TEJIDO MUSCULAR
Anuncio

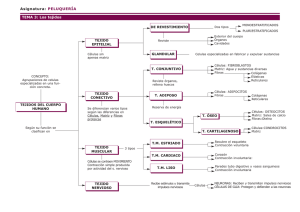
Tejido muscular: Concepto, generalidades y tipos. Tejido muscular estriado esquelético: concepto y generalidades. Fibra muscular estriada. Tipos de fibras. Tejido muscular estriado cardiaco: concepto y generalidades. Fibra muscular cardiaca. Tejido de conducción cardiaco: fibra de Purkinje. Tejido muscular liso: concepto y generalidades. Fibra muscular lisa. El tejido muscular es un tejido muy especializado constituido por células que contienen gran cantidad de proteínas contráctiles que le dan la capacidad de contraerse de forma coordinada en una determinada dirección para producir un movimiento. Las células musculares reciben el nombre de fibras musculares o miofibras, y se disponen de forma paralela constituyendo fascículos de fibras orientadas en el sentido de la contracción. Se diferencian tres tipos de tejido muscular atendiendo a las características morfológicas y funcionales así como a la disposición de sus células: • Tejido muscular estriado esquelético: forman parte del sistema musculo esquelético y de ciertos órganos como lengua y globo ocular. Sus fibras se caracterizan por presentar una estriación transversal en su citoplasma. Se trata de un tejido muscular de contracción voluntaria. • Tejido muscular estriado cardiaco: constituye la musculatura del corazón. Sus fibras presentan en su citoplasma una estriación transversal. Se trata de un tejido muscular de contracción involuntaria y rítmica. • Tejido muscular liso: se encuentra en diversas localizaciones, entre otras, forma parte de las vísceras y los vasos sanguíneos. Sus fibras no presentan estriación transversal en su citoplasma. Se trata de un tejido muscular de contracción involuntaria. En general, cuando se estudia el tejido muscular, algunas de las estructuras que constituyen las fibras reciben un nombre especial, precedido siempre del prefijo sarco- (carne en griego), así, la membrana plasmática se denomina sarcolema, el citoplasma sarcoplasma, el retículo endoplásmico retículo sarcoplásmico y la mitocondria sarcosoma. TEJIDO MUSCULAR ESTRIADO ESQUELÉTICO Está constituido por fibras de morfología cilíndrica con un grosor que varía entre 10-100 µm de diámetro y con una longitud que puede llegar a ser tan larga como los músculos de los que forman parte. En el tamaño de las fibras musculares influirá la edad, el ejercicio, el estado nutricional, el sexo y la posición de cada fibra dentro del músculo. Cada fibra muscular se encuentra rodeada por una delgada capa de tejido conectivo reticular que contiene numerosos capilares y fibras nerviosas y denominada endomisio. Las fibras musculares se agrupan paralelamente en fascículos cuyo grosor varía según la actividad muscular y cada fascículo está rodeado por septos de tejido conectivo con abundantes fibras de colágeno y que también presenta abundantes capilares y fibras nerviosas y que se denomina perimisio. Los fascículos se agrupan para formar los músculos y se encuentran rodeados por una banda de tejido conectivo denso irregular con abundantes fibras de colágeno tipo I y II y fibras elásticas que se denomina epimisio, que se continua con los tendones e inserciones musculares. A través del epimisio penetran las arterias y los nervios en los músculos, ramificándose por el perimisio y endomisio para llevar el riego y la inervación a las fibras musculares. El origen de las fibras musculares estriadas esqueléticas está en la fusión de numerosos mioblastos, que son células inmaduras procedentes del mesodermo y que presentan morfología fusiforme. La morfología de las fibras musculares estriadas esqueléticas en corte transversal en fresco es redondeada, pero cuando se fijan la aparecen de morfología poligonal. Los núcleos son ovalados y se encuentran periféricamente, alineados bajo el sarcolema. En las proximidades de las uniones tendinosas es normal encontrar algunos núcleos en el interior del sarcoplasma. El sarcolema se encuentra delimitado periféricamente por una lámina basal y presenta invaginaciones denominadas túbulos T, cada uno de los cuales se encuentra asociado a dos cisternas terminales del retículo sarcoplásmico liso, constituyendo una estructura denominada tríada. Estas estructuras presentan un papel importante en la contracción muscular, permitiendo que la despolarización del sarcolema llegue a las regiones profundas del sarcoplasma, induciendo así la salida del calcio de las cisternas terminales del retículo sarcoplásmico liso, iniciándose así el mecanismo de contracción. Las triadas se disponen abrazando a los haces de miofibrillas a intervalos regulares, a nivel en las transiciones entre las bandas A e I. El sarcolema presenta caveolas que se corresponden con vesículas de pinocitosis. También presenta unas depresiones donde el botón terminal de la neurona realiza la sinapsis nerviosa con las fibras musculares, constituyendo las unidades motoras, y otras depresiones donde se localizan las células satélite. Las células satélite presentan un núcleo ovalado y un citoplasma escaso donde aparecen unas pocas mitocondrias y un escaso sistema tubular y se caracterizan porque tienen la capacidad de dividirse y fusionarse a las fibras musculares para que aumenten su grosor y porque son capaces de regenerar a las fibras musculares dañadas, siempre y cuando estas no presentan dañada la lámina externa. Las fibras regeneradas se diferencian de las normales porque presentan los núcleos en el centro en vez de en la periferia. La población de células satélite es elevada en los animales jóvenes y va disminuyendo con la edad. El sarcoplasma presenta una tinción acidófila intensa y contiene como organoides más importantes el retículo sarcoplásmico, abundantes sarcosomas y gránulos β de glucógeno, que son la fuente de energía para llevar a cabo el mecanismo de contracción muscular. Además, también pueden observase veces vacuolas de lípidos que pueden ser utilizadas también como fuente de energía y pequeños aparatos de Golgi situados junto a cada núcleo. Dentro del sarcoplasma también podemos encontrar proteínas no contráctiles como albumina y moléculas de mioglóbina que son las que le proporcionan al músculo el color rojo característico. La función de estas moléculas de mioglobina almacenar oxígeno que será utilizado en la glucólisis anaerobia para la obtención de energía durante la contracción muscular. También podemos encontrar sales minerales. La mayor parte del sarcoplasma (60-70% del volumen) está ocupado por las miofibrillas, cuya medida es de 1-2 µm de diámetro. En cortes longitudinales las miofibrillas presentan estriaciones transversales muy refringentes y que se tiñen fuertemente con la hematoxilina férrica. En cortes transversales se agrupan en áreas poligonales denominadas campos de Conheim, que se consideran un artefacto de la contracción más que una estructura funcional. Estas miofibrillas están constituidas por proteínas contráctiles: actina y miosina. La actina forma parte de los miofilamentos delgados (unos 5-6 nm de grosor) y la miosina de los miofilamentos gruesos (unos 15 nm de grosor). La ordenación de los miofilamentos en el músculo esquelético y en el cardiaco da lugar a la aparición de las características estriaciones transversales. Cuando se observan las fibras musculares con un microscopio de luz polarizada aparece una estriación con bandas anisótropas, muy refringentes que se denominan bandas A, y bandas isótropas, sin refringencia denominadas bandas I. Con el microscopio electrónico, la banda A es electro densa y está compuesta por miofilamentos gruesos y delgados, mientras que la banda I es poco electro densa y está compuesta solamente por miofilamentos delgados. En el centro de la banda I aparece una línea electro densa que se denomina línea Z y que se corresponde con el lugar donde se fijan los miofilamentos delgados mediante una proteína denominada α-actinina. La estructura que está comprendida entre dos líneas Z se denomina sarcómero y que se corresponde con la unidad de contracción muscular. En el centro de la banda A aparece una banda más estrecha y menos electro densa denominada banda H o de Héller, que está constituida solamente por miofilamentos gruesos. En el centro de la banda H aparece otra línea electro densa denominada línea M o Mesofragma, que es el lugar donde se anclan los miofilamentos gruesos a través de proteínas. En un corte transversal del sarcómero: • Línea M: está constituida por filamentos gruesos de miosina unidos entre ellos por filamentos M, unidos por puentes M. • Banda H: está compuesta solamente por filamentos gruesos. • Banda A: cada filamento grueso está rodeado por 6 delgados. • Banda I: aparecen solamente filamentos delgados que se disponen en los vértices de un hexágono. • Línea Z: cada filamento de actina se divide en cuatro filamentos, por lo que presenta el aspecto de un entramado reticular. Los filamentos gruesos presentan una forma como de palo de golf de unos 15nm de diámetro y 1,5 µm de longitud, y está formada por unas 200 moléculas de miosina. La cabeza esta representada por la meromiosina pesada y la cola por la meromiosina ligera. Para constituir los miofilamentos gruesos, se disponen 6 filamentos de miosina con las cabezas dispuestas en un giro completo de 360º y alrededor de los mismos se dispone una proteína elástica denominada titina, cuya función es la de mantener la posición de las moléculas de miosina, impedir el sobre estiramiento de los sarcómeros y permitir que vuelvan a su posición inicial tras la contracción. Los filamentos delgados están formando dos por moléculas de actina fibrosa o F constituida por moléculas de actina globular o G que polimerizan formando una doble hélice. Cada molécula de actina G presenta un sitio activo de unión con la miosina. A lo largo de la doble hélice de actina aparece una proteína fibrilar denominada tropomiosina, que en la situación de reposo aparece tapando los lugares activos de unión con la miosina. A intervalos regulares también aparece una proteína globular denominada troponina, constituida por tres poli péptidos globulares, el Tn.T, que se une a la tropomiosina, el Tn.C, que presenta gran afinidad por el calcio, y el Tn.I que inhibe la unión actina-miosina. Otra proteína denominada nebulina, es la que ayuda a mantener la disposición de la actina en el sarcómero. Según la estructura y características morfológicas y funcionales se pueden diferenciar varios tipos de fibras musculares, que se organizan en unidades motoras que presentan diferentes tipos de motoneuronas y un número variable de miofibrillas (10-100). Los somas están en el sistema nervioso central (SNC) y los axones llegan para formar sinapsis con el músculo: • Unidad motora S o lenta: está constituida por fibras musculares tipo I o intermedias, que son fibras musculares que presentan un contenido de mioglobina intermedio, son de contracción lenta y sostenida durante largos periodos de tiempo y son fibras no fatigables. Este tipo de fibras son ricas en mitocondrias, presentan una línea Z gruesa y un escaso REL. • Unidad motora FR: es resistente a la fatiga. Está constituida por fibras tipo IIA o rojas, ya que presentan un alto contenido en mioglobina, y son de contracción rápida. Estas fibras son ricas en mitocondrias, su línea Z es gruesa y escaso REL. • Unidad motora FF: es fácilmente fatigable. Está constituida por fibras tipo IIB o blancas, ya que presentan un bajo contenido en mioglobina, son de contracción rápida e intervienen en movimientos de fuerza rápidos. Estas fibras presentan escasas mitocondrias, su línea Z es delgada y tienen muy desarrollado del retículo sarcoplásmico. TEJIDO MUSCULAR ESTRIADO CARDIACO Está constituido por fibras musculares estriadas de morfología cilíndrica de unos 80-100 µm de longitud y unos 5-15 µm de diámetro, frecuentemente ramificadas y anastomosadas con otras por medio de uniones o bandas escaleriformes, visibles con el microscopio óptico, especialmente con la tinción de hematoxilina férrica. Estas fibras musculares son de contracción involuntaria y rítmica, a modo de sincitios y podemos diferenciar dos tipos de fibras: contráctiles y conductoras. Fibras contráctiles Las fibras musculares cardiacas son cilíndricas y frecuentemente ramificadas y aparecen dispuestas formando fascículos paralelos rodeados por tejido conectivo laxo rico en vasos sanguíneos. Las células aparecen rodeadas por un tejido conectivo laxo reticular rico en capilares sanguíneos, pero carecen de células satélite, por lo que estas fibras musculares no pueden regenerarse y cuando se destruyen son sustituidas por tejido conectivo. Al igual que las fibras musculares esqueléticas, tienen una estriación transversal, pero solamente presentan 1 ó 2 núcleos ovalados y vesiculosos de posición central con 1 ó 2 nucléolos. En un corte transversal las fibras aparecen con morfología redondeada con el núcleo situado en el centro de la fibra. El sarcolema también presenta invaginaciones (túbulos T), de mayor diámetro que en el músculo estriado y con invaginaciones de la lámina basal que sólo aparecen asociadas a una cisterna del retículo sarcoplásmico liso, constituyendo díadas, que presentan una orientación transversal y longitudinal, constituyendo el sistema tubular transversal axial. Dentro del sarcoplasma podemos encontrar numerosos sarcosomas de pequeño tamaño que ocupan casi la mitad del volumen citoplasmático, también aparecen abundantes gránulos de glucógeno y pequeñas gotas con lípidos. Las fibras musculares cardiacas de la aurícula derecha y en menor medida las de la izquierda contienen gránulos de secreción electro densos esféricos de unos 0,30,4 µm de diámetro que contienen el factor natriurético auricular, que es una hormona que se vierte a la sangre y actúa regulando el volumen sanguíneo y disminuyendo la presión arterial. La constitución de los miofilamentos es similar al de las fibras musculares estriadas esqueléticas, diferenciándose solamente en las líneas Z, que se localizan en las bandas escaleriformes y parece que cruzan de una célula a otra. Las bandas escaleriformes, también llamadas discos intercalares, en un corte transversal aparecen constituidas por uniones tipo adherens (fascia adherens y mácula adherens) y en las caras laterales presentan uniones tipo Gap. La función de estas bandas es la de transmitir la onda de despolarización para conseguir que todas las fibras musculares cardiacas se contraigan al mismo tiempo. Fibras conductoras Estas fibras se encargan de la conducción de los impulsos nerviosos y se localizan en los nódulos sinoaruricular y auriculoventricular y se continúan por el haz auriculoventricular o fascículo de His. Presentan escasa cantidad de miofilamentos y gran cantidad de gránulos de glucógeno. Su citoplasma es pálido y poco coloreado. Carecen de túbulos T y de discos intercalares, presentando solamente pequeños desmosomas. Podemos diferenciar tres tipos de fibras conductoras: • Fibras nodales: presentan pequeño tamaño (5-10 µm de diámetro) y se localizan en los nódulos. • Fibras fasciculares o de Purkinje: son más grandes (30-50 µm de diámetro) e irregulares. • Fibras transicionales: tienen características intermedias y transitan por el tejido conectivo subendocárdico. TEJIDO MUSCULAR LISO Está constituido por células fusiformes mono nucleares de contracción involuntaria (a excepción de la vejiga de la orina), lenta y sostenida. Aparece en numerosas localizaciones, como formando parte de las vísceras de los aparatos digestivo, respiratorio y genitourinario, paredes de arterias, venas y vasos linfáticos, dermis, músculo erector del pelo, cápsula y tabiques esplénicos, ligamento ancho y pleura visceral. Las fibras musculares lisas están delimitadas por una lámina basal, tienen un tamaño que varía entre unos 20-500 µm de longitud y unos 5-20 µm de diámetro (las dimensiones mayores se pueden observar en el útero grávido). El núcleo es central, con morfología fusiforme, presenta uno ó dos nucléolos y aparece durante la contracción a modo de sacacorchos. No presentan túbulos T, pero si tienen cavéolas que contienen iones calcio. El sarcoplasma es acidófilo y con escasos organoides localizados alrededor del núcleo (Golgi, sarcosomas, glúcogeno y retículo sarcoplásmico rugoso y liso). Carecen de estriación transversal en los cortes longitudinales. Entre células adyacentes presentan uniones tipo Gap. Los miofilamentos se disponen formando fascículos, a manera de espiral, de unos 300 nm de diámetro. Los filamentos de actina son los más numerosos, no presentan troponina y en algunas localizaciones también carecen de tropomiosina. Se agrupan 15 filamentos de actina por cada uno de miosina y se fijan al sarcolema por medio de placas de unión o placas de anclaje, y en el citoplasma forman los cuerpos densos, actuando de un modo similar a la línea Z del músculo. Durante la contracción muscular provocan la invaginación del sarcolema. Los filamentos de miosina están constituidos por meromiosina pesada en toda su extensión, se disponen al azar y en un corte transversal ocupan los vértices de un rectángulo. Los filamentos intermedios ocupan el centro y la periferia de la fibra muscular lisa, alrededor de las placas de anclaje y cuerpos densos. Estos filamentos son de vimentina y desmina en los vasos sanguíneos y de desmina en el resto de localizaciones. Las fibras musculares lisas tiene capacidad de división, por lo que pueden regenerarse y, en algunos casos, pueden generarse a partir de la diferenciación de los pericitos.