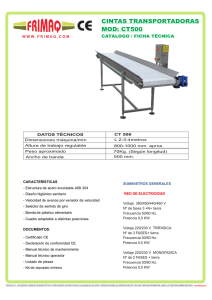
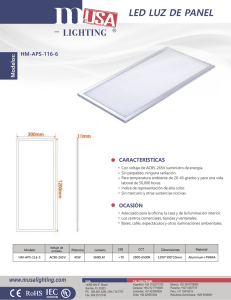
universidad de chile facultad de ciencias físicas y
Anuncio

UNIVERSIDAD DE CHILE FACULTAD DE CIENCIAS FÍSICAS Y MATEMÁTICAS DEPARTAMENTO DE INGENIERÍA ELÉCTRICA ANÁLISIS Y CONTROL DE UN MODELO DINÁMICO DE UNA NEURONA TALÁMICA MEMORIA PARA OPTAR AL TITULO DE INGENIERO CIVIL ELECTRICISTA CHRIST ALEJANDRA DEVIA MANRIQUEZ PROFESOR GUÍA: MANUEL DUARTE MERMOUD MIEMBROS DE LA COMISIÓN: HECTOR AGUSTO ALEGRIA MARÍA DE LA LUZ AYLWIN OSTALE SANTIAGO DE CHILE OCTUBRE 2007 RESUMEN DE LA MEMORIA PARA OPTAR AL TÍTULO DE INGENIERO CIVIL ELECTRICISTA POR: CHRIST DEVIA MANRIQUEZ FECHA: 18 DE OCTUBRE DE 2007 PROF. GUÍA: SR. MANUEL DUARTE MERMOUD “ANÁLISIS Y CONTROL DE UN MODELO DINÁMICO DE UNA NEURONA TALÁMICA” Actualmente existen dispositivos de estimulación cerebral intracraneal, aplicados principalmente a pacientes con desordenes de movimiento. Estos dispositivos estimulan con una corriente eléctrica débil, grupos de neuronas en regiones específicas del cerebro, como el núcleo subtalámico y Globo Pálido. Los parámetros del estímulo aplicado son sintonizados por el médico tratante a lo largo del tiempo, considerando como meta detener o minimizar los síntomas, sin causar con este proceso trastornos físicos o conductuales al paciente. En ese contexto, este Trabajo de Título hace una abstracción del problema y propone que la sintonización de los parámetros de estimulación se realice en forma automática, considerando que el grupo de neuronas que se desea controlar debería descargar (responder) de igual modo que poblaciones de neuronas o modelos de referencia. El objetivo general de la memoria es diseñar y aplicar una estrategia de control, que permita que la neurona sobre la cual se aplica la acción de control, descargue en el mismo modo que otra neurona de referencia, independiente del estímulo que reciba. Se logrará así un esquema denominado de seguimiento neuronal. Para ello se estudió y determinó, mediante simulaciones, las características de entrada/salida de un modelo dinámico simple de una neurona talámica. Esto permitió determinar cuáles son las características que permiten clasificar su modo de repuesta y estimar, a partir de la salida, la magnitud del estímulo que recibe. Basados en el estudio anterior se diseñó un sistema de control que genera una corriente apropiada, considerando que se tiene acceso sólo al voltaje de membrana de la neurona. De esta manera el esquema de control propuesto consigue el objetivo de identificar el modo de descarga de una neurona y modificarlo en caso de ser necesario, generando una acción de control permanente en presencia de estímulos del tipo constante, sinusoidales y simuladores de corrientes sinápticas. Se ha logrado entonces en esta memoria diseñar un sistema de control que, a partir de las señales de voltaje de membrana del modelo de dos neuronas, identifique su modo de respuesta y defina la acción de control necesaria aplicar para que ambas estén en igual modo de repuesta. i Agradecimientos En primer lugar a mi Tomi por todo su apoyo. Por estar ahí siempre. A toda toda mi familia, a mi Mamá, Papá, Fany, Javier, Lin y Dieguito, en especial a la Pa, al Tipi y al Manolo, quienes me brindaron su ayuda de forma incondicional. Al tío Ivan por, prácticamente, adoptarme en su casa. A Hardy por almorzar conmigo todo el Verano y mostrarme que pocas cosas merecen mi estrés. A la Marce, Pedro y Fabián, por ayudarme, a veces sin darse cuenta, a encontrar la solución a varios de mis problemas. Al Profe Javier por su paciencia. A Francisco de Borja Rodriguez y Richard Adams por su interés y amabilidad. Gregory Smith y Jeff Groff por la buena disposición para contestar mis preguntas sobre el modelo. Finalmente, al Profe Manuel y a la Profe Marilú por su ayuda, tiempo y atención, muchísimas gracias. Sin todos los aquí mencionados, nada habría sido lo mismo. Muchas gracias por todo. ii Índice general Agradecimientos ii Índice general 1 Índice de figuras 3 Capítulo 1 6 Introducción 1.1. Estimulación Cerebral Profunda .....................................................................................6 1.2. Objetivos.............................................................................................................................9 1.3. Estructura de la memoria ...............................................................................................11 Capítulo 2 Neuronas y Modelos 12 2.1. Características y estructura de las neuronas.................................................................12 2.2. Modelos neuronales ........................................................................................................18 2.3. Neuronas talámicas y modelos ......................................................................................24 Capítulo 3 Análisis del Modelo IFB 38 3.1. Análisis dinámico.............................................................................................................43 3.2. Características de la respuesta........................................................................................48 Capítulo 4 Diseño y Aplicación de la Estrategia de Control 66 4.1. Diseño de la estrategia de control .................................................................................67 4.2. Resultado Simulaciones ..................................................................................................73 4.3. Robustez del controlador ...............................................................................................90 4.4. Control realimentado ......................................................................................................96 1 Capítulo 5 Conclusiones y Trabajo Futuro Referencias 101 105 Anexo A Modelo IFB 113 Anexo B Sistema de control lazo abierto 115 Anexo C Sistema de control realimentado 118 2 Índice de figuras Figura 1.1 Esquema del dispositivo de DBS implantado. Adaptada de [ 3]..................................7 Figura 1.2 Formas de onda en terapias de electroestimulación. Adaptada de [ 7]........................8 Figura 1.3 Áreas de estimulación en la terapia DBS. Adaptada de [ 6]. .........................................9 Figura 2.1 Esquema de neurona motora (A) y neurona sensitiva (B). Adaptada de [ 14]. ....... 13 Figura 2.2 Esquema de distintos tipos de neuronas. Adaptada de [ 11]. .................................... 13 Figura 2.3 Esquema de la membrana neuronal. Adaptada de [ 11]. ............................................ 14 Figura 2.4 Formas de onda del potencial de membrana. .............................................................. 16 Figura 2.5 Esquema de transmisión. Adaptada de [ 11]. ............................................................... 16 Figura 2.6 Esquema de sinapsis química. Adaptada de [ 11]. ....................................................... 17 Figura 2.7 Potencial presináptico, corriente presináptica y potencial postsinápco. .................. 18 Figura 2.8. Análogo eléctrico de una sección de membrana. Adaptada de [ 15]. ...................... 19 Figura 2.9 Gráfica de las variables del modelo HH. ...................................................................... 23 Figura 2.10 Fases de un potencial de acción. .................................................................................. 24 Figura 2.11 Vista sagital del encéfalo adulto. Adaptada de [ 11]................................................ 25 Figura 2.12 Imagen del tálamo y del núclo lateral geniculado. Adaptada de [ 12]..................... 26 Figura 2.13 Neurona talámica. Adaptada de [ 13]. ......................................................................... 26 Figura 2.14 Respuesta experimental de una TRN.......................................................................... 28 Figura 2.15 Actividad oscilatoria del voltaje de membrana, burst. Adaptada de [ 22].............. 29 Figura 2.16 Resultados obtenidos con el modelo de HM............................................................. 33 Figura 2.17 Modelo multicompartimiento de McIntyre. Adaptada de [ 9]................................. 35 Figura 3.1 Comparación de distintos modelos. .............................................................................. 39 Figura 3.2 Idealización de m∞ y h. Adaptada de [ 12]. ................................................................... 40 Figura 3.3 Verificación de resultados del modelo IFB. ................................................................. 42 3 Figura 3.4 Tipos de repuestas modelo IFB. Adaptada de [ 59].................................................... 43 Figura 3.5 Variables en respuesta burst. .......................................................................................... 44 Figura 3.6 Variables en respuesta tónica.......................................................................................... 45 Figura 3.7 Sincronización de la variables en descargas burst........................................................ 46 Figura 3.8 Diagrama de fase modelo IFB........................................................................................ 48 Figura 3.9 Descarga en modo burst. ................................................................................................ 49 Figura 3.10 Corriente de reobase, modo tónico. ............................................................................ 50 Figura 3.11 Corriente de cronaxia, modo tónico............................................................................ 50 Figura 3.12 Corrientes aplicadas en modo tónico. ......................................................................... 51 Figura 3.13 Corriente de reobase, modo burst. .............................................................................. 51 Figura 3.14 Corriente de cronaxia, modo burst.............................................................................. 52 Figura 3.15 Corrientes aplicadas en modo burst. ........................................................................... 52 Figura 3.16 Relación entre respuesta y frecuencia de estímulo. ................................................... 54 Figura 3.17 Repuesta tónica ante AC. .............................................................................................. 55 Figura 3.18 Repuesta tónica ante AC y DC. ................................................................................... 56 Figura 3.19 Respuesta burst ante AC. .............................................................................................. 57 Figura 3.20 Corriente sináptica. ........................................................................................................ 58 Figura 3.21 Proceso de llegadas de Poisson. ................................................................................... 59 Figura 3.22 Corriente de estímulo. ................................................................................................... 59 Figura 3.23 Respuesta tónica frente a entrada sináptica sintética. ............................................... 60 Figura 3.24 Respuesta tónica de frecuencia irregular..................................................................... 60 Figura 3.25 Respuesta burst frente a entrada sináptica sintética. ................................................. 61 Figura 3.26 Repuesta tónica entrada de alta frecuencia................................................................. 62 Figura 3.27 Respuesta modo burst ante entrada de alta frecuencia............................................. 63 Figura 3.28 Respuesta burst ante entrada de alta frecuencia. ....................................................... 64 Figura 3.29 Relación frecuencia de estímulo frecuencia de descarga. ......................................... 65 Figura 4.1 Esquema del sistema de control propuesto.................................................................. 67 Figura 4.2 Registro extracelular de descargas. Adaptada de [ 61]. ............................................... 68 Figura 4.3 Codificación en frecuencia.............................................................................................. 70 Figura 4.4 Relación frecuencia amplitud, modo tónico................................................................. 71 Figura 4.5 Relación frecuencia amplitud de estímulo. ................................................................... 72 Figura 4.6 Respuesta en modo burst para varias amplitudes de estímulo. ................................. 72 Figura 4.7 Relación dV/dt y amplitud, modo burst....................................................................... 73 4 Figura 4.8 Ejemplo de corrientes de estímulo DC......................................................................... 74 Figura 4.9 Voltaje de membrana en lazo abierto............................................................................ 74 Figura 4.10 Control de modo tónico a burst, estímulo DC.......................................................... 75 Figura 4.11 Voltaje de membrana en lazo abierto.......................................................................... 76 Figura 4.12 Control de modo burst a tónico, estímulo DC.......................................................... 77 Figura 4.13 Ejemplo de casos en que no se genera acción de control, estímulo DC. .............. 79 Figura 4.14 Estímulos sinusoidales aplicados. ................................................................................ 80 Figura 4.15 Repuesta en lazo abierto, estímulo AC. ...................................................................... 80 Figura 4.16 Control del modo tónico al burst, estímulo AC. ....................................................... 81 Figura 4.17 Control del modo burst al tónico, estímulo AC. ....................................................... 82 Figura 4.18 Ejemplo de casos en que no se genera acción de contro, estímulo AC................. 83 Figura 4.19 Control del modo tónico a burst, estímulo ISS. ........................................................ 86 Figura 4.20 Control del modo burst a tónico, estímulo ISS. ........................................................ 87 Figura 4.21 Ejemplos donde no controló, estímulo ISS. .............................................................. 89 Figura 4.22 Ejemplo donde no pudo controlar, estímulo ISS...................................................... 90 Figura 4.23 Variación de gL, de tónico a burst................................................................................ 92 Figura 4.24 Variación de gL, de burst a tónico................................................................................ 93 Figura 4.25 Variación de gT, de tónica a burst. ............................................................................... 94 Figura 4.26 Variación de gT, de burst a tónico................................................................................ 95 Figura 4.27 Diagrama de estado del sistema de control. ............................................................... 97 Figura 4.28 Control de burst a tónico, estímulo DC. .................................................................... 98 Figura 4.29 Casos en que no aplicó acción de control, estímulo DC.......................................... 99 5 Capítulo 1 Introducción En el siglo XX los avances en el área de la salud han sido sorprendentes. Elementos como el marcapasos y el ecógrafo, que actualmente son una efectiva herramienta para el tratamiento y diagnóstico de enfermedades respectivamente, fueron inventados en la década de 1950 y se masificaron en la década del 70. Como estos ejemplos hay muchos otros que ilustran el aporte que la ingeniería eléctrica ha realizado a la salud humana en el siglo recién pasado. Estos aportes no se han limitado a la invención y diseño de máquinas e instrumentos, sino que además han abarcado el área de la fisiología, en especial la modelación de sistemas fisiológicos. La electroestimulación es un área de la medicina que se encarga de la estimulación muscular y neuronal mediante impulsos eléctricos, ya sea como terapia de rehabilitación para el tratamiento del dolor o para el control de patologías. Actualmente muchos equipos de electroestimulación, en particular de neuroestimulación, se encuentran en su etapa de pruebas de prototipos [ 1]. Es en esta área en que el uso de modelos fisiológicos es parte fundamental, pues permite diseñar, optimizar y probar estímulos y electrodos, previo a su aplicación en seres vivos. 1.1. Estimulación Cerebral Profunda La Estimulación Cerebral Profunda o Deep Brain Stimulation (DBS) es una técnica que consiste en implantar electrodos en áreas específicas del cerebro y mediante estos electrodos estimular dicha área con trenes de pulsos de alta frecuencia (sobre 100 [Hz]), provenientes de un 6 generador de pulsos similar al marcapasos cardiaco [ 2]. En la Figura 1.1 se muestra un esquema de un dispositivo DBS implantado. A. Generador de señales, B. Electrodos de estimulación. Figura 1.1 Esquema del dispositivo de DBS implantado. Adaptada de [ 3]. La estimulación cerebral profunda (DBS) fue aplicada por primera vez al hipotálamo para tratar el dolor crónico en 1950 por el Dr. Pool [ 4]. Treinta años después, en 1980, esta técnica fue aplicada por el Dr. Alim-Louis Benabid del Grenoble Universty Hospital en Francia para tratar el Temblor Esencial y la Enfermedad de Parkinson estimulando áreas como el Núcleo Subtalámico (STN) o el Globo Pálido interno (GPi). En ambas enfermedades el principal síntoma es el temblor rítmico e involuntario de las extremidades y la DBS permite disminuir e incluso eliminar este y otros síntomas asociados [ 5]. Actualmente existen dispositivos de DBS aprobados por la FDA1 para tratar ambas enfermedades, y son más de 30.000 los pacientes en el mundo que se han sometido a una cirugía de este tipo [ 6]. Los dispositivos de DBS disponibles en el comercio están constituidos por el neuroestimulador (ver Figura 1.1 A), y uno o más electrodos (ver Figura 1.1 B). Los voltajes generados por el neuroestimulador son programados por el médico después de que el dispositivo 1 De su sigla en inglés Food and Drug Administration, Agencia del gobierno de Estados Unidos que regula alimentos y medicamentos en dicho pais, dependiente del Departamento de Salud de Estados Unidos (U.S. Department of Health and Human Services). 7 ha sido implantado. Los electrodos pueden ser programados como ánodo (positivo) o cátodo (negativo), en configuración monopolar o bipolar. En configuración monopolar, la cubierta de titanio del neuroestimulador se usa como ánodo, mientras el electrodo se sitúa cerca del área a estimular. En configuración bipolar ambos electrodos, ánodo y cátodo, se sitúan cerca de la zona a estimular. Los electrodos tienen una impedancia entre 600 – 1000 [Ω]. Para desórdenes de movimiento los parámetros del estímulo son entre 2 – 4 [V], lo que equivale a 2 – 4 [mA] si el dispositivo es de 1000 [Ω] y un ancho de pulso entre 90 – 180 [µseg] y 100 – 185 [Hz] [ 1]. En la Figura 1.2 se muestra la forma de onda en distintas terapias de electroestimulación, donde la primera fila corresponde a DBS. El hecho de que se programe el voltaje de estímulo y no la corriente, se debe a que el potencial daño que puede sufrir el tejido depende de la densidad de corriente aplicada sobre el tejido, por ello los dispositivos comerciales (e.g. Medtronic Inc. [ 3]) permiten programar el voltaje aplicado. La corriente de estímulo debida a este voltaje dependerá de la impedancia del tejido y es por ello que el médico tarda varios meses en estabilizar los parámetros del estímulo aplicado. ECT: Terapia electroconvulsiva (electroconvulsive therapy). VNS: Estimulación del nervio vago (vagus nerve stimulation). Figura 1.2 Formas de onda en terapias de electroestimulación. Adaptada de [ 7]. El mecanismo de acción de la DBS aún no se conoce completamente, pero los resultados que se obtienen son similares que con una lesión quirúrgica del área estimulada. Es por ello que a la fecha se han hecho pruebas clínicas en que se aplica DBS para tratar otras enfermedades para las 8 cuales las lesiones quirúrgicas son una alternativa. Hasta ahora la DBS se ha aplicado para tratar la Enfermedad de Parkinson, el Temblor Esencial, Distonía [ 4], el Trastorno Obsesivo Compulsivo (TOC), la Depresión, el Síndrome de Tourette, la Epilepsia, estados de mínima conciencia y jaquecas [ 6]. Las zonas que se estimulan comúnmente son la corteza cerebral para el TOC, el Tálamo para los desordenes de movimiento y la cápsula interna para la depresión tal como se ve en la Figura 1.3. Figura 1.3 Áreas de estimulación en la terapia DBS. Adaptada de [ 6]. 1.2. Objetivos Reconociendo que la DBS es una terapia que aún no se ha desarrollado completamente, muchos equipos de investigación concuerdan en que los desarrollos a través de simulaciones permitirán no sólo diseñar terapias y electrodos, sino además avanzar en la comprensión del mecanismo de acción de la DBS [ 1], [ 8], [ 9]. Es en el área de desarrollo de terapias en que se enmarca esta memoria. Se propone diseñar un sistema de control que permita, mediante estímulos de corriente, controlar el modo de descarga de un modelo dinámico de neurona talámica (Neurona Planta), bajo la restricción de que esta acción de control debe ser tal que lleve a la Neurona Planta al modo de descarga de una Neurona de Referencia. 9 1.2.1. Objetivos generales Los objetivos generales que se propone lograr son: Analizar un modelo dinámico de una neurona talámica del núcleo lateral geniculado para fines de control. Diseñar un esquema de control que permita obtener el modo de respuesta del modelo y determinar la acción de control necesaria según sea el caso. 1.2.2. Objetivos específicos Los objetivos específicos que se propone alcanzar con la realización de este trabajo son: o Realizar una revisión de los modelos de neuronas talámicas disponibles a la fecha. o Realizar una revisión de los estados fisiológicos de la neurona talámica en relación a las variables y parámetros del sistema. o Determinar las condiciones que permiten que la neurona descargue en uno u otro modo de respuesta (modo tónico o burst). o Analizar mediante simulaciones y teoría de sistemas dinámicos, el modelo de neurona. o Diseñar un sistema de control que permita modificar el modo de descarga de una neurona, para que se iguale al modo de descarga de otra neurona de referencia. 10 1.3. Estructura de la memoria El presente trabajo está dividido en cinco capítulos los cuales se resumen a continuación. En el Capítulo 1 se presenta una motivación al tema, se plantea el problema a tratar y se presenta además los objetivos de esta memoria En el Capítulo 2 se introduce la neurona desde el punto de vista biológico, tanto su estructura como su funcionamiento. Luego se indican los modelos matemáticos de las mismas, sus bases biofísicas y sus limitaciones. En este contexto se indican los modelos dinámicos de neuronas talámicas disponibles actualmente y se presenta el modelo con que se trabajará en esta memoria. En el Capítulo 3 se realiza un análisis dinámico del modelo y se determinan las características y modo de respuesta ante distintos estímulos. En el Capítulo 4 se diseña y aplica un esquema de control que permite determinar el modo de descarga de 2 neuronas y determinar la acción de control necesaria para que la neurona Planta tenga el mismo modo de descarga definido por la neurona Referencia En el Capítulo 5 se presentan las conclusiones sobre el trabajo realizado, además de proponer diversos trabajos futuros a desarrollar en el área. 11 Capítulo 2 Neuronas y Modelos Este Capítulo se divide en 2 partes; en la primera se presentan las neuronas y los conceptos básicos sobre éstas, se introducen las neuronas en general, su estructura física y su funcionamiento. En la segunda parte se presentan modelos dinámicos de neuronas, partiendo desde el modelo clásico de Hodgkin - Huxley [ 15]. 2.1. Características y estructura de las neuronas Las neuronas son la unidad funcional y estructural del Sistema Nervioso. Su función es generar, conducir y trasmitir los impulsos nerviosos desde las distintas zonas del cuerpo hasta el cerebro (también desde el cerebro hacia el resto del cuerpo) y dentro del cerebro. Para entender como ocurre la transmisión de estos impulsos nerviosos es necesario entender la estructura de las neuronas y sus mecanismos de transmisión. En general, las neuronas, como todas las células, son una única unidad funcional, especializadas en conducir y transmitir las señales eléctricas en el Sistema Nervioso. Desde el punto de vista físico se pueden distinguir 3 partes: soma (o cuerpo), axón y dendritas, las que se muestran en la Figura 2.1 para una neurona motora (A) y una neurona sensitiva (B) [ 10], [ 11]. La morfología de las neuronas puede variar según su función específica, tal como se puede ver en la Figura 2.2, donde se muestran neuronas de la retina, de Purjinke y piramidales, entre otras. 12 Figura 2.1 Esquema de neurona motora (A) y neurona sensitiva (B). Adaptada de [ 14]. Figura 2.2 Esquema de distintos tipos de neuronas. Adaptada de [ 11]. 13 Las dendritas son prolongaciones neuronales que reciben las aferencias sinápticas, que son las señales provenientes de otras neuronas. El soma es por definición la zona que contiene al núcleo. El axón hace las veces de un cable de transmisión, pues puede llevar información en forma de potenciales de acción desde el soma a las dendritas y viceversa. Es decir, es posible la transmisión en ambos sentidos. Será la estructura física de las neuronas y el tipo de sinapsis que realizan lo que determina si la transmisión de la señal eléctrica es uni o bi-direccional [ 10], [ 11]. 2.1.1. Membrana neuronal Para entender el mecanismo de transmisión a través de la neurona, en particular en el axón, es necesario comprender como está constituida su membrana. En la Figura 2.3 se muestra un esquema de membrana, donde se muestra la capa bilipídica, que es la membrana e inserta en ella los canales iónicos y las bombas transportadoras de iones. Tanto los canales como las bombas participan en los movimientos de iones a través de la membrana. Las bombas mueven los iones activamente en contra de su gradiente de concentración, por lo que establecen los gradientes iónicos. Los canales permiten el paso de ciertos iones a través de la membrana a favor de su gradiente, por lo que se dice que le confieren a la membrana la capacidad de tener permeabilidad selectiva [ 11]. B. Canales iónicos A. Bombas iónicas. Figura 2.3 Esquema de la membrana neuronal. Adaptada de [ 11]. Los canales iónicos pueden estar en 2 estados; abierto o cerrado. Si bien los mecanismos que permiten que estén en uno u otro estado son complejos, y escapan a los alcances de este trabajo, es importante distinguir entre canales dependientes de ligando y canales dependiente de voltaje. En los canales dependientes de ligando, como su nombre lo dice, su estado de apertura o cierre 14 depende de que un ligando específico se encuentre o no adherido en su correspondiente dominio en el canal. Mientras que los canales voltaje dependientes su estado dependerá del voltaje de membrana. 2.1.2. Potenciales de acción La membrana plasmática aísla el medio interno de la neurona del medio externo. Es debido a la membrana, sus canales y bombas, que la concentración de iones en el medio externo será distinta a la concentración de iones en el medio interno de la neurona. Esta diferencia de concentración produce una corriente de iones cuando los canales de dicho ión así lo permiten (cuando están abiertos), está corriente se traduce en que se puede medir una diferencia de potencial entre el exterior y el interior de la neurona, esto es lo que llamaremos potencial de membrana. Cuando la neurona no está transmitiendo información, como potenciales de acción, su potencial de membrana tendrá un valor fijo que llamaremos potencial de reposo, este potenciales se debe a la corriente de fuga o leak. Por otro lado, cuando transmite información lo hace a través de una forma de onda definida, que se llama potencial de acción. Al recibir un estímulo, por ejemplo una corriente, la neurona lo traduce como un cambio en su potencial de membrana, es así como toda vez que una neurona reciba un estímulo variará su potencial de membrana. Pero no siempre el estímulo aplicado gatillará un potencial de acción. Es por esta propiedad que se dice que el modo de respuesta de las neuronas es de todo o nada. El estímulo debe superar una valor umbral para gatillar un potencial de acción, si no supera este umbral, el estímulo se reflejará como una variación del potencial de membrana desde el reposo. En la Figura 2.5 se muestra evolución en el tiempo del potencial de membrana cuando el estímulo es subumbral (Figura 2.5 – A) y cuando el estímulo es supraumbral (Figura 2.5 – B). 15 A. Respuesta pasiva (arriba) ante estímulo subumbral (abajo). Corriente máxima de estímulo 2 [µA]. B. Espiga (arriba) ante estímulo supraumbral (abajo). Corriente máxima de estímulo 2.5 [µA]. Figura 2.4 Formas de onda del potencial de membrana. En una neurona, los potenciales de acción fluyen desde el soma hacia las terminaciones sinápticas, a través del axón (ver Figura 2.1). El modo en que se transmite esta señal es a través de potenciales de acción, los cuales se propagan desde el punto de inicio de la transmisión en el cono axónico hasta el extremo de los terminales sinápticos. En la Figura 2.5 se muestra un esquema de propagación de los potenciales de acción dentro del axón de una neurona en función del tiempo y el largo. Figura 2.5 Esquema de transmisión. Adaptada de [ 11]. 16 2.1.3. Potenciales sinápticos Para transmitir el impulso nervioso de una neurona a otra debe ocurrir una sinapsis entre las dos neuronas. Las sinapsis se dividen en 2 grandes grupos, las sinapsis eléctricas y las sinapsis químicas. La sinapsis química es la más frecuente en el Sistema Nervioso Central, y consiste en transmitir la información desde una neurona a otra mediante neurotransmisores [ 11]. En la Figura 2.6 se muestra un diagrama que muestra como ocurre la transmisión química. Figura 2.6 Esquema de sinapsis química. Adaptada de [ 11]. Una vez que un potencial de acción alcanza los terminales sinápticos de la neurona, este provoca el influjo de iones de Calcio a través de canales de Calcio voltaje dependientes, los cuales permiten la liberación de neurotransmisores hacia el medio externo. Éstos difunden por el medio, interactúan con los receptores de membrana postsinápticos que gatillan la apertura de canales iónicos dependientes de ligando, lo que provoca un flujo de iones en la membrana postsináptica. Esta corriente de iones provoca un potencial postsináptico excitatorio (EPSP) o inhibitorio (IPSP) [ 10], [ 11]. En la Figura 2.7- A se puede ver el potencial de membrana presináptico, la 17 corriente presináptica que provoca (Figura 2.7 – B) y el potencial postsináptico asociado (Figura 2.7 – C). A. Potencial de membrana presináptica [mV]. B. Corriente de Calcio presináptica [µA/cm]. C. Potencial de membrana postsináptico [mV] Imagen adaptada de [ 11]. Figura 2.7 Potencial presináptico, corriente presináptica y potencial postsinápco. 2.2. Modelos neuronales Uno de los modelos fisiológicos que ha suscitado mayor interés es el de los científicos Alan Hodgkin y Andrew Huxley. Presentado en 1952 como parte de una serie de experimentos que explicaban los mecanismos iónicos de la membrana neuronal. Fueron los primeros en entregar un sistema de ecuaciones que modelara el potencial de membrana del axón, a partir de la dinámica del movimiento de los iones ante un estímulo de corriente [ 15]. Este modelo fue la conclusión de una serie de papers publicados en 1952 en el Journal of Physiology [ 16], [ 17], [ 18], donde describían los experimentos realizados en el axón gigante de calamar. Fue por estas investigaciones que Hodgkin y Huxley ganaron el Premio Nobel de Medicina en 1963 [ 19]. Este trabajo de Hodgkin y Huxley es reconocido como uno de los modelos fisiológicos más importante, pues fue el primer modelo elaborado a partir de resultados experimentales, capaz de predecir muchas características del potencial de membrana, tales como los cambios de permeabilidad, la respuesta todo o nada del voltaje y el período refractario de membrana. Si bien al pasar los años se han encontrado limitaciones al modelo, debidas principalmente a los avances en los conocimientos moleculares de la membrana, no han sido capaces de invalidar el modelo. 18 Por sobre las limitaciones que pueda tener, el modelo presenta características físicas de la dinámica celular, que son consideradas en la mayoría de los modelos posteriores, como [ 20]: 1. El cambio en el sentido de INa e IK a ENa y EK, respectivamente. 2. La comunicación entre canales se produce a través de la membrana mediante el voltaje y producto de un campo eléctrico, y no producto de alguna reacción química. 3. La activación depende fuertemente del potencial de membrana. El sistema de ecuaciones de Hodgkin-Huxley (HH) modela una sección infinitesimal de membrana del axón gigante de calamar como el circuito de la Figura 2.8. Este modelo supone que el voltaje de membrana viene dado por la suma de 3 corrientes iónicas, dos de ellas variables debidas a los iones de Sodio (INa) y Potasio (IK), mientras la tercera es la llamada corriente de fuga o ‘leak current’ (Il) producto de la actividad residual de los canales de la membrana. La membrana se modela como un condensador Cm en paralelo con las conductancias g Na , g K y g l que dan cuenta de los cambios de permeabilidad de la membrana en función del voltaje. A su vez estas conductancias están en serie con las fuentes de voltaje ENa, EK y El respectivamente, que representan el voltaje debido a la diferencia entre la concentración interna y externa de iones, y que dan origen al movimiento pasivo de iones debido a su gradiente electroquímico. En la Figura 2.8 se indica como RNa el inverso de la conductancia gNa, lo mismo para el Potasio y la conductancia de fuga. El signo de los potenciales de reversa ENa, EK y El que se indica en la Figura 2.8, depende de si la mayor concentración de iones está dentro o fuera de la membrana. Figura 2.8. Análogo eléctrico de una sección de membrana. Adaptada de [ 15]. 19 El circuito de la Figura 2.8 queda descrito por la ecuación diferencial (2.1). CM dE = I ionica + I dt (2.1) donde, I ionica =I Na + I K + I l I ionica = g Na ( E − E Na ) + g K ( E − E K ) + g l ( E − El ) (2.2) Los canales iónicos de Sodio y Potasio son dependientes del voltaje. Para modelar el cambio de permeabilidad de la membrana, mediante la variación del estado de los canales, en función del voltaje de la misma, las conductancias g Na y g K se definen como: g Na = g Na m 3 h gK = gK n4 (2.3) La variable m3 indica la proporción de canales de Na+ abiertos en cierto instante, y puede variar en el rango continuo [0,1]. El valor 0 indica que ningún canal está activo, mientras que el valor 1 indica que el número máximo de canales de Na+ está activo. La variable (1-h) es el número de canales de Na+ inactivo en un cierto instante, también se mueve en el rango continuo [0,1]. Luego si (1-h) alcanza el valor 1, 1 − h = 1 ⇒ h = 0 quiere decir que el número máximo de canales está inactivado, mientras que si h = 1 significa que ningún canal se encuentra inactivado [ 21]. La variable n4 da cuenta de la proporción de canales de K+ activos en un cierto instante. Toma valores en el rango [0,1], representando el valor 0 que ningún canal está abierto y 1 que el número máximo de canales está abierto [ 21]. Si bien en el paper de Hodgkin-Huxley [ 15] se designan las variables n, m y h como una probabilidad, y actualmente se sabe que los canales están en los estados abierto, inactivo o cerrado con una cierta probabilidad, se evitó designarla así en este texto pues en el modelo no se definen estas variables como una variable aleatoria, luego sólo confundiría el hecho de definirla como una probabilidad en este caso. La ecuación diferencial genérica de activación e inactivación de canales en el modelo HH es [ 15]: 20 dn = α (Vm )·(1 − n ) − β (Vm )n dt (2.4) donde α(Vm) y β(Vm) son tasas de transferencia de iones desde afuera hacia adentro de la membrana y viceversa, respectivamente, son dependientes del voltaje y están en unidades de 1/[mseg]. Si se define: n∞ = α (Vm ) α (Vm ) + β (Vm ) τn = 1 α (Vm ) + β (Vm ) ⇒ dn n∞ − n = dt τn (2.5) Finalmente las ecuaciones del modelo de HH son (notar que V está medido respecto al potencial de reposo del axón): dV 1 = ( I ext − g Na m 3 h(V − V Na ) − g K n 4 (V − Vk ) − g l (V − Vl )) dt C m dm = α m (1 − m) − β m m dt dh = α h (1 − h) − β h h dt dn = α n (1 − n) − β n n dt (2.6) donde: g Na : gK : gl : VNa , VK , Vl : Cm : n: m: h: Conductancia máxima de sodio [mS/cm2]. Conductancia máxima de potasio [mS/cm2]. Conductancia de fuga [mS/cm2]. Voltaje de reversa [mV]. Capacitancia debida a la membrana [µF/cm2]. Dinámica de activación de los canales de potasio. Dinámica de activación de los canales de sodio. Dinámica de inactivación de los canales de sodio. 21 Las variables αm, βm, αh, βh, αn y βn de (2.6) se definen como: αm = − 0.1(V + 25) ⎛ (V + 25) ⎞ 1 − exp⎜ ⎟ ⎝ 10 ⎠ βh = ⎛V ⎞ α h = 0.07 exp⎜ ⎟ ⎝ 20 ⎠ αn = ⎛V ⎞ ⎟ ⎝ 18 ⎠ (2.7) 1 ⎛ V + 30 ⎞ 1 + exp⎜ ⎟ ⎝ 10 ⎠ (2.8) β m = 4 exp⎜ − 0.01(V + 10) ⎛ V + 10 ⎞ 1 − exp⎜ ⎟ ⎝ 10 ⎠ ⎛V ⎞ ⎟ ⎝ 80 ⎠ β n = 0.125 exp⎜ (2.9) En la Tabla 2.1 se muestran los valores de los parámetros para el modelo de HH, presentados por los autores en su trabajo de 1952 [ 21]. En la Figura 2.9 se muestra la gráfica de las variables del modelo HH para los valores de la Tabla 2.1. En esta Figura y en lo que sigue en este texto se definirán los gráficos con unidades de [mV] para voltaje, [mseg] para el tiempo y [µA] para la corriente. VARIABLE VALOR UNIDADES VARIABLE VALOR UNIDADES g Na 120 [mmho/cm2] VNa -115 [mV] gK 36 [mmho/cm2] VK +12 [mV] gl 0.3 [mmho/cm2] Vl -10.6 [mV] Cm 1 [µF/cm2] Tabla 2.1 Parámetros del modelo HH. En la Figura 2.9 – A se muestra el estímulo aplicado en el milisegundo 200, en la Figura 2.9 – B se ve la espiga o potencial de acción que produjo este estímulo. En la Figura 2.9 – C se puede ver como evoluciona la variable n ante el estímulo, se puede ver que su máximo se produce un poco antes de los 210 [mseg]. En la Figura 2.9 – D se puede ver las variables m y h, el máximo de m se produce unos instantes antes que el mínimo de h, y ambos preceden el máximo de n. La secuencia temporal de la dinámica de n, m y h, es importante pues en ellas subyace el mecanismo de generación del potencial de acción en este modelo. Cuando llega un estímulo, se produce una apertura de los canales de Na+, con lo que se produce una corriente de Na+cuyo máximo viene definido por m y h. Esta corriente eleva el voltaje de membrana hasta un valor cercano a VNa, esto es lo que se llama depolarización o fase ascendente del potencial. Tras un período de tiempo los 22 canales de K+ se activan (con cierto retardo) y los canales de Na+ se inactivan (mínimo de la curva h), con lo que empieza el descenso del potencial de membrana, a esta fase se le llama fase descendente. En cierto instante los canales de K+ activos son muchos más que los canales de Na+ activos (de hecho la mayoría de canales Na+ se encuentran inactivos), con lo que el voltaje de membrana se acerca al voltaje VK, esto corresponde al mínimo del voltaje de membrana de la Figura 2.9 – B. Como en esta fase se alcanzan voltajes bajo el voltaje de reposo de la neurona se llama fase refractaria y se denomina este voltaje como hiperpolarización. En respuesta a la hiperpolarización del voltaje de membrana se cierran los canales de K+ con lo que el voltaje de membrana vuelve al reposo. Las fases del potencial de acción y sus nombres se muestran en la Figura 2.10. A. Corriente de estímulo. Va de 0 a 2.5 [µA] en el milisegundo 200. B. Voltaje de membrana. C. Variable de activación n. D. Variable de activación m y de inactivación h. Figura 2.9 Gráfica de las variables del modelo HH. 23 Figura 2.10 Fases de un potencial de acción. 2.3. Neuronas talámicas y modelos A continuación se presentan las neuronas talámicas y sus características. Luego se hace un resumen de los modelos de estas neuronas disponibles en la literatura actualmente. 2.3.1. Características fisiológicas y función El tálamo, esta constituido por una colección de núcleos y es el componente principal del diencéfalo. Su principal función es la de transmitir la información sensitiva proveniente de los centros sensoriales inferiores (vista y tacto por ejemplo) hacia la corteza cerebral [ 11]. Es decir, el tálamo pasa las señales sensoriales desde la periferia hacia el lugar donde se integra con otros componentes, para generar la respuesta a esa señal en la corteza cerebral. Así mismo, las señales de respuesta al estímulo que envía la corteza, pasan por el tálamo. Es por ello que se denomina a las neuronas del tálamo neuronas talámicas de pasada o de paso (TRN del inglés Thalamic Relay Neuron). En la Figura 2.11 se muestra en rojo la zona que ocupa el tálamo en una vista sagital del encéfalo adulto. 24 Figura 2.11 Vista sagital del encéfalo adulto. Adaptada de [ 11]. De aquí en adelante se tratará el núcleo lateral geniculado (LGN), que es la zona del tálamo que transmite la información desde la retina hacia la corteza visual, ver Figura 2.12. Nos restringiremos a esta zona pues es la más estudiada del tálamo y porque el modelo con que se trabajará utiliza neuronas del LGN. Existen 2 clases de neuronas de paso en el LGN: Las X o células β y las Y o células α. Si bien existen diferencias morfológicas y fisiológicas entre estas dos clases de neuronas, tienen distinta participación en los circuitos neuronales que forman [ 10]. Al igual que todas las neuronas del tálamo son eléctricamente compactas, es decir se pueden aproximar como una unidad olvidando las propiedades distribuidas de éstas. Aunque una explicación más detallada del tema escapa a los objetivos de este trabajo, se debe tener presente que existen diferencias entre ambas clases. En la Figura 2.13 se puede ver un ejemplo de neurona del núcleo ventrobasal de una rata. 25 Figura 2.12 Imagen del tálamo y del núclo lateral geniculado. Adaptada de [ 12]. Figura 2.13 Neurona talámica. Adaptada de [ 13]. Las neuronas talámicas, en general, poseen dos modos de descarga (respuesta) que se describen más adelante. Existe una gama de conductancias y corrientes de membrana en las neuronas talámicas, son estas corrientes las que le permiten 2 modos de descarga. Si bien a medida que avanzan las investigaciones en el tema se encuentran nuevas conductancias [ 10], [ 22], [ 23] que podrían venir asociadas a nuevas corrientes, las corrientes más importantes y estudiadas son las de Na+, K+ y Ca2+, pues son las responsables de generar los distintos tipos de respuesta. 26 Corrientes de Na+: Estas corrientes se deben principalmente a dos conductancias de Na+ dependientes del voltaje del tipo Hodgkin-Huxley. Una persistente (INap) asociada a los canales de Sodio Nav1.6 y la otra rápida (INa de HH) asociada al canal de Sodio Nav1.2, las cuales generan la depolarización necesaria para inactivar IA e IT [ 24], [ 25]. Corrientes de K+: Se conocen muchas conductancias de iones de K+, algunas dependientes de voltaje y/o ligando. Las más importantes son el rectificador de K+ (IK) asociada a los canales Kv1, que repolariza la membrana después de INa. La corriente de fuga IKLeak y la corriente de K+ IA, esta última está asociada a los canales Kv4.2 – 4.3. La corriente IA al igual que IT, está inactiva a voltajes depolarizantes y se activa a niveles de voltaje hiperpolarizantes, lo que significa que se opone a la depolarización, pero como es más lenta que IT, esta última logra activarse y producir una espiga antes de que IA se active [ 25]. Corrientes de Ca2+: Se conocen dos conductancias de Ca2+, una de bajo umbral asociada a los canales Cav3.1 – 3.3 y otra de alto umbral asociada a los Cav2.1. Si bien en algunos modelos se consideran ambas corrientes, se sabe que es la corriente de bajo umbral IT la que juega un papel principal en la generación de distintos modos de respuesta en las neuronas talámicas de paso (TRN). Esto dado que si dicha corriente se encuentra activa la repuesta de las TRN será del tipo burst, mientras que si IT está inactiva la repuesta será tónica2. Es decir, de esta corriente depende el modo de repuesta de las TRN [ 26]. Corriente de hiperpolarización: Debida a iones K+(Ih), principalmente, es una corriente depolarizante que está activa a voltajes de hiperpolarización, con lo que eleva el voltaje lo suficiente para que se des-inactive IT, con lo que se depolariza la membrana, la cual es repolarizada por corrientes de iones K+. Esto último vuelve a activar a Ih iniciándose nuevamente el ciclo de bursting. Es decir, esta corriente es responsable de la respuesta rítmica de la neurona. La única forma de interrumpir este ritmo de disparo es aplicando una excitación suficientemente larga y depolarizante como para llevar la repuesta al modo tónico [ 27], [ 28]. Existen muchas enfermedades (mutaciones de los genes asociados principalmente) que afectan a los canales iónicos, y hacen que las corrientes aquí mencionadas tengan una dinámica 2 Los conceptos descargas en modo burst y tónico se explican en los párrafos siguientes. 27 distinta de la esperada en organismos sanos. A si mismo estas enfermedades a los canales iónicos se asocian con algunos tipos de epilepsia, parálisis, algunos tipos de convulsiones, arritmias y miotonía [ 29], [ 30], [ 31]. Ahí el interés en caracterizar las corrientes mencionadas. Es debido a las corrientes de Na+, K+, Ca2+ e hiperpolarización que las TRN pueden presentar dos estados o modos de descarga [ 22]: 1. Ondas de actividad rítmica sincronizada. Estas ondas lentas se presentan en estados de somnolencia o sueño. Se caracterizan por patrones de onda tipo burst o estallido (en lo siguiente se denominará indistintamente como burst o estallido), en un rango de frecuencias entre 0.5 – 4 Hz. Ver Figura 2.14 – B. 2. Actividad tónica, caracterizada por espigas regulares dependientes del estímulo, la cual se presenta durante la vigilia y el sueño de movimientos oculares rápidos (REM). Su rango de frecuencia es el de las ondas γ. Ver Figura 2.14 – A. Al pasar de un estado a otro cambia no sólo el modo de descarga de estas neuronas sino también la respuesta de éstas a los estímulos externos [ 22]. En la Figura 2.14 se muestra los potenciales de acción generados en cada estado y las corrientes de estímulo respectivas. Los registros fueron tomados in vitro de trozos de tálamo dorsal de conejillo de indias. En Figura 2.14 - A los pulsos rectangulares corresponden a las corrientes aplicadas a la muestra, (1a) y (2a), los registros de voltaje se indican como (1b) y (2b), respectivamente. En la Figura 2.14 – A se muestra una respuesta subumbral (1b) ante un pulso de corriente (1a). El registro (2b) muestra la respuesta tónica ante una corriente de igual forma de onda que (1a) a la que se le a sumado una componente continua positiva (2a) [ 23]. La Figura 2.14 – B muestra la respuesta en modo burst (arriba). A. Descarga pasiva (1b) ante estímulo subumbral (1a). Descarga en modo tónico (2b) ante estímula supraumbral (2a). Misma escala que B. B. Descarga en modo burst. Arriba, el voltaje de descarga, abajo la corriente de estímulo. Imagen extraída de [ 23]. Figura 2.14 Respuesta experimental de una TRN. 28 Si las neuronas talámicas son depolarizadas desde potenciales sobre -55mV, ya sea por una corriente artificial o por un potencial postsináptico excitatorio, la descarga observada es tónica (Figura 2.14 – A). Esta descarga se debe a la corriente de sodio INa y a la corriente del rectificador de potasio IK [ 23]. Mientras que si la neurona está hiperpolarizada durante un tiempo suficiente, es decir si se lleva a voltajes bajo los -65 mV, por medio de un potencial postsináptico inhibitorio o por la inyección de una corriente como se muestra en la Figura 2.14 – B, la descarga será de tipo burst [ 22]. Este potencial que permite ocurra un estallido es generado por la corriente de Calcio de bajo umbral IT, si el potencial de membrana es depolarizado desde un potencial hiperpolarizado, IT se activa al comienzo y luego se inactiva lentamente. Esto es lo que se llama una espiga de bajo umbral de Calcio (LTS: Low-threshold spike). Esta LTS lleva el potencial de membrana a un valor de depolarización suficiente (sobre umbral) como para disparar un burst compuesto de 3-8 potenciales de acción rápidos. Al inactivarse IT se repolariza la membrana hasta alcanzar un valor de hiperpolarización lo que activa la corriente Ih, la cual a su vez depolariza la membrana lo suficiente para que se produzca otro LTS que produzca un nuevo burst (ver Figura 2.15). De la sucesión de corrientes necesarias para tener una descarga tónica o burst, y de diversas investigaciones se ha concluido que es imposible tener un burst debido a una entrada sináptica desde el potencial de reposo, pues los canales de Calcio que generan la corriente IT no están activados [ 22]. Figura 2.15 Actividad oscilatoria del voltaje de membrana, burst. Adaptada de [ 22]. 29 2.3.2. Modelo de neurona talámica El modelo de Huguenard y McCormick (HM) [ 35], [ 34] es un análogo del modelo eléctrico presentado por Hodgkin y Huxley (HH) en 1952 [ 15], basado en resultados experimentales en neuronas talámicas. El modelo de HM considera 10 corrientes iónicas, a diferencia de las 3 que consideraba el modelo de HH. En este modelo el voltaje de membrana queda definido por la ecuación (2.11), I ext = Vm C m + I Na + I Nap + I L + I T + I C + I A + I K 2 + I h + I KLeak + I NaLeak Vm = − (I ext + I Na + I Nap + I L + I T + I C + I A + I K 2 + I h + I KLeak + I NaLeak ) Cm (2.10) (2.11) donde cada corriente viene dada por: I Na = g Na (Vm − E Na )(m Na ) hNa (2.12) I Nap = g Nap (Vm − E Na ) m Na (2.13) 3 I L = p L ·(m L ) ·z 2 2 [ ] [ ] Vm F 2 Ca 2+ i − Ca 2+ o exp(− zFVm / RT ) RT 1 − exp( zFVm / RT ) I T = pT (mT )(hT )z 2 [ ] [ ] Vm F 2 Ca 2+ i − Ca 2+ o exp(− zFVm / RT ) RT 1 − exp( zFVm / RT ) I C = g C (Vm − E K )mC [ (2.14) (2.15) (2.16) ] I A = 0.6· g A1 ·(m A1 ) ·h A1 + 0.4·g A2 ·(m A2 ) ·h A2 ·(Vm − E K ) (2.17) I K 2 = [0.6· g a , K 2 ·m a , K 2 ·ha , K 2 + 0.4· g b , K 2 ·mb , K 2 ·hb , K 2 ]·(Vm − E K ) (2.18) I h = g h ·m h ·(Vm − E eq ) (2.19) 4 4 En la Tabla 2.2 se indican los valores de los parámetros para las corrientes de iones de Ca2+, IL e IC. PARÁMETRO UNIDADES VALOR F, constante de Faraday Coulombs 96 485.3415 R, constante de los gases J·K-1·mol-1 8.314 472 T, temperatura C 35.5º z, valencia del Ca2+ adimensional 2 Tabla 2.2. Valores de los parámetros para las corrientes IC e IL. 30 Como se ve de las ecuaciones anteriores (desde la 2.12 a la 2.19), las corrientes vienen definidas por variables de activación, inactivación y la concentración de Calcio, las cuales dependen del voltaje de membrana como se presenta a continuación. En lo que sigue se designará [Ca2+] como χC y χL para cada corriente IC e IL, respectivamente. 0.091(Vm + 38) (1 − m Na ) + 0.062(Vm + 38) m Na ⎛ V + 38 ⎞ ⎛ V + 38 ⎞ 1 − exp⎜ m 1 − exp⎜ m ⎟ ⎟ ⎝ −5 ⎠ ⎝ 5 ⎠ (2.20) 2.07 ⎛ V + 55 ⎞ hNa = 0.016 exp⎜ m hNa ⎟(1 − hNa ) − ⎛ Vm − 17 ⎞ ⎝ − 15 ⎠ 1 + exp⎜ ⎟ ⎝ − 21 ⎠ (2.21) ⎛V ⎞ ⎛ −V ⎞ m C = 2.5·10 5 ·χ C ·exp⎜ m ⎟(1 − mC ) − 0.1exp⎜ m ⎟mC ⎝ 24 ⎠ ⎝ 24 ⎠ (2.22) m Na = χ C = m L = χ L = m T = − 5.18·10 −3 I L − χC 2.9·10 3 (2.23) 1 .6 (1 − m L ) + 0.02·(Vm − 1.31) m L 1 + exp(− 0.072(Vm − 5)) ⎛ V − 1.31 ⎞ 1 − exp⎜ m ⎟ ⎝ 5.36 ⎠ (2.24) − 5.18·10 −3 (I L + IT ) − χ L 2.9·10 3 (2.25) ⎡ ⎛ Vm + 57 ⎞⎤ ⎢1 + exp⎜ − 6.2 ⎟⎥ ⎝ ⎠⎦ ⎣ −1 − mT −1 ⎡ ⎛ Vm + 312 ⎞ ⎛ Vm + 16.8 ⎞⎤ ⎢exp⎜ − 16.7 ⎟ + exp⎜ 18.2 ⎟⎥ + 0.612 ⎠ ⎝ ⎠⎦ ⎣ ⎝ (2.26) −1 ⎡ ⎛ Vm + 81 ⎞⎤ ⎢1 + exp⎜ 4 ⎟⎥ − hT ⎝ ⎠⎦ hT = ⎣ τ hT (Vm ) ⎛ Vm + 467 ⎞ ⎟ ⎝ 66.6 ⎠ Vm < −80 mV τ h = exp⎜ Vm < −80 mV τ h = exp⎜ ⎛ Vm + 22 ⎞ ⎟ + 28 ⎝ − 10.5 ⎠ 31 (2.27) m A1 = ⎡ ⎛ Vm + 60 ⎞ ⎤ ⎟ + 1⎥ ⎢exp⎜ − ⎣ ⎝ 8.5 ⎠ ⎦ −4 − m A1 −1 (2.28) −1 (2.29) ⎡ ⎛ Vm + 35.8 ⎞ ⎛ Vm + 79.7 ⎞⎤ ⎢exp⎜ 19.7 ⎟ + exp⎜ − 12.7 ⎟⎥ + 0.37 ⎝ ⎠ ⎠⎦ ⎣ ⎝ ⎡ ⎛ Vm + 36 ⎞ ⎤ ⎢exp⎜ − 20 ⎟ + 1⎥ ⎠ ⎦ ⎣ ⎝ m A 2 = −4 − m A2 ⎡ ⎛ Vm + 35.8 ⎞ ⎛ Vm + 79.7 ⎞⎤ ⎢exp⎜ 19.7 ⎟ + exp⎜ − 12.7 ⎟⎥ + 0.37 ⎠ ⎝ ⎠⎦ ⎣ ⎝ h A1 = h A2 = ⎡ ⎛ Vm + 78 ⎞ ⎤ ⎢exp⎜ 6 ⎟ + 1⎥ ⎠ ⎦ ⎣ ⎝ −4 − h A1 (h A2 ) τ h , A1 (τ h , A 2 ) τ h, A1 = Vm < −63 mV 1 ⎛ V + 46 ⎞ ⎛ V + 238 ⎞ exp⎜ m ⎟ + exp⎜ m ⎟ ⎝ 5 ⎠ ⎝ − 37.5 ⎠ Vm ≥ −63 mV τ h , A1 = 19 Vm < −73 mV τ h , A 2 = τ h , A1 Vm ≥ −73 mV τ h , A 2 = 60 ⎡ ⎛ Vm + 43 ⎞ ⎤ ⎢exp⎜ − 17 ⎟ + 1⎥ ⎠ ⎦ ⎣ ⎝ m K 2 = −4 − mK 2 −1 ⎡ ⎛ Vm − 81 ⎞ ⎛ Vm + 132 ⎞⎤ ⎢exp⎜ 25.6 ⎟ + exp⎜ − 18 ⎟⎥ + 9.9 ⎝ ⎠⎦ ⎠ ⎣ ⎝ ha , K 2 = hb , K 2 = ⎡ ⎛ Vm + 58 ⎞ ⎤ ⎢exp⎜ 10.6 ⎟ + 1⎥ ⎠ ⎦ ⎣ ⎝ (2.31) −1 − ha , K 2 −1 ⎡ ⎛ Vm − 1.329 ⎞ ⎛ V + 130 ⎞⎤ ⎟ + exp⎜ m ⎟⎥ + 120 ⎢exp⎜ 200 ⎠ ⎝ − 7 .1 ⎠ ⎦ ⎣ ⎝ ⎡ ⎛ Vm + 58 ⎞ ⎤ ⎢exp⎜ 10.6 ⎟ + 1⎥ ⎠ ⎦ ⎣ ⎝ (2.30) (2.32) −1 − hb , K 2 τ h ,K 2 (2.33) b Vm < −70 mV τ h ,K 2 = τ h Vm ≥ −70 mV τ h , K 2 = 8900 a ,K 2 b b −1 ⎡ ⎛ Vm + 75 ⎞ ⎤ ⎢exp⎜ 5.5 ⎟ + 1⎥ − m h ⎠ ⎦ ⎣ ⎝ m h = [exp(− 0.0863·Vm − 14.59) + exp(0.0701·Vm − 1.87 )]−1 32 (2.34) Las unidades de las variables y parámetros presentados en las ecuaciones anteriores se muestran en la Tabla 2.3. VARIABLE O PARÁMETRO VARIABLE O UNIDADES PARÁMETRO UNIDADES Vm [mV] Ix [nA] [Ca2+], χx [M] Cm [nF] gx [µS] px [cm3/seg] mx Sin unidades hx Sin unidades τx [mseg] Tabla 2.3. Unidades de las variables y parámetros del modelo de HHM. En la Figura 2.16 se muestran ejemplos del voltaje de membrana Vm y la corriente de estímulo presentado por los autores de este modelo en [ 35]. La Figura 2.16 – A se muestra una respuesta tipo burst o estallido, mientras en Figura 2.16 – B se muestra la respuesta tónica. Figura 2.16 Resultados obtenidos con el modelo de HM. Debido a que HM es un modelo basado en la biofísica de las neuronas, da cuenta de 10 corrientes iónicas distintas. La dependencia de estas corrientes de ecuaciones diferenciales de activación, inactivación y concentración de Calcio, hacen que se llegue a un total de 16 ecuaciones 33 diferenciales no lineales. Esto significa una carga computacional importante al momento de simular este modelo. 2.3.3. Otros modelos Existen otros modelos biofísicos de las neuronas talámicas de paso. Dependiendo de las propiedades que quiera mostrar el modelo, descargas en estallido o tónicas u oscilaciones de éstas, es la complejidad y el número de corrientes que debe incluir. En la Tabla 2.4 se resumen los distintos modelos de neuronas talámicas de paso, para cada uno se indica las corrientes que considera y el tipo de respuesta que es capaz de producir. Los signos (+) indican que dicha corriente está presente en el modelo, los signos (–) indican que dicha corriente no es considerada en el modelo y los casilleros con signos (*) indican que se consideran en el modelo corrientes similares a la indicada en la casilla. PAPER INA INAP IK2 IC IT IL IH IA ILEAK BURST TONICA Huguenard 1992 + + + + + + + + + + + Destexhe 1993 - - + - + - + - - - + Golomb 1996 - - - - + - + - + - + Destexhe 1998 - - - - + - - - + + + McIntyre 2004 + - * - + - + - - + + Rhodes 2005 + - * + + * + + - + + Tabla 2.4 Resumen de las características de cada modelo. El modelo HM [ 35] (Hueguenard 1992 en la Tabla 2.4) tiene las 9 corrientes que se presentaron en la sección anterior y supone una neurona de un único compartimiento, es decir supone que una neurona se puede representar como un circuito RC similar al de la Figura 2.8. Reproduce respuestas del tipo estallido y espigas tónicas, al modificar sus parámetros es posible obtener estallido rítmicos ante entrada nula. Destexhe et al [ 13] en 1993 proponen un modelo compuesto de 3 compartimientos, es decir 3 circuitos RC como el de Hodgkin-Huxley conectados en paralelo a través de resistencias, cada compartimiento representa una zona de la neurona talámica. Comparan este modelo de 3 compartimientos y un modelo un compartimiento con resultados experimentales para la descarga en modo burst. Golomb et al [ 36] proponen un 34 modelo de neurona talámica en el contexto de un estudio sobre cómo se propagan las señales a través de poblaciones de neuronas talámocorticales acopladas con poblaciones de neuronas reticulotalámicas. En este modelo cada neurona es representada por un compartimiento. Destexhe et al [ 37] en 1998 usan un modelo de neurona talámica con el propósito de estudiar los mecanismos que producen las oscilaciones de ondas lentas, consideraron una neurona como un compartimiento. McIntyre et al [ 9] estudió los efectos de la DBS en un modelo de neurona talámica multicompartimiento (ver Figura 2.17) inmerso en un campo eléctrico debido al electrodo de estimulación. En 2005 Rhodes y Llinás [ 38] definen una neurona de 219 compartimientos con la que reproducen las descargas tónicas, estallidos y oscilaciones, y estudian cómo afecta la distribución de los canales iónicos a lo largo de la neurona en la generación de este tipo de señales. Figura 2.17 Modelo multicompartimiento de McIntyre. Adaptada de [ 9]. Es evidente, a partir de lo dicho en esta sección, que la simulación de neuronas de uno o varios compartimientos no es sencilla. No sólo por la carga computacional que representa sino por la necesidad de contrastar los resultados con datos experimentales. 2.3.4. Análisis dinámico y otros sistemas de control Luego de que en 1952 se presentara el modelo de HH aparecieron en la literatura especializada publicaciones que analizaban el sistema de HH en base a la teoría de los sistemas dinámicos. 35 Análisis sobre el modelo de HH tal como este fue presentado en su publicación de 1952 [ 15] existen en la literatura bajo distintas condiciones. En 1979 Rinzel y Millar [ 39] estudiaron la dinámica del sistema bajo distintos estímulos de corriente constante. Su trabajo permitió observar de forma global el ciclo límite inestable que se producía bajo cierto estímulo de corriente y que otros autores previos no habían podido describir debido a dificultades con el cálculo computacional. Más aún los autores encontraron una región de histéresis en que el sistema se alternaba entre uno de los dos puntos de estabilidad dependiendo si la corriente iba aumentando o disminuyendo. En el trabajo de Aihara y Matsumoto de 1983 [ 40] se presenta una síntesis de los puntos de interés del sistema de HH para el sistema autónomo variando la concentración externa de potasio. Es así como encuentra: Un rango en que la solución es única y asintóticamente estable. Otro en que hay un único punto inestable y un ciclo límite estable. Y una zona en que el ciclo límite estable y el equilibrio asintóticamente estable coexisten. Bedrov et al [ 41], estudiaron el tipo de solución que se obtenía al variar las conductancias de Sodio y Potasio, la corriente de estímulo, y la dependencia del voltaje de m, h, n. Debido a la dificultad de estudiar un espacio multiparamétrico, realizaron análisis de la matriz Jacobiana sobre combinaciones de 3 parámetros, así encontraron zonas donde se presentan soluciones estables y otras donde se generaban oscilaciones estable. Guckenheimer y Laboriau [ 42] presentaron un análisis del modelo HH donde describieron cómo la solución variava desde equilibrios a oscilaciones periódicas en función de la intensidad del estímulo y el voltaje de reversa del Potasio. En los trabajos mencionados en el párrafo anterior el énfasis estuve en el análisis del sistema HH desde el punto de vista de sistemas dinámicos, sin considerar el significado fisiológico de los resultados obtenidos, ni de los estímulos o parámetros utilizados. Las investigaciones de Hodgkin y Huxley de 1952 fueron hechas en la fibra nerviosa del calamar, pues tenía 800 [mm] de diámetro por lo que permitía insertar los electrodos disponibles en aquella época, a diferencia del axón de mamífero que, con un diámetro de 2 [mm], hacia impracticables los experimentos. Fue en 1976 que Adrian y Marshall [ 43] publicaron los registros de los parámetros numéricos suficientes para usar el modelo de HH aplicado al músculo extensor largo de los dedos (extensor digitorum longus) de la rata. Si bien existen otros modelos matemáticos más exactos para fibras musculares, como el de Falk y Fatt [ 44], muchas de las aplicaciones del modelo de HH a mamífero se basan en modificar los parámetros originales de 36 dicho modelo según sus análogos en estudios como el de Adrian y Marshall , Falk y Fatt o Pappone [ 46]. En general, a los resultados obtenidos a partir del modelo de HH con parámetros numéricos para mamíferos se les intenta dar un significado fisiológico. Lo usual es mencionar que los resultados se pueden asociar con estados epilépticos o desórdenes obsesivos compulsivos pero sin detallar y justificar de manera más precisa en qué se basan dichos supuestos. Terada et al estudian la dinámica del modelo ante variaciones en la conductancia de fuga y un parámetro de la conductancia de Sodio. Wang et al [ 47], [ 48], [ 49], quienes en su trabajo analizan el comportamiento del sistema ante una perturbación debida a un campo eléctrico externo de baja frecuencia, este campo eléctrico se representa como una fuente de voltaje en serie con las fuentes debidas a las concentraciones iónicas. Al variar el voltaje de esta fuente encuentran una bifurcación de Hopf que hace al sistema inestable [ 47], [ 48]. Esta misma situación la analizan frente a la variación de otros parámetros como conductancia de fuga la fuerza electromotriz del Potasio [ 49]. También han propuesto controles para el sistema. Para una bifurcación de Hopf que se produce ante una variación de la conductancia de fuga [ 50], presentan un control que mueve el punto en que se produce dicha bifurcación hasta un punto fisiológicamente imposible de alcanzar. Para una bifurcación de Hopf que se produce ante determinados valores de la corriente de estímulo también proponen un sistema de control [ 51], [ 52]. 37 Capítulo 3 Análisis del Modelo IFB El modelo integrate-and-fire-or-burst (IFB), fue propuesto por Smith, Cox, Sherman y Rinzel en el año 2000 [ 55]. Se basa en un modelo clásico de integrate-and-fire neuron (integrar y disparar) al cual se le agregó una variable lenta que representa la desinactivación de IT. Este modelo que fue construido para simplificar la carga computacional, permite reproducir la dependencia de la entrada que tiene la respuesta en modo burst y en modo tónico. Si bien en este modelo se disminuye la carga computacional en base a disminuir las características biofísicas modeladas por el sistema, la mayoría de las características biológicas de entrada y salida se mantienen inalteradas. El gráfico de la Figura 3.1 compara la eficiencia computacional con las características biológicas que reproduce el modelo. El modelo IFB (línea roja) está entre los más eficientes, pues toma entre 9 y 13 operaciones de punto flotante simularlo por 1 [mseg], y las características biológicas que reproduce son buenas. Por otro lado el modelo de Hodgkin y Huxley (línea verde) es el que mejor reproduce las características biológicas pero a un costo de 1200 operaciones por milisegundo de simulación. 38 Características biológicas que reproduce el modelo versus costo de implementación. Evaluado para una simulación de 1 [mseg]. Se llamó FLOPS al número aproximado de operaciones de punto flotante. Imagen extraída de [ 59]. Figura 3.1 Comparación de distintos modelos. El modelo IFB propuesto por Smith et al [ 55] se sintonizó a partir de registros de trozos de tálamo, en particular del núcleo lateral geniculado (LGN) de gatos entre 5 y 8 semanas de vida. La ecuación de balance de corriente para este modelo viene dada por [ 55]: C dV = I APP − I L − I T dt (3.1) La corriente aplicada viene definida por IAPP, mientras que IL e IT representan la corriente de fuga y la corriente de Calcio de bajo umbral, respectivamente. La corriente IL depende de una conductancia gL constante. La corriente IT depende de una variable lenta h y de una rápida, m∞. I L = g L (V − VL ) (3.2) I T = g T m∞ h(V − VT ) (3.3) En la ecuación (3.3) las variables h y m∞, dependen del voltaje V en relación con Vh, siendo Vh el voltaje límite en que se produce la activación de la descarga burst. Las ecuaciones (3.4) y (3.5) definen m∞ y h, respectivamente. ⎧1 m∞ = ⎨ ⎩0 V − Vh ≥ 0 V − Vh < 0 ⎧ −h ⎪⎪ τ − h = ⎨ h (1 − h) ⎪ ⎪⎩ τ h + (3.4) V ≥ Vh V < Vh 39 (3.5) La definición de h en (3.5) y m∞ en (3.4), es una idealización de la dinámica de IT a partir de la dinámica basada en datos experimentales de inactivación y activación de IT descrita en el modelo HM [ 34], ver Figura 3.2. La inactivación h depende de los parámetros τh- y τh+. El parámetro τhdetermina la duración que debe tener la hiperpolarización para producir un burst, mientras τh+ determina la duración del estallido. Estas variables se traducen en que para valores de voltaje V>Vh el valor de h se va a cero con una constante de tiempo τh-, y cuando V<Vh tiende a uno con una constante de tiempo τh+. A. Inactivación y activación. B. Idealización de la inactivación y activación. C. Activación. D. Idealización de la activación. Figura 3.2 Idealización de m∞ y h. Adaptada de [ 12]. Toda vez que el voltaje de membrana V(t) sea menor Vθ, la ecuación que rige al modelo es (3.1). Si V(t) es igual a Vθ entonces se produce un potencial de acción. Inmediatamente después de un potencial de acción el voltaje de membrana tiende al valor de reset, Vreset. La ecuación (3.6) describe el voltaje de membrana en el tiempo. 40 ⎧ dVm ⎪C dt = I APP − I L − I T ⎪ Vm (t ) = ⎨ Vspike ⎪ V reset ⎪ ⎩ si Vm (t − ) < Vθ si Vm (t − ) = Vθ si Vm (t − ) = Vspike (3.6) Los parámetros del modelo son los presentados en la Tabla 3.1. En la Figura 3.3 se muestra en la primera columna los resultados experimentales, en la segunda columna los resultados presentados en el paper y obtenidos con el modelo IFB y la tercera columna muestra los resultados obtenidos con el modelo implementado en Matlab 6.5 y resuelto mediante el método de Runge – Kutta [ 56] para esta memoria, ver Anexo A. Para los resultados experimentales presentados en el paper (ver Figura 3.3 - A) la magnitud de la corriente de estímulo en microampere no es la misma que fue aplicada al modelo (ver Figura 3.3 - B) para obtener una respuesta con idéntica forma de onda que la experimental. Es decir, para obtener una respuesta determinada, en el experimento y en simulaciones del modelo, las corrientes de estímulo tienen la misma forma de onda y sus magnitudes tienen una relación, pero no se aplica el mismo estímulo en ambos casos. En el ejemplo de la Figura 3.3 la corriente aplicada en el experimento para obtener una descarga tónica fue de -58 [pA], 230 [pA] y 430 [pA], mientras que el estímulo aplicado al modelo para obtener la misma descarga fue de 0.67 [µA], 1.33 [µA] y 0.67 [µA]. PARÁMETRO VALOR UNIDADES PARÁMETRO VALOR UNIDADES Vθ -50 mV gL 0.035 mS/cm2 Vh -70 mV gT 0.8 mS/cm2 Vreset -60 mV τh+ 100 mseg Vspike 30 mV τh- 20 mseg VL -75 mV C 2 µF/cm2 VT 120 mV Tabla 3.1 Parámetros usados en el modelo para las simulaciones. 41 A. Resultados experimentales, imagen adaptada de [ 55]. B. Resultados obtenidos con el modelo presentados en el paper, adaptada de [ 55]. C. Resultados obtenidos con el modelo IFB implementado en Matlab. Figura 3.3 Verificación de resultados del modelo IFB. Como se puede ver en la Figura 3.3 el modelo es capaz de generar descargas tónicas (o tonic spiking) y estallidos de rebote (o rebound burst). Las descargas tónicas se obtienen al aplicar un estímulo que depolariza el voltaje de membrana, es decir una corriente sobreumbral, mientras los estallidos de rebote se obtienen al liberar la neurona de una hiperpolarización, por eso se llama de rebote pues se producen al quitar el estímulo que hiperpolarizaba el voltaje de membrana. Se concluye que el modelo presentado, con los parámetros de la Tabla 3.1, puede presentar dos modos de respuesta dependiendo de la entrada aplicada. La validez del modelo IFB en modelar la topología de una neurona talámica de paso mediante un análogo puntual definido por una sistema de ecuaciones en el tiempo independientes de la variable espacial, radica en resultados experimentales que indican que las neuronas talámicas de paso X e Y son eléctricamente compactas, lo que quiere decir que entrads sinápticas distantes pueden tener efectos en el soma y el axón [ 57], [ 58]. Para modificar los modos de respuesta del modelo existe otra posibilidad, además de modificar la entrada, ésta es variar los parámetros del sistema [ 59]. Variando los parámetros del sistema se pude obtener también respuestas del tipo de una espiga en fase (o phasic spiking), un 42 estallido en fase (o phasic bursting), adaptación de frecuencia, excitabilidad clase 1, integración, espiga de rebote, biestabilidad, depolarización después de potencial (o depolarizing after-potentials DAP) y espigas inducidas por inhibición. En la Figura 3.4 se ilustra cada uno de estos modos de respuesta sobre la corriente de estímulo. A. Espiga en fase. B. Estallido en fase. C. Adaptación en frecuencia. D. Excitabilidad clase 1. E. Integrador. F. Espiga de rebote. G. Biestabilidad. H. Depolarización después de potencial. I. Espigas inducidas por inhibición. Figura 3.4 Tipos de repuestas modelo IFB. Adaptada de [ 59]. Como el objetivo de esta memoria es diseñar un sistema de control, no se calibrarán los parámetros para obtener otro tipo de respuesta sino que se usaran los indicados en la Tabla 3.1. 3.1. Análisis dinámico En la Figura 3.3 - C se muestra la respuesta del voltaje en función de la corriente de entrada, sin considerar cómo era la dinámica de h y m∞. En la Figura 3.5 se muestra la respuesta tipo burst 43 y el comportamiento de las variables V(t), IL, IT, h y m∞ frente a una entrada IAPP del tipo pulso igual a la aplicada en la Figura 3.3. Mientras que en la Figura 3.6 se muestran las mismas variables para la respuesta tónica. A. Corriente de estímulo. B. Voltaje de membrana. C. Corriente IT. D. Corriente IL E. Variable h. F. Variable m∞. Figura 3.5 Variables en respuesta burst. 44 A. Corriente de estímulo. B. Voltaje de membrana. C. Corriente IT. D. Corriente IL E. Variable h. F. Variable m∞. Figura 3.6 Variables en respuesta tónica. De la Figura 3.5 – E y Figura 3.5 – F se puede ver que mientras la corriente está en -0.16 [µA] h aumenta exponencialmente y m∞ se hace cero; una vez que la corriente ha aumentado a 0.34 [µA] h comienza a decaer a cero de manera exponencial en el mismo instante en que m∞ se hace uno (ver también Figura 3.7 – A donde la transición desde -0.16 [µA] a 0.34 [µA] ocurre en el milisegundo 500). Las variaciones de estas dos variables se traducen en que la corriente IT disminuye bruscamente de 0 a -110 [µA] en el instante en que comienza a decaer h (cuando m∞ es uno), hasta hacerse cero cuando h se hace cero. La descarga burst ocurre en sincronía con la 45 corriente IT. Cuando IT comienza a moverse desde cero hacia valores negativos comienza la descarga de espigas, estas espigas finalizan cuando IT alcanza el cero (ver sincronización Figura 3.7 - B). La corriente IL, ver Figura 3.5 – D, depende del voltaje de membrana de manera lineal, por lo tanto su forma de onda es similar y en sincronía con el voltaje de membrana. B. Voltaje de membrana (gris claro) y corriente IT (gris oscuro). A. Variables h (gris oscuro) y m∞ (gris claro). Figura 3.7 Sincronización de la variables en descargas burst. En el caso de la Figura 3.6, en los gráfico E y F se puede ver que las variables h y m∞ permanecen constantes en cero y uno, respectivamente, cuando la descarga es tónica. Por lo tanto la corriente IT permanece nula, sólo la corriente IL participa en la descarga tónica, ver Figura 3.6 – C y Figura 3.6 - D. Considerando como sistema dinámico la ecuación (3.1) y la ecuación (3.5), cuando V>Vh se obtiene el sistema de ecuaciones (3.7). Cuando V<Vh el sistema de ecuaciones es el (3.8). CV = I APP − g L (V − VL ) − g T m∞ h(V − VT ) −h h = − (3.7) τh CV = I APP − g L (V − VL ) − g T m∞ h(V − VT ) 1− h h = + (3.8) τh Ambos sistemas tienen 2 variables de estado (V,h). Para V>Vh la variable h es nula en el equilibrio mientras que V es igual a VL. En el caso en que V<Vh la variable h es uno y m∞ es nula 46 en el equilibrio, entonces el voltaje de equilibrio es nuevamente VL. De modo general ante una entrada arbitraria no nula el punto de equilibrio dependerá de esta corriente aplicada IAPP y de la relación entre V y Vh como lo muestra la ecuación (3.9) y (3.10). 0 ⎞ ⎛ h1 ⎞ ⎛⎜ I Si V ≥ Vh ⇒ ⎜⎜ ⎟⎟ = ⎜ APP + V ⎟⎟ L ⎟ ⎝V1 ⎠ ⎜⎝ g L ⎠ (3.9) ⎞ ⎛ h2 ⎞ ⎛⎜ I 1 Si V < Vh ⇒ ⎜⎜ ⎟⎟ = ⎜ APP + V ⎟⎟ L ⎝ V2 ⎠ ⎝ g L ⎠ (3.10) Los puntos de equilibrio (3.9) y (3.10), están restringidos a la zona de dominio de la ecuación (3.7) y (3.8), respectivamente. En la Figura 3.8 - C se muestra el diagrama de fase Vm v/s h para una entrada sinusoidal de frecuencia 2 [Hz], amplitud de la componente continua y de la sinusoidal 0.1 [µA]. En esta Figura se indican 6 puntos del diagrama y su correspondiente posición en los gráficos de voltaje de membrana (Figura 3.8 - A) y la variable h en función del tiempo (Figura 3.8 - B). Esta Figura permite comprender cómo influye h en la duración y número de espigas que genera el modelo en una descarga de estallido, pues el tiempo que demora h en ir desde su máximo (punto 3) hasta su mínimo (punto 6) es el período donde se producirán las descargas. Si la respuesta fuera pasiva o tónica h nunca dejaría de ser nula con lo que en el diagrama de fase se vería sólo una línea horizontal en cero. De los párrafos anteriores se puede concluir que una vez hiperpolarizado el voltaje de membrana por debajo de cierto umbral, en el caso del modelo IFB bajo Vh, se desencadenará una descarga de espigas en estallido, siendo sólo este modo de respuesta posible. Por otra parte existe otro umbral (Vθ en este modelo) que si es sobrepasado se producirá inmediatamente una o varias espigas que en conjunto forman una respuesta tónica. Si el voltaje se mantiene por sobre el umbral de burst y por debajo del umbral de respuesta tónica, permanecerá en dicho estado sin generar espigas. Luego queda claro que ante una entrada dada en un instante dado sólo uno de estos tres tipos de respuesta son posibles en este modelo, y cada tipo de respuesta es excluyente de la otra. 47 A. Voltaje de membrana. B. Dinámica de h. C. Diagrama de fase, en el eje X la variable h en el eje Y el voltaje de membrana. Las coordenadas de los puntos son (aprox): Punto 1 (-70,0) Punto 2 (-80,0.85) Punto 3 (-70,1) Punto 4 (30,0.9) Punto 5 (30,0.2) Punto 6 (-55,0) Figura 3.8 Diagrama de fase modelo IFB. 3.2. Características de la respuesta Hasta ahora se ha caracterizado el modelo desde el punto de vista de sistemas dinámicos. Es necesario ahora caracterizarlo desde el punto de vista de su respuesta entrada-salida. Esto se hará frente a entradas de tipo escalón (llamada DC), sinusoidales (llamada AC) y sinápticas. 3.2.1. Modos de respuesta Para los análisis que continúan es conveniente enfatizar que la respuesta en modo tónico se producirá toda vez que el estímulo haga que el voltaje de membrana supere Vθ. Mientras que para que se produzca una descarga tipo burst, como se explicó en la Sección 2.3.1, se debe aplicar una 48 corriente que hiperpolarice el voltaje de membrana bajo Vh y que luego le permita alcanzar la condición V>Vh. Si esto no ocurre y se mantiene el voltaje hiperpolarizado la descarga burst no ocurrirá. En la Figura 3.9 – A se muestra un ejemplo de descarga burst de rebote. Cuando el estímulo es igual a 0.1 [µA] hiperpolariza el voltaje de membrana, en el milisegundo 400 el estímulo aumenta a 0.67 [µA] y se produce la descarga burst. Lo mismo ocurre en la Figura 3.9 – B, la diferencia está en que la descarga se produce en el milisegundo 800 pues en ese instante el estímulo deja de ser hiperpolarizante. A. Estímulo hiperpolarizante (0.67; 0.1; 0.67) el burst se genera en 400 mseg. B. Estímulo hiperpolarizante (0.67; 0.1; 0.67) el burst se genera en 800 mseg. Figura 3.9 Descarga en modo burst. 3.2.1.1. Entradas tipos escalón, DC Para la respuesta tipo escalón se determinará la corriente de reobase y de cronaxia para la respuesta en modo tónico y burst. La corriente de reobase es por definición la mínima corriente necesaria para causar excitación nerviosa, suponiendo que esta corriente dura varios milisegundos. La corriente de cronaxia es un pulso cuadrado de mínima duración necesaria para producir excitación considerando una amplitud de corriente el doble de la reobase [ 14]. En la Figura 3.10 - B se muestra el voltaje de membrana para una corriente de reobase de amplitud máxima 0.8051[µA] con 800[mseg] de duración, esta corriente produce una única espiga. Mientras en la gráfica A de la misma Figura se puede ver como al disminuir la corriente en 0.0001 49 [µA] no se obtiene descarga alguna. En la Figura 3.11 – B se muestra la respuesta para una corriente de 0.9402 [µA] y 145 [mseg] de duración, lo que produce una única y rápida espiga. En contraste en el gráfico A de la misma Figura la duración del pulso se disminuyó a 144[mseg], en este caso no se produce descarga. Las corrientes de reobase y cronaxia aplicadas se muestran en la Figura 3.12. A. Amplitud máxima de la corriente aplicada 0.8050[µA]. Duración del pulso 800 [mseg]. B. Amplitud máxima de la corriente aplicada 0.8051[µA]. Duración del pulso 800[mseg]. Figura 3.10 Corriente de reobase, modo tónico. A. Amplitud mínima de la corriente aplicada 0.9402 [µA]. Duración del pulso 144[mseg]. B. Amplitud mínima de la corriente aplicada 0.9402 [µA]. Duración del pulso 145[mseg]. Figura 3.11 Corriente de cronaxia, modo tónico. 50 Corriente de reobase (línea continua) de 0.8051 [µA] y 800[mseg]. Corriente de cronaxia (línea punteada) 0.9402 [µA] y 145[mseg]. Figura 3.12 Corrientes aplicadas en modo tónico. Para la descarga burst se consideró la corriente de cronaxia igual a menos el doble de la la amplitud de la corriente de reobase de burst. La Figura 3.13 - B y la Figura 3.14 – B, muestran la respuesta a una corriente de reobase de 0.1749 [µA] y 800[mseg] de duración, y para una corriente de cronaxia de -0.3498 [µA] y 86 [mseg] de duración, respectivamente. En la Figura 3.13 – A y la Figura 3.14 – A se grafica la respuesta pasiva obtenida para corrientes de menor amplitud que la de reobase y para corrientes de menor duración que la de cronaxia. En la Figura 3.15 se grafican la corriente de reobase y de cronaxia aplicadas. A. Amplitud mínima de la corriente aplicada 0.1750 [µA]. Duración del pulso 800[mseg]. B. Amplitud mínima de la corriente aplicada 0.1749[µA]. Duración del pulso 800[mseg]. Figura 3.13 Corriente de reobase, modo burst. 51 A. Amplitud mínima de la corriente aplicada -0.3498[µA]. Duración del pulso 85[mseg]. B. Amplitud mínima de la corriente aplicada -0.3498[µA]. Duración del pulso 86[mseg]. Figura 3.14 Corriente de cronaxia, modo burst. Corriente de reobase (línea continua) de 0.1749 [µA] y 800[mseg]. Corriente de cronaxia (línea punteada) 0.3498 [µA] y 86[mseg]. Figura 3.15 Corrientes aplicadas en modo burst. La corriente de reobase para la descarga tónica y en modo burst se determinó para un ancho de pulso de 800[mseg]. En caso de disminuir el ancho del pulso se requiere una mayor amplitud de pulso para producir la misma descarga, el caso extremo aquí definido es la corriente de cronaxia. Si para la corriente de cronaxia se disminuye el ancho del pulso se deberá aumentar la corriente. De la Figura 3.12 y la Figura 3.15 se puede concluir que los parámetros de la corriente de reobase y cronaxia para obtener descargas en modo burst o modo tónico son las indicadas en la Tabla 3.2. 52 MODO DE DESCARGA CORRIENTE DE REOBASE CORRIENTE DE CRONAXIA DURACIÓN AMPLITUD DURACIÓN AMPLITUD Tónico 800 [mseg] 0.8051 [µA] 145 [mseg] 0.9402 [µA] Burst 800 [mseg] 0.1749 [µA] 86 [mseg] -0.3498 [µA] Tabla 3.2 Parámetros de reobase y cronaxia, modo tónico y burst. 3.2.1.2. Entradas sinusoidal, AC Como se vio en la Sección anterior, la respuesta no sólo depende de la amplitud de la señal sino que también de su duración. La Figura 3.16 ilustra cómo afecta la frecuencia de la señal al tipo de respuesta que se pueda obtener. Los gráficos A1 – A3 muestran el estímulo aplicado, la variable h y el voltaje de membrana respectivamente, este estímulo no provoca descargas de ningún tipo. Si se disminuye la frecuencia en 0.1 [Hz] (gráficos B1 – B3), sin modificar los demás parámetros del estímulo, se genera una descarga tónica. Finalmente si se disminuye la frecuencia a 3.5 [Hz] o menos (gráficos C1 – C2), se mantiene la espiga que se había obtenido pero además se generan descargas de modo burst. Para esta Figura se escogió como valor continuo de la señal 0.49 [µA], pues este es el valor medio entre la corriente de reobase de descarga tónica y burst. 53 A1. Estímulo, amplitud DC 0.49[µA], AC 0.5[µA], f = 6.6 [Hz]. B1. Estímulo, amplitud DC 0.49[µA], AC 0.5[µA], f = 6.5 [Hz]. C1. Estímulo, amplitud DC 0.49[µA], AC 0.5[µA], f = 3.5 [Hz]. A2. Variable h. B2. Variable h. C2. Variable h. A3. Voltaje de membrana. B3. Voltaje de membrana. C3. Voltaje de membrana. Figura 3.16 Relación entre respuesta y frecuencia de estímulo. La respuesta tónica ante entradas AC se muestra en la Figura 3.17, donde sólo se varió la frecuencia de la señal entre los gráficos A y B. En la Figura 3.17 – B se muestra que para frecuencias de estímulo menores a 24 [Hz], el modelo responde con descargas tónicas a cada máximo de la sinusoide, para frecuencias mayores o iguales a 24 [Hz] no se alcanza a producir una o más espigas por máximo del ciclo de la señal. En particular para 24 [Hz] la corriente de estímulo tiene 24 máximos (Figura 3.17 – B1) y se producen sólo 22 espigas (Figura 3.17 – B2). 54 A1. Estímulo, amplitud DC 1[µA], AC 0.67 [µA], f = 1 [Hz]. B1. Estímulo, amplitud DC 1[µA], AC 0.67 [µA], f = 24 [Hz]. A2. Voltaje de membrana. B2. Voltaje de membrana. Figura 3.17 Repuesta tónica ante AC. Como se ve en la Figura 3.17 para frecuencias sobre 23 [Hz] la respuesta en voltaje se satura, es decir no sigue aumentando su frecuencia de manera lineal con la frecuencia del estímulo. El caso extremo se muestra en la Figura 3.18 donde se verifica que para frecuencias iguales (Figura 3.18 - B) o mayores a 130 [Hz] (Figura 3.18 - C), la frecuencia de descarga es la misma que en el caso en que la corriente aplicada es continua de máximo 1 [µA]. Esto indica que para frecuencias sobre los 130 [Hz] deja de responder a la sinusoidal y considera como estímulo sólo la componente continua de la corriente aplicada. 55 A1. Estímulo, amplitud DC 1[µA], sin componente AC. A2. Voltaje de membrana, de 19 espigas. A1 A2 B1. Estímulo, amplitud DC 1[µA], AC 0.67 [µA], f = 130 [Hz]. B2. Voltaje de membrana, de19 espigas. B1 B2 C1. Estímulo, amplitud DC 1[µA], AC 0.67 [µA], f = 300 [Hz]. C2. Voltaje de membrana, de 19 espigas. C1 C2 Figura 3.18 Repuesta tónica ante AC y DC. La respuesta en burst ante corrientes sinusoidales es mostrada en la Figura 3.19, donde se observan 3 casos donde la corriente de estímulo es siempre de igual amplitud y componente continua, lo único que cambia entre un ejemplo y otro es la frecuencia de la sinusoidal. En la Figura 3.19 – A se puede ver un caso en que la sinusoide es de baja frecuencia ocurriendo 5 mínimos de la señal en el gráfico (Figura 3.19 – A1), de la Figura 3.19 – A2 se pude ver que se generan los 5 estallidos correspondientes. Se determinó que para un estímulo de amplitud AC 0.67 [µA] la máxima frecuencia de estímulo es 5 [Hz], para frecuencias mayores el número de estallidos en la señal de voltaje no tiene una correspondencia uno a uno con los mínimos de la señal de estímulo. El caso extremo se presenta en la Figura 3.19 – B, para una frecuencia de 31 56 [Hz] el modelo responde como si la señal de entrada fuese una constante, es decir genera un único estallido de rebote al liberarlo del estímulo hiperpolarizante. Esta situación se revierte para frecuencias sobre 35 [Hz] hasta 40 [Hz] donde responde con varios estallidos a la señal sinusoidal, Figura 3.19 – C. Al continuar aumentando la frecuencia de la señal hasta 110 [Hz] se alternaron las respuestas B3 y C3, obteniéndose para 30, 45, 60, 85 y 110 [Hz] la respuesta del gráfico B3. A1. Estímulo, amplitud DC nula, AC 0.67[µA], f = 5 [Hz]. B1. Estímulo, amplitud DC nula, AC 0.67[µA], f = 31 [Hz]. C1. Estímulo, amplitud DC nula, AC 0.67[µA], f = 35 [Hz]. A2. Variable h. B2. Variable h. C2. Variable h. A3. Voltaje de membrana. B3. Voltaje de membrana. C3. Voltaje de membrana. Figura 3.19 Respuesta burst ante AC. Se concluye de esta sección que frente a un estímulo sinusoidal de amplitud DC 1 [µA] y AC 0.67 [µA], la frecuencia máxima bajo la cual no se satura la descarga tónica es de 23 [Hz]. Para una 57 señal sinusoidal de amplitud AC 0.67 [µA] y DC nula, la frecuencia máxima sobre la cual se satura la señal es de 5 [Hz]. 3.2.1.3. Entradas sinápticas Las entradas tipo escalón y sinusoidales puras, como las presentadas en la Sección 3.2.1.1 y 3.2.1.2 sólo son posibles cuando se trabaja in vitro, es decir se pueden realizar sólo dentro del laboratorio. La corriente de estímulo que recibe una neurona in vivo es más ruidosas, pues una neurona puede tener miles de contactos sinápticos que la estimulan. Así la corriente de estímulo que ve la neurona es una suma simple de todas las entradas sinápticas. Para caracterizar la corriente sináptica se considerará el modelo de corrientes sinápticas sintéticas propuesto en [ 60]. Este considera que una corriente sináptica unitaria (CSU) es una curva de decaimiento exponencial, ver Figura 3.20, pues cuando un contacto sináptico libera neurotransmisores estos se difunden en el medio cercano. Por otro lado la llegada u ocurrencia de CSU obedece a un proceso de Poisson como los mostrados en la Figura 3.21. Finalmente la corriente sináptica sintética que vería una neurona es la mostrada en la Figura 3.22, donde se consideró una tasa de ocurrencia de 2.4 llegadas por milisegundo. Corriente sináptica unitaria. Amplitud máxima 0.1, constante de tiempo 3 [mseg]. Figura 3.20 Corriente sináptica. 58 A. Tasa de eventos λ = 2.4 sucesos/milisegundo B. Tasa de eventos λ = 0.6 sucesos/milisegundo Figura 3.21 Proceso de llegadas de Poisson. A. CSU de 0.05[µA] y τ = 3[mseg]. Ocurrencia de CSUs con λ = 2.4 sucesos/milisegundo. Media de la corriente 0.41[µA]. B. CSU de 0.1[µA] y τ = 3[mseg]. Ocurrencia de CSUs con λ = 2.4 sucesos/milisegundo. Media de la corriente 0.86[µA]. Figura 3.22 Corriente de estímulo. En la Figura 3.23 se muestra la respuesta tónica del modelo. En los casos A y B los parámetros son los mismos y las diferencias se deben al proceso de Poisson. En la Figura 3.24 se puede ver cómo la corriente sináptica produce una descarga tónica de frecuencia irregular. Mientras que en la Figura 3.25 se pude ver cómo bajo los mismos parámetros de corriente se puede desencadenar o no un estallido. 59 A1 B1 A2 B2 Componente DC -0.12[µA]. CSU de 0.1[µA] y τ = 3[mseg]. λ = 2.4 sucesos/milisegundo. Figura 3.23 Respuesta tónica frente a entrada sináptica sintética. Entrada de componente DC 0.1[µA]. CSU de 0.1[µA] y τ = 3[mseg]. λ = 2.4 sucesos/milisegundo. Figura 3.24 Respuesta tónica de frecuencia irregular. 60 A1 B1 A2 B2 Componente DC -0.12[µA]. CSU de 0.1[µA] y τ = 3[mseg]. λ = 2.4 sucesos/milisegundo. Figura 3.25 Respuesta burst frente a entrada sináptica sintética. 3.2.1.4. Entrada DBS A continuación se muestra la respuesta del modelo ante entrada tipo DBS. Como se indicó en la sección 1.1, el rango de parámetros para DBS aplicado a desórdenes de movimiento es 2-4 [V] (que equivale a 2-4 [mA] para una impedancia de electrodo de 1000 [Ω]), anchos de pulso entre 90 y 180 [µs] y 100-185 [Hz] [ 1]. En la Figura 3.26 y Figura 3.27 se puede ver la repuesta del modelo ante entradas tipo tren de pulsos, la frecuencia y el ancho del pulso se fijaron dentro de los valores usados para DBS en desórdenes de movimiento. No se pudo fijar el valor de la corriente pues se desconoce cuál es el valor que ve cada neurona proveniente del electrodo, además dependerá de la distancia entre la neurona y éste, por lo que es difícil de estimar en este caso. Debido a esto último se estudió cuál era la corriente mínima que se debía aplicar para producir descargas en modo tónico (ver Figura 3.26) y en modo burst (ver Figura 3.27). En este último caso era difícil determinar a partir del 61 voltaje si el modelo estaba produciendo descargas en modo burst, por ello se determinó a partir de la variable h el modo de descarga. A1. Estímulo, amplitud DC 0.67[µA], amplitud del pulso 5[µA], ancho 100 [µseg] y 182 [Hz]. A2. Voltaje de respuesta pasiva. A1 membrana, A2 B1. Estímulo, amplitud DC 0.67[µA], amplitud del pulso 6.97[µA], ancho 100 [µseg] y 182 [Hz]. B2. Voltaje de membrana, 1 espiga. B1 B2 C1. Estímulo, amplitud DC 0.67[µA], amplitud del pulso 9[µA], ancho 100 [µseg] y 182 [Hz]. C2. Voltaje de membrana, 6 espigas. C1 C2 Figura 3.26 Repuesta tónica entrada de alta frecuencia. De los ejemplos de la Figura 3.26 se puede ver que la relación entre la amplitud del estímulo y la respuesta es lineal. En el gráfico A de la Figura 3.26 se puede ver, de la forma de onda del voltaje, que el tren de pulsos de alta frecuencia está actuando como un equivalente de entrada constante, es decir el modelo no ve que el estímulo sube y baja, pues la depolarización de la señal comienza en el instante en que se aplica el estímulo (50 [mseg]) y termina en el instante en que cesa el estímulo de alta frecuencia (850 [mseg]). Luego para un estímulo de alta frecuencia aplicado por 800 [mseg], de ancho de pulso 10 [µseg] y frecuencia 182 [Hz], la amplitud de 62 corriente de reobase se fija con un máximo de 7.64 [µA] (máximo del pulso más componente DC) y mínimo de 0.67 [µA] (mínimo del pulso es 0 más la amplitud DC). En el caso de la descarga en modo burst, se fijó la corriente DC en 0.3 [µA], un ancho de pulso de 100 [µseg] y una frecuencia de 182 [Hz], así la corriente de reobase tiene un mínimo de 0.3 [µA] y un máximo de -10.3 [µA]. A1. Estímulo, amplitud DC 0.3 [µA], amplitud del pulso -8[µA], ancho 100 [µseg] y 182 [Hz]. A2. Voltaje de membrana. A1 A2 B1. Estímulo, amplitud DC 0.3 [µA], amplitud del pulso 10[µA], ancho 100 [µseg] y 182 [Hz]. B2. Voltaje de membrana. B1 B2 C1. Estímulo, amplitud DC 0.3 [µA], amplitud del pulso 15[µA], ancho 100 [µseg] y 182 [Hz]. C2. Voltaje de membrana. C2 C1 Figura 3.27 Respuesta modo burst ante entrada de alta frecuencia. En la Figura 3.27 – B, para determinar si la descarga era en modo burst se consideró el valor de la variable h. En este caso la variable era no nula pero de un valor mucho menor que 0.1, en la Figura 3.28 - A se muestra el voltaje de membrana cuando la amplitud del pulso es de -14 [µA], y 63 en la misma Figura gráfico B se muestra la variable h correspondiente, se puede ver que cuando se produce la primera descarga que es de corta duración (como en la Figura 3.27 - B) el valor de h es menor que 0.1, en cambio cuando se produce la segunda descarga burst la variable h alcanza una valor cercano a 1. A. Voltaje de membrana frente a un estímulo de amplitud DC 0.3 [µA], amplitud del pulso -14[µA], ancho 100 [µseg] y 182 [Hz]. B. Variable h. B A Figura 3.28 Respuesta burst ante entrada de alta frecuencia. De la Figura 3.27 se concluye que para amplitudes de pulso menores (más negativos) o iguales a -15 [µA] la respuesta del modelo es equivalente al caso en que recibe un estímulo hiperpolarizante constante de 800 [mseg] de duración. Si para el caso de la Figura 3.26 – C se varía la frecuencia de la señal dejando fija la amplitud del pulso en 9 [µA], el ancho del pulso en 100 [µseg] y la amplitud DC en 0.67 [µA], se obtienen los resultados de la Figura 3.29. Esta Figura muestra cómo incide la frecuencia del estimulo en el modo de respuesta, estableciéndose que al disminuir la frecuencia del estímulo va disminuyendo la depolarización que produce, lo que se traduce en una menor frecuencia de descarga. A. Voltaje de membrana. Estímulo, amplitud DC 0.67[µA], amplitud del pulso 9[µA], ancho 100 [µseg] y 100 [Hz] 64 B. Voltaje de membrana. Estímulo, amplitud DC 0.67[µA], amplitud del pulso 9[µA], ancho 100 [µseg] y 120 [Hz] C. Voltaje de membrana. Estímulo, amplitud DC 0.67[µA], amplitud del pulso 9[µA], ancho 100 [µseg] y 140 [Hz] D. Voltaje de membrana. Estímulo, amplitud DC 0.67[µA], amplitud del pulso 9[µA], ancho 100 [µseg] y 160 [Hz] Figura 3.29 Relación frecuencia de estímulo frecuencia de descarga. 65 Capítulo 4 Diseño y Aplicación de la Estrategia de Control En este Capítulo se diseña y aplica un sistema de control para el modelo IFB de la neurona talámica. El objetivo de este sistema de control es que considerando 2 neuronas independientes, cada una modelada por modelos IFB de iguales parámetros, el sistema determina la acción de control que es necesario aplicar para que ambas descarguen en el mismo modo. Así se definirá una neurona como Neurona de Referencia, de ahora en adelante llamada Referencia, y la otra como Neurona a Controlar, de ahora en adelante llamada Planta. La Referencia definirá el modo de descarga que se desea tenga la neurona Planta, es decir, el control debe ser tal que permita que la neurona Planta siga el modo de descarga que posee la neurona de Referencia. En un sistema real (aplicación in vitro del sistema de control) no es posible medir la corriente sináptica que estimula a una determinada neurona, sólo se puede tener acceso al voltaje de membrana de la misma. Es así como este sistema de control debe ser capaz de definir la acción de control necesaria considerando como única variable medible el voltaje de membrana intracelular (i.e. dentro de la neurona). 66 4.1. Diseño de la estrategia de control El sistema de control propuesto está compuesto de 2 bloques, tal como se muestra en la Figura 4.1. El Bloque 1 o Clasificador debe determinar el modo respuesta de la neurona de Referencia y Planta, a partir de la señal de voltaje de cada una. Este bloque puede arrojar 4 respuestas; que la neurona está en modo burst, modo tónico, respuesta pasiva o indeterminada. El Bloque 2 o Controlador debe determinar, en base a la información del Bloque 1 sobre cada neurona, si es necesario aplicar una acción de control sobre la neurona Planta. De ser así estima la corriente estímulo que debe estar recibiendo la neurona Planta y calcula la acción de control que se le debe aplicar para llevarla al modo de descarga de la neurona de Referencia. En la Figura 4.1 se muestra un esquema del sistema de control propuesto. Figura 4.1 Esquema del sistema de control propuesto. 67 El sistema diseñado tiene por objetivo que dos neuronas en distinto modo de descarga, descarguen en igual modo como consecuencia de la acción de control aplicada. En este contexto se considerará como modos de respuesta las descargas tónicas y los estallidos. Por lo que toda vez que alguna de las 2 neuronas o ambas simultáneamente generen una respuesta pasiva, no se considerará como una situación que se deba controlar. El sistema de control aquí propuesto se basa en registros intracelulares (que son sólo posibles in vitro), los cuales permiten distinguir las respuestas pasivas de los otros modos de descarga, pero se debe considerar aquí que in vivo (en organismos completos) sólo se tiene acceso a los registros extracelulares. Considerando esto último, son 2 los motivos para no aplicar el controlador cuando alguna de las neuronas está en respuesta pasiva: El primero es que en registros extracelulares (i.e. aquellos en que el electrodo está fuera de la neurona, posibles en organismos vivos) se distingue el modo burst del tónico mediante métodos estadísticos [ 62], ver Figura 4.2, donde dentro de los períodos análizados hay lapsos de tiempo en que la respuesta es pasiva, sin significar ello que el registro completo se clasifique como tal. El segundo motivo para no especificar un control para la respuesta en modo pasivo, es que son las descargas tónicas y en estallido, además de la periodicidad con que ocurren, las que transmiten información, por lo que son estos modos de descarga los que se asocian a patologías y disfuncionalidades.[ 61] de la vía talámica. Registro extracelular de descarga espontanea de dos neuronas del núcleo lateral geniculado de un mono despierto. Los asteriscos (registro en rojo) muestran 3 burst, los demás potenciales de acción son descargas tónicas. Figura 4.2 Registro extracelular de descargas. Adaptada de [ 61]. 4.1.1. Diseño del Bloque 1 El Bloque 1 o Clasificador debe determinar el modo de respuesta de la neurona Planta y Referencia a partir de la información de voltaje de cada neurona. Para identificar el modo de respuesta se consideró el hecho de que la respuesta en modo burst, pasivo o tónico dependerá del 68 nivel de voltaje de membrana. Como se estableció en la Sección 2.3.1, Figura 2.14, existen dos voltajes límite, uno bajo el cual se gatilla el burst y otro sobre el cual la descarga es tónica. Entre ambos límites la respuesta es pasiva. Se estableció como criterio para determinar el modo de respuesta el nivel de voltaje. Si el vector de voltaje tiene algún coeficiente sobre el valor umbral para gatillar potenciales de acción, entonces se considera que la respuesta es tónica. Si tiene valores bajo el nivel que gatilla un burst se considera que la neurona está en modo burst Si el vector de voltaje tiene sus valores bajo el umbral de espigas tónicas y sobre el de burst, se considera en respuesta pasiva. Bastará con que exista un único valor del vector de voltaje sobre el umbral tónico para que clasifique la neurona como en descargas tónicas, de igual modo si existe un valor bajo el umbral burst se determina que el modelo está descargando en modo estallido. Los coeficientes del vector de voltaje se leen en orden temporal, es decir, desde el valor más antiguo hasta el más reciente. En esta lectura secuencial cuando un coeficiente supera el umbral es suficiente para clasificar la respuesta en modo burst o tónico, según el umbral superado. G G Traducir estas reglas a variables del modelo, considerando V un vector de voltaje y V (i ) el elemento i-ésimo de éste, significa que: G Si V (i ) ≤ Vh entonces respuesta está en modo burst. G Si V (i ) ≥ Vθ entonces la respuesta es de descarga tónica. G Si Vh < V (i ) < Vθ entonces la respuesta es pasiva. Si bien por la formulación del modelo usado los voltajes límites son directos para cada modo de descarga, en el caso general es posible realizar pruebas para determinar el voltaje umbral Vθ y Vh. 4.1.2. Diseño del Bloque 2 El Bloque 2 o Controlador determina la acción de control que se debe aplicar toda vez que el modo de descarga de la Planta sea distinto al de la Referencia. En ese caso, a partir del vector de 69 G G voltaje de la Referencia y la Planta, V REF y VPLA , se estima la corriente que debe estar estimulando a cada neurona, IEST_REF e IEST_PLA, y se define la acción de control como: I CONTROL =I EST _ REF − I EST _ PLA (4.1) Para estimar la corriente de estímulo de las neuronas, no fue posible determinar una regla que se aplicara para ambos modos de respuesta, por lo que se definió un estimador para el modo burst y otro para el modo tónico. El estimador de corriente de modo tónico considera el hecho que, en general, las neuronas codifican en frecuencia, es decir a mayor amplitud de estímulo más alta es la frecuencia de respuesta [ 11]. En la Figura 4.3 se ejemplifica esta propiedad de las neuronas. A. Corriente de estímulo 0.67 – 0.9 -0.67 [µA], lo cambios se producen en 200 [mseg] y 800 [mseg]. B. Corriente de estímulo 0.67 – 1.1 -0.67 [µA], lo cambios se producen en 200 [mseg] y 800 [mseg]. Figura 4.3 Codificación en frecuencia. La Figura 4.4 muestra la relación entre frecuencia de descarga y amplitud del estímulo, para el modelo de neurona usado. 70 Aproximación cuadrática (línea gruesa) y lineal (línea delgada) de la relación entre frecuencia de la descarga y corriente de estímulo. Figura 4.4 Relación frecuencia amplitud, modo tónico. Como se observa de la Figura 4.4 la aproximación cuadrática se ajusta mejor a los datos de baja frecuencia. Luego, si se conoce la frecuencia media de la señal f med se puede calcular la corriente de estímulo como muestra la ecuación (4.2). La frecuencia media, f med , se obtiene como el inverso del promedio del tiempo entre espigas para una señal dada. ~ 2 I TONICA = 40.2 * f med + 16.85 * f med + 0.67 (4.2) Para el caso de la respuesta en modo burst, el estimador de corriente se basa en el hecho que la relación entre la amplitud de la señal y la frecuencia media de descarga no es significativa, como en el caso de la respuesta tónica (ver Figura 4.5). Lo que cambia en la respuesta en modo burst en directa relación con la amplitud del estímulo es la derivada del voltaje, tal como se puede ver en la Figura 4.6 – A, donde se observa como disminuye el voltaje de hiperpolarización alcanzado. En la Figura 4.6 (B) se puede ver con más detalle la curva que cae al voltaje de hiperpolarización. 71 Frecuencia versus amplitud máxima de corriente Imax entre -2 y 2. Se pude ver que hay una gran diferencia entre la máxima y la mínima frecuencia de las señales cuando está en modo burst Imax <0.1, zona anaranjada. Mientras que en modo tónico la diferencia entre dichas corrientes es mucho menor Imax > 0.8, zona verde. Figura 4.5 Relación frecuencia amplitud de estímulo. B A Figura 4.6 Respuesta en modo burst para varias amplitudes de estímulo. Para establecer una relación entre la deriva algebraica del voltaje y la amplitud del estímulo, se definió un valor límite para la derivada. Es decir, se considerará para la estimación de la corriente la derivada del voltaje en el punto en que es menor o igual que -0.25 [mV]. Esta restricción sobre la derivada del voltaje asegura una relación lineal entre la corriente y la derivada del voltaje. Si bien se podría haber considerado otra cota para la derivada, fue con este valor que se consiguió una relación lineal que fuera independiente del error numérico cometido al hacer el cálculo de la estimación. En la Figura 4.7 se muestra la relación entre la derivada del voltaje y la corriente de estímulo. A partir de la regresión lineal de los datos del gráfico se obtiene que la corriente en modo burst se puede estimar como (4.3). 72 Regresión lineal sobre los datos que relacionan la derivada del voltaje con la corriente de estímulo que recibe la neurona en modo burst. Figura 4.7 Relación dV/dt y amplitud, modo burst. ~ I BURST = 2 * V + 0.6 (4.3) 4.2. Resultado Simulaciones En esta sección se presentan los resultados al aplicar el sistema de control, descrito en la sección anterior, ante corrientes de estímulo DC, AC y sinápticas sintéticas. En todas las simulaciones las condiciones iniciales para la neurona de Referencia y de Planta son V = -60 [mV] y h = 0. En el Anexo B se presenta el código para Matlab 6.5 que implementa el control para las Secciones 4.2.1, 4.2.2 y 4.2.3. En estos casos la acción de control se calcula después de los 200 [mseg] de simulación, a partir de la información de voltaje acumulada hasta ese momento. Una vez calculada la corriente ICONTROL que corresponde aplicar , se mantiene en este valor hasta el fin de la simulación, es decir no se verifican los resultados obtenidos a partir de dicha acción de control. 4.2.1. Control ante estímulos DC En este caso se definirá la acción de control como una corriente tipo tren de pulsos que variará entre 0.49 [µA] e ICONTROL, con un ancho de pulso de 300 [mseg] y un ciclo de 600 [mseg]. 73 El valor de ICONTROL lo determina el controlador restando a la corriente de la neurona de Referencia, IREF, la corriente de Planta, IPLA (ver Figura 4.1). El valor de 0.49 [µA] se define pues es la media entre la corriente de reobase de descarga tónica y burst, se escogió este valor y no 0 [µA], pues a 0 [µA] ya se produce una descarga en modo burst. Para las simulaciones de esta sección las corrientes de estímulo de la Planta se aplicaron desde el milisegundo 50 hasta el 2500, para la Referencia la corriente se aplicó desde el milisegundo 50 hasta el 1000. El tiempo total de las simulaciones es de 4000 [mseg]. En la Figura 4.8 se muestra un ejemplo de las corrientes de estímulo aplicadas a la Referencia (línea oscura) y a la Planta (línea clara). Línea oscura, corriente de estímulo aplicada a la Referencia entre 50 – 1000 [mseg]. Línea clara, corriente de estímulo aplicada a la Planta entre 50 – 2500 [mseg]. Figura 4.8 Ejemplo de corrientes de estímulo DC. En la Figura 4.9 se muestra el voltaje de membrana de la neurona Referencia y Planta en lazo abierto (cuando no se aplica acción de control) para dos corrientes IPLA distintas, la Referencia está en modo burst y la Planta en modo tónico. En la Figura 4.10 se muestra el resultado al aplicar el sistema de control en el caso A y B de la Figura 4.9. En el milisegundo 200 se aplica la acción de control a la Planta y comienza a descargar en modo burst. A. Referencia en modo burst, IREF = 0.1 [µA]. Planta en modo tónico, IPLA = 0.9 [µA]. B. Referencia en modo burst, IREF = 0.1 [µA]. Planta en modo tónico, IPLA = 1.1 [µA]. Figura 4.9 Voltaje de membrana en lazo abierto. 74 A1. Estímulo y corriente de control. IEST_REF = 0.0949 [µA], IEST_PLA = 0.8842 [µA]; IREF = 0.1 [µA], IPLA = 0.9 [µA]. B1. Estímulo y corriente de control. IEST_REF = 0.0949 [µA], IEST_PLA = 1.1049 [µA]; IREF = 0.1 [µA], IPLA = 1.1 [µA]. A2. Voltaje de membrana. B2. Voltaje de membrana. A3. Variable h. B3. Variable h. Figura 4.10 Control de modo tónico a burst, estímulo DC. 75 A continuación se aplica el sistema de control al caso en que la Referencia se encuentra en modo tónico y la Planta en modo burst. En la Figura 4.11 se muestra la respuesta de cada neurona en lazo abierto, en la Figura 4.11 – A la planta es estimulada por una corriente de 0.1 [µA], en B es estimulada por una corriente de -0.2 [µA]. En la Figura 4.12 se muestra el resultado obtenido al aplicar el sistema de control a estos ejemplos. A. Referencia en modo tónico, IREF = 0.9 [µA]. Planta en modo burst, IPLA = 0.1 [µA]. B. Referencia en modo tónico, IREF = 0.9 [µA]. Planta en modo burst, IPLA = -0.2 [µA]. Figura 4.11 Voltaje de membrana en lazo abierto. 76 A1. Estímulo y corriente de control. IEST_REF = 0.8842 [µA], IEST_PLA = 0.0949 [µA]; IREF = 0.9 [µA], IPLA = 0.1 [µA]. B1. Estímulo y corriente de control. IEST_REF = 0.8842 [µA], IEST_PLA = -0.2025 [µA]; IREF = 0.9 [µA], IPLA = -0.2 [µA]. A2. Voltaje de membrana. B2. Voltaje de membrana. A3. Variable h. B3. Variable h. Figura 4.12 Control de modo burst a tónico, estímulo DC. 77 Como se puede apreciar de la Figura 4.10 y Figura 4.12, el sistema de control cumple el objetivo de modificar el modo de respuesta de la neurona Planta para llevarlo al modo de la Referencia. Pero en todos estos casos se puede ver que en el milisegundo 2500, cuando varía el estímulo de la Planta, el sistema de control, que se mantiene en el valor determinado para las corrientes estimadas en el milisegundo 500, no es el apropiado. Si bien en los 4 casos mantiene la respuesta de la Planta en el modo de respuesta de la Referencia, en el caso tónico (ver Figura 4.12) la frecuencia de descarga de la Planta no es la misma frecuencia de la Referencia pasado los 2500 [mseg]. Lo mismo ocurre para cuando se lleva la Planta al modo burst (ver Figura 4.10), donde después del milisegundo 2500 la corriente de control hace que la hiperpolarización previa al estallido sea menor de lo que es la hiperpolarización de la Referencia. En la Figura 4.13 se muestran 3 casos en que no se genera acción de control. En A no se genera acción de control pues ambas neuronas presentan una respuesta pasiva, en B no se aplica el control pues la Referencia está en modo pasivo, en el caso C no se genera acción de control pues ambas neuronas están descargando en el mismo modo. 78 A1. Estímulo y corriente de control. IREF = 0.7 [µA], IPLA = 0 [µA]. A2. Voltaje de membrana. B1. Estímulo y corriente de control. IREF = 0.9 [µA], IPLA = 0.3 [µA]. B2. Voltaje de membrana. C1. Estímulo y corriente de control. IREF = 1 [µA], IPLA = 0.9 [µA]. C2. Voltaje de membrana. Figura 4.13 Ejemplo de casos en que no se genera acción de control, estímulo DC. 79 4.2.2. Control ante estímulos AC En este caso la corriente de estímulo no se supuso constante, sino que sinusoidal. La corriente de control (ICONTROL) se definió como un tren de pulsos de 0.49 [µA] como en la Sección anterior, con la diferencia que el ancho del pulso es de 300 [mseg] y el ciclo del pulso es de 400 [mseg] En la Figura 4.14 se muestra un ejemplo de las corrientes de estímulo. Línea oscura, corriente de estímulo aplicada a la Referencia. Línea clara, corriente de estímulo aplicada a la Planta. Figura 4.14 Estímulos sinusoidales aplicados. En la Figura 4.15 se muestra el voltaje de membrana sin control para dos corrientes de estímulo distintas, en ambos casos la Referenica está en modo burst y la Planta en modo tónico. En la Figura 4.16 se muestra el resultado al aplicar el sistema de control a los ejemplos de la Figura 4.15. Del mismo modo para el caso en que la Referencia está descargando en modo tónico y la Planta en modo burst se aplica el sistema de control, los resultados se muestran en la Figura 4.17. A. Referencia en modo burst, amplitud DC del estímulo 0.1 [µA], AC 0.5 [µA], frecuencia 1.9 [Hz]. Planta en modo tónico, amplitud DC del estímulo 0.9 [µA], AC 0.5 [µA] y frecuencia 1.9 [Hz]. B. Referencia en modo burst, amplitud DC del estímulo 0.1 [µA], AC 0.5 [µA], frecuencia 1.4 [Hz]. Planta en modo tónico, amplitud DC del estímulo 0.9 [µA], AC 0.5 [µA] y frecuencia 1.4 [Hz]. Figura 4.15 Repuesta en lazo abierto, estímulo AC. 80 A1. Estímulo y corriente de control. IEST_REF = 0.1849 [µA], IEST_PLA = 1.3355 [µA]. Referencia: DC 0.5 [µA], AC 0.1 [µA], 1.9 [Hz]. Planta: AC 0.9 [µA], igual DC y f. B1. Estímulo y corriente de control. IEST_REF = 0.1849 [µA], IEST_PLA = 1.3355 [µA]. Referencia: DC 0.5 [µA], AC 0.1 [µA], 1.4 [Hz]. Planta: AC 0.9 [µA], igual DC y f. A2. Voltaje de membrana. B2. Voltaje de membrana. A3. Variable h. B3. Variable h. Figura 4.16 Control del modo tónico al burst, estímulo AC. 81 A1. Estímulo y corriente de control. IEST_REF = 1.3355 [µA], IEST_PLA = 0.1849 [µA]. Referencia: DC 0.5 [µA], AC 0.9 [µA], 1.9 [Hz]. Planta: AC 0.1 [µA], igual DC y f. B1. Estímulo y corriente de control. IEST_REF = 1.3267 [µA], IEST_PLA = 0.1832 [µA]. Referencia: DC 0.5 [µA], AC 0.9 [µA], 1.9 [Hz]. Planta: AC 0.1 [µA], igual DC y f. A2. Voltaje de membrana. B2. Voltaje de membrana. A3. Variable h. B3. Variable h. Figura 4.17 Control del modo burst al tónico, estímulo AC. 82 A1. Estímulo y corriente de control. Referencia: DC 0.3 [µA], AC 0.5 [µA], 1.9 [Hz]. Planta: DC 0.3 [µA], AC 0.5 [µA], 1.4 [Hz]. A2. Voltaje de membrana. B1. Estímulo y corriente de control. Referencia: DC 0.5 [µA], AC 0.9 [µA], 1.9 [Hz]. Planta: AC 0.1 [µA], igual DC y frecuencia. B2. Voltaje de membrana. C1. Estímulo y corriente de control. Referencia: DC 0.5 [µA], AC 0.9 [µA], 1.9 [Hz]. Planta: DC 0.5 [µA], AC 1.1 [µA], 1.4 [Hz]. C2. Voltaje de membrana. Figura 4.18 Ejemplo de casos en que no se genera acción de contro, estímulo AC. 83 En la Figura 4.18 se muestra el caso en que no se genera acción de control pues ambas neuronas presentan una respuesta pasiva (A), un caso (B) donde no se aplica acción de control pues la Referencia está en modo pasivo y otro caso (C) donde no se genera la acción de control pues ambas neuronas están descargando en modo tónico. En la Figura 4.16 se vio el caso en que la Planta estaba descargando en modo tónico y la Referencia en estallidos para una señal de frecuencia 1.9 (Figura 4.16 – A) y 1.4 [Hz] (Figura 4.16 – B). En ambos casos la acción de control logró que la Planta comenzara a descargar estallidos a partir del milisegundo 500 (cuando se inició la acción de control). En ambos casos la fase de hiperpolarización del potencial previo al estallido fue irregular, ver Figura 4.16 A2 – A3 y B2 – B3, debido a que la señal del pulso de control estaba en desfase con la señal sinusoidal de estímulo. En ninguno de los casos esto significó perder el objetivo de control. Para el control del modo burst se muestran 2 casos de distinta frecuencia en la Figura 4.17, en ambos casos se consiguió el objetivo de control, llevar la Planta a descargas burst de frecuencia similar a la de la Referencia, ver Figura 4.17 – A y B3. Nuevamente, debido a que la señal de control no está en fase con la señal sinusoidal de estímulo la periodicidad del conjunto de descargas tónicas en la Planta (un conjunto de descargas por ciclo del estímulo) no es la misma que en la Referencia. Se verificó además que el sistema de control no se aplica en los casos en que la Referencia o Planta, o ambas, están en respuesta pasiva (ver Figura 4.18 – A y B), o están en igual modo descarga, ver Figura 4.18 – C. 4.2.3. Control ante estímulos sinápticos sintéticos Para las simulaciones de esta sección las corrientes sinápticas sintéticas (ISS) se generaron como se indicó en la Sección 3.2.1.3. La entrada a la Planta se aplicó hasta el milisegundo 2500, mientras que la entrada a la Referencia se aplicó hasta el milisegundo 1000. En la Figura 4.19 se muestra un ejemplo en que controla el modo tónico de la Planta, mientras en la Figura 4.20 se muestra el caso en que controla la repuesta en modo burst. 84 En el caso del control del modo tónico (Figura 4.19) se muestran 2 ejemplos, en la Figura 4.19 – A la amplitud de la corriente sináptica unitaria (CSU) de la corriente sináptica es de 0.1 [µA], mientras que en el caso de la Figura 4.19 – B la amplitud del CSU es de 0.05 [µA], en este último caso se debió aumentar además la amplitud de la corriente DC sobre la que se montó la corriente sináptica de la Planta, para que se provocara la descarga tónica de espigas. En ambos casos se consiguió el objetivo de control (ver Figura 4.19 – A3 y B3) de llevar la Planta al modo burst. La hiperpolarización del voltaje de membrana de la Planta en ninguno de los 2 casos corresponde con la hiperpolarización del voltaje de la Referencia, esto se debe al carácter aleatorio del proceso de Poisson por el que se generan las corrientes de estímulo, comparar Planta y Referencia de la Figura 4.19 – A2 y B2. Si se compara la corriente estimada para la Planta y la Referencia en cada caso, se observa que el estimador de corriente en respuesta tónica hace una estimación mucho mejor que el de respuesta en modo burst para este tipo de entrada, ver Tabla 4.1. Esto se debe a que el estimador en modo tónico usa la frecuencia media de la señal de voltaje disponible, mientras que el estimador de modo burst al usar sólo un punto de la señal derivada de voltaje es más sensible a las variaciones de la señal. CASO A CASO B REFERENCIA PLANTA REFERENCIA PLANTA CORRIENTE ESTIMADA -0.6982 1.0316 -0.6982 0.9196 CORRIENTE MEDIA 0.1102 1.0525 -0.2749 0.9209 Tabla 4.1 Comparación de estimadores, estímulo ISS. 85 A1. Estímulo y corriente de control. IEST_REF = -0.6982 [µA], IEST_PLA = 1.0316 [µA]. Amplitud CSU Referencia 0.1 [µA], DC -0.7 [µA], media 0.1102 [µA], amplitud CSU Planta 0.1 [µA], DC 0.2 [µA], media 1.0525 [µA]. B1. Estímulo y corriente de control. IEST_REF = -0.6982 [µA], IEST_PLA = 0.9196 [µA]. Amplitud CSU Referencia 0.05 [µA], DC -0.7 [µA], media -0.2749 [µA], amplitud CSU Planta 0.05 [µA], DC 0.5 [µA], media 0.9209 [µA]. A2. Voltaje de membrana. B2. Voltaje de membrana. A3. Variable h. B3. Variable h. Figura 4.19 Control del modo tónico a burst, estímulo ISS. 86 A1. Estímulo y corriente de control. IEST_REF = 1.0370 [µA], IEST_PLA = -0.6982 [µA]. Amplitud CSU Referencia 0.1 [µA], DC 0.2 [µA], media 1.0539 [µA], amplitud CSU Planta 0.1 [µA], DC -0.7 [µA], media 0.1484 [µA]. B1. Estímulo y corriente de control. IEST_REF = 0.8989 [µA], IEST_PLA = -0.6982 [µA]. Amplitud CSU Referencia 0.05 [µA], DC 0.5 [µA], media 0.9326 [µA], amplitud CSU Planta 0.05 [µA], DC -0.7 [µA], media -0.2771 [µA]. A2. Voltaje de membrana. B2. Voltaje de membrana. A3. Variable h. B3. Variable h. Figura 4.20 Control del modo burst a tónico, estímulo ISS. El caso en que la Planta se encuentra en modo burst y se quiere llevar al modo tónico se muestra en la Figura 4.20. Se puede ver que para los 2 ejemplos de dicha Figura se cumple el 87 objetivo de control (ver Figura 4.20 – A3 y B3), pero no se consigue que la frecuencia de descarga de la Planta sea similar a la frecuencia de descarga de la Referencia. Esto pues, al igual que el caso en que se controla el modo tónico, la estimación de corriente en el modo burst es sensible a las variaciones del estímulo. CASO A CASO B REFERENCIA PLANTA REFERENCIA PLANTA CORRIENTE ESTIMADA 1.0370 -0.6982 0.8989 -0.6982 CORRIENTE MEDIA 1.0539 0.1484 0.9326 -0.2721 Tabla 4.2 Comparación de corrientes estimadas, estímulo ISS. En la Figura 4.21 se muestran 3 casos en que el controlador no generó acción de control. En el caso A no controló porque ambas neuronas estaban en respuesta pasiva. En B no controló porque la referencia estaba en descarga pasiva. Y en C no controló porque ambas neuronas presentaban igual modo de descarga. 88 A1. Estímulo y corriente de control. Amplitud CSU Referencia 0.05 [µA], DC 0 [µA], media 0.4195 [µA], amplitud CSU Planta 0.1 [µA], DC -0.2 [µA], media 0.6479 [µA]. A2. Voltaje de membrana. B1. Estímulo y corriente de control. Amplitud CSU Referencia 0.05 [µA], DC -0.5 [µA], media -0.0817 [µA], amplitud CSU Planta 0.1 [µA], DC -0.6 [µA], media 0.2342 [µA]. B2. Voltaje de membrana. C1. Estímulo y corriente de control. Amplitud CSU Referencia 0.05 [µA], DC 0 [µA], media 0.4392 [µA], amplitud CSU Planta 0.1 [µA], DC -0.6 [µA], media 0.2441 [µA]. C2. Voltaje de membrana. Figura 4.21 Ejemplos donde no controló, estímulo ISS. 89 Si bien se probó que el sistema de control funciona cuando la entrada es sináptica sintética, no siempre es aplicable pues cerca de un 40% de las veces el Bloque Clasificador declara que no puede clasificar la señal de voltaje que recibió (ver ejemplo Figura 4.22). Si bien esta tasa disminuye a medida que la amplitud DC del estímulo se hace más depolarizante (se aumenta) o más hiperpolarizante (se disminuye), en este trabajo se presentan casos límites, es decir aquellos estímulos cercanos a la corriente de reobase para ambos modos. A. Estímulo y corriente de control. Amplitud CSU Referencia 0.1 [µA], DC -0.7 [µA], media 0.1718 [µA], amplitud CSU Planta 0.1 [µA], DC 0.2 [µA], media 1.0438 [µA]. B. Voltaje de membrana. Figura 4.22 Ejemplo donde no pudo controlar, estímulo ISS. 4.3. Robustez del controlador En esta sección se presentan los resultados obtenidos al aplicar el sistema de control diseñado a neuronas a las cuales se le cambiaron los parámetros de conductancia, para así tener una idea de la robustez del controlador frente a cambios en los parámetros de la Planta. Se variarán para este estudio sólo las conductancias gL y gT, ya que la densidad de canales iónicos puede variar a lo largo de la vida, producto de enfermedades que afectan a los canales iónicos [ 31], [ 26] o por traumatismos [ 62]. La capacitancia del modelo está en directa relación con el área de la neurona, luego es difícil estimar qué tipo de variaciones se podría obtener. Los voltajes de reversa VL y VT dependen de la concentración de iones del medio y quedan definidos por la ecuación de potencial de Nerst, luego también es difícil estimar variaciones para estos valores. 90 Las conductancias se variaron en ± 20% como muestra la Tabla 4.3. Se definió una variación de pequeña magnitud pues como se dijo en la Sección 3.2.1 es posible obtener otros modos de respuesta bajo otra sintonización de parámetros, para evitar la aparición de estos otros modos de respuesta se decidió realizar sólo estas pequeñas variaciones. PARÁMETRO VALOR IFB gL [mS] 0.035 0.042 0.028 gT [mS] 0.8 0.96 0.64 + 20% -20% Tabla 4.3 Variación de conductancias. En la Figura 4.23 se muestran los resultados obtenidos al variar la conductancia gL. Como se puede ver al aumentar esta conductancia a 0.042 [mS], gráficos A1, A2 y A3, el sistema de control hace una mala estimación de la corriente de estímulo de la Planta, por lo que la acción de control no logra llevar el sistema de descargas tónicas a modo burst, excepto hasta el instante 2500 [mseg] donde la corriente de estimula de la Planta se va a 0.67 [µA]. Mientras que en el caso B donde se disminuye la conductancia gL, el sistema de control logra modificar el modo de descarga de la Planta. La diferencia entre ambos resultados se debe probablemente al aumento de la frecuencia de descarga de la Planta cuando gL es 0.028 [mS]. En el caso del control en que se quiere llevar la Planta de descargas burst a tónico, ver Figura 4.24, consigue su objetivo de control. Esto pues la respuesta en modo burst no se ve afectada mayormente por la variación de la conductancia, la única variación evidente es la amplitud máxima de la variable h. Cuando se aumenta o disminuye la conductancia gT, tanto para el caso en que se controla el modo tónico (ver Figura 4.25) y el modo burst (Figura 4.26), se consigue modificar el modo de respuesta de la Planta al aplicar la acción de control. Se concluye que el controlador es mucho más sensible ante disminuciones de la conductancia gL que frente a aumentos de la misma. Mientras que es robusto ante variaciones en gT dentro del rango explorado. 91 Gráficos para gL = 0.042 [mS]. Gráficos para gL = 0.028 [mS]. A1. Estímulo y corriente de control. IEST_REF = 0.0949 [µA], IEST_PLA = 0.9522 [µA]; IREF = 0.1 [µA], IPLA = 1.1 [µA]. B1. Estímulo y corriente de control. IEST_REF = 0.0949 [µA], IEST_PLA = 1.2554 [µA]; IREF = 0.1 [µA], IPLA = 1.1 [µA]. A2. Voltaje de membrana. B2. Voltaje de membrana. A3. Variable h. B3. Variable h. Figura 4.23 Variación de gL, de tónico a burst. 92 Gráficos para gL = 0.042 [mS]. Gráficos para gL = 0.028 [mS]. A1. Estímulo y corriente de control. IEST_REF = 1.1049 [µA], IEST_PLA = 0.0498 [µA]; IREF = 1.1 [µA], IPLA = 0.1 [µA]. B1. Estímulo y corriente de control. IEST_REF = 1.1049 [µA], IEST_PLA = 0.1573 [µA]; IREF = 1.1 [µA], IPLA = 0.1 [µA]. A2. Voltaje de membrana. B2. Voltaje de membrana. A3. Variable h. B3. Variable h. Figura 4.24 Variación de gL, de burst a tónico. 93 Gráficos para gT = 0.96 [mS]. Gráficos para gT = 0.64 [mS]. A1. Estímulo y corriente de control. IEST_REF = 0.0949 [µA], IEST_PLA = 1.1049 [µA]; IREF = 0.1 [µA], IPLA = 1.1 [µA]. B1. Estímulo y corriente de control. IEST_REF = 0.0949 [µA], IEST_PLA = 0.0949 [µA]; IREF = 0.1 [µA], IPLA = 1.1 [µA]. A2. Voltaje de membrana. B2. Voltaje de membrana. A3. Variable h. B3. Variable h. Figura 4.25 Variación de gT, de tónica a burst. 94 Gráficos para gT = 0.96 [mS]. Gráficos para gT = 0.64 [mS]. A1. Estímulo y corriente de control. IEST_REF = 1.1049 [µA], IEST_PLA = 0.0949 [µA]; IREF = 1.1 [µA], IPLA = 0.1 [µA]. B1. Estímulo y corriente de control. IEST_REF = 1.1049 [µA], IEST_PLA = 0.0949 [µA]; IREF = 1.1 [µA], IPLA = 0.1 [µA]. A2. Voltaje de membrana. B2. Voltaje de membrana. A3. Variable h. B3. Variable h. Figura 4.26 Variación de gT, de burst a tónico. 95 4.4. Control realimentado En esta sección se presentan los resultados obtenidos al modificar el sistema de control anterior realimentándolo con la salida. Esta mejora se plantea con el fin de solucionar las discrepancias en frecuencia (cuando descarga en tónico) e hiperpolarización (cuando descarga en burst) que presenta la neurona Planta en comparación con la neurona de Referencia una vez que el estímulo que recibe la neurona Planta a sesado, ver Sección 4.2.1. Para implementar este sistema de control realimentado se definieron 4 estados en los que puede estar el sistema. • Estado 0 o estado inicial. Es éste el estado en que parte la simulación del sistema de control y en el que está después de pasar por el Estado 3. En el Estado 0 espera que pasen 500 [mseg] antes de intentar controlar, si generó acción de control pasa al Estado 1. La estimación de corrientes de estímulo la realiza a partir del vector de voltaje correspondiente a los 500 [mseg] anteriores al tiempo en que se está calculando el control. • Estado 1 o de actuación. Este estado permite que se aplique la acción de control diseñada en el Estado 0 por 500 [mseg]. • Estado 2 o de verificación. En este estado se verifica que la acción de control actual haya llevado la repuesta de la Planta al estado de la Referencia, de ser así verifica también la precisión de la corriente aplicada y la corrige de ser necesario. • Estado 3 o de liberación. Cuando está en este estado el sistema es liberado de cualquier acción de control por 500 [mseg]. Entre estos 4 estados existen sólo algunas transiciones posibles, las que se muestran en la Figura 4.27. En el Anexo C se detalla el código implementado en Matlab 6.5. 96 Figura 4.27 Diagrama de estado del sistema de control. En la Figura 4.28 se muestra el resultado al aplicar el control con realimentación cuando la Planta está en modo burst y la Referencia está en modo tónico. Mientras que en la Figura 4.29 se muestran algunos casos donde no se debe aplicar acción de control. 97 A1. Estímulos y corrientes de control. Planta 0.1 [µA] entre 50 y 2000 [mseg]. Referencia 0.9 desde los 50 [mseg]. B1. Estímulos y corrientes de control. Planta 0.1 [µA] desde 50 [mseg]. Referencia 0.9 desde los 50 [mseg]. A2. Voltaje de membrana. B2. Voltaje de membrana. A3. Variable h. B3. Variable h. Figura 4.28 Control de burst a tónico, estímulo DC. 98 A1. Estímulos y corrientes de control. Planta 0.6 [µA] desde 50[mseg] a 3900[mseg]. Referencia 0.4 entre 50 y 3500 [mseg]. A2. Voltaje de membrana. B1. Estímulos y corrientes de control. Planta 0.9 [µA] desde 50 [mseg]. Referencia 0.2 entre 50 y 2000 [mseg]. B2. Voltaje de membrana. C1. Estímulos y corrientes de control. Planta 0.1 [µA] desde 50 [mseg] a 3900 [mseg]. Referencia -0.1 entre 50 y 3500 [mseg]. C2. Voltaje de membrana. Figura 4.29 Casos en que no aplicó acción de control, estímulo DC. 99 Como se puede ver en la Figura 4.28 el sistema de control con realimentación logró su objetivo de control, llevar la Planta de descargas en modo burst al modo tónico. Si se compara este resultado con el mostrado en la Figura 4.12, en la cual al cambiar el estímulo de la Planta en el milisegundo 2500 el control si bien mantenía el modo tónico no mantenía la frecuencia de descarga de la Referencia (Figura 4.12 – A2) sino que aumentaba. Mientras que en la Figura 4.28 – A2 se puede ver cómo el control con realimentación consigue su objetivo en el milisegundo 1600 aproximadamente, luego en el milisegundo 2500 cambia la señal de estímulo de la Planta, con lo que se produce un reajuste de la corriente de control que dura 500 [mseg], es decir en el milisegundo 3000 ya a superado la variación en el estímulo de la Planta y esta descarga a la misma frecuencia que la Referencia. En la Figura 4.29 se verifica que el sistema de control realimentado mantienen las características del sistema de control de lazo abierto. Es decir, no aplica el control si alguna de las dos neuronas está en respuesta pasiva, y tampoco lo aplica si están en el mismo modo de descarga. 100 Capítulo 5 Conclusiones y Trabajo Futuro A continuación se procede a extraer las principales conclusiones correspondientes al trabajo realizado en esta memoria. Además, hacia el fin del capítulo, se mencionan las líneas de investigación que deberían seguir los trabajos futuros. Con el fin de aproximar el umbral de corriente a partir del cual se produce la descarga en estallido o la descarga tónica, y la duración que debe tener este estímulo, se determinó en la Sección 3.2.1.1 para el modelo IFB con los parámetros de la Tabla 3.1, que la corriente reobase para la descarga tónica es de 0.8 [µA] aplicada por 800 [mseg ] y la corriente de cronaxia es de 0.94 [µA] aplicada por 145 [mseg]. Para la descarga en modo burst, la corriente de reobase se determinó como 0.17 [µA] aplicada por 800 [mseg] y la corriente de cronaxia de una amplitud de 0.35 [µA] aplicada por 86 [mseg]. Es decir, para corrientes de larga duración el umbral de descarga tónica está cercano a los 0.8 [µA]. Mientras que el umbral de estallidos está cercano a los 0.17 [µA]. Respecto al tipo de respuestas ante corrientes variables en el tiempo, se estableció que para corrientes sinusoidales y tipo tren de pulsos de alta frecuencia, el modelo se satura y la respuesta del modelo no aumenta de manera lineal con la frecuencia del estímulo, lo que si ocurre para señales de baja frecuencia. En particular, en la Sección 3.2.1.2 se estableció que para una señal sinusoidal de amplitud AC 0.67 [µA], amplitud DC 1 [µA] y frecuencia igual o mayor a 24 [Hz], la respuesta es de distinta frecuencia que la del estímulo (ver Figura 3.17). Para frecuencias mayores a 130 [Hz] (ver Figura 3.18) la respuesta del modelo es la misma que si se aplica una señal 101 continua (comparar respuestas del gráfico A con B y C en la Figura 3.18). Respecto a entradas del tipo tren de pulsos con parámetros dentro del rango establecido para estimulación cerebral profunda, se estableció que la respuesta del modelo IFB supone que está siendo estimulada por una corriente constante. En particular se estableció (ver Figura 3.26) que para un estímulo de amplitud DC 0.67 [µA], amplitud 6.97 [µA], ancho de pulso 100 [µseg] y frecuencia 182 [Hz], la salida es muy parecida al caso de la corriente de reobase de la Figura 3.10 - B. Se concluye entonces que estimular con una forma de onda tipo tren de pulso o sinusoidal de alta frecuencia, es equivalente en este modelo a estimular con una corriente constante. Se cumplió así uno de los objetivos de este trabajo, el de analizar desde el punto de vista de entrada/salida la respuesta de un modelo dinámico de neurona talámica, lo que facilitó el diseño del sistema del control. En base a estas pruebas de entrada/salida, se diseñaron dos estimadores de corriente. Estos estiman la amplitud de la señal de corriente que estimula a una neurona en base al voltaje de membrana que produce este estímulo. Para la respuesta en modo tónico se determinó, ecuación (4.2), que es posible estimar la corriente de estímulo en base a la frecuencia media de descarga de la señal. (Ver Figura 4.4). En el caso de la respuesta en modo burst, el estimador usa la derivada algebraica de la señal de voltaje (ecuación (4.3)) pues tiene una relación lineal con la amplitud de la señal de estímulo. (Ver Figura 4.7). De las pruebas hechas al controlador en la Sección 4.2.3 ante entradas sinápticas sintéticas, se verificó que el estimador de modo tónico es poco sensible a variaciones aleatorias de la señal de estímulo. Por otro lado, el estimador de modo burst es altamente sensible a variaciones de la señal. Como se detalla en el siguiente párrafo, estos estimadores de corriente son la base del sistema de control utilizado. Para determinar en que modo estaba descargando una neurona, se consideraron los umbrales de descarga tónica y burst. Así, si una señal de voltaje alcanzaba niveles de hiperpolarización bajo el umbral del estallido, entonces se definía dicha neurona como en modo de descarga burst. Por otro lado, si la señal de voltaje presentaba una depolarización por sobre el umbral de la descarga de espigas tónicas, se definía que dicha neurona estaba en descargas tónicas. En base a todo el conocimiento descrito anteriormente, se diseñó una estrategia de control en lazo abierto que, a partir del vector de voltaje, de largo 200 [mseg], clasifica el modo de respuesta 102 para cada neurona, estima la corriente de estímulo y calcula la corriente de control que debe aplicar (ver esquema en la Figura 4.1). En todas las pruebas hechas al controlador ante entradas tipo escalón y sinusoidales (ver Secciones 4.2.1 y 4.2.2) se cumplió el objetivo de control. Para entradas sinápticas en cambio, no siempre fue posible aplicar el controlador, pues el bloque clasificador no fue capaz de clasificar el modo de respuesta. Sin embargo, en los casos en que si pudo clasificar el modo de respuesta y se aplicó el controlador, se consiguió el objetivo de control. En la Sección 4.3 se estudió la robustez de este controlador frente a variaciones de los parámetros de la Planta. Se determinó que frente a variaciones de la conductancia gT y aumentos en la conductancia gL el estimador es robusto, sin embargo es muy sensible a disminuciones de gL. Una variante del control propuesto en la Sección 4.1 y descrito en el párrafo anterior, es el control realimentado descrito en la Sección 4.4. En este sistema de control realimentado, cada 500 [mseg] se actualiza la corriente de control. De no ser posible, se libera el sistema de neuronas de la corriente de control por un período de tiempo, para luego intentar aplicar nuevamente el control. Se verificó (ver Figura 4.28) que este sistema de control permite que la Planta descargue en el mismo modo que la Referencia, incluso cuando el estímulo de la Planta varía a lo largo del tiempo. Con los resultados anteriores, se cumplió el otro objetivo de este trabajo; diseñar y aplicar un sistema de control que permita que una neurona Planta descargue en el mismo modo que una neurona de Referencia. Lo anterior se logró a partir de la información de voltaje de membrana de ambas neuronas. En relación a las líneas de investigación futuras que se advierten ahora ya finalizado este trabajo, queda pendiente determinar cuál sería el desempeño y cuáles serían los ajustes necesarios hacer al controlador, para aplicarlo a una población de neuronas. En una primera etapa se debería ajustar el controlador para que trabaje con el voltaje de membrana extracelular, aquí el principal cambio en el controlador tiene que ver con el bloque clasificador pues debería determinar el tipo de respuesta en base a métodos estadísticos, como se indicó en la Sección 4.1. Como segunda etapa, para simular la población de neuronas se propone seguir la idea propuesta por Huertas y Smith [ 63], donde cada neurona se define como un modelo IFB y se defina una función de densidad de probabilidad sobre las variables de estado de la población de neuronas. Este modelo de poblaciones genera un amplio rango de respuestas dependiendo de los parámetros usados, 103 entre ellas estallidos rítmicos. Es posible también simular poblaciones no mediante un modelo de densidad de probabilidad, sino que agregando a la ecuación (3.1) de balance de corriente, corrientes sinápticas que representen los estímulos provenientes de otras neuronas. En ese caso es recomendable consultar la publicación de Huertas et al [ 64] donde simula una red neuronal a partir del modelo IFB. Si bien en las publicaciones mencionadas en la Sección 2.3.3 también se indica como construir poblaciones de neuronas, cada una de un compartimiento, los valores de los parámetros para las corrientes sinápticas (conductancias) no serán los mismos si se usa el modelo IFB. Otra línea de continuación que deja abierta este trabajo es la de determinar, en base a un modelo multicompartimiento de una neurona, las consecuencias del sistema de control aplicado a distintas zonas de la neurona, para el caso en que la estimulación se realiza mediante un microelectrodo intracelular y el caso en que se usa uno extracelular. Determinando en qué parte de la neurona tiene mayores ventajas posicionar el electrodo de muestreo y estimulación en cada caso. La importancia de las dos líneas de investigación futura ya planteadas y el sistema de control presentado en este trabajo, radica en la posibilidad de realizar estimulación y muestreo de voltaje mediante arreglos de microelectrodos extracelulares basados en nanocables [ 65]. Esto con el propósito de realizar una estimulación más localizada que la que se puede realizar con los electrodos actuales. Lo cual posibilitaría en un futuro próximo nuevas terapias in vivo aplicables a patologías que afectan al sistema nervioso central y periférico. 104 Referencias [ 1] Testerman, R., Rise, M. and Stypulkowski, P., “Electrical stimulation as therapy for neuroligical disorders”. IEEE Engineering in Medicine and Biology 25(5): 74-78. 2006. [ 2] Wichmann, T. and DeLong, M., “Deep Brain Stimulation for neurologic and neuropsychiatric disorders”. Neuron 52: 197-204, October 2006. [ 3] Medtronic [en línea] <http://www.medtronic.es/ES/> [Consulta 17 de Agosto de 2007]. [ 4] Anderson, W.S. and Lenz, F.A., “Surgery Insigth: Deep brain stimulation for movements disorders”. Nature Clinical Practice Neurology 2(6): 310-320, June 2006. [ 5] The NIH Record Archives, Volumen LIII(5), Marzo 6, 2001 [en línea] <http://nihrecord.od.nih.gov/> [Consulta 17 de Agosto de 2007]. [ 6] Abbott, A., “Deep in thought”. Nature 436: 18 – 19, July 2005. [ 7] Hardesty, D. and Sackeim, H., “Deep brain stimulation in movement and psychiatric disorders”. Biol Psychiatry 61: 831 – 835, 2007. 105 [ 8] McIntyre, C., Grill, W., Sherman, D., and Thakor, N., “Model-based analysis of deep brain stimulation of the thalamus”. Proceedings of the Second Joint EMBS/BMES Conference (Houston, TX, USA): 2047 - 2048, October 2002. [ 9] McIntyre, C., Grill, W., Sherman, D., and Thakor, N., “Cellular Effects of deep brain stimulation: Model –based analysis of activation an inhibition”. Journal of Neurophysiology 91: 1457-1469, 2004. [ 10] Shepherd, Gordon, edited by. The Synaptic Organization of the Brain. 5th ed. New York, Orford University Press, 2003. 660p. [ 11] Purves, D., Agustine, G., Fitzpatrick, D., Katz, L., LaMantia, A. y McNamara, J., Invitación a la neurociencia. 1ª ed. Argentina. Médica Panamericana, 2004. 680p. Disponible en el Bookshelf de PubMed [en línea], [Consulta Noviembre de 2006] < http://www.ncbi.nlm.nih.gov/sites/entrez?db=Books&itool=toolbar>. [ 12] Smith, G, “Inhibition and bursting in thalamocortical relay neurons and network models of the visual thalamus” [en línea]. http://as.wm.edu/Faculty/Smith/downloads/downloads/seminars/SmithInhibiti onAndBurstingTCNetworks.pdf [Consulta 14 de Junio de 2007, 12:55 hrs]. [ 13] Destexhe, A., Neubig, M. Ulrich, D. and Huguenard, J., “Dendritic low-threshold calcium currents in thalamic relay cells”. The Journal of Neuroscience 18(10): 3574 – 3588, May 1998. [ 14] Dorf, R. edited by, The Electrical Engineering Handbook. Second Edition, CRC Press, 2000. 2719 p. [ 15] Hodgkin, A. And Huxley, A., “A quantitative description of membrane currents and its application to conduction an excitation in nerve”. Journal of Physiology 117: 500 – 544, 1952. 106 [ 16] Hodgkin, A.L. and Huxley, A.F., “Currents carried by sodium and potassium ions through the membrane of the giant axon of Loligo”. Journal of Phsiology 116: 449-472, 1952. [ 17] Hodgkin, A.L. and Huxley, A.F., “The components of membrane conductance in the giant axon of Loligo”. Journal of Phsiology 116: 473-496, 1952. [ 18] Hodgkin, A.L. and Huxley, A.F., “The dual effect of membrane potential on sodium conductance in the giant axon of Loligo”. Journal of Phsiology 116: 497506, 1952. [ 19] Nobelprize.org. The Nobel Prize in Physiology or Medicine 1963. [en línea] <http://nobelprize.org/nobel_prizes/medicine/laureates/1963/> [Consulta 26 de Marzo de 2007] [ 20] Hille, Bertil, Ionic channels of excitable membranes. 2ª ed. Sunderland, Mass, Sinauer, 1992. 607 p. [ 21] Hodgkin, Alan, The conduction of the nervous impulse. 3ª ed. England. Liverpool University Press, 1967. 108 p. [ 22] McCormick, D. and Bal, T., “Sleep and arousal: Thalamocortical mechanisms”. Annual Review Neuroscience 20: 185-215, 1997. [ 23] Llinás, R. and Steriade, M., “Bursting of thalamic neurons and states of vigilance”. Journal of Neurophysiology 95: 3297 – 3308, 2006. [ 24] Vega-Saenz de Miera, E.C., Rudy, B., Sugimori, M., and Llinas, R., ”Molecular characterization of the sodium channel subunits expressed in mammalian cerebellar Purjinke cells”. Proceedings of the National Academy of Science 94: 7059 – 7064, 1997. 107 [ 25] Jahnsen, H., Llinás, R., “Ionic basis for electroresponsiveness and oscillatory properties of guinea pig thalamic neurons in vitro”. Journal of Physiology 349: 227 – 247, 1984. [ 26] Kullman, D., “The neuronal channelophaties”. Brain 125:1177-1195, 2002. [ 27] McCormick, D.A. and Pape, H.C., “Properties of hyperpolarization-activated cation current and its role in rhythmic oscillation in thalamic relay neurons”. Journal of Physiology 431: 291 – 318, 1990. [ 28] Entrez Gene [en línea] http://www.ncbi.nlm.nih.gov/sites/entrez [Consulta 10 de Octubre de 2007]. [ 29] McNamara, J.O., “Cellular and molecular basis of epilepsy”. The Journal of Neuroscience 14(6): 3413 – 3425, June 1994. [ 30] Weinreich, F. and Jentsch, T.J., “Neurological diseases caused by ion-channel mutations”. Current Opinion in Neurobiology 10: 409 – 415, 2000. [ 31] Kullman, D. and Hanna, M., “Neurological disorders caused by inherited ionchannel mutations”. The Lancet Neurology 1: 157-166, July 2002. [ 32] Llinas, R., Ribary, U., Jeanmonod, D., Kronberg, E., and Mitra, P., “Thalamocortical dysrhythmia: A neurological and neuropsychiatric syndrome characterized by magnetoencephalography”. Proceedings of National Academy of Science 96(26): 15222 – 15227, December 1999. [ 33] Keener, James and Sneyd, James., Mathematical Physiology. New York, SpringerVerlag, 1991. 766 p. (Interdisciplinary applied mathematics; v.8). [ 34] McCormick, D. and Huguenard, R., “Simulation of the currents involved in phythmic oscillations in thalamic relay neurons”. Journal of Neurophysiology 68(4): 1373-1383, October 1992. 108 [ 35] McCormick, D. and Huguenard, R., “A model of the electrophysiological properties of thalamocortical relay neurons”. Journal of Neurophysiology 68(4): 1384-1400, October 1992. [ 36] Golomb, D., Wang, X. and Rinzel, J., “Propagation of spindle waves in a thalamic slice model”. Journal of Neurophysiology 75(2): 750 - 769, February 1996. [ 37] Destexhe, A., Babloyantz, A. and Senjnowski, J., “Ionic mechanisms for intrinsic slow oscillations in thalamic relay neurons”. Biophysical Journal 65: 1538 – 1552, October 1993. [ 38] Rhodes, P. and Llinas, R., “A model of thalamocortical relay cells”. Journal of Physiology 565(3): 765 – 781, 2005. [ 39] Rinzel J., Millar R., “Numerical calculation of stable and instable periodic solutions to the Hodgkin-Huxley equations”. Mathematical Biosciences 49: 27 – 59, 1980. [ 40] Aihara K, Matsumoto G., ”Two stable steady status in the Hodgkin-Huxley axons”. Biophysical Journal 41: 87 – 89, 1983. [ 41] Bedrov, Y.A., Akoev, G.N. and Dick,O.E., “Partition of the Hodgkin-Huxley type model parameter space into the regions of qualitatively different solutions”. Biological Cybernetics 66: 413 – 418, 1992. [ 42] Guckenheimer, J. and Laboriau, I.S., “Bifurcation of the Hodgkin and Huxley equations: A new Twist”. Bulletin of Mathematical Biology 55(5): 937 – 952, 1993. [ 43] Adrian R., Marshall M., “Sodium currents in mammalian muscle”. Journal of Physiology 268: 223-250, 1977. [ 44] Falk, G. and Fatt, P., “Linear electrical properties of striated muscle fibres observed with intracellular electrodes”. Journal of Physiology 268: 223-250, 1977. 109 [ 45] Terada, K., Yoshizawa, S., Nishimura, C., “Dynamical behavior oh the Hodgkin Huxley equations for muscles”. IEEE Systems, Man, and Cybernetics 1999 Conference Proceedings 2:371 – 376, 12-15 October 1999. [ 46] Pappone, P., “Voltag-clamp experiments in normal and denervated mammalian skeletal muscle fibres”. Journal of Physiology 306: 377 – 410, 1980. [ 47] Wang J., Zhang H., Tsang, K.M., “Hopf bifurcation in the Hodgkin-Huxley model exposed to ELF electrical field”. Proceedings of the 25th Annual Conference of the IEEE EMBS Cancun Mexico: 17 – 21, 2003. [ 48] Wang J., Kai Ming T., Zhang H., “Hopf bifurcation in the Hodgkin-Huxley model exposed to ELF electrical field”. Chaos, Solitions & Fractals 20: 759 – 764, 2004. [ 49] Wang J., Che, Y., Fei, X.,Li, L., “Multiparameter Hopf bifurcation in HodgkinHuxley model exposed to ELF external electric field”. Chaos, Solitions & Fractals 26: 1221 – 1229, 2005. [ 50] Wang J., Chen L., Fei X., “Analysis and control of the bifurcation of HodgkinHuxley model”. Chaos, Solitions & Fractals 31: 247 – 256, 2007. [ 51] Wang J., Chen L., Fei X., “Bifurcation control of the Hodgkin-Huxley equations”. Chaos, Solitions & Fractals 33: 217 – 224, July 2007. [ 52] Fei X, Wang J., Chen L., “Bifurcation control of the Hodgkin-Huxley model of nerve system”. Proceedings of the 6th World Congress on Intelligent Control and Automation Dalian, China: 9406 – 9410, 21 – 23, June 2006. [ 53] Huguenard, R. and Prince, A., “Slow inactivation of a TEA-sensitive K current in acutely isolated rat thalamic relay neurons”. Journal of Neurophysiology 66(4): 1316 – 1328, October1991. 110 [ 54] Abbott, A., “Deep in thought”. Nature 436:18-19, July 2005. [ 55] Smith, G., Cox, C., Sherman, Murray and Rinzel, J., “Fourier analysis of sinusoidally driven thalamocortical relay neurons and a minimal integrate-and-fireor-burst model”.Journal of Neurophysiology 83: 588 – 610, 2000. [ 56] Shampine, L.F. and Reichelt, M.W. “ The Matlab Ode Suite”. The MathWorks [en línea] http://www.mathworks.com/ [Consulta el 10 de Diciembre de 2006]. [ 57] Bloomfield, S.A., Hamos, J.E. and Sherman, S.M., “Passive cable properties and morphological correlates of neurons in the lateral geniculate nucleus of the cat”. Journal of Physiology 383: 653 – 692, 1987. [ 58] Bloomfield, S.A. and Sherman, S.M., “Dendritic current flow in relay cells and interneurons of the cat’s lateral geniculate nucleus”. Proceedings of National Academy of Science 86: 3911 – 3914, May 1989. [ 59] Izhikevich, E., “Wich model to use for cortical spiking neuron?”. IEEE Transactions on Neural Networks 15(5), September 2004. [ 60] Stevens, C.F. and Zador, A.M., “Input synchrony and the irregular firing of cortical neurons”. Nature Neuroscience 1(3):210-217. July 1998. [ 61] Sherman, S.M., “Tonic and burst firing: dual modes of thalamocortical relay”. Trends in Neuroscience 24(2): 122 – 126, February 2001. [ 62] Hains, B.C., Saab, C.Y. and Waxman, S.G., “Alterations in burst firing of thalamic VPL neurons and reversal by Nav1.3 antisense after spinal cord injury”. Journal of Neurophysiology 95: 3343 – 3352, 2006. [ 63] Huertas, M and Smith, G., “A multivariate population density model of the dLGN/PGN relay”. Journal of Computational Neuroscience 21(2): 171 – 189, 2006. 111 [ 64] Huertas, M. Groff, J. and Smith, G., “Feedback inhibition nd throughput properties of integrate-and-fire-or-burst network model of retinogeniculate transmission”. Journal of Computational Neuroscience 19: 147 – 180, 2005. [ 65] Yu, Z., McKnight, T.E., Nance Ericson, M., Melechko, A.V., Simpson, M.L. and Morrison III, B., “Vertically Aligned Carbon Nanofiber Arrays Record Electrophysiological Signals from Hippocampal Slices”. Nano Letters 7(8): 2188 – 2195, August 2007. 112 Anexo A Modelo IFB A continuación se detalla el código, para Matlab 6.5, donde se implementa el modelo IFB. Para implementar el sistema (3.6) se definió la función IFBdif; que recibe de parámetros un vector de tiempo (t), un vector de condiciones iniciales (y) y 2 parámetros, el voltaje anterior (v) y la corriente de estímulo (Iapp), como muestra la siguiente línea. function dydt = IFBdif(t,y,v,Iapp) v_h = -70; if v >= v_h dydt = [(0.5*Iapp-0.5*0.035*(y(1)+75)-0.5*0.8*y(2)*(y(1)-120)); (-y(2)/20)]; else dydt = [(0.5*Iapp-0.5*0.035*(y(1)+75)); ((1-y(2))/100)]; end La función IFBdif se resuelve mediante el método ode45 de Matlab6.5, que aplica el método Runge-Kutta para resolver ecuaciones diferenciales. El resultado almacenado se guarda como una variable de tiempo (t) y otra de voltaje llamada (y) como muestra la siguiente línea. Donde para un instante de tiempo i, se resuelve el sistema de ecuaciones a partir de la información de las condiciones iniciales y el voltaje y la corriente anterior. Esto se hace para cada paso de tiempo, es decir para cada instante entre i-1 e i. [t,y] = 1),estimulo(i)); ode45(@IFBdif,[T_total(i-1) T_total(i)],[V_integral h_integral],[],V(i- Cada vez que se genera un spike se debe llevar el voltaje de membrana al voltaje de reset. Para ello se implementó un if en que los argumentos que le entrega al ode45 varian dependiendo 113 del voltaje actual. A continuación se muestran las líneas de código que implementan esta condición. % Caso en que el voltaje esta bajo el umbral de espiga if V(i-1) < V_teta -e [t,y] = ode45(@IFBdif,[T_total(i-1) T_total(i)],[V_integral 1),estimulo(i)); % Actualiza las variables V_acumulado = y(length(y),1); h_acumulado = y(length(y),2); % CI de la siguiente integral V_integral = V_acumulado; h_integral = h_acumulado; % Caso en que supero el umbral y va a generar una espiga elseif V(i-1) > (V_teta - e) & V(i-1) ~= V_spike [t,y] = ode45(@IFBdif,[T_total(i-1) T_total(i)],[V_integral 1),estimulo(i)); % Actualiza las variables V_acumulado = V_spike; h_acumulado = y(length(y),2); % CI de la integral V_integral = V_acumulado; h_integral = h_acumulado; % Caso en que vuelve de haber generado una espiga elseif V(i-1) == V_spike [t,y] = ode45(@IFBdif,[T_total(i-1) T_total(i)],[V_integral 1),estimulo(i)); % Actualiza las variables V_acumulado = V_reset; h_acumulado = y(length(y),2); % CI de la integral V_integral = V_acumulado; h_integral = h_acumulado; h_integral],[],V(i- h_integral],[],V(i- h_integral],[],V(i- end Este if se ejecuta para cada paso de tiempo de la simulación, el voltaje y la variable h en función del tiempo, se almacenan en los vectores V y H como, V = [V; V_acumulado]; H = [H; h_acumulado]; 114 Anexo B Sistema de control lazo abierto En este caso, y de modo análogo a lo indicado en el Anexo A, se implementa un for que en cada paso de la simulación ejecuta un if para la neurona Planta, otro para la neurona de Referencia y otro para el módulo controlador. Los if de la neurona Planta y Referencia son iguales a los del Anexo A, pero los parámetros que reciben son distintos y además se guardan en distintos vectores V y H, La neurona Planta recibe como estímulo el valor aux2, que corresponde al estímulo aplicado (estimulo_pla) menos la corriente de control aplicada (con_pulso), % Neurona Planta aux2 = estimulo_pla(i) + con_pulso; [t,y] = ode45(@IFBdif,[T_total(i-1) T_total(i)],[VI_pla hI_pla],[],V_pla(i-1),aux2); V_pla = [V_pla; Vacu_pla]; H_pla = [H_pla; hacu_pla]; Mientras, para la neurona de Referencia el estímulo aplicado corresponde únicamente al estímulo que recibe en ese instante. % Neurona Referencia [t,y] = ode45(@IFBdif,[T_total(i-1) 1),estimulo_ref(i)); V_ref = [V_ref; Vacu_ref]; H_ref = [H_ref; hacu_ref]; T_total(i)],[VI_ref hI_ref],[],V_ref(i- Cuando el tiempo de simulación supera los 200 [mseg], se debe clasifica el modo de respuesta, lo cual se hace a través de las siguientes líneas, clas_pla = clase_modo(V_pla) 115 clas_ref = clase_modo(V_ref) Estas líneas llaman a la función clase_modo que determina en qué modo está la respuesta. La función clase_modo se muestra en la siguientes línes de código: function clase_mod = clase_modo(vector_voltaje); % La respuesta que da es: % 0 = si no pudo clasificar el modo % 1 = si esta en modo burst % 2 = si esta en modo tonico % 3 = si esta en modo pasivo % Si esta en burst donde_burst = find(vector_voltaje <= -70); % Si esta en tonico donde_tonico = find(vector_voltaje >= -50); % Es burst y no tonico if isempty(donde_burst) ~= 1 & isempty(donde_tonico) == 1 clase_mod = 1; % Es tonico y no burst elseif isempty(donde_tonico) ~= 1 & isempty(donde_burst) == 1 clase_mod = 2; % No es tonico ni es burst --> es pasivo elseif isempty(donde_tonico) == 1 & isempty(donde_burst) == 1 clase_mod = 3; % si no supo que era else clase_mod = 0; 'No clasifico el modo de respuesta' end Luego de clasificar la respuesta, se estima la corriente de estímulo y se calcula la acción de control, todo esto se hace mediante el siguiente if: % Si ambos esta en el mismo modo no genera accion de control if clas_pla == clas_ref I_control = 0; % Si no pudo clasificar uno tampoco genera accion de control elseif clas_pla == 0 & clas_ref ~= 0 I_control = 0; elseif clas_pla ~= 0 & clas_ref == 0 I_control = 0; % Si alguno de los 2 esta en modo pasivo elseif clas_pla == 3 | clas_ref ==3 % Si clasifico y ninguno es pasivo genera accion de control else t_aux = T_total(1:i); Iest_ref = estimador_corriente(clas_ref,V_ref,t_aux); Iest_pla = estimador_corriente(clas_pla,V_pla,t_aux); I_control = Iest_ref - Iest_pla; end 116 Como se puede ver la corriente de control es la resta simple entre la corriente estimada para la Planta y la corriente estimada para la Referencia. Para estimar dichas corrientes si usa la función estimador_corriente, que recibe como argumento el modo de respuesta, el voltaje en el tiempo y el vector de tiempo. La función estimador_corriente se implementa como: function estimada = estimador_corriente(clase,vec_voltaje,tie_voltaje); % Si esta en modo burst if clase == 1 der_voltaje = derivada_de(vec_voltaje,tie_voltaje); aux1 = find(der_voltaje <= -0.25); aux1 = der_voltaje(aux1(1)) estimada = 2*aux1 + 0.6; % Si esta en modo tonico elseif clase == 2 [f1,f2,f3] = frecuencia(vec_voltaje,tie_voltaje); estimada = 40.2*f3*f3 + 16.85*f3 + 0.67; end Las funciones derivada_de y frecuencia que llama el estimador de corriente, calculan la derivada algebraica del vector de voltaje y la frecuencia de las espigas de la señal de voltaje, respectivamente. 117 Anexo C Sistema de control realimentado El control realimentado se implementa igual que el control en lazo abierto, con la diferencia que en vez de implementar el if que hace la estimación y calcula la acción de control condicionado a que se calcule una vez después del milisegundo 200. Esta se calcula o no dependiendo del estado en que se encuentre el sistema. Los estados se difinieron como el siguiente if, % ESTADO 0: En este estado parte y clasifica despues de liberar if Estado_actual(1) == 0 if i>500 V_pla_aux = V_pla(i-500:i); V_ref_aux = V_ref(i-500:i); clas_pla = clase_modo(V_pla_aux); clas_ref = clase_modo(V_ref_aux); % Si ambos esta en el mismo modo no genera accion de control if clas_pla == clas_ref I_control = 0; Estado_actual(1) = 0; % Si no pudo clasificar uno no genera accion de control elseif clas_pla == 0 & clas_ref ~= 0 I_control = 0; Estado_actual(1) = 0; elseif clas_pla ~= 0 & clas_ref == 0 I_control = 0; Estado_actual(1) = 0; % Si alguno de los 2 esta en pasivo no genera accion de control elseif clas_pla == 3 | clas_ref ==3 I_control = 0; Estado_actual(1) = 0; % Si clasifico y ninguno es pasivo genera accion de control else t_aux = T_total(i-500:i); Iest_ref_1 = estimador_corriente(clas_ref,V_ref_aux,t_aux); Iest_pla_1 = estimador_corriente(clas_pla,V_pla_aux,t_aux); if isempty(Iest_ref_1) == 1 | isempty(Iest_pla_1) == 1 I_control = 0; Estado_actual(1) = 3; 118 Estado_actual(2) = i; else I_control = Iest_ref_1 - Iest_pla_1; % Como genero accion de control lo deja por 500 mseg Estado_actual(1) = 1; Estado_actual(2) = i; end end % En cualquier otro caso vuelve a empezar else Estado_actual(1) = 0; end % ESTADO 1: Deja que actue el control por 500mseg sin chequearlo elseif Estado_actual(1) == 1 aux101 = Estado_actual(2) + 500; if aux101 > i % Sigue en estado E1 Estado_actual(1) = 1; else % Se va al estado E2 Estado_actual(1) = 2; Estado_actual(2) = i; end % ESTADO 2: Chequea el control actual elseif Estado_actual(1) == 2 V_pla_aux = V_pla(i-500:i); V_ref_aux = V_ref(i-500:i); clas_pla_2 = clase_modo(V_pla_aux); clas_ref_2 = clase_modo(V_ref_aux); % Si no pudo clasificar el modo deja el control que tiene para % recalcular mas tarde if clas_pla_2 == 0 | clas_ref_2 == 0 Estado_actual(1) = 1; Estado_actual(2) = i; % Si clasifico pero una respuesta es pasiva se va al estado 3 elseif clas_pla_2 == 3 | clas_ref_2 == 3 Estado_actual(1) == 3; Estado_actual(2) == i; I_control = 0; % Si clasifico el modo y ninguna es pasiva chequea el resultado else t_aux = T_total(i-500:i); Iest_ref_2 = estimador_corriente(clas_ref_2,V_ref_aux,t_aux); Iest_pla_2 = estimador_corriente(clas_pla_2,V_pla_aux,t_aux); % Le resta el control a la corriente estimada para la planta if isempty(Iest_pla_2) == 0 Iest_pla_2 = Iest_pla_2 - I_control; end % Si no pudo estimar la corriente se queda con la que tenia if isempty(Iest_ref_2) == 1 Iest_ref_2 = Iest_ref_1; elseif isempty(Iest_pla_2) == 1 Iest_pla_2 = Iest_pla_1; end % Si esta en un modo distinto al de la referencia inicial % libera el sistema de cualquier accion de control if clas_pla_2 ~= clas_ref | clas_ref_2 ~= clas_ref Estado_actual(1) = 3; Estado_actual(2) = i; I_control = 0; % Si esta en igual modo que la referencia inicial chequea % la precision del resultado 119 else aux102 = abs(Iest_ref_2 - Iest_ref_1); aux103 = abs(Iest_pla_2 - Iest_pla_1); I_control_2 = Iest_ref_2 - Iest_pla_2; % Si esta bien la estimacion de corrientes if aux102<5 & aux103<5 Iest_ref_1 = Iest_ref_2; Iest_pla_1 = Iest_pla_2; I_control = I_control_2; Estado_actual(1) = 1; Estado_actual(2) = i; % Si está mal la estimacion de corrientes hecha else Estado_actual(1) = 3; Estado_actual(2) = i; I_control = 0; end end end % ESTADO 3: Libera el modelo por 500mseg elseif Estado_actual(1) == 3 aux104 = Estado_actual(2) + 500; % se mantiene en estado E3 if aux104 > i Estado_actual(1) = 3; % se va al estado E0 else Estado_actual(1) = 0; Estado_actual(2) = i; end end 120