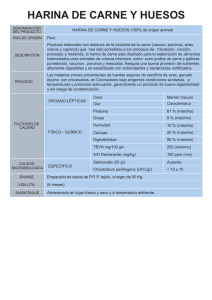
EVALUACIÓN DE LA CALIDAD NUTRICIONAL, CONSUMO Y
Anuncio

EVALUACIÓN DE LA CALIDAD NUTRICIONAL, CONSUMO Y DIGESTIBILIDAD IN VIVO DE LOS HENOS DE PASTO VIDAL (Bothriochloa saccharoides), COLOSOANA (Bothriochloa pertusa) Y ANGLETON CLIMACUNA (Dichanthium annulatum) JAIRO ANDRES PARDO GUZMAN MARY YURIANA QUINTERO PUENTES Trabajo de grado presentado como requisito parcial para optar al título de Médico Veterinario Zootecnista Director JAIRO RICARDO MORA DELGADO Ph.D. En Sistemas de Producción Agrícola Tropical Sostenible Codirector: ROBERTO PIÑEROS VARÓN M.Sc. En Ciencias Pecuarias UNIVERSIDAD DEL TOLIMA FACULTAD DE MEDICINA VETERINARIA Y ZOOTECNIA MEDICINA VETERINARIA Y ZOOTECNIA IBAGUE – TOLIMA 2014 DEDICADO A: A nuestras familias por su apoyo incondicional y a todas las personas que contribuyeron en la realización de este proyecto. 3 AGRADECIMIENTOS: A Roberto Piñeros Varón por su guía, enseñanzas, dedicación y paciencia en desarrollo de este trabajo de investigación. Al Doctor Jairo Ricardo Mora Delgado, por su orientación y apoyo incondicional durante el desarrollo de esta tesis. Al Centro Universitario Regional del Norte del Tolima (CURDN) de la Universidad del Tolima y a su director en el momento, Rodrigo Serrano por la colaboración prestada en el transcurso de la investigación. Al Doctor Héctor Vidal Perdomo por el respaldo y aporte en la consecución del pasto Vidal. A Román Castañeda por su apoyo y colaboración en todo momento. Al Comité Central de Investigaciones de la Universidad del Tolima. 4 CONTENIDO INTRODUCCIÓN 10 1. REFERENTES CONCEPTUALES 11 1.1 DIGESTIÓN DE LOS DIFERENTES NUTRIENTES EN RUMIANTES 11 1.1.1 Digestión de carbohidratos 11 1.1.2 Digestión de proteínas en el rumen 11 1.1.3 Digestión de Lípidos 12 1.2 DIGESTIBILIDAD 13 1.2.1 Factores que afectan la digestibilidad 14 1.3 MÉTODOS DE DETERMINACIÓN DE LA DIGESTIBILIDAD 15 1.3.1 Digestibilidad in vivo 15 1.3.2 Método in vitro 16 1.3.3 Método in situ 16 1.4 GRAMÍNEAS DEL TRÓPICO SECO 17 1.4.1 Pasto Vidal (Bothriochloa saccharoides) 17 1.4.2 Colosoana (Bothriochloa pertusa) 19 1.4.3 Angleton Climacuna (Dichanthium annulatum) - Angleton mono (Dichanthium aristatum) 20 1.5 CONSUMO VOLUNTARIO 21 1.5.1 Factores que afectan el consumo 23 2. OBJETIVOS 24 2.1 OBJETIVO GENERAL 2.2 OBJETIVOS ESPECÍFICOS 3. METODOLOGÍA 25 3.1 TRATAMIENTOS EVALUADOS 25 5 3.2 DIGESTIBILIDAD POR COLECTA TOTAL DE HECES 26 3.2.1 Periodo de adaptación 26 3.2.2 Periodo en las jaulas 26 3.2.3 Muestreo 26 3.3 CONSUMO DE MATERIA SECA 27 3.4 ANÁLISIS BROMATOLÓGICOS 27 3.5 FRACCIONAMIENTO PROTEICO 28 3.6 METABOLITOS SECUNDARIOS 29 3.7 MODELO ESTADÍSTICO 29 4. RESULTADOS Y DISCUSIÓN 30 4.1 BROMATOLÓGICOS 30 4.1.1 Componentes estructurales 30 4.1.2 Proteína cruda 31 4.1.2.1 Fraccionamiento Nitrogenado 32 4.1.3 Extracto Etéreo 33 4.2 CONSUMO VOLUNTARIO 34 4.3 METABOLITOS SECUNDARIOS 36 4.4 DIGESTIBILIDAD APARENTE 38 4.4.1 Digestibilidad de la MS y MO 38 4.4.2 Digestibilidad aparente de la proteína 40 4.4.3 Digestibilidad aparente de la FDN 42 5. CONCLUSIONES 43 RECOMENDACIONES 45 REFERENCIAS 47 6 LISTA DE TABLAS Tabla 1. Composición química y digestibilidad de la MS En diferentes etapas del ciclo fenológico del Bothriochloa saccharoides. Tabla 2. Análisis bromatológico del pasto B. pertusa en tres tiempos diferentes de corte. Tabla 3. Distribución de los ovinos en los tratamientos y periodos experimentales. Tabla 4. Composición Bromatológica del Bothriochloa saccharoides, Bothriochloa pertusa y Dichanthium annulatum. Tabla 5. Fracciones nitrogenadas, con respecto al porcentaje total de la proteína de cada una de las gramíneas. Tabla 6. Consumo de MS con respecto al peso vivo y el peso metabólico de las de las diferentes gramíneas evaluadas. Tabla 7. Metabolitos secundarios de las tres especies de Bothriochloa saccharoides, Bothriochloa pertusa y Dichanthium annulatum. Tabla 8. Digestibilidad aparente (DA) y nutrientes digestibles totales (NDT) de henos de Pasto Vidal (Bothriochloa saccharoides), Colosoana (Bothriochloa pertusa) y Angleton Climacuna (Dichanthium annulatum). Tabla 9. Digestibilidad de las fracciones nitrogenadas B3 y C en las gramíneas B. saccharoides, D. annulatum y B. pertusa. 7 RESUMEN Las producciones ganaderas de América Tropical se basan en el suministro de forrajes de baja calidad nutricional que son convertidos en alimentos de alta calidad proteica como carne y leche. El objetivo de este estudio fue determinar la calidad nutricional, el consumo y la digestibilidad aparente de tres gramíneas henificadas (B. saccharoides, B. pertusa, D. annulatum). El experimento se desarrolló en el Centro Universitario Regional del Norte del Tolima en el municipio de Armero Guayabal Tolima, a una altura de 275 m.s.n.m; temperatura de 26 °C y precipitación anual de 1732 mm. El diseño experimental fue Cuadrado Latino 3*3. Para la determinación de la digestibilidad aparente se utilizó el método in vivo colecta total de heces; se emplearon 12 ovinos machos mestizos que fueron alojados en jaulas metabólicas. De igual forma se empleó esta fase para realizar mediciones de consumo. Las muestras de gramíneas y heces recolectadas se evaluaron en el laboratorio de Ecofisiología de la Universidad del Tolima y la Universidad de Nariño. Los datos se analizaron a partir del software Infostat®, realizando ANOVA y Tukey (p= 0.05). El resultado del análisis bromatológico, fue similar para las tres gramíneas incluyendo los metabolitos secundarios; se observaron datos de digestibilidad superiores para B. saccharoides seguido por el B. Pertusa y por último D. annulatum. Sin embargo los pastos B. pertusa y D. annulatum presentaron mejor consumo con respecto a B. saccharoides. Palabras claves: Consumo, Digestibilidad, Pasto vidal, Colosoana, Angleton climacuna 8 ABSTRACT Livestock production in tropical America are based on the supply of fibrous foods that are converted to high quality food protein as meat and milk, which is why studies seeking to confirm an improvement in digestibility and intake of grasses ensuring greater conversion nutritional management based on a more accurate. The present study aimed to determine the nutritional quality, consumption and apparent digestibility three grasses (B. saccharoides, B. pertusa, D. annulatum) the dry tropics of northern Tolima. The pilot phase was developed in the CURDN center in Armero-Tolima, (South America or Colombia) at 275 meters below the sea level temperature of 26 ° C and annual rainfall of 1732 mm. The experimental design was a Latin Square 3 * 3, with three treatments and three experimental periods was used. For determining the in vivo digestibility was used method through total feces collection; 12 crossbreed male sheep were housed 14 days to adaptation and 3 days to the experimental period in metabolic cages. Collection and weighing of food and feces declined in each of the experimental periods cages were used. Likewise this phase is used to perform power measurements, weighing the quantity of feed and 24 hours after rejections. Samples collected grasses and feces were evaluated in the laboratory of Ecophysiology of Tolima University and the University of Nariño. Data were analyzed from Infostat® software, performing Anova and Tukey (p = 0.05). The nutritional analysis was similar for the three grasses including secondary metabolites; higher digestibility data B. saccharoides followed by B. pertusa and finally D. annulatum observed. However pertusa pastures B. and D. annulatum had better consumption relative to B. saccharoides. Keywords : Consumption , Digestibility, Vidal Pasto, Colosoana , Angleton climacuna 9 INTRODUCCIÓN Actualmente se busca establecer especies forrajeras que se adapten a las condiciones climáticas del trópico, sean de fácil obtención, buena digestibilidad y que contribuyan con un gran aporte nutricional a los rumiantes. El pasto Vidal (Bothriochloa saccharoides) no es muy conocido en nuestro medio, pero recientemente se han realizado algunos estudios que han permitido conocer un poco más sobre sus bondades, determinando características fenotípicas y tolerancias a las sombras, demostrando que puede ser una muy buena opción forrajera para los animales del trópico. En este estudio se busca evaluar la calidad nutricional, el consumo y la digestibilidad del pasto Vidal (Bothriochloa pertusa), Colosoana (Bothriochloa pertusa) y Climacuna (Dichanthium annulatum) que con el paso del tiempo han colonizado en gran medida las praderas del trópico seco. 10 1. REFERENTES CONCEPTUALES La mayoría de los compuestos orgánicos de los alimentos se encuentran en forma de grandes moléculas insolubles, que deben degradarse en compuestos más sencillos, para así poder absorberse en la mucosa del tracto digestivo e integrarse a la sangre o a la linfa. El proceso de degradación, se denomina digestión (McDonald et al., 2006), en los rumiantes la base de su digestión son los microorganismos que habitan el rumen y su importancia radica en la degradación de fibras provenientes de diferentes forrajes (Brautigan, 2007). Según Relling y Mattioli, (2003) el estómago de los rumiantes está formado por cuatro divertículos, con el mismo origen embrionario. El rumen es el primero y más importante donde ocurre el proceso de fermentación a partir de los microorganismos, el retículo es un compartimiento más pequeño que retiene partículas grandes y comparte la función del rumen, el Omaso el cual filtra y absorbe agua de los alimentos, por último el abomaso que se encarga de la secreción de Ácido Clorhídrico (HCl) y otros zimógenos para el desdoblamiento de las proteínas (McDonald et al., 2006). 1.1 DIGESTIÓN DE LOS DIFERENTES NUTRIENTES EN RUMIANTES. 1.1.1 Digestión de carbohidratos. el desdoblamiento de los carbohidratos estructurales y no estructurales en el rumen se da por acción de las enzimas de los microorganismos que habitan este órgano, los cuales convierten estos compuestos en AGV (ácidos grasos volátiles) acético, propiónico y butírico, principalmente para ser absorbidos y utilizados como fuente de energía en el rumiante (Estrada, 2002). 1.1.2 Digestión de proteínas en el rumen. el nitrógeno en los alimentos puede dividirse en dos grupos principales: nitrógeno proteico (NP) y nitrógeno no proteico (NNP), obviando los ácidos nucleicos y otras formas de NNP (Estrada, 2002); pero también se pueden clasificar según (Shimada, 2005) en cuatro grupos: NNP soluble, proteína de rápida 11 degradación, proteína de lenta degradación y proteína no degradable; en los forrajes el contenido de ácidos nucleicos es insignificante, pero los productos fermentados ricos en microorganismos, pueden contener cantidades apreciables de estos (Estrada, 2002). La degradación proteica en el medio ruminal se inicia básicamente con la acción de las enzimas extracelulares de origen bacteriano y la fagocitosis que llevan a cabo los protozoos (Shimada, 2005); los microorganismos hidrolizan, las proteínas, los péptidos y los aminoácidos, algunos de los cuales posteriormente se degradan en ácidos orgánicos, amoniaco y dióxido de carbono (CO2) (Ramírez, 2005). Las bacterias, los protozoarios y hongos separan el grupo amino del aminoácido y lo liberan al medio ruminal como un producto de desecho, y emplean la cadena carbonada para obtener energía como si se tratara de un hidrato de carbono. Por otro lado, los grupos amino (-NH2) libres se convierten, en amoniaco (NH3) y luego en amonio (NH4) (Wattiaux, 2014; Relling & Mattioli, 2003 ). Pero así como los microorganismos cubren una parte de sus requerimientos energéticos desdoblando aminoácidos, las bacterias y los hongos pueden resintetizarlos siguiendo el camino inverso, uniendo cadenas carbonadas, proveniente especialmente de los hidratos de carbono, con grupos amino provenientes NH4 o desde otra fuente de nitrógeno no proteico NNP. los microorganismos salen del rumen, y su desdoblamiento y absorción se da en el estómago e intestino delgado aportando así los requerimientos proteicos que el animal necesita (Relling & Mattioli, 2003). 1.1.3 Digestión de Lípidos. Los lípidos se encuentran normalmente en bajas cantidades en los alimentos de origen vegetal. Los forrajes frescos poseen lípidos celulares (fosfolípidos, glucolipidos) y de superficie (ceras y cutina) (Relling & Mattioli, 2003). Garton et al. (1961) y Dawson et al. (1974) aseguran que los galactolipidos son los más abundantes en el forraje de los rumiantes (Citados por Vargas, 2012), contienen gran cantidad de ácidos grasos poliinsaturados (linoleico y linolénico), estos se hidrolizan al igual que los fosfolípidos en un alto grado por acción de las lipasa bacterianas. Una vez liberados, el glicerol puede ser convertido a un AGV y los ácidos grasos son hidrogenados 12 hasta ser convertidos en ácido esteárico (McDonald et al., 2006). A nivel de producción animal, es de gran interés, efectuar mediciones que indiquen o se aproximen al verdadero aprovechamiento de los nutrientes de un alimento, a través del tubo digestivo; a lo cual se le denomina digestibilidad. 1.2 DIGESTIBILIDAD. Es definida como la fracción de un alimento suministrado que no es excretado en las heces, es decir, la cantidad que se pierde o se digiere a través de su recorrido en el tracto gastrointestinal del animal y su estimación se da comúnmente en relación con la materia seca, coeficiente o en porcentaje, de tal forma se considera que este proceso de absorción influye en la valoración nutricional de los alimentos (Church, 1997; Shimada, 2005; McDonald et al., 2006). Según Duarte, (2011) la digestibilidad se halla principalmente con el fin de hacer análisis comparativos de los alimentos, las dietas o ingredientes que hacen parte de éstas. La determinación se realiza a través de diferentes métodos, uno de ellos es la colecta total de heces que tiene un mayor control del alimento ingerido y las excretas, tornándose dispendioso y complejo. Särkijärvi et al. (2012) señalan que para obtener la digestibilidad de la materia orgánica de los forrajes, la colecta total de heces es exacta, sin embargo tiene algunos inconvenientes al emplearlo en animales grandes como los caballos. Otros autores consideran que el estudio con animales in vivo es la prueba predilecta para comparar con otras técnicas que buscan hallar la digestibilidad de los alimentos ya que proporciona datos más precisos, sin embargo presenta desventajas al requerir bastante tiempo, personal capacitado, mayores recursos económicos y análisis químicos (Flores et al., 2003; Church., 1997). Entre las técnicas indirectas se encuentra las relaciones empíricas entre la composición química del forraje y digestibilidad, digestión in vitro e in situ de forrajes y la estimación por el método de espectroscopia de reflactancia en el infrarrojo cercano (NIRS) (Flores et al., 2003). Valencia et al. (2010) determinaron la digestibilidad in vivo en ovinos utilizando dietas a base de forrajes tropicales y establecieron la importancia de conocer la digestibilidad y las 13 propiedades nutricionales de los diferentes alimentos, afirmando que los nutrientes que posee la planta no son el total de nutrientes asimilables dentro del proceso de digestión en los rumiantes, considerando de esta forma retomar las decisiones en la utilización de cultivos y forrajes que no son comunes y que se pueden incluir en las dietas de los animales. Conocer la digestibilidad de un alimento es un objetivo primordial para el desarrollo de dietas balanceadas y su respectivo análisis de calidad. Una dieta con mayor digestibilidad, aporta más nutrimentos que los proporcionados por una dieta de baja digestibilidad (Särkijärvi et al., 2012). 1.2.1 Factores que afectan la digestibilidad. La digestibilidad de un alimento puede ser afectada por factores propios de éste como lo son sus componentes nutricionales o por aspectos relacionados con los animales que lo consumen (Church, 1993; Shymada, 2003). Hay forrajes con gran variabilidad en su digestibilidad y esto se debe en gran medida a su estado de madurez, entre más maduro este el forraje tendrá menor contenido de proteína y azúcares con mayor contenido de fibra (celulosa y lignina), lo que puede disminuir progresivamente su digestibilidad. La presentación de los alimentos también influyen en la digestibilidad, cuando estos se ven expuestos a procesos tales como el molido, pastillas u hojuela se aumenta la velocidad de pasaje por el tubo digestivo, reduciendo el tiempo de digestión, aumentando el consumo y mejorando la respuesta animal. Es importante mencionar que la digestibilidad varía entre especies así es como los rumiantes tienen mayor capacidad para digerir alimentos fibrosos con bajo contenido de proteína a diferencia de los monogástricos; sin embargo entre animales de una misma especies también existen cambios gracias a los hábitos y requerimientos de cada etapa productiva (Shymada, 2003). Changwony et al. (2014) evaluaron el consumo, la digestibilidad y la calidad de diferentes gramíneas a través de una cronosecuencia de años (15, 20, 25, 30) de pastoreo con ovinos, allí se evidenció que no hubo cambios significativos en el consumo, pero si en la digestibilidad de la materia seca (MS) que disminuyó con el pasar de 20 años de uso de 14 la tierra, debido en gran parte a que las fracciones de fibra FDN y FDA fueron menores y aumentó la cantidad de componentes indigeribles como la lignina (LDA). Särkijärvi et al. (2012) realizaron un estudio evaluando la calidad de dos especies de gramíneas con tres periodos de corte diferente, utilizando los métodos de digestibilidad in vivo e in vitro pepsina-celulasa en ovejas, en éste se comprobó que las diferentes especies de gramíneas y el tiempo de corte de éstas si intervienen en la digestibilidad, de modo que al retrasar el corte o la cosecha de las gramíneas estas se hacían menos digestibles por el aumento de lignificación de las paredes celulares, dando como resultado las diferencias de digestibilidad de la (MO) en las dos especies forrajeras. Otros argumentos sobre el efecto de la madurez en la digestibilidad tienen en cuenta la relación hoja-tallo, pero en este trabajo no hace referencia esta explicación ya que el forraje que tuvo mayor digestibilidad fue la que tenía menos hojas. Entre algunas dificultades que se presentan en la comparación de calidad de diferentes gramíneas, se encuentran la alta variedad de especies, que se realizan en diferentes etapas de crecimiento, las condiciones agroclimáticas y las distintas metodologías utilizadas en los laboratorios (Särkijärvi et al., 2012). La presencia de metabolitos secundarios como los compuestos fenólicos entre ellos la lignina mencionada anteriormente y los taninos, pueden afectar la digestibilidad de las gramíneas, debido a que estos no son normalmente absorbidos en el tracto gastrointestinal, permaneciendo en estado libre o formando complejos en el rumen, que interfieren en la digestibilidad de las proteínas y las paredes celulares de las plantas a través de los grupos inhibidores de la celulasas (McLeod, 1974; Meirelles et al., 2005). 1.3 MÉTODOS DE DETERMINACIÓN DE LA DIGESTIBILIDAD 1.3.1 Digestibilidad in vivo. Navarathne (1990) afirma que es una técnica empleada desde el siglo XIX para determinar la digestibilidad en rumiantes y monogástricos, basada comúnmente en el método de colecta total de heces (Citado por Ruiz, 2011). Según lo expuesto por Bondi, (1989), la colecta total de heces (CTH), involucra la medición de la 15 ingestión del alimento por parte de los animales y su excreción en un periodo de tiempo determinado. Muestras del forraje ofrecido y rechazado, de orina y heces son analizadas en laboratorio, para evaluar el balance de nutrientes ingeridos y excretados, como base de la determinación de la digestibilidad de los nutrientes en estudio. Esta es normalmente representada por un coeficiente de digestibilidad aparente (CDA), expresado en forma porcentual que se calcula mediante la ecuación descrita por Bondi, (1989) 𝐶𝐶𝐶𝐶𝐶𝐶 (%) = 𝑐𝑐𝑐𝑐𝑐𝑐𝑐𝑐𝑐𝑐𝑐𝑐𝑐𝑐 − 𝑒𝑒𝑒𝑒𝑒𝑒𝑒𝑒𝑒𝑒𝑒𝑒𝑒𝑒ó𝑛𝑛 𝑓𝑓𝑓𝑓𝑓𝑓𝑓𝑓𝑓𝑓 × 100 𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶 CDA: coeficiente de digestibilidad aparente 1.3.2 Método in vitro. Busca imitar artificialmente los procesos fisiológicos que ocurren en el sistema digestivo del animal, realizando los procesos mecánicos y enzimáticos (pepsina, tripsina, celulasa y líquido ruminal) del alimento en el laboratorio, exponiéndolo a periodos de incubación, por lo cual se espera que la cantidad de alimento que desaparece por acción de las enzimas sea la cantidad digestible. Se desarrolla fácilmente, es más económico, los resultados obtenidos son confiables al compararlo con la técnica in vivo, sin embargo tiene mayor probabilidad de error al reemplazar los animales (Church, 1993; Caravaca, et al, 2003). 1.3.3 Método in situ. Esta prueba emplea animales con cánulas en distintos segmentos del tracto digestivo, dejando en las cánulas bolsas de nylon que contienen específica cantidad de alimento por un tiempo establecido, el tiempo puede variar dependiendo de la calidad o tratamiento de conservación del forraje (Caravaca, et al, 2003). Otro de los métodos para conocer la digestibilidad es a través de sustancias indigestibles que pueden estar en el propio alimento y son expulsadas en las heces, entre ellos están los cromógenos extraíbles con acetona, la lignina, y hay otros que se pueden mezclar con el alimento como el óxido de cromo. También existen los métodos predictivos por medio de las ecuaciones de regresión en el cual hay algunos parámetros químicos obtenidos en 16 laboratorio como la FB en monogástricos, la FAD y lignina en rumiantes (Caravaca, et al, 2003). 1.4 GRAMÍNEAS DEL TRÓPICO SECO 1.4.1 Pasto Vidal (Bothriochloa saccharoides). Crece en forma menuda y tupida, presenta una raíz fuerte y rústica, tiene un tamaño de 1,2-1,5 m, hojas planas lanceoladas de 10 cm. a 30 cm. con 2 mm a 8 mm de ancho (Vidal et al., 2007). Este pasto se ha reportado en diferentes regiones de Colombia de difíciles condiciones climáticas, como el bosque seco de la costa Caribe, el bosque subhúmedo tropical de la meseta del Tolima y el clima oceánico de San Andrés y Providencia (Piñeros et al., 2011), igualmente presenta alto grado de rusticidad, muy adaptable a laderas, con una excelente capacidad de invasión, favoreciendo la reducción de arvenses en el potrero y con capacidad de competir con el pasto Colosoana (Bothriochloa pertusa) (Vidal et al., 2007). Estudios reportados sobre esta gramínea son pocos; Piñeros et al. (2009) hallaron valores para la proteína cruda (PC) de 8,3 y 12,9 para época húmeda y seca respectivamente a libre exposición y sin fertilización, además de esto concluyeron que el pasto Vidal tienen un buen comportamiento bajo sombra lo cual es favorable al implementarlo en sistemas agroforestales (Piñeros et al., 2009), igualmente Piñeros et al. (2010) realizaron un trabajo donde determinaron las características nutricionales más relevantes durante el desarrollo del ciclo del pasto y encontraron que en estado de prefloración la proteína cruda (PC) es de 5,24%; digestibilidad in vitro de materia seca (DIVMS) 54,21%; 45,8% de fibra detergente ácida (FDA) y 8,3% de lignina (tabla 1). Según los autores estas diferencias entre los dos trabajos se podrían explicar por las variaciones de las condiciones agroclimáticas presentes en cada zona donde se realizaron cada uno de los estudios. 17 Tabla 1. Composición química y digestibilidad de la MS en diferentes etapas del ciclo fenológico del Bothriochloa saccharoides. DIVMS EB PC EE Ceniz FDN FDA Lig CHO Hem Cel. (%) (Kcal/ (%) (%) a (%) (%) (%) (%) S (%) . (%) Etapas Germinación Día 27 Vegetativa Día 91 Reproductiva Día 114 Senescencia Día 128 (%) Kg) 56,12 3,87 54,21 3,97 53,90 3,78 53,22 3,98 6,1 1,4 5 9 5,2 1,9 4 7 5,1 1,2 2 6 3,5 2,0 6 2 12,61 11,63 9,87 9,15 70,4 43,3 7,0 1 6 2 72,1 45,8 1 10,34 8,3 11,24 9,4 13,99 74,5 46,2 6 1 75,3 47,0 9,7 6 8 3 13,83 27,0 35,0 5 6 25,8 36,8 9 1 24,0 37,3 4 5 22,1 38,7 9 9 DIVMS: digestibilidad de la materia seca; EB: energía bruta; PC: proteína cruda; FDN: fibra detergente neutra; FDA: fibra detergente acida; Lig: lignina; CHOS: carbohidratos no fibrosos; Hem: hemicelulosa; Cel.: celulosa Fuente: Piñeros et al. (2010) En un experimento realizado por Arcos et al. (s.f.) evaluaron seis gramíneas (Pasto CORPOICA 6 Vidal -Bothriochloa saccharoides- ; Colosoana -Bothriochloa pertusa-; pasto Angleton Climacuna -Dichamthium annulatum-; Pasto Angleton mono o común Dichamthium aristatum-; Pasto Braquiaria la Libertad -Brachiaria brizantha- y Pasto Llanero -Brachiaria dictyoneura-), establecidas empleando material vegetativo (cepas de diez tallos, aproximadamente) y una leguminosa (Campanita o zapatico de reina Clitoria ternatea ), determinaron que el pasto que tuvo el mejor porcentaje de prendimiento del material vegetativo (cepa) fue el pasto Vidal con 99,3 % seguido del Colosoana, climacuna, Angleton mono, pasto llanero y por ultimo Brachiaria la Libertad. En el mismo estudio evaluaron la producción de Materia seca (MS) de las mismas especies en época húmeda y seca respectivamente, siendo el pasto Vidal el de la mayor producción de lo cual afirman que merece especial observación debido al gran potencial evidenciado en este estudio. 18 1.4.2 Colosoana (Bothriochloa pertusa). Esta es una especie perenne, comúnmente con estolones rosados o rojos. Tallos desde los 50 cm hasta 90 cm de largo. Las hojas son de color verde grisáceo, de menos de 30 cm de largo y de 2 a 6 mm de ancho. Inflorescencia subdigitada o digitada, color púrpura de 3 a 13 racimos, de hasta 7 cm de largo. Floración depende del ecotipo. Produce cosechas significativas de semilla y desarrolla estolones fuertes. Se puede lograr cosecha de semillas de 70 a 100 kg/ha (Corpoica & Universidad Nacional de Colombia, 2013a). El Colosoana es una gramínea de productividad aceptable, posee crecimiento vegetativo, con una amplia adaptación a condiciones de suelos de baja fertilidad en áreas de escasa precipitación y buena tolerancia al pisoteo, características que la convierten en una gramínea agresiva. A pesar de esto se reporta que este pasto tiene suceptibilidad al mión o Salivazo (Sierra et al., 1988). En un estudio realizado por Piñeros, Tobar, y Mora, (2011) uno de los objetivos fue determinar la calidad nutricional del pasto Colosoana en tres tiempos diferentes, bajo un monocultivo y sistema silvopastoril (SSP) con Leucaena (Leucaena leucocephala) (tabla 2), y la cantidad de biomasa de los dos tratamientos; en los resultados encontraron que la calidad nutricional y la producción de biomasa en el monocultivo con respecto al sistema SSP fue superior, por lo que concluyeron que la baja productividad del B. pertusa en el SSP, se puede explicar por la poca tolerancia de esta especie a la sombra. En otro trabajo realizado por Sánchez et al. (2005) en el cual determinan la cinética de degradación de algunas gramíneas y leguminosas tropicales, hallaron para el pasto Colosoana valores de Pc del 2.77 % cortado a los 35 días. Además reportaron dos fracciones para proteína del Cornell Net Carbohydrate and Protein System (CNCPS): proteína insoluble en detergente neutro (PIDN) y proteína insoluble en detergente ácido (PIDA). PIDN fue de 30.49 % y PIDA de 25.33 % con respecto al porcentaje total de proteína. La DIVMS reportada para este pasto es de 73.1% y 69.85% en época de lluvia y de sequía respectivamente; en cuanto al consumo de MS con respecto al peso vivo (PV) de novillos 19 en pastoreo está alrededor de 2.01% en época de lluvia y 0.9 % en época seca (Chamorro, Carulla, & Cuesta, 2005). Tabla 2. Análisis bromatológico del pasto B. pertusa en tres tiempos diferentes de corte. Tratamiento Control (SM) Tiempo PB FDN FDA Cel Hcel Lignina EM EE Cenizas 7 15,44 72,91 40,28 23,9 27,66 4,9 1344,6 1,38 14,33 21 9,44 72,16 38,3 26,33 29,16 5,6 1378,6 1,71 13,28 28 8,31 77,98 50,51 23 22,77 13,09 1168,9 1,27 12,98 7 15,44 73,4 37,13 24,58 31,54 1,51 13,98,7 1,59 13,15 Leucaena 21 8,06 75,51 37,46 22,7 33,68 4,14 13,930 1,46 12,56 (SSP) 28 6,81 78,57 45,64 24,16 28,73 9,3 1252,5 1,53 11,84 PB: proteína bruta; FDN: fibra detergente neutra; FDA: fibra detergente acida; Cel: celulosa; Hcel: hemicelulosa; EM: energía metabolizable (kcal/kg); EE: extracto etéreo. Fuente: Varón et al., (2011). 1.4.3 Angleton Climacuna (Dichanthium annulatum) - Angleton mono (Dichanthium aristatum). el Angleton climacuna es una planta perenne estolonífera en ocasiones. Tallos geniculados ascendentes con anillos pronunciados de 3.5 mm de largo sobre los nudos y macollas fértiles de aproximadamente 1 m. Hojas lanceoladas de cerca de 30 cm y de 2 a 7 mm de ancho, márgenes ligeramente pubescentes (Corpoica & Universidad Nacional de Colombia, 2013b) crece en matojos, su mejor desarrollo lo presenta en una textura franco arcilloso, es una gramínea rastrera pero que se asocia bien con leguminosas lo que puede mejorar significativamente su desempeño, se ha difundido especialmente en Valle del Sinú y en el Alto Magdalena (Vanegas & Riveros, 1999). En estado de prefloración reporta valores: proteína de 10.3%, DIVMS del 69%, FDN del 64% y lignina del 3.4 % indicadores de un buen valor nutritivo para especies tropicales (Corpoica, 2005). El Pasto Climacuna se utiliza en una gran proporción para la fabricación de heno como alternativa de conservación de forrajes en época de sequía, su producción es mayor comparada con al Angleton mono debido una mayor relación hoja/tallo (Cuesta et al., s.f.). El Angleton mono es una especie perenne de crecimiento erecto o semierecto, con buen macollamiento. Se propaga por semilla y por tallos. Se adapta a diferentes tipos de suelo, pero los mejores rendimientos se obtienen en suelos francos y fértiles con buena humedad, bien drenados. Crece bien a alturas entre 0 y 1.000 msnm, con precipitaciones 20 entre 750 y 2.200 mm al año. Tolera la quema, el pisoteo y la sequía; sin embargo, los rendimientos de forraje se reducen considerablemente en la época seca, por lo cual se recomienda la aplicación de riego (Corpoica, 2005). Presenta PC del 10-16%, digestibilidad de la MS de 65-75% (Corpoica & Universidad Nacional de Colombia, 2013c), FDN 74%, FDA 43.4%, cenizas 9.6 % y EE de 1% (Ruiz, 2011). Sánchez et al., (2005), reporta valores de PC 6.5 %, FDN 69.07 %. FDA 40.09 %, EE 0.94 %, Czas % 11,78%, CNET 11.66% y PIDN fue de 40.65 % y PIDA de 18.29% con respecto al porcentaje total de proteína. En estudios realizados en el Sur del departamento del Tolima por Arcos et al., (s.f.), en la cual evaluaron la producción de MS de Dichanthium Annulatum y Dichanthium Aristatum, llegaron a la conclusión que el Angleton mono no se recomienda para la región, ya que bajo las condiciones de estudio, durante la época de máxima precipitación presentó la menor producción de materia seca y, durante la época de mínima precipitación, desapareció. Por esta razón no es recomendable para la zona. Entre tanto la utilización de Angleton climacuna aparece como una alternativa para suelos en los cuales sea posible la utilización de riego. 1.5 CONSUMO VOLUNTARIO El consumo voluntario de un alimento, se define como la cantidad de alimento consumido por un animal o grupo de animales durante un periodo de tiempo, en el cual los animales tienen libre acceso al alimento (Ramírez, 2005). El consumo voluntario es probablemente el factor más importante desde el punto de vista de productividad pecuaria, ya que todos los parámetros del comportamiento animal como son la ganancia de peso, la secreción de leche, el crecimiento de la lana, etc, dependen de forma directa del factor en cuestión; en términos generales se busca que el animal consuma más, ya que en individuos sanos esto se traduce en mayor producción (Shimada, 2005). 21 El control del apetito a largo y a corto plazo está relacionado con la energía más que con cualquier otro nutriente (Church, 1997). Los mecanismos que regulan la ingestión de los alimentos en los rumiantes, operan en tres niveles. A nivel metabólico, a partir de la concentración de nutrientes metabolitos y hormonas que actúan sobre el sistema nervioso pueden provocar que los animales empiece o terminen de comer (McDonald et al., 2006). A nivel del sistema digestivo, los niveles de productos de la digestión pueden determinar que los animales consuman más cantidad de alimentos. Por último, las influencias externas como las variables climáticas o la facilidad con que puedan ser ingeridos los alimentos, también influyen sobre el consumo (McDonald et al., 2006). La cantidad necesaria de nutrientes necesarios para el mantenimiento y producción en rumiantes, dependerá de la ingesta de materia seca. Esto necesita ser predicho en una forma precisa para lograr un alto rendimiento y para evitar gastos innecesarios con la alimentación, ya que este componente representa mayores costos para la producción animal (Vieira et al., 2013). Según Shimada (2005) el consumo de MS puede medirse de dos maneras: 1) Por medición directa del forraje consumido (Alimento suministrado-Rechazo). 2) Por el uso de marcadores y colecta total de heces. El consumo de MS se puede expresar con respecto al peso vivo o al peso metabólico; la primera sirve para hacer comparaciones entre la misma especie y la segunda para realizar confrontaciones entre diferentes especies; esta se halla elevando la masa en kilogramos a la 0.75 (Kg 0.75) lo cual se refiere a la producción de calor en función a la superficie corporal (Brautigan, 2007). Maloiy et al., (2008) evaluaron el consumo con respecto al peso metabólico en especies de rumiantes domésticos y no domésticos pudiendo hacer comparaciones entre ellos a partir de la superficie corporal. Igualmente (Riaz et al., 2014), realizaron un experimento donde midieron el consumo de MS entre pequeños y grandes rumiantes y encontraron diferencias pronunciadas con exponentes más bajos para las ovejas seguido de las cabras y más altos para los bovinos y búfalos; esta medicion la hicieron a partir de la estimacion del peso metabólico. 22 1.5.1 Factores que afectan el consumo. un trabajo realizado por Riaz et al., (2014) compararon la digestibilidad y el consumo de materia seca de bovinos, caprinos, ovinos y búfalos. Encontraron que la cantidad de proteína presente en la dieta influye positivamente sobre el consumo de materia seca en las cuatro especies. La presencia de alto contenido de FDN y FDA en la dieta, afecto negativamente el consumo en todas las especies mencionadas, menos en el búfalo donde no se detectaron cambios. El efecto más marcado se presenta en ovinos y bovinos. Maloiy et al., (2008) compararon especies de rumiantes domésticos y no domésticos en Sabana Africana y encontraron que la deshidratación afectaba el consumo de alimento en las cabras, seguidas por los bovinos (Cebú). La temperatura (40°C) también fue un factor que afecto significativamente el consumo de forrajes especialmente en animales domésticos y la interacción entre los dos factores medioambientales redujo significativamente el consumo a un 62%. Otro factor que puede alterar el consumo, es la presencia de metabolitos secundarios, como los taninos, importantes en las dicotiledóneas y de éstas, en las leguminosas, sin embargo también están presentes en casi todos los géneros vegetales como las gramíneas (McLeod, 1974). La presencia de estos puede alterar el consumo de alimento debido a su efecto astringente que disminuye la palatabilidad. 23 2. OBJETIVOS 2.1 OBJETIVO GENERAL Evaluar la calidad nutricional, digestibilidad y consumo de gramíneas henificadas: Pasto Vidal (Bothriochloa saccharoides), Colosoana (Bothriochloa pertusa) y Angleton Climacuna (Dichanthium annulatum). 2.2 OBJETIVOS ESPECÍFICOS Evaluar la calidad nutricional de las gramíneas henificadas: Vidal (Bothriochloa saccharoides), Colosoana (Bothriochloa pertusa) y Angleton Climacuna (Dichanthium annulatum). Comparar la digestibilidad in vivo de las gramíneas evaluadas. Determinar el consumo de Materia seca de las diferentes gramíneas henificadas en ovinos de pelo Identificar metabolitos secundarios de las gramíneas Bothriochloa saccharoides, Bothriochloa pertusa y Dichanthium annulatum. Estimar el fraccionamiento de proteína Cornell, de los henos evaluados 24 3. METODOLOGÍA El estudio se realizó en el valle cálido del Magdalena Medio, en la Granja del Centro Universitario Regional del Norte del Tolima (CURDN) de la Universidad del Tolima ubicada en Armero Guayabal (5°01′50″N -74°53′08″O) -Tolima, que cuenta con las siguientes condiciones ambientales: altitud de 275 m.s.n.m, temperatura de 26ºC y precipitación anual de 1732 mm, parámetros que lo clasifican según Holdridge, et al., (1971) como Bosque Seco Tropical. 3.1 TRATAMIENTOS EVALUADOS Los tratamientos consistieron en dietas a base de gramíneas henificadas, las cuales fueron picadas hasta alcanzar un tamaño de partícula 4 -7 cm aproximadamente. T1: heno de pasto Vidal (Bothriochloa saccharoides) T2: heno de pasto Angleton Climacuna (Dichanthium annulatum) T3: heno de pasto Colosoana (Bothriochloa pertusa) El heno del pasto Vidal se obtuvo de la Hacienda “La Estrella” del Dr. Héctor Vidal Perdomo, ubicada en el municipio de Venadillo-Tolima, a 348 m.s.n.m. con una temperatura promedio de 26 °C; correspondiente a una zona de vida de bosque subhúmedo tropical. El heno de los pastos Angleton Climacuna y Colosoana, fueron adquiridos en la Granja el CURDN de la Universidad del Tolima, en la cual se realizó el experimento. La edad del rebrote donde se hizo el corte de los pastos evaluados, fue a los 60 - 70 días aproximadamente. Se utilizaron 12 ovinos de pelo machos enteros mestizos con un peso promedio de 26 ± 4,49 Kg, de los cuales 9 fueron empleados en el diseño experimental y los otros 3 se manejaron como reemplazos en caso de presentarse alguna eventualidad. 25 El método utilizado para determinar la digestibilidad de las gramíneas fue la colecta total de heces (CTH), Cada periodo experimental fue de 17 días: 14 días para adaptación a las dietas experimentales y 3 días de evaluación (alimentos ofrecidos, rechazos, heces y orina) completándose así la fase experimental en 51 días desde el 14 de Octubre hasta el 3 de Diciembre del 2013. 3.2 DIGESTIBILIDAD POR COLECTA TOTAL DE HECES Este método se realizó teniendo en cuenta la normativa bioética para experimentación animal contemplada por la Universidad del Tolima. 3.2.1 Periodo de adaptación. El tiempo de acostumbramiento a la dieta se realizó en corrales independientes para cada tratamiento; el tiempo dedicado para este periodo fue de 14 días. Se suministró a cada grupo de animales 4 kg de heno del tratamiento correspondiente con base a un consumo de materia seca del 3% del peso vivo, ajustándolo para obtener 5-10 % de rechazos del alimento total ofrecido en el transcurso de cada periodo hasta estandarizar el consumo del alimento; también se suministró diariamente agua a voluntad. 3.2.2 Periodo en las jaulas. Para el experimento se emplearon jaulas metabólicas individuales con dimensiones de 1 m de ancho, 1 m de alto y 1,5 m de largo, equipadas con colectores, separadores de heces y orina, con comederos y bebederos individuales. El tiempo de permanencia de los animales en las jaulas metabólicas fue de tres (3) días concernientes a la duración de la recolección de las muestras, esto basado en una previa estandarización. 3.2.3 Muestreo. Al iniciar la toma de datos de cada periodo, se pesaba el alimento (1 kg) propio de cada tratamiento y se registraba para posteriormente suministrarlo a cada animal en dos tiempos 9:00 am y 3:00 pm. Pasadas 24 horas se recogían los rechazos y las heces en bolsas para luego pesarlos y posteriormente almacenarlos. De esta forma se registraban los datos durante tres días en cada uno de los periodos. Al igual que en el periodo de adaptación, para esta fase los animales tuvieron a disposición agua voluntad. 26 Para realizar los análisis bromatológicos y determinar la digestibilidad in vivo de los nutrientes de cada uno de los forrajes, se obtuvieron muestras de los pastos henificados y las heces. Las muestras fueron almacenadas en un refrigerador hasta terminar el periodo de colecta (3 días) para luego ser enviadas al laboratorio de Ecofisiología de la Universidad del Tolima. 3.3 CONSUMO DE MATERIA SECA. Simultáneamente a la evaluación de la digestibilidad, se realizó la medición del consumo, esta consistió en pesar la cantidad de alimento suministrado a cada uno de los animales y 24 horas después pesar los rechazos para así obtener un consumo aparente de los henos. Posteriormente fue analizado el porcentaje de MS de los forrajes y este porcentaje se aplicaba a la cantidad consumida durante el experimento para así obtener el consumo de MS. Este valor fue representado en gramos por animal, % MS con respecto al peso vivo y con respecto al peso metabólico (kg W 0,75) para cada uno de los tratamientos evaluados. 3.4 ANÁLISIS BROMATOLÓGICOS Las muestras colectadas de pasto y heces fueron analizadas, determinando el valor de materia seca (MS), materia orgánica (MO), proteína cruda, extracto etéreo y cenizas, mediante análisis químico proximal según los métodos establecidos por la AOAC (2000); fibra detergente neutra y fibra detergente ácida por medio del protocolo propuesto por (Van Soest, Robertson, & Lewis, 1991). En el momento que se tuvieron todos los análisis bromatológicos tanto de pastos como las heces, se procedió a determinar la digestibilidad aparente (ecuación 1) de la materia seca, materia orgánica, proteína cruda, Fibra detergente neutra, y extracto etéreo. Ecuación 1. CDA: Coeficiente digestibilidad aparente. 𝐶𝐶𝐶𝐶𝐶𝐶 (%) = 𝑐𝑐𝑐𝑐𝑐𝑐𝑐𝑐𝑐𝑐𝑐𝑐𝑐𝑐 − 𝑒𝑒𝑒𝑒𝑒𝑒𝑒𝑒𝑒𝑒𝑒𝑒𝑒𝑒ó𝑛𝑛 𝑓𝑓𝑓𝑓𝑓𝑓𝑓𝑓𝑓𝑓 × 100 𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶 27 Los nutrientes digestibles totales (NDT) de las gramíneas, fueron estimados a partir de la ecuación descrita por NRC, (2001). Ecuación 2. Nutrientes digestibles totales. NDT (%) = (CNF*dCNF) + (Pc*dPc)+ (FDN*dFDN) + (EE*dEE) x 2,25 – 7 Donde: dCNF: digestibilidad de los carbohidratos no fibrosos dPc: Digestibilidad de la proteína cruda dFDN: digestibilidad de la fibra detergente neutra dEE: digestibilidad del extracto etéreo 3.5 FRACCIONAMIENTO PROTEICO Las fracciones proteicas CNCPS fueron determinadas de acuerdo con el procedimiento estandarizado por Licitra, Hernandez, & Van Soest, (1996). La fracción A (NNP) fue medida a partir tungstato de sodio, el cual precipita la proteína y permite cuantificar el NNP; la fracción B1, soluble en tampón fosfato fue cuantificado utilizando el tampón borato fosfato menos el nitrógeno soluble en la fracción A; el nitrógeno insoluble en detergente neutro B3 (NIDN) y el nitrógeno insoluble en detergente ácido, C (NIDA) fueron determinados como el contenido de nitrógeno residual, posterior a los procedimientos de detergente neutro y detergente ácido respectivamente. La fracción B2 insoluble en el tampón borato fosfato, pero soluble en detergente neutro, fue calculada como la diferencia entre el resto de fracciones. Los resultados fueron presentados como porcentaje con respecto al total de la proteína cruda. Se determinó la digestibilidad de dos de las fracciones proteicas (B3- C) por su relación con taninos, celulosa y lignina; para esto se hallaron estas fracciones tanto en el pasto como en las heces y se reemplazó en la ecuación de digestibilidad aparente descrita anteriormente. 28 3.6 METABOLITOS SECUNDARIOS La determinación cualitativa de metabolitos secundarios (fenoles, saponinas, esteroles y alcaloides) se realizó mediante la metodología de Domínguez, (1973) en el Laboratorio de Nutrición Animal de la Universidad de Nariño. 3.7 MODELO ESTADÍSTICO Se hizo un diseño experimental de cuadrado Latino 3*3, con tres tratamientos y tres periodos experimentales. El número de repeticiones fue n= 27. Se realizó un análisis de varianza ANOVA para establecer las diferencias estadísticas entre tratamientos. La diferencia de medias se determinaron por una prueba de honestidad de TUKEY (p= 0.05). Todos estos cálculos se desarrollaron por medio del programa Infostat®. El modelo estadístico fue: Yijk=µ+Ai +Pj +Tk+eijk Dónde: Yijk=respuestas al tratamiento k, del animal i, en el período j; µ=media de los tratamientos; Ai =efecto del animal i, (1...4); Pj =efecto del periodo j, (1...4); Tk=efecto del tratamiento k, (1...3); eijk=error residual. 29 4. RESULTADOS Y DISCUSIÓN 4.1 BROMATOLÓGICOS El porcentaje de MS encontrada para los pastos Bothriochloa saccharoides, Bothriochloa pertusa y Dichanthium annulatum, vario entre 87% a 88% (tabla 4) lo cual está dentro del rango esperado si se tiene en cuenta que estas gramíneas fueron henificadas para facilitar las condiciones del experimento, entre tanto, un heno de cualquier gramínea de trópico seco, normalmente tiene un contenido de MS que esta próximo 85% (Sánchez, Wills, & Figueroa, 2014). Tabla 4. Composición Bromatológica del Bothriochloa saccharoides, Bothriochloa pertusa y Dichanthium annulatum. Nutriente TRATAMIENTOS B. saccharoides D. annulatum B. pertusa MS 88,1 ± 2,5 87,6 ± 1,2 87,9 ± 1,6 MO 93,6 ± 0,9 90,2 ± 1 91,1 ± 0,6 PC 6,7 ± 0,3 7,1 ± 0,9 6,7 ± 0,9 FDN 79,6 ± 1 72,6 ± 0,5 73,3 ± 0,3 FDA 54,3 ± 0,6 53 ± 1,6 54,2 ± 1 Hem. 25,2 ± 1,5 19,5 ± 2,1 19 ± 1 Cel. 41,5 ± 2,7 43,0 ± 2,1 42,8 ± 2,5 Lig. 12,8,9 ± 2,4 10 ± 1,7 11,4 ± 2,7 EE 1,6 ± 0,4 3,5 ± 1,4 4,8 ± 1,6 Czas 6,3 ± 0,9 9,7 ± 1 9,3 ± 0,6 MS: materia Seca; MO: materia orgánica; PC: proteína cruda; FDN: fibra detergente neutra; FDA: fibra detergente ácida; Cel: Celulosa; Hem: hemicelulosa; Lig: lignina; EE: extracto etéreo. Czas: cenizas. Fuente: autores 30 4.1.1 Componentes estructurales. Entre las tres gramíneas evaluadas, el pasto B. saccharoides presentó el mayor porcentaje de FDN (79,6%), dado a la cantidad de hemicelulosa presente en sus fracciones estructurales (tabla 4). Con respecto a las gramíneas B. pertusa y D. annulatum, la FDN fue inferior, 73,3 % y 72,6%, variable atribuida, posiblemente a la etapa del ciclo fenológico en el que se encontraban los pastos cuando fueron cortados. (Piñeros et al., 2010; Krämer et al., 2012). La FDA fue similar para las tres gramíneas estudiadas (tabla 4); como se mencionó anteriormente, por la mayor presencia de hemicelulosa que es excluida al ser sometida a la solución de detergente ácido (Van Soest et al., 1991; Herrick et al., 2012). Debido a esta similitud, la FDA (celulosa y lignina) no es un factor que se pueda considerar una causa diferencial de la digestibilidad en este estudio. (Jung, Mertens, & Payne, 1997). Del mismo modo la cantidad de lignina encontrada tuvo valores semejantes en los tres tipos de pastos (tabla 4), lo cual es un indicativo de madures de las plantas, que se da como mecanismo de adaptación a las condiciones medioambientales presentadas en los lugares donde se cosecharon los henos evaluados (Fukushima et al., 2015). Entre los resultados expuestos anteriormente para las tres gramíneas, se destaca el alto contenido de componentes estructurales, los cuales fueron más altos a los reportados por otros autores (Piñeros et al., 2010; Vanegas & Riveros, 1999; Corpoica, 2005). A partir de estos datos se puede inferir que estas gramíneas no fueron colectadas en la fase indicada del ciclo vegetativo, donde la expresión de los nutrientes está en un mejor nivel de garantía para la planta y el animal (Corson, Rotz, & Skinner, 2007). 4.1.2 La proteína cruda. (Pc) de los henos de B. saccharoides, D. annulatum y B. pertusa oscilo entre 6,6% y 7,1% (tabla 4). Piñeros et al., (2009) hallaron valores de Pc para B. saccharoides de 8,3 y 12,9 para época húmeda y seca respectivamente a libre exposición y sin fertilización. Para el D. annulatum Vanegas & Riveros, (s.f.) y Corpoica, (2005) describen en estado de prefloración una proteína igual al 10,3%. Entre tanto para pasto B. pertusa Sánchez et al., (2005) reportan un contenido de Pc de 2,77% a los 35 días de rebrote; sin embrago Varón et al., (2011) hallaron valores más altos de Pc para los días 31 21 y 28 equivalentes a 9,44 y 8,31% respectivamente. Estas diferencias presentadas en los valores proteicos de esta gramínea, pueden estar relacionados, con su ciclo fenológico, el cual es más corto para esta especie y que por desconocimiento no se realiza el corte o el pastoreo en el periodo indicado donde los valores nutricionales están en una mejor expresión (Varón et al., 2011). 4.1.2.1 Fraccionamiento Nitrogenado. En el análisis de las diferentes fracciones, determinó que la fracción A, fue predominante en los pastos analizados con un porcentaje que varió de 49 a 63% (tabla 5) en relación al porcentaje total de la proteína que oscilo 6,7 a 7,1%. Esto puede ser atribuido a la alta absorción de componentes no proteicos por parte de las plantas para el crecimiento y formación de sus estructuras (Jampeetong et al., 2014). Sin embargo es poca la información desarrollada en la evaluación y composición de las fracciones nitrogenadas de las plantas de trópico seco. Tabla 5. Fracciones nitrogenadas, con respecto al porcentaje total de la proteína de cada una de las gramíneas. Fracciones B. saccharoides D. annulatum B. pertusa A (% PC) 58,14 ± 2,83 63,4 ± 0,41 49,01 ± 0,64 B1 (% PC) 1,98 ± 0,80 2,01 ± 1,33 4,37 ± 0,15 B2 (% PC) 11,3 ± 1,15 6,47 ± 1,46 15,82 ± 0,55 B3 (% PC) 1,58 ± 0,08 2,88 ± 3,06 4,09 ± 1,00 C (% PC) 26,9 ± 1,81 26,31 ± 0,88 28,04 ± 2,4 A= fracción asociada con NNP altamente degrada en el rumen; B1, B2, B3 = rápidamente, moderadamente, lentamente, degradadas en el rumen; C= fracción no degradada en el rumen e indigestible en el intestino. Fuente: autores La fracción proteica B1, presentó un bajo porcentaje para las tres gramíneas (tabla 5) en relación a la totalidad de la proteína cruda, lo que la hace un componente poco influyente en la digestibilidad total del nitrógeno. 32 La gramínea B. pertusa registró el mayor contenido de la fracción B2 (15,82%) seguido por B. saccharoides (11,3%) y el contenido más bajo lo obtuvo el D. annulatum (6,47%). En cuanto a la fracción B3 (tabla 5) tuvo una baja participación dentro del porcentaje total de la proteína de los pasto B. saccharoides (1,58%); B. pertusa (4,09%) y D. annulatum (2,88%). Los estudios en fraccionamiento nitrogenado de pastos tropicales son escasos. Sin embargo Sánchez et al., (2005), reportan porcentajes de proteína indigestible en detergente neutro (B3), para B. pertusa de 30,49% lo cual difiere de lo encontrado en el presente trabajo, posiblemente por la madures del pasto B. pertusa utilizado en ese estudio . Del mismo modo, se han reportaron valores de la fracción B3 para las gramíneas Dichanthium aristatum (40.65), Brachiaria decumbens (29.9%) y Cynodon dactylon (23,18%) (Sánchez et al., 2005; Branco et al., 2012). Las gramíneas estudiadas, presentaron un contenido similar de fracción C (tabla 5), que osciló entre 26,9 a 28,04. En el trabajo hecho por Sánchez et al., (2005), encontrarón valores de proteina indigestible en detergente ácido (fracción C) para B. pertusa de 25,33%, lo cual coincide con lo hallado en el presente trabajo. En el mismo estudio encontraron para D. aristatum y B. decumbens, 18,29% y 2,8% respectivamente para fracción C. Estos valores evidenciados, al parecer son proporcionales al contenido de lignina de cada planta en función del tiempo y las condiciones ambientales (Cong et al., 2013). 4.1.3 Extracto Etéreo. B. Saccharoides para este estudio presentó un EE de 1,6% (tabla 4) lo cual concuerda con valores descritos por Piñeros et al., (2010) que encontró datos de 1,26 % y 2,02 en senescencia. El EE del D. annulatum fue de 3,5% (tabla 4). Cardona et al., (2002), para el D. aristatum encuentran valores del 2,2%; Posada, (2011) reporta 1%. El B. Pertusa evidenció 4,8% de EE (tabla 4) el cual no se relaciona con el valor hallado por Varón et al., (2011) puesto que reportan 1,27% para el día 28 de su ciclo fenológico, similar a lo descrito por Sánchez et al., (2005) que reporta 1,02 % de EE. Según un estudio realizado por Kaul y Vats, (1998), el B. pertusa siendo una gramínea oleaginosa y aromática en su composición química de aceites esenciales evidencia 33 aproximadamente 28 compuestos entre ellos están los sesquiterpenos oxigenados, intermedeol y acorenone-B como compuestos principales, de igual forma pero en menor proporción se hallaron otras sustancias como el kessane que estimula el gusto en el ganado, por esta razón y la baja concentración de acorenone-B (reduce palatabilidad) se explica que esta gramínea sea de mayor aceptación por parte de los animales y también permite relacionar el mayor valor de EE obtenido en el presente estudio con su propiedad oleaginosa. Sin embargo la determinación del extracto etéreo es algo inespecífica debido a que se desprecian algunos componentes importantes y además se asume la presencia de triglicéridos predominando en las gramíneas los galactolípidos que no tienen el mismo contenido energético (Estrada, 2002). 4.2 CONSUMO VOLUNTARIO El consumo de MS de las diferentes gramíneas con respecto al peso vivo y al peso metabólico (W kg 0,75) en los diferentes periodos, medido a los animales en el trascurso del experimento, presentó diferencias estadísticamente significativas (p˂0,05) entre los pastos B. saccharoides frente a D. annulatum y B. Pertusa, destacando este último como el forraje de mejor consumo durante los periodos experimentales (tabla 6). 34 Tabla 6. Consumo de MS con respecto al peso vivo y el peso metabólico de las gramíneas evaluadas. B. D. saccharoides annulatum B. pertusa EEM PValor ----- ----- Peso vivo (kg) 25,03 ± 3,83 25,03 ± 3,83 25,03 ± 3,83 Peso metabólico 11,17 ± 1,28 11,17 ± 1,28 11,17 ± 1,28 ----- ----- 1,74a 2,06ab 2,20b 0,11 0,0109 3,87a 4,58ab 4,60b 0,23 0,0462 W 0,75 kg Consumo de MS (% de PV) Consumo de MS kg / kg W 0,75 (%) EEM: error estándar de la media; medias con una letra común no son significativamente diferentes (p˃0,05). Fuente: autores Estas diferencias presentadas, pueden estar relacionadas con el contenido de FDN del B. saccharoides (79,6%), con respecto al FDN del D. annulatum (72,6 %) y el B. pertusa (73,3 %), donde el alto contenido de los componentes estructurales de la fibra dentro de la dieta de los animales, afecta considerablemente el consumo de alimento debido a su lenta degradabilidad ruminal y la baja tasa de pasaje (Barahona & Sánchez, 2005; Riaz et al., 2014). 35 A pesar de haber encontrado estas diferencias en el consumo de MS (p˂0,05) (tabla 6) de los diferentes forrajes, se evidencia que estos consumos en general son bajos si se comparan con otros trabajos donde reportan consumos de gramíneas superiores al 3% con respecto al peso vivo (Ngwa, Nsahlai, & Iji, 2002); (Misra et al., 2006). Además de los factores nutricionales, los ambientales, también ocupan un papel importante en el consumo; el manejo experimental, la temperatura, humedad entre otras condiciones, ocasionan en los animales un grado de estrés, el cual puede llegar a reducir el consumo hasta en un 60% como lo reporta Maloiy et al., (2008). El estrés puede llegar a producir un incremento en los niveles de diferentes sustancias químicas, entre ellas el cortisol, inhibiendo la producción de insulina, permitiendo que la glucosa quede circulante en sangre y el animal experimente una sensación de saciedad, evitando de este modo el consumo de alimento (Marai et al ., 2007). Otro factor que puede influir en el bajo consumo de los forrajes, es la presencia de metabolitos secundarios, que se presentan en las plantas como un mecanismo de defensa ante los diferentes factores medioambientales y los cuales pueden intervenir sobre el consumo y la digestibilidad. 4.3 METABOLITOS SECUNDARIOS Metabolitos secundarios obtenidos (tabla 7) evidencia que las tres especies presentaron contenido de fenoles y esteroles, excepto el B. saccharoides en el que se halló además de estas, un contenido bajo de saponinas. Tabla 7. Metabolitos secundarios de las gramíneas Bothriochloa saccharoides, Bothriochloa pertusa y Dichanthium annulatum. Especies Fenoles Saponinas Alcaloides Esteroles a b c d e f g H I J K L + + + - + - - - - + - + B. pertusa ++ + + - - - - - - ++ - + D. annulatum ++ + + - - - - - - ++ - - B. saccharoides 36 Indicadores de presencia o ausencia: (-) Negativo, (+) Bajo, (+ +) Moderado (+ + +) Abundante. Pruebas cualitativas para la determinación de Fenoles: Cloruro férrico ( a ), Acetato de plomo ( b) y Gelatina-sal ( c ); Saponinas: Espuma ( d ), Rosenthler Vainillina ácido ortofosfórico ( e ) y Molish ( f ); Alcaloides: Dragendorff ( g ), Wagner ( h ) y Mayer ( i ); Esteroles: Liebermann ( j ), Rosenheim ( k ) y Salkowiski ( l ). Fuente: autores La presencia de fenoles en baja y moderada cantidad como se evidencia en las gramíneas (tabla 7), entre los que se destaca la lignina y los taninos, puedo influenciar el consumo de alimento como la digestibilidad. Entre tanto la lignina genera un llenado en el espacio físico del rumen, retrasando así el pasaje del alimento hacia los otros compartimientos del aparato digestivo y alterando el consumo (López, Maiztegui, & Cabrera, 1998). La posible presencia de taninos dentro del grupo de los fenoles (tabla 7), puede ser posible en este tipo de gramíneas (McLeod, 1974; Meirelles et al., 2005). Estos también pueden reducir de manera considerable el consumo de alimento debido a su efecto astringente y a la formación de complejos en el rumen, interfiriendo en la digestibilidad de las proteínas, las paredes celulares de las plantas y con grupos inhibidores de celulasas (Febres, 2008). La presencia de esteroles (tabla 7) gramíneas es bajo. Para el caso de los forrajes evaluados, se observó un contenido moderado de esteroles para B. pertusa y D. annulatum y un contenido bajo para B. saccharoides, lo cual se puede relacionar con el extracto etéreo observado (tabla 4). El pasto B. saccharoides, fue el único que presentó saponinas (tabla 7) (baja presencia); los efectos antinutricionales de esta se han descrito principalmente en los no-rumiantes, porque en los rumiantes, estos compuestos pueden ser degradados por las bacterias ruminales (Febres, 2008). Sin embargo se ha documentado que la presencia de saponinas puede generar efectos positivos sobre la flora ruminal, reducción de la concentración de amonio y de la emisión de metano en el rumen (Patra & Yu, 2014; Inamdar et al., 2015), 37 lo cual puede ser un atributo importante para cualquier alimento orientado hacia la nutrición de rumiantes. 4.4 DIGESTIBILIDAD APARENTE Fueron observadas diferencias estadísticas (p˂0,05) en la digestibilidad de algunos de los componentes nutricionales evaluados para cada una de las gramíneas. 4.4.1 Digestibilidad de la MS y MO. De modo general la digestibilidad de la MS, fue superior (p˂0,05) para el B. saccharoides (58,05%) seguido por el B. Pertusa (50,84%) y por último D. annulatum (49,64%). Al igual que la MS, la materia orgánica tuvo un comportamiento similar donde la digestibilidad más alta la obtuvo de nuevo el B. saccharoides (61,48%) seguido por B. pertusa (54,10%) y D. annulatum (52,67%) (tabla 8). Además, se observó una amplia diferencia entre la digestibilidad de la MS y la MO que puede estar dada por la presencia de un alto contenido de material inorgánico dentro de la MS de las heces, producto de la excreción endógena, lo que conlleva a subestimar la digestibilidad de la MS. Las diferencias estadísticas expuestas anteriormente, para la digestibilidad de la MS y la MO entre las diferentes gramíneas, normalmente se puede relacionar con su contenido de FDA, la cual por su composición, dificulta la degradación del alimento en el rumen y por ende la absorción y digestibilidad del alimento (Krämer et al., 2012); sin embargo, los análisis de detergente ácido realizados y observados anteriormente, evidenciaron valores similares para las tres gramíneas, que oscilaron entre 53% a 54,3% (tabla 8) lo cual no es un factor que se pueda considerar determinante para justificar las diferencias estadísticas halladas. Probablemente existen varios factores que influyeron en las diferencias encontradas entre la digestibilidad de los tres tratamientos, sin embargo el consumo de alimento por parte de los animales del experimento puede ser el factor más determinante en el momento de considerar las diferencias de la digestibilidad aparente (Riaz et al., 2014), del B. 38 saccharoides, B. pertusa y D. annulatum, si se tiene en cuenta que los mayores consumos tuvieron una relación inversa a la digestibilidad. El menor consumo de MS fue presentado por el B. saccharoides (tabla 6), al mismo tiempo este obtuvo la mejor digestibilidad (tabla 8). Van soest, (1994) señala que el consumo de MS se puede correlacionar negativamente con el tiempo de retención ruminal, lo que demuestra que entre menos MS consumida más tiempo es retenido en el rumen y mejor aprovechado por parte del animal. Otro factor que pudo influir en la alta digestibilidad de estos componentes, fue la tasa de pasaje, la cual es limitada por la alta cantidad de FDN presentada en el B. saccharoides, lo cual interviene en el tiempo de ocupación del alimento en el rumen, lo que conlleva a que la degradación sea más completa por parte del complejo de enzimas secretadas por los microorganismos y de este modo aumentar la digestibilidad aparente de estos nutrientes (López et al., 1998). Tabla 8. Digestibilidad aparente (DA) y nutrientes digestibles totales (NDT) de henos de B. saccharoides, B. pertusa y D. annulatum. B. pertusa EEM P- Valor 49,64b 50,84b 1,66 0,0010 61,48a 52,67b 54,10b 2,30 0,0181 PC 45,68 45,80 41.19 1,92 0,1594 FDN 64,31a 49,86b 53,11b 1,55 ˂0,0001 CEL 60,28a 48,73b 50,52b 2,24 0,0009 HEM 91,45a 79,92b 90,62a 1,60 ˂0,0001 EE 62,55c 76,90b 87,84a 2,08 ˂0,0001 B. saccharoides D. annulatum MS 58,05a MO 39 NDT 62,13a 50,90c 55,91b 1,33 ˂0,0001 MS: materia Seca; MO: materia orgánica; PC: proteína cruda; FND: fibra detergente neutra; Cel: celulosa; Hem: hemicelulosa; EE: extracto etéreo; CNF, carbohidratos no fibrosos; NDT; nutrientes digestibles totales. EEM: error estándar de la media; Filas que difieren de letra son estadísticamente significativas (p<0,05). Fuente: autores 4.4.2 Digestibilidad aparente de la proteína. La comparación realizada para la digestibilidad aparente de la proteína cruda no arrojo diferencias significantes (p˃0,05) entre las tres especies de pastos (tabla 8). Estos presentaron valores que variaron entre 41,1 a 45,8 %. Esta baja digestibilidad hallada puede ser atribuida a la presencia de compuestos fenólicos como se evidenció en el bromatológico (tabla 4) y en el análisis de metabolitos secundarios (tabla 7) (Askar et al., 2014), que forman complejos con algunas proteínas de estas gramíneas y las hacen inaccesibles para las proteasas microbianas y componente enzimático del animal (Cong et al., 2013). Esta afirmación fue demostrada a partir del fraccionamiento nitrogenado Cornell, en el cual se halló una gran proporción de fracción C para las tres gramíneas estudiadas. Esta hipótesis fue reconfirmada a partir de la digestibilidad aparente, realizada para las fracciones B3 y C como se aprecia en la tabla 9. Tabla 9. Digestibilidad de las fracciones nitrogenadas B3 y C en las gramíneas B. saccharoides, D. annulatum y B. pertusa. B. saccharoides D. annulatum B. pertusa EEM P- Gramínea Heces Gramínea Heces Gramínea Heces Proteína cruda (%) 6,70 9,47 7,11 7,54 6,72 8,29 ------ ------ Proteína cruda (g) 32,24 17,56 36,34 19.80 37.52 22,12 ------ ------ Fracción B3 (% PC) 1,58 0,00 2,88 0,00 4,09 0,00 ------ ------ Fracción C (% PC) 26,9 46,04 26,31 43,46 28,04 48,31 ------ ------ 1,92 0,1594 Digestibilidad Pc 45,68 45,80 (%) 40 41.19 valor Digestibilidad 100 100 100 0,00 Sd 2,03a 4,58a 2,29a 0,97 0,2016 fracción B3 (%) Digestibilidad Fracción C (%) EEM: error estándar de la media. Columnas que difieren de letra son estadísticamente significativas (p<0,05) Fuente: autores La digestibilidad de la fracción B3, no presentó diferencias estadísticas entre el B. saccharoides, B. pertusa y D. annulatum, como se aprecia en la tabla 9, donde se observa una digestibilidad total de los componentes insolubles en detergente neutro (B3). Sniffen et al., (1992), afirman que está fracción, debido a su disponibilidad en el rumen e intestino, puede tener una digestibilidad del 80%. Como lo describe Alzueta et al., (2001) esta proteína insoluble en detergente neutro es potencialmente degradable y digestible, sin embargo todo depende del tiempo empleado en el rumen, lo cual está relacionado con la tasa de pasaje del alimento (López et al., 1998) ; entre menor sea esta tasa mayor va a ser el tiempo empleado por los microorganismos para la digestión de esta fracción. Dentro de las gramíneas estudiadas, existen dos factores importantes que puede afectar este pasaje, que son el contenido de FDN, el cual por sus componentes retardan el flujo del alimento por el rumen (Riaz et al., 2014) y el bajo consumo de MS, el cual tiene una relación directa con la tasa de pasaje (López et al., 1998). En la tabla 9, también se encuentran citados los valores de proteína cruda, en pasto y heces, evidenciados para cada una de las gramíneas estudiadas. A partir de estos datos, se determinó la digestibilidad de este elemento; de lo cual se puede inferir que la proteína indigestible de los tres pastos en promedio fue superior al 50%. De este 50% aproximadamente el 25% se le atribuye a la fracción C, la cual tuvo un digestibilidad que oscilo entre 2,03% a 4,05% sin presentar diferencias estadísticas (p˃0,05) entre las gramíneas evaluadas (tabla 9), valor que coincide con lo reportado por Sniffen et al., (1992) el cual afirma que la fracción C es casi indigestible para el rumiante. El 25 % restante de proteína indigestible puede ser atribuida, a proteína de tipo endógeno producto 41 de algunos procesos metabólicos y de restitución como lo es el caso de vellosidades intestinales; a otros componentes orgánicos como microorganismos que se encuentran posterior al intestino delgado donde se imposibilita su absorción (Stern et al., 1997). Estos factores mencionados, pueden contribuir a subestimar la proteína del alimento, aparentemente digestible para el caso de las gramíneas suministradas (Stern et al; 1997; Gargallo et al., 2006). 4.4.3 Digestibilidad aparente de la FDN. El pasto B. saccharoides presentó el mayor porcentaje de digestibilidad de la FDN (p˂0,05), con respecto al B. pertusa y D. annulatum (tabla 8). La mayor digestibilidad de FDN puede estar asociada, al mayor contenido de hemicelulosa que posee el B. Saccharoides con respecto al B. pertusa y D. annulatum; característica que es de gran importancia si se considera que este elemento posee una alta degradabilidad en el rumen e influye positivamente en la digestibilidad del resto de componentes de la FDN (Herrick et al., 2012). Entre tanto la hemicelulosa tuvo una alta digestibilidad para los pastos B. saccharoides (91,45%) y B. pertusa (90,62%) en comparación con el D. annulatum, con el cual hubo diferencias significativas (p˂0,05) (tabla 8); patrón que fue similar en la digestibilidad de la celulosa sin embargo para este nutriente las diferencias estadísticas (p˂0,05) se presentaron para el B. saccharoides en relación a las otras dos gramíneas (tabla 8), lo cual es similar a lo reportado por Herrick et al., (2012) sobre la influencia positiva que puede ejercer la hemicelulosa sobre el resto de componentes de la FDN (Viola et al., 2008). Los valores de digestibilidad del EE para los forrajes evaluados fueron B. saccharoides 62,5 %, D. annulatum 76,9 % y B. pertusa 87,8 % (tabla 8), diferencias que demuestran una estrecha relación con los datos de EE obtenidos en el análisis bromatológico descrito así 1.6%, 3,5% y 4,8% respectivamente para las especies de pastos mencionados anteriormente. 42 5. CONCLUSIONES Las gramíneas evaluadas evidenciaron una baja calidad nutricional en comparación con los forrajes que se colectan en el momento indicado, lo cual se asocia al manejo que realizan normalmente los productores de esta región. Los altos contenidos de FDN y FDA evidenciados en los análisis bromatológicos de los pastos evaluados, afectan considerablemente el consumo y la digestibilidad de los henos evaluados. La gramínea B. saccharoides, es la especie que presentó la mayor digestibilidad, lo cual la convierte en una buena opción para alimentación de animales de trópico. La presencia de la fracción C, redujo significativamente la digestibilidad de la proteína de forrajes evaluados, lo cual es un dato interesante y obliga a desviar la atención hacia este componente tan influyente dentro de la digestibilidad de los alimentos Después de haber analizado el consumo de MS y la digestibilidad aparente de las diferentes gramíneas, se concluye, que los animales pueden optimizar el aprovechamiento de nutrientes, cuando estos no tienen un contenido nutricional aceptable que contribuya por lo menos en su mantenimiento. El consumo de MS de los diferentes henos evaluados, está directamente influenciado por la cantidad de FDN; como lo fue evidenciado en el pasto B. saccharoides donde un mayor contenido de celulosa, hemicelulosa y lignina influyó inversamente en el consumo. Las gramíneas analizadas presentaron un contenido moderado de compuestos fenólicos lo cual era lo esperado debido a la edad y etapa de corte de estás. Por otro lado la cantidad de esteroles presentados por los pastos B. pertusa D. annulatum está directamente relacionados con la cantidad de grasa que poseen estas gramíneas. 43 Según los aportes nutricionales y la digestibilidad presentada en este estudio, se concluye que estos pastos no alcanzaron a cumplir con los requerimientos básicos necesitados por los animales. 44 RECOMENDACIONES Es importante seguir evaluando la calidad nutricional y disponibilidad de biomasa de las tres gramíneas (B. Saccharoides, B. pertusa, D. annulatum) en diferentes días de rebrote, para determinar en qué tiempo se puede alcanzar una adecuada producción de biomasa que no altere la calidad nutricional, consumo y digestibilidad de éstas. Es necesario tener presente que en este estudio se analizaron las especies de gramíneas como única base alimenticia, sin embargo, se recomienda realizar trabajos que integren en los tratamientos, leguminosas o suplementos alimenticios que permitan alcanzar los requerimientos nutricionales adecuados. Se debe realizar la determinación de metabolitos secundarios en las diferentes gramíneas por medio de pruebas cuantitativas que permitan obtener mayor relación con los valores de consumo y digestibilidad obtenidos. La información encontrada acerca de la relación de proteína indigestible con fracciones nitrogenadas, específicamente la fracción C, es muy escasa, sin embargo su estimación es muy importante si se quiere obtener un valor más acertado de proteína digestible y de los factores que pueden afectar su utilización en el rumiante. El pasto B. saccharoides, se muestra como una buena opción de pastura a instaurar en las regiones tropicales colombianas; sin embargo son pocos los núcleos de semilla del Pasto Vidal en todo el país, de acuerdo a esto se recomienda crear centros de germoplasma y cultivos de esta especie. Se evidencia la importancia de realizar el pastoreo o periodo de corte de las gramíneas de acuerdo con su ciclo fenológico impidiendo que lleguen a un mayor estado de madurez donde los cambios químicos efectuados disminuyen la calidad nutricional, por ende el consumo y la digestibilidad, aspectos relevantes en la nutrición animal. 45 Se debe tener en cuenta que los animales empleados en experimentos y aún más los ovinos, manejan altos niveles de estrés, lo cual influye en el consumo y digestibilidad de los forrajes, por lo cual se recomienda trabajar en mejorar las condiciones a las que éstos son expuestos. Se requiere avanzar en el uso de estrategias que mejoren la degradabilidad y digestibilidad ruminal, como la rotación y cosecha adecuada de praderas, empleo de microorganismos con mayor acción fibroliticas, el mejoramiento genético, uso de enzimas que modifican el ecosistema ruminal, fertilización, manejo silvopastoril, entre otras alternativas. 46 REFERENCIAS Alzueta, C., Caballero, R., Rebolé, A., Treviño, J., Gil, A., Rebole, A., & Trevin, J. (2001). Crude protein fractions in common vetch ( Vicia sativa L .) fresh forage during pod filling. Journal of Animal Science, 79, 2449–2455. AOAC, & Chemist, A. of O. A. (2000). Official methods of analysis. (A. International, Ed.) (17th ed.). Virginia. Arcos, J., Romero, H., Vanegas, M., & Riveros, E. (s.f.). OVINOS COLOMBIANOS DE PELO. ALTERNATIVA PRODUCTIVA PARA EL SUR DEL DEPARTAMENTO DEL TOLIMA. Askar, a. R., Salama, R., El-Shaer, H. M., Safwat, M. a., Poraei, M., Nassar, M. S., … Raef, O. (2014). Evaluation of the use of arid-area rangelands by grazing sheep: Effect of season and supplementary feeding. Small Ruminant Research, 121(2-3), 262–270. Bach, A., & Calsamiglia, S. (2006). La fibra en los rumiantes ¿ Química o física? FEDNA, 99–114. Barahona, R., & Sánchez, S. (2005). Limitaciones físicas y químicas de la digestibilidad de pastos tropicales y estrategias para aumentarla. Revista Corpoica, 6, 69–82. Bondi, A. (1989). Nutrición animal. (A. S.A., Ed.) (p. 546). Zaragoza. Brautigan, I. (2007). Nutrición Animal (Universisd). Costa Rica. Cardona, M., Sorza, J., Posada, S., Carmona, J., Ayala, S., & Alvarez, O. L. (2002). Establecimiento de una base de datos para la elaboración de tablas de contenido nutricional de alimentos para animales, 15, 240–246. Chamorro, D., Carulla, J., & Cuesta, P. (2005). Cararacterización nutricional de dos asociaciones gramínea - leguminosa en con novillas en pastoreo en el Alto Magdalena. Revista Corpoica, 37–51. Church, D. (1997). Fisiología Digestiva y Nutrición de los Rumiantes Vol.2 - Nutrición. (E. Acribia, Ed.). España. Cong, F., Diehl, B. G., Hill, J. L., Brown, N. R., & Tien, M. (2013). Covalent bond formation between amino acids and lignin: Cross-coupling between proteins and lignin. Phytochemistry, 96, 449–456. 47 Corpoica. (2005). Producción y utilización de recursos forrajeros en sistemas de producción bovina de las regiones Caribe y valles interandinos.de Recursos Forrajeros 1–108. Corpoica, & Universidad Nacional de Colombia. Bothriochloa pertusa (2013). Corpoica, & Universidad Nacional de Colombia. (2013b). Dichanthium annulatum ( Pasto climacuna , Angleton mono ), 8041. Corpoica, & Universidad Nacional de Colombia. (2013c). Dichanthium aristatum ( Pasto Angleton ). Corson, M. S., Rotz, C. A., & Skinner, R. H. (2007). Evaluating warm-season grass production in temperate-region pastures: A simulation approach. Agricultural Systems, 93, 252–268. Cuesta, P., Echeverría, H., Santana, M., & Henriquez, J. (s.f.). IV ESTRATEGIAS DE MANEJO DE PRADERAS PARA MEJORAR LA PRODUCTIVIDAD DE LA GANADERÍA EN LAS REGIONES CARIBE Y VALLES INTERANDINOS. Corpoica.org.co, 1–24. Domínguez, A. (1973). Métodos de Investigación Fitoquímica. (Limusa, Ed.) (pp. 149– 159.). México D.F. Duarte, J. (2011). Comparación de los marcadores internos : fibra en detergente neutro ( FDNi ) y ácido ( FDAi ) indigestibles y colecta total de heces para estimar la digestibilidad en ovinos de pelo. Revista Colombiana de Ciencia Animal, 4(1), 53–60. Estrada, J. (2002). PASTOSY FORRAJES PARA EL TROPICO COLOMBIANO (Universida). Manizales. Febres, O. A. (2008). Factores antinutricionales en los alimentos para ganado vacuno. In Desarrollo sostenible de la ganaderia de doble proposito. (pp. 411–421). Flores, A., Gonzáles, J., Castro, M., Cardelle, B., Fernandez, L., & Alonso, V. (2003). Evaluación de métodos de laboratorio para la predición de la digestibilidad in vivo de la materia orgánica de ensilajes de hierba y planta entera de maíz. Revista PASTOS, 33(5), 3–22. Fukushima, R. S., Kerley, M. S., Ramos, M. H., Porter, J. H., & Kallenbach, R. L. (2015). Comparison of acetyl bromide lignin with acid detergent lignin and Klason lignin and correlation with in vitro forage degradability. Animal Feed Science and Technology, 201, 25–37. 48 Gargallo, S., Calsamiglia, S., & Ferret, a. (2006). Technical note: A modified three-step in vitro procedure to determine intestinal digestion of proteins. Journal of Animal Science, 84, 2163–2167. Herrick, K. J., Hippen, A. R., Kalscheur, K. F., Anderson, J. L., Ranathunga, S. D., Patton, R. S., & Abdullah, M. (2012). Lactation performance and digestibility of forages and diets in dairy cows fed a hemicellulose extract. Journal of Dairy Science, 95(6), 3342– 53. Holdridge, L. R., Grenke W. C., Hatheway W. H., Liang, T., T. J. J. A. (1971). Forest environments in tropical life zones: a pilot study. Forest environments in tropical life zones: a pilot study. Inamdar, A. I., Chaudhary, L. C., Agarwal, N., & Kamra, D. N. (2015). Effect of Madhuca longifolia and Terminalia chebula on methane production and nutrient utilization in buffaloes. Animal Feed Science and Technology, 201, 38–45. Jampeetong, A., Brix, H., & Kantawanichkul, S. (2014). Effects of inorganic nitrogen form on growth, morphology, N uptake, and nutrient allocation in hybrid Napier grass (Pennisetum purpureum × Pennisetum americanum cv. Pakchong1). Ecological Engineering, 73, 653–658. Jung, H. G., Mertens, D. R., & Payne, A. J. (1997). Correlation of acid detergent lignin and Klason lignin with digestibility of forage dry matter and neutral detergent fiber. Journal of Dairy Science, 80(8), 1622–8. Kaul, V. K., & Vats, S. K. (1998). Essential oil composition of Bothriochloa pertusa and phyletic relationship in aromatic grasses. Biochemical Systematics and Ecology, 26, 347–356. Krämer, M., Weisbjerg, M. R., Lund, P., Jensen, C. S., & Pedersen, M. G. (2012). Estimation of indigestible NDF in forages and concentrates from cell wall composition. Animal Feed Science and Technology, 177(1-2), 40–51. Licitra, G., Hernandez, T. M., & Van Soest, P. J. (1996). Standardization of procedures for nitrogen fractionation of ruminant feeds. Animal Feed Science and Technology, 57(4), 347–358. López, A., Maiztegui, J., & Cabrera, R. (1998). Voluntary intake and digestibility of forages with different nutritional quality in alpacas (Lama pacos). Small Ruminant Research, 29, 295–301. Maloiy, G. M. O., Kanui, T. I., Towett, P. K., Wambugu, S. N., Miaron, J. O., & Wanyoike, M. M. (2008). Effects of dehydration and heat stress on food intake and dry matter digestibility in East African ruminants. Comparative Biochemistry and Physiology. Part A, Molecular & Integrative Physiology, 151(2), 185–90. 49 Marai, I. F. M., El-Darawany, a. a., Fadiel, a., & Abdel-Hafez, M. a M. (2007). Physiological traits as affected by heat stress in sheep-A review. Small Ruminant Research, 71, 1– 12. McDonald, P., Edwards, R., Greenhalgh, J., & Morgan, C. (2006). Nutrición animal. (E. Acribia, Ed.) (Sexta, pp. 395–396). McLeod, M. N. (1974). Plant tannins-their role in forage quality. Nutr. Abstr and Rev, 44, 803–815. Meirelles, P., Luiz, R., Batista, G., Lempp, B., & Costa, C. (2005). QUANTIFICAÇÃO E DISTIBUIÇÃO DE TANINOS EM GRAMÍNEAS FORRAGEIRAS TROPICAIS. Reunião Anual Da Sociedade Brasileira de Zootecnia, 5–8. Misra, A. K., Mishra, A. S., Tripathi, M. K., Chaturvedi, O. H., Vaithiyanathan, S., Prasad, R., & Jakhmola, R. C. (2006). Intake, digestion and microbial protein synthesis in sheep on hay supplemented with prickly pear cactus [Opuntia ficus-indica (L.) Mill.] with or without groundnut meal. Small Ruminant Research, 63(1-2), 125–134. National Research Council-NRC. (2001). Nutrient Requirements of Dairy Cattle. (7th ed.). Revisada. Washington, D. C. USA. Ngwa, A. T., Nsahlai, I. V, & Iji, P. A. (2002). Effect of supplementing veld hay with a dry meal or silage from pods of Acacia sieberiana with or without wheat bran on voluntary intake , digestibility , excretion of purine derivatives , nitrogen utilization , and weight gain in South African Merino she, 77, 253–264. Patra, A. K., & Yu, Z. (2014). Combinations of nitrate, saponin, and sulfate additively reduce methane production by rumen cultures in vitro while not adversely affecting feed digestion, fermentation or microbial communities. Bioresource Technology, 155, 129–135. Piñeros, R., Delgado, J., & Holguín, V. (2011). Respuesta del pasto Bothriochloa saccharoides a diferentes intensidades de sombra simulada en el valle cálido del Magdalena en el Tolima (Colombia). Revista CORPOICA. Ciencia Y …, 12, 42–50. Piñeros, R., Silva, K., Sánchez, M., & Delgado, J. (2010). Calidad nutricional del pasto Vidal ( Bothriochloa saccharoides ) bajo condiciones del trópico seco Nutritional quality of Vidal grass ( Bothriochloa saccharoides ) under dry tropical conditions, 3(1), 50–55. Piñeros, R., Silva, K., Sánchez, M. I., Delgado, J., & Holguín, V. A. (2009). INDICADORES AGRONÓMICOS DEL PASTO VIDAL (BOTHRIOCHLOA SACCHAROIDES) BAJO SOMBRA SIMULADA EN EL VALLE CÁLIDO DEL MAGDALENA, TOLIMA (COLOMBIA). Revista Luna Azul, (29), 32–36. 50 Posada, R. (2011). Comparación de dos metodos in vitro para estimar la digestibilidad de pastos tropicales en rumiantes. CITESCA, 2(2), 13–25. Ramírez, R. (2005). NURICIÓN DE RUMIANTES. (Trillas, Ed.) (Primera, pp. 131–132). México. Relling, E., & Mattioli, G. (2003). FISIOLOGIA DIGESTIVA Y METABOLICA DE LOS RUMIANTES (EDULP, pp. 1–72). La Plata. Riaz, M. Q., Südekum, K.-H., Clauss, M., & Jayanegara, a. (2014). Voluntary feed intake and digestibility of four domestic ruminant species as influenced by dietary constituents: A meta-analysis. Livestock Science, 1–10. Ruiz, R. (2011). Comparación de dos metodos in vitro para determinar la digestibilidad de pastos tropicales. Revista Citecsa, 13–25. Sánchez, D., Arreaza, L., & Abadía, B. (2005). Estudio de la cinética de degradación in vitrode cuatro forrajes tropicales y una leguminosa de clima templado, 6, 58–68. Sánchez, L., Wills, A., & Figueroa, J. (2014). Efecto de la población microbial sobre las pruebas de digestibilidad in vitro e in situ. Revista de Facultad de Medicina Veterinaria Y de Zootecnia Universidad Nacional ., 46(2), 16–20. Särkijärvi, S., Sormunen, R., Heikkilä, T., Rinne, M., & Saastamoinen, M. (2012). Effect of grass species and cutting time on in vivo digestibility of silage by horses and sheep. Livestock Science, 144(3), 230–239. Shimada, A. (2005). Nutrición animal. (Trillas, Ed.) (Primera, p. 64). México. Sierra, O., Bedoya, J., Monsalve, D., & Orozco; J. (1988). Observaciones sobre Colosua (Bothiochloa, Pertusa (L). Camus)mnen la Costa Atlantica de Colombia. Sniffen, C. J., O’Connor, J. D., Van Soest, P. J., Fox, D. G., & Russell, J. (1992). A net carbohydrate and protein system for evaluating cattle diets.1. Ruminal fermentation. J. Anim. Sci., 70, 3551–3561. Stern, M. D., Bach, A., & Calsamiglia, S. (1997). Alternative Techniques for Measuring Nutrient Digestion in Rumi. Journal of Animal Science, 75, 2256–2276. Valencia, L., Restrepo, J., Cerón, E., & Herrera, W. (2010). utilizando dietas a base de forrajes tropicales. Revista de Investigación Agraria Y Ambiental, 1(1), 25–29. Van soest, P. (1994). NUTRITIONAL ECOLOGY OF THE RUMINANT (2nd ed.). New York. 51 Van Soest, P., Robertson, J. B., & Lewis, B. A. (1991). Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. Journal of Dairy Science, 74(10), 3583–97. Vanegas, M., & Riveros, E. (1999). Producción de semilla del pasto Angleton climacuna (Dichanthium annulatum)en el alto Magdalena. Corpoica Org, 1–6. Vargas, A. (2012). Cinética de biohidrogenación in vitro de ácidos grasos poliinsaturados en fluido ruminal. Universidad Nacional de Colombia. Varón, R. P., Tobar, V., & Mora, J. (2011). Evaluación agronómica y zootécnica del pasto Colosoana ( Bothriochloa pertusa ) en el trópico seco del Tolima . Agronomic and zootechnical assessment of Colosoana grass ( Bothriochloa pertusa ) in dry tropics of Tolima Resumen Indicadores Agronómicos Indi, 4(1), 36–40. Vidal, H., Mora Delgado, J., & Vanegas, M. (2007). El pasto Vidal: nace hasta en las piedras. Revista de FEDEGAN. Vieira, P. A. S., Pereira, L. G. R., Azevêdo, J. A. G., Neves, A. L. A., Chizzotti, M. L., dos Santos, R. D., … Chaves, A. V. (2013). Development of mathematical models to predict dry matter intake in feedlot Santa Ines rams. Small Ruminant Research, 112(13), 78–84. Viola, E., Zimbardi, F., Cardinale, M., Cardinale, G., Braccio, G., & Gambacorta, E. (2008). Processing cereal straws by steam explosion in a pilot plant to enhance digestibility in ruminants. Bioresource Technology, 99, 681–689. 52