 1356-1365 [35] Cruz DM, Ivirico JL, Gomes MM, Ribelles JL, Sánchez MS, Reis RL, Mano JF. Chitosan microparticles as injectable scaffolds for tissue engineering. J Tissue Eng Regen Med 2008; 2: 378-380 144 [36] Garcia-Giralt N, Izquierdo R, Nogués X, Perez-Olmedilla M, Benito P, Gómez-Ribelles JL, Checa MA, Suay J, Caceres E, Monllau JC. A porous PCL scaffold promotes the human chondrocytes redifferentiation and hyalinespecific extracellular matrix protein synthesis. J Biomed Mater Res A 2008; 85: 1082-1089 [37] Lebourg M, Sabater Serra R, Más Estellés J, Hernández Sánchez F, Gómez Ribelles JL, Suay Antón J. Biodegradable polycaprolactone scaffold with controlled porosity obtained by modified particle-leaching. J Mater Sci Mater Med 2008; 19: 2047-2053 [38] Más Estellés J, Vidaurre A, Meseguer Dueñas JM, Castilla Cortázar I. Physical characterization of polycaprolactone scaffolds J Mater Sci Mater Med 2008; 19: 189-195 [39] Zhang R, Ma PX. Processing of polymer scaffolds: phase separation Atala A, Lanza R (Eds.), Methods of Tissue Engineering, Academic Press, San Diego, CA 2001: 715– 724 [40] Chong W, Min W. Electrospun multifunctional tissue engineering scaffolds. Front Mater Sci 2014; 8: 3-19 [41] Stupp SI. Self-assembly and biomaterials. Nano Lett 2010; 10: 4783–4786 [42] Liu C, Xia Z, Czernuszka JT. Design and development of three-dimensional scaffolds for tissue engineering. Chem Eng Res Des 2007; 85: 1051-1064 145 [43] Butler MJ, Sefton MV. Poly(butyl methacrylate-co- methacrylic acid) tissue engineering scaffold with proangiogenic potential in vivo. J Biomed Mater Res A. 2007; 82: 265-273 [44] Shimko DA. Nauman EA. Development and characterization of a porous poly(methyl methacrylate) scaffold with controllable modulus and permeability. J Biomed Mater Res B Appl Biomater. 2007; 80: 360-369 [45] Perez Olmedilla M, Garcia-Giralt N, Monleón Pradas M, Ruiz PB, Gómez Ribelles JL, Cáceres Palou E, Monllau García JC. Response of human chondrocytes to a nonuniform distribution of hydrophilic domains on poly (ethyl acrylate-co-hydroxyethyl methacrylate) copolymers. Biomaterials 2006; 27: 1003-1012 [46] Brandrup J, Immergut EH. In polymer handbook. 3rd ed. New York: John Wiley & Sons; 1989. Chapter II [47] Mayo FR, Lewis FM. Copolymerization I. A basis for comparing the behavior of monomers in copolymerization; the copolymerization of styrene and methyl methacrylate. J Am Chem Soc 1944; 66: 1594–1601 [48] Alfrey T, Goldfinger G. The mechanism of copolymerization. J Chem Phys 1944; 12: 205–209 [49] Ward IM (1979) Mechanical properties of solid polymers. Wiley, New York [50] Mc Crum NG, Buckley CP, Bucknall CB (2001) Principles of polymer engineering. Oxford University Press, New York 146 [51] Gedde UW. Polymer physics. Chapman & Hall, London, 1995. p.: 82 [52] Gómez Ribelles JL, Vidaurre Garayo A, Cowie JMG, Ferguson R, Harris S, McEwen IJ. Polymer, 1998: 40: 183192 [53] Cowie JMG. Polymers: Chemistry and Physics of Modern Materials. Blackie & Son Ltd, Glasgow & London. Chapman & Hall, New York, 1991 [54] Gómez Ribelles JL, Meseguer Dueñas JM, Monleón Pradas M. Dielectric relaxations in poly(methyl acrylate), poly(ethyl acrylate) and poly(butyl acrylate). J Appl Polym Sci 1989; 38: 1145-1157, 1989 [55] Kremer F, Schonhals A, Luck W, Broadband Dielectric Spectroscopy, Springer-Verlag, 2002 [56] Gómez Ribelles JL, Meseguer Dueñas JM, Monleón Pradas M. Dielectric relaxations in poly(hydroxy ethyl acrylate): influence of the absorbed water. Polymer 1988; 29: 11241127 [57] Couchman PR. Compositional variation of glass-transition temperatures. 2. Application of the thermodynamic theory to compatible polymer blends. Macromolecules 1978; 11: 1156–1161 [58] Chandler D. Interfaces and the driving force of hydrophobic assembly. Nature 2005; 437: 640–647 [59] Debenedetti PG. In metastable liquids. Princeton, NJ: Princeton University Press; 1996 147 [60] Ben-Naim A. In hydrophobic interactions. New York: Plenum Press; 1980 [61] Grinberg NV, Dubovik AS, Grinberg VY, Kuznetsov DV, Makhaeva EE, Grosberg AY, Tanaka T. Studies of the thermal volume transition of poly(N-isopropylacrylamide) hydrogels by high-sensitivity differential scanning microcalorimetry. 1. Dynamic effects. Macromolecules 1999; 32: 1471–1475 [62] Tokuhiro T, Amiya T, Mamada A, Tanaka T. NMR study of poly(Nisopropylacrylamide) gels near phase transition. Macromolecules 1991; 24: 2936–2943 [63] Shibayama M, Mizutani S, Nomura S. Thermal properties of copolymer gels containing N-isopropylacrylamide. Macromolecules 1996; 29: 2019–2024 [64] Shibayama M, Morimoto M, Nomura S. Phase separation induced mechanical transition of poly(N- isopropylacrylamide)/water isochore gels. Macromolecules 1994; 27: 5060–5066 [65] Shibayama M, Suetoh Y, Nomura S. Structure relaxation of hydrophobically aggregated poly(N-isopropylacrylamide) in water. Macromolecules 1996; 29: 6966–6968 [66] Suetoh Y, Shibayama M. Effects of non-uniform solvation on thermal response in poly(N-isopropylacrylamide) gels. Polymer 2000; 41: 505–510 [67] Wu C. A comparison transition of linear between the ‘coil-to-globule’ chains and the ‘‘volume phase transition” of spherical microgels. Polymer 1998; 39: 4609– 4619 148 [68] Tsunashima Y, Kawanishi H, Horii F. Reorganization of dynamic selfassemblies of cellulose diacetate in solution: dynamical critical-like fluctuations in the lower critical solution temperature system. Biomacromolecules 2002; 3: 1276–1285 [69] Zhang YX, Da AH. A fluorocarbon-containing hydrophobically associating polymer. J Polym Sci Polym Part C. Polym Lett 1990; 28: 213–218 [70] Zhang YX, Da AH, Butler GB, Hogen-Esch TE. A fluorine-containing hydrophobically associating polymer I. Synthesis and acrylamide solution and properties of fluorine-containing copolymers of acrylates or methacrylates. J Polym Sci Part A 1992; 30: 1383–1391 [71] Xie X, Hogen-Esch TE. Copolymers dimethylacrylamide and of N,N2-(N- ethylperfluorooctanesulfonamido)ethyl acrylate in aqueous media and in bulk. Synthesis and properties. Macromolecules 1996; 29: 1734–1745 [72] Ng WK, Tam KC, Jenkins RD. Rheological properties of methacrylic acid/ethyl acrylate co-polymer: comparison between an unmodified and hydrophobically modified system. Polymer 2001; 42: 249–259 [73] Wang GJ, Engberts JBFN. Fluorescence probing of the formation of hydrophobic domains by cross-linked poly(alkylmethyldiallylammonium bromides) in aqueous solution. Recl Trav Chim Pays-Bas 1994; 113: 390–393 149 [74] Salmerón Sánchez M, Monleón Pradas M, Gómez Ribelles JL. Thermal transitions of benzene in copolymers and interpenetrating polymer networks based on hydrophilic and hydrophobic components. J Polym Sci Part B Polym Phys 2003; 41: 1713–1721 [75] Werner C, Pompe T, Katrin P. Modulating Extracellular Matrix at Interfaces of Polymeric Materials. Adv Polymer Sci 2006; 203: 63-93 [76] Tsapikouni T, Missirlis Y. Protein–material interactions: From micro-to-nano scale. Mat Sci Eng B. 2008; 152: 2-7 [77] Song W, Chen H. Protein adsorption on materials surfaces with nano-topography. Chin Sc. Bull 2007; 52: 3169-3173 [78] Sipe JD. Tissue engineering and reparative medicine. Ann NY Acad Sci 2002; 961: 1-9 [79] Schmidt DR, Waldeck H, Kao WJ. Protein Adsorption to Biomaterials. D.A. Puleo, R. Bizios, Editors. 2009, 1-18 [80] Salmerón-Sánchez M, Altankov G. Cell-Protein-Material interaction in tissue engineering. D. Eberli, Editor. 2010, In-Tech: 77-102 [81] Roach P, Eglin D, Rohde K, Perry CC. Modern biomaterials: a review – bulk properties and implications of surface modifications. J Mater Sci: Mater Med 2007; 18: 1263-1277 [82] Planell JA, Navarro M, Altankov G, Aparicio C, Engel E, Gil J, Ginebra MP, Lacroix D. Materials Surface Effect on Biological Interactions. Shastri VP, Altankov G, Lendlein A, Editors. 2010, Springer Netherlands: Dordrecht. 233-252 150 [83] Noh H, Vogler EA. Volumetric interpretation of protein adsorption: competition from mixtures and the Vroman effect. Biomaterials, 2007; 28: 405-422 [84] McKenzie JL, Webster TJ. Protein Interactions at Material Surfaces. R. Narayan, Editor. 2009, Springer US: Boston, MA. p. 215-237 [85] Lord MS, Foss M, Besenbacher F. Influence of nanoscale surface topography on protein adsorption and cellular response. Nano Today 2010; 5: 66-78 [86] Liu H, Webster TJ. Nanomedicine for implants: A review of studies and necessary experimental tools. Biomaterials 2007; 28: 354-369 [87] Grinnell F. Focal adhesion sites and the removal of substratum-bound fibronectin. J. Cell Biol 1986; 103: 26972706 [88] Griffith LG, Naughton G. Tissue engineering--current challenges and expanding opportunities. Science 2002; 295: 1009-1014 [89] García AJ. Interfaces to Control Cell-Biomaterial Adhesive Interactions. Adv Polym Sci 2006; 203: 171-190 [90] Chen H, Yuan L, Song W, Wu Z, Li D. Biocompatible polymer materials: Role of protein–surface interactions. Prog Polym Sci 2008; 33: 1059-1087 151 [91] Ballet T, Boulange L, Brechet Y, Bruckert F, Weidenhaupt M. Protein conformational changes induced by adsorption onto material surfaces : an important issue for biomedical applications of material science. Bull. Pol. Ac.: Tech 2010; 58: 303-315 [92] Anselme K, Ponche A, Bigerelle M. Relative influence of surface topography and surface chemistry on cell response to bone implant materials. Part 2: biological aspects. Proc. IMechE. Part H: J. Engineering in Medicine, 2010. 224, 1487-1507 [93] Hynes RO. Integrins: bidirectional, allosteric signaling machines. Cell, 2002; 110: 673-687 [94] Geiger B, Bershadsky A, Pankov R, Yamada KM. Transmembrane crosstalk between the extracellular matrix and the cytoskeleton. Nat Rev Mol Cell Bio 2001; 2: 793805 [95] García AJ. Get a grip: integrins in cell-biomaterial interactions. Biomaterials 2005; 26: 7525-7529 [96] Wong JY, Leach JB, Brown XQ. Balance of chemistry, topography, and mechanics at the cell–biomaterial interface: Issues and challenges for assessing the role of substrate mechanics on cell response. Surf Sci 2004; 570: 119- 133 152 [97] Rodríguez Hernández JC, Salmerón Sánchez M, Soria JM, Gómez Ribelles JL, chemistry-dependent Monleón Pradas conformations of M. Substrate single laminin molecules on polymer surfaces are revealed by the phase signal of atomic force microscopy. M. Biophys. J. 2007; 93: 202-207 [98] Rico P, Rodríguez Hernández JC, Moratal D, Altankov G, Monleón Pradas M, Salmerón-Sánchez M.. Substrate induced assembly of fibronectin into networks: influence of surface chemistry and effect on osteoblast adhesion. Tissue Eng Part A. 2009; 15: 3271-3281 [99] Rico P, González-García C, Petrie TA, García AJ, Salmerón-Sánchez M. Molecular assembly and biological activity of a recombinant fragment of fibronectin (FNIII(710)) on poly(ethyl acrylate). Colloid Surf B 2010; 78: 310316 [100] Gugutkov D, Altankov G, Rodríguez Hernández JC, M. Salmerón-Sánchez. Fibronectin activity on substrates with controlled --OH density. J Biomed Mater Res A 2010; 92: 322-331 [101] Brizuela Guerra N, González-García C, Llopis V, Rodríguez Hernández JC, Moratal D, Rico P, SalmerónSánchez M. Subtle variations in polymer chemistry modulate substrate stiffness and fibronectin activity. Soft Matter 2010; 6: 4748-4755 [102] Cristina González García’s thesis. Biological activity of fibronectina at the cell-material interface. Universitat Politècnica de València (Spain), 2012 153 [103] Laschke MW, Harder Y, Amon M, Martin I, Farhadi J, Ring A, Torio-Padron N, Schramm R, Rücker M, Junker D, Häufel JM, Carvalho C, Heberer M, Germann G, Vollmar B, Menger MD. Angiogenesis in tissue engineering: breathing life into constructed tissue substitutes. Tissue Eng. 2006; 12: 2093–2104 [104] Rouwkema J, Rivron NC, van Blitterswijk CA. Vascularization in tissue engineering. Trends Biotechnol. 2008; 26: 434–441 [105] Druecke D, Langer S, Lamme E, Pieper J, Ugarkovic M, Steinau HU, Homann HH. Neovascularization of poly(ether ester) blockcopolymer scaffolds in vivo: long-term investigations using intravital fluorescent microscopy. J Biomed Mater Res A 2004; 68: 10–18 [106] Yang S, Leong KF, Du Z, Chua CK. The design of scaffolds for use in tissue engineering. Part I. Traditional factors. Tissue Eng. 2001; 7: 679–689 [107] Karageorgiou V, Kaplan D. Porosity of 3D biomaterial scaffolds and osteogenesis. Biomaterials 2005; 26: 5474– 5491 [108] Hollister SJ. Porous scaffold design for tissue engineering. Nat. Mater. 2005; 4: 518–524 [109] Hutmacher DW, Sittinger M, Risbud MV. Scaffold-based tissue engineering: rationale for computer-aided design and solid free-form fabrication systems. Trends Biotechnol 2004; 22: 354–362 154 [110] Richardson TP, Peters MC, Ennett AB, Mooney DJ. Polymeric system for dual growth factor delivery. Nat. Biotechnol. 2001; 19: 1029–1034 [111] Hirschi KK, Skalak TC, Peirce SM, Little CD. Vascular assembly in natural and engineered tissues. Ann. N.Y. Acad. Sci. 2002; 961: 223–242 [112] Zisch AH, Lutolf MP, Hubbell JA. Biopolymeric delivery matrices for angiogenic growth factors. Cardiovasc Pathol 2003; 12: 295–310 [113] Carmeliet P. Mechanisms of angiogenesis and arteriogenesis. Nat Med 2000; 6: 389–395 [114] Chen RR, Silva EA, Yuen WW, Mooney DJ. Spatiotemporal VEGF and PDGF delivery patterns blood vessel formation and maturation. Pharm Res 2007; 24: 258–264 [115] Pola R, Ling LE, Silver M, Corbley MJ, Kearney M, Blake Pepinsky R, Shapiro R, Taylor FR, Baker DP, Asahara T, Isner JM. The morphogen Sonic hedgehog is an indirect angiogenic agent upregulating two families of angiogenic growth factors. Nat Med 2001; 7: 706–711 [116] Déry MA, Michaud MD, Richard DE. Hypoxia-inducible factor 1: regulation by hypoxic and non-hypoxic activators. Int. J. Biochem. Cell Biol. 2005; 37: 535–540 [117] Deckers MM, van Bezooijen RL, van der Horst G, Hoogendam J, van Der Bent C, Papapoulos SE, Löwik CW. Bone morphogenetic proteins stimulate angiogenesis through osteoblast-derived vascular endothelial growth factor A. Endocrinology 2002; 143: 1545–1553 155 [118] Kneser U, Polykandriotis E, Ohnolz J, Heidner K, Grabinger L, Euler S, Amann KU, Hess A, Brune K, Greil P, Stürzl M, Horch RE. Engineering of vascularized transplantable bone tissues: induction of axial vascularization in an osteoconductive matrix using an arteriovenous loop. Tissue Eng. 2006; 12: 1721–1731 [119] Black AF, Berthod F, L'heureux N, Germain L, Auger FA. In vitro reconstruction of a human capillarylike network in a tissue-engineered skin equivalent. FASEB J. 1998; 12: 1331–1340 [120] Tremblay PL, Hudon V, Berthod F, Germain L, Auger FA. Inosculation of tissue-engineered capillaries with the host’s vasculature in a reconstructed skin transplanted on mice. Am. J. Transplant. 2005; 5: 1002–1010 [121] Shepherd BR, Enis DR, Wang F, Suarez Y, Pober JS, Schechner JS. Vascularization and engraftment of a human skin substitute using circulating progenitor cell-derived endothelial cells. FASEB J. 2006; 20: 1739–1741 [122] Levenberg S, Rouwkema J, Macdonald M, Garfein ES, Kohane DS, Darland DC, Marini R, van Blitterswijk CA, Mulligan RC, D'Amore PA, Langer R. Engineering vascularized skeletal muscle tissue. Nat. Biotechnol. 2005; 23: 879–884 [123] Choong CS, Hutmacher DW, Triffitt JT. Co-culture of bone marrow fibroblasts and endothelial cells on modified polycaprolactone substrates for enhanced potentials in bone tissue engineering. Tissue Eng. 2006; 12: 2521–2531 156 [124] Rouwkema J, de Boer J, Van Blitterswijk CA. Endothelial cells assemble into a 3-dimensional prevascular network in a bone tissue engineering construct. Tissue Eng. 2006; 12: 2685–2693 [125] Unger RE, Sartoris A, Peters K, Motta A, Migliaresi C, Kunkel M, Bulnheim U, Rychly J, Kirkpatrick CJ. Tissuelike self-assembly in cocultures of endothelial cells and osteoblasts structures and on the formation three-dimensional of microcapillarylike porous biomaterials. Biomaterials 2007; 28: 3965–3976 [126] Fuchs S, Hofmann A, Kirkpatrick C. Microvessel-like structures from outgrowth endothelial cells from human peripheral blood in 2-dimensional and 3-dimensional cocultures with osteoblastic lineage cells. Tissue Eng. 2007; 13: 2577–2588 [127] Caspi O, Lesman A, Basevitch Y, Gepstein A, Arbel G, Habib IH, Gepstein L, Levenberg S. Tissue engineering of vascularized cardiac muscle from human embryonic stem cells. Circ. Res. 2007; 100: 263–272 [128] Kelm JM, Djonov V, Hoerstrup SP, Guenter CI, Ittner LM, Greve F, Hierlemann A, Sanchez-Bustamante CD, Perriard JC, Ehler E, Fussenegger M. Tissue-transplant fusion and vascularization of myocardial microtissues and macrotissues implanted into chicken embryos and rats. Tissue Eng. 2006; 12: 2541–2553 [129] West-Mays JA, Dwivedi DJ. The keratocyte: corneal stromal cell with variable repair phenotypes. Int J Biochem Cell Biol 2006; 38: 1625-1631 157 [130] Dalby MJ, Riehle MO, Sutherland DS, Agheli H, Curtis AS. Fibroblast response to a controlled nanoenvironment produced by colloidal lithography. J Biomed Mater Res A 2004; 69: 314-322 [131] Lambert CA, Nusgens BV, Lapiere ChM. Mechanosensing and mechano-reaction of soft connective tissue cells. Adv Space Res 1998; 21: 1081-1091 [132] Kühnel. Atlas Color de Citología e Histología (11ª ed). Ed. Médica. Panamericana, 2005 [133] Histology: a Text And Atlas; 6th ed; Ed. Wolters Kluver, 2011 [134] Atala A, Lanza R, Thomson JA, Nerem R. Principles of Regenerative Medicine. Ed. Elsevier, 2011 [135] Latkany R, Sheu MS, Loh IH, Trinkaus-Randall V. Plasma surface modification of artificial corneas for optimal epithelialization. J Biomed Mater Res 1997; 36: 29–37 [136] Aucoin L, Griffith CM, Pleizier G, Deslandes Y, Sheardown H. Interactions of corneal epithelial cells and surfaces modified with cell adhesion peptide combinations. J Biomater Sci Polym Ed 2002; 13: 447-462 [137] Jacob JT, Rochefort JR, Bi J, Gebhardt BM. Corneal epithelial cell growth over tethered-protein/peptide surfacemodified hydrogels. J Biomed Mater Res B Appl Biomater 2005; 72: 198-205 158 [138] Merrett K, Griffith CM, Deslandes Y, Pleizier G, Sheardown H. Adhesion of corneal epithelial cells to cell adhesion peptide modified pHEMA surfaces. J Biomater Sci Polym Ed 2001; 12: 647-671 [139] Kobayashi H, Ikada Y. Corneal cell adhesion and proliferation on hydrogel sheets bound with cell adhesive proteins. Curr. Eye Res., 1991; 10: 899-908 [140] Wallace C, Jacob JT, Stoltz A, Bi J, Bundy K. Corneal epithelial adhesion strength to tethered protein peptide modified hydrogel surfaces. J Biomed Mater Res A 2005; 72: 19-24 [141] Diebold Y, Calonge M, Enríquez de Salamanca A, Callejo S, Corrales RM, Sáez V, Siemasko KF, Stern ME. Characterization of a Spontaneously Immortalized Cell Line (IOBA-NHC) from Normal Human Conjunctiva. Invest Ophthalmol Vis Sci 2003; 44: 4263-4274 [142] Owens DK, Wendt RC. Estimation of the surface free energy of polymers. J Appl Polym Sci 1969; 13: 1741-1747 [143] Greenspan L, Humidity Fixed Points of Binary Saturated Aqueous Solutions. J. Res. Nat. Bur. Stand. (US), 1977; 81A: 89-96 [144] Jaffe EA, Nachman RL, Becker CG Minick C.R. Culture of human endothelial cells derived from umbilical veins: identification by morphologic and immunologic criteria. J Clin Invest 1973; 52: 2745-2756 159 [145] Peters K, Schmidt H, Unger RE, Otto M, Kamp G, Kirkpatrick CJ. Software-supported image quantification of angiogenesis in an in vitro culture system: application to studies of biocompatibility. Biomaterials 2002; 23: 34133419 [146] Unger RE, Wolf M, Peters K, Motta A, Migliaresi C, James Kirkpatrick C. Growth of human cells on a nonwoven silk fibroin net: a potential for use in tissue engineering. Biomaterials 2004; 25: 1069-1075 [147] Unger RE, Peters K, Wolf M, Motta A, Migliaresi C, Kirkpatrick CJ. Endothelialization of a non-woven silk fibroin net for use in tissue engineering: growth and gene regulation of human endothelial cells. Biomaterials 2004; 25: 5137-5146 [148] Flory PJ. Statistical Mechanics of Chain Molecules. C. Hanser Verlag, Munich, 1985 (orig.: J. Wiley, N.Y., 1969) [149] Tziampazis E, Kohn J, Moghe PV. PEG-variant biomaterials as selectively adhesive protein templates: model surfaces for controlled cell adhesion and migration. Biomaterials 2000; 21: 511-520 [150] Salmerón Sánchez M, Gallego Ferrer G, Monleón Pradas M, Gómez Ribelles JL. Influence of the hydrophobic phase on the thermal transitions of water sorbed in a polymer hydrogel based on interpenetration of a hydrophilic and a hydrophobic network. Macromolecules 2003; 36: 860–866 [151] Johari GP, Hallbrucker A, Mayer E. The glass–liquid transition of hyperquenched water. Nature 1987; 330: 552– 553 160 [152] Kwei TK. The effect of hydrogen-bonding on the glasstransition temperatures of polymer mixtures. J Polym Sci Polym Part C. Polym Lett 1984; 22: 307–313 [153] Kwei TK, Pearce EM, Pennacchia JR, Charton M. Correlation between the glass transition temperatures of polymer mixtures and intermolecular force parameters. Macromolecules 1987; 20: 1174–1176 [154] Krakovský I, Salmerón Sánchez M. Thermal transitions and structure of epoxy networks based on a,x-diamino terminated poly(propyleneoxide)-block-poly(ethyleneoxide)block-poly(propylene oxide) swollen in water. J Polym Sci Part B Polym Phys 2005; 43: 699–708 [155] Donth E. In relaxation and thermodynamics in polymersglass transition. Berlin: Akademie Verlag; 1992 [156] Salmerón Sánchez M, Monleón Pradas M, Gómez Ribelles JL. Thermal transitions of benzene in a poly(ethyl acrylate) network. J Non-Cryst Solids 2002; 307: 750–757 [157] Alves NM, Mano JF, Balaguer E, Meseguer Dueñas JM, Gómez Ribelles JL. Glass transition and structural relaxation in semi-crystalline poly(ethylene terephthalate): a DSC study. Polymer 2002; 43: 4111–4122 [158] Wang Y, Gómez Ribelles JL, Salmerón Sánchez M, Mano JF. Morphological contributions to glass transition in poly(L-lactic acid). Macromolecules 2005; 38: 4712–4718 161 [159] Monleón Pradas M, Gómez Ribelles JL, Serrano Aroca A, Gallego Ferrer G, Suay Antón J, Pissis P. Interaction between water and polymer chains in poly(hydroxyethyl acrylate) hydrogels. Colloid Polym Sci 2001; 279: 323–330 [160] Flory PJ. In principles of polymer chemistry. Ithaca, NY: Cornell University Press; 1966 [161] Siegfried DC, Thomas DA, Sperling LH. A reexamination of polystyrene/polystyrene homo interpenetrating polymer networks: aspects of relative network continuity and internetwork coupling. Macromolecules 1979; 12: 586–589 [162] Lydon, MJ, Minett TW, Tighe BJ. Cellular interactions with synthetic polymer surfaces in culture. Biomaterials 1985; 6: 396-402 [163] Horbett TA, Schway MB, Ratner BD. Hydrophilichydrophobic copolymers as cell substrates: Effect on 3T3 cell growth rates. J. Colloid. Interface Sci. 1985; 104: 28-39 [164] Kiremitçi M, Pulat M, Şenvar C, Şerbetçi Aİ, Pişkin E. Structural and cellular characterization of solvent-casted polyurethane membranes. Clin. Mater. 1990; 6: 227-237 [165] Rosen JJ, Schway MB. Kinetics of cell adhesion to a hydrophilic-hydrophobic copolymer model system Polym. Sci. Technol. 1980; 12: 667-686 [166] Horbett TA, Waldburger JJ, Ratner BD, Hoffman AS. Cell adhesion to a series of hydrophilic-hydrophobic copolymers studied with a spinning disc apparatus. J Biomed Mater Res 1988; 22: 383-404 162 [167] Schneider GB, English A, Abraham M, Zaharias R, Stanford C, Keller J. The effect of hydrogel charge density on cell attachment. Biomaterials 2004; 25: 3023-3028 [168] Soria JM, Martínez Ramos C, Bahamonde O, García Cruz DM, Salmerón Sánchez M, García Esparza MA, Casas C, Guzmán M, Navarro X, Gómez Ribelles JL, García Verdugo JM, Monleón Pradas M, Barcia JA. Influence of the substrate’s hydrophilicity on the in vitro Schwann cells viability. J Biomed Mater Res A 2007; 83: 463-470 [169] Unger RE, Peters K, Huang Q, Funk A, Paul D, Kirkpatrick CJ. Vascularization and gene regulation of human endothelial cells growing on porous polyethersulfone (PES) hollow fiber membranes. Biomaterials 2005; 26: 3461-3469 [170] Anderson JS, Price TM, Hanson SR, Harker LA. In vitro endothelialization of small-caliber vascular grafts. Surgery 1987; 101: 577-586 [171] Kaehler J, Zilla P, Fasol R, Deutsch M, Kadletz M. Precoating substrate and surface configuration determine adherence and spreading of seeded endothelial cells on polytetrafluoroethylene grafts. J Vasc Surg 1989; 9: 535-541 [172] Zilla P, Fasol R, Preiss P, Kadletz M, Deutsch M, Schima H, Tsangaris S, Groscurth P. Use of fibrin glue as a substrate for in vitro endothelialization of PTFE vascular grafts. Surgery 1989; 105, 515-522 163 [173] Campillo-Fernandez AJ, Pastor S, Abad-Collado M, Bataille L, Gomez-Ribelles JL, Meseguer-Dueñas JM, Monleon-Pradas M, Artola A, Alio JL, Ruiz-Moreno JM. Future design of a new keratoprosthesis. physical and biological analysis of polymeric substrates for epithelial cell growth. Biomacromolecules 2007; 8: 2429-2436 [174] Soria JM, Martínez Ramos C, Salmerón Sánchez M, Benavent V, Campillo Fernández A, Gómez Ribelles JL, García Verdugo JM, Pradas MM, Barcia JA. Survival and differentiation of embryonic neural explants on different biomaterials. J Biomed Mater Res A 2006: 79: 495-502 [175] Magnusson MK, Mosher DF. Fibronectin: structure, assembly and cardiovascular implications. Arterioscler Thromb Vasc Biol 1998; 18: 1363-1370 164 Publicaciones [1] Ivashchenko S, Escobar Ivirico JL, García Cruz DM, Campillo-Fernández AJ, Gallego Ferrer G, Monleón Pradas M. Bioactive organic–inorganic poly(CLMA-co-HEA)/silica nanocomposites. J Biomater Appl. In press. [2] Seras-Franzoso J, Steurer C, Roldán M, Vendrell M, Vidaurre-Agut C, Tarruella A, Saldaña L, Vilaboa N, Parera M, Elizondo E, Ratera I, Ventosa N, Veciana J, Campillo-Fernández AJ, García-Fruitós E, Vázquez E, Villaverde A. Functionalization of 3D scaffolds with protein-releasing biomaterials for intracellular delivery. J Control Release 2013; 171: 63-72 [3] Campillo-Fernández AJ, Unger RE, Peters K, Halstenberg S, Santos M, Salmerón Sánchez M, Meseguer Dueñas JM, Monleón Pradas M, Gómez Ribelles JL, Kirkpatrick CJ. Analysis of the Biological Response of Endothelial and Fibroblast Cells Cultured on Synthetic Scaffolds with Various Hydrophilic/Hydrophobic Ratios: Influence of Fibronectin Adsorption and Conformation. Tissue Eng Part A. 2009; 15:1331-1341 [4] Lluch AV, Fernández AC, Ferrer GG, Pradas MM. Bioactive Scaffolds Mimicking Natural Dentin Structure. J Biomed Mater Res B Appl Biomater 2009; 90: 182-194 [5] Campillo-Fernández AJ, Salmerón Sánchez M, Sabater i Serra R, Meseguer Dueñas JM, Monleón Pradas M, Gómez Ribelles JL. Water-induced (nano)organization in poly(ethyl acrylate-co-hydroxyethyl acrylate) networks. Eur Polym J 2008; 44; 1996-2004 165 [6] Campillo-Fernandez AJ, Pastor S, Abad-Collado M, Bataille L, Gomez-Ribelles JL, Meseguer-Dueñas JM, Monleon-Pradas M, Artola A, Alio JL, Ruiz-Moreno JM. Future design of a new keratoprosthesis. physical and biological analysis of polymeric substrates for epithelial cell growth. Biomacromolecules 2007; 8: 2429-2436 [7] Soria JM, Martínez Ramos C, Salmerón Sánchez M, Benavent V, Campillo Fernández A, Gómez Ribelles JL, García Verdugo JM, Pradas MM, Barcia JA. Survival and differentiation of embryonic neural explants on different biomaterials. J Biomed Mater Res A 2006: 79: 495-502 [8] Campillo Fernández AJ, Arredondo Zamudio A, Gallego Ferrer G, Monleón Pradas M, Carda Batalla MC. Polymeric scaffolds for guided dentine regeneration. Int J Artif Organs 2005; 28: 362 [9] Serrano Aroca A, Campillo Fernández AJ, Gómez Ribelles JL, Monleón Pradas M, Gallego Ferrer G, Pissis P. Porous poly(2-hydroxyethyl acrylate) hydrogels prepared by radical polymerization with methanol as diluents. Polymer 2004; 45: 8949-8955.