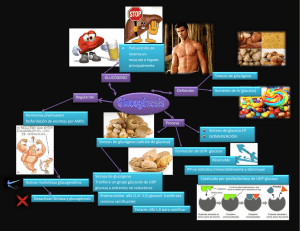
TEMA II METABOLISMO ENERGETICO I. METABOLISMO
Anuncio

Ir al índice Ir al menú principal TEMA II METABOLISMO ENERGETICO I. METABOLISMO ENERGETICO Y SU RELACION CON LA NUTRICION 2.1 INTRODUCCIÓN AL METABOLISMO ENERGETICO Para una mejor comprensión de este módulo dedicado a la Nutrición del deportis ta de alto rendimiento, y para un conocimiento general y global de los principios en los que se basa esa Nutrición, exponemos a continuación las bases teóricas sobre los procesos metabólicos mediante los cuales el organismo obtiene la energía de los alimentos y la transforma en energía utilizable por el músculo en ejercicio. Algunos conceptos y apartados se repetirán de una manera más práctica en los capítulos posteriores. 51 Centro Olímpico de Estudios Superiores En la realización de cualquier actividad física, el organismo humano necesita energía para; la síntesis de material celular nuevo que reponga el que se ha degradado, el transporte de sustancias contra gradientes de concentración, el mantenimiento de una correcta temperatura corporal y por último y más importante, para la realización del trabajo mecánico, principalmente a nivel muscular. Esta energía, que en circunstancias de ejercicio intenso y/o de larga duración es muy alta, es obtenida fundamentalmente de los alimentos, mediante una serie de procesos metabólicos. El principal medio de almacenar y de intercambiar energía en el organismo humano es el Adenosin Trifosfato (ATP), que libera una gran cantidad de energía al hidrolizarse y soltar sus iones fosfato (Pi), así como otros compuestos con uniones de fósforo de alta energía. Pero hay que reseñar que la cantidad de ATP y de los otros compuestos de alta energía, que puede almacenar el cuerpo humano, es limitada y relativamente baja. Por lo que hay que resintetizarlo cada vez que es utilizado. Para su resíntesis se necesita también energía, que se obtiene de la ruptura escalonada (vías metabólicas) de moléculas más complejas, provenientes del medio ambiente (los alimentos). Las dos vías responsables fundamentalmente de proveer energía para la resíntesis de ATP son; a) la ruptura sin oxígeno de la Glucosa y el Glucógeno, hasta Piruvato o Lactato y b) la oxidación de las grasas, los hidratos de carbono y las proteínas. Desde un punto de vista más bioquímico, las vías de resíntesis de ATP serían (aparte del propio “sistema de los fosfágenos": La glucolítica oxígeno- independiente (anaeróbica). La glucolítica aeróbica. La lipolítica (y proteolítica) aeróbica. 52 Centro Olímpico de Estudios Superiores Si bien ya desde estudios de von Pettenkofer y Voit en 1866 se conocía el papel menor que como substrato energético ejercían las proteínas, estudios posteriores realizados en los años 20 por Krogh y Lindhard y en los años 30 por Christensen y colaboradores demostraron la utilización de una manera diferenciada de las grasas o los hidratos de carbono como substratos energéticos, depend iendo de la duración o intensidad del ejercicio y del tipo de dieta, así como la mayor o menor concentración de ácido láctico en sangre durante ejercicio dependiendo del nivel de condición física (Bang, 1936).Aunque la falta de medios no permitió a estos investigadores, el poder profundizar en el conocimiento mas profundo de los mecanismos que regulaban esos procesos energéticos. El uso de técnicas más complejas e invasivas (sobre todo el uso de la biopsia muscular por aguja en humanos) ha permitido que en la actualidad se tenga un conocimiento mayor de los procesos metabólicos que se producen en nuestro organismo, para obtener energía durante el ejercicio físico, así como de su regulación y sus adaptaciones al ejercicio continuado. 2.1.1 LOS FOSFATOS DE ALTA ENERGÍA. La forma básica de obtención de energía en las células vivas, son los "compuestos de fosfato de alta energía", que son formas de energía química potencial fácilmente convertibles y asequibles para proveer los procesos metabólicos celulares, que sin esa energía no se llevarían a cabo. Se almacenan en la fibra muscular y si no se resintetizaran, darían energía para ejercicios muy intensos (repeticiones máximas de fuerza y/o velocidad), que duren hasta unos 6 segundos aproximadamente Los principales ejemplos de "compuestos de fosfato de alta energía son la Adenosina Trifosfato (ATP) y la Fosfocreatina (PCr), también llamados "fosfágenos de alta energía". La cantidad de estos fosfágenos almacenada en el músculo esquelético humano es muy limitada (60-70 gramos aprox. en el músculo de un deportista entrenado), por lo que en cualquier ejercicio que dure más de 6 seg. la clave estará en la resíntesis de estos compuestos. Las células han desarrollado una amplia variedad de reacciones metabólicas para producir y mantener niveles suficientes de energía química (cuya moneda fundamental son los fosfatos de alta energía). También ha desarrollado mecanismos para dirigir y utilizar esa energía en los procesos y momentos que la célula necesita. La energía puede estar almacenada en la célula en forma de fosfágenos de alta energía o en otros substratos (p.e. glucógeno muscular, triglicéridos) de los que tendrá que transferirse a formas más básicas y asequibles, como son los fosfágenos. 53 Centro Olímpico de Estudios Superiores Una de las características que distinguen a la célula muscular de las otras células, es que su actividad metabólica puede aumentar mucho en muy poco tiempo, como ocurre en el paso del estado de reposo al de contracción. Así por ejemplo el músculo sartorio de la rana puede pasar de unos requerimientos energéticos de unos 7 nMol de O2/gr.peso fresco/min. a tener en pocos segundos de una contracción tetánica isométrica un requerimiento de energía de unos 7000 nMol O2/gr peso fresco/min. Teniendo en cuenta que ese gran aumento ya se inicia a los pocos milisegundos de comenzar la contracción. Es interesante destacar que el paso contrario, es decir, la transición de un estado de elevada actividad metabólica (ejercicio intenso) al estado basal (reposo), dura mucho más tiempo. Debido a todo ello, unos de los problemas fundamentales para el músculo en ejercicio es el poder disponer rápidamente de grandes cantidades de energía, para poder reciclar los fosfágenos que va utilizando. De una manera básica, se podría decir que durante una contracción muscular que se mantiene durante algún tiempo, se aprecia una disminución de los niveles de Fosfatos de alta energía, que es mayor cuanto más intensa sea la contracción y que va acompañado de un aumento en los niveles de Pi y de una disminución casi proporcional del pH intracelular. Nada mas acabada la contracción (es decir, durante el período de recuperación), los niveles de Fosfatos de alta energía aumentan hasta llegar a sus valores basales, siendo mas lento ese aumento cuanto más intensa fue la contracción y más disminuyeron los niveles. El pH intracelular como se mencionó anteriormente, también disminuye cuando la contracción es intensa, llegando incluso a valores de pH 6.0. Al final de la contracción el pH tiende a disminuir algo mas aun (al inicio de la recuperación) y después vuelve lentamente a sus valores basales. 54 Centro Olímpico de Estudios Superiores 2.2. VÍAS METABÓLICAS PRINCIPALES Las vías metabólicas que fundamentalmente aportan energía durante el ejercicio físico son; la glucolisis y la fosforilacion oxidativa. 2.2.1 Glucolisis Esta vía metabólica se volverá a describir con más detalle en el apartado relativo a la degradación de los Hidratos de Carbono, ya que esta vía metabólica produce energía de la ruptura de Hidratos de Carbono sin usar el oxígeno, por eso algunos autores la llaman oxígeno-independiente. (Hawley y Burke, 1998). Y como veremos más adelante aunque en lenguaje “coloquial” se le llama glucolisis “anaeróbica”, en realidad no funciona en ausencia de oxígeno siempre sino que es “independiente de la presenc ia de oxígeno. Esta vía se activa muy rápido cuando se realiza ejercicio intenso, así por ejemplo en ejercicios intensos de unos 30 seg. se calcula que aporta aproximadamente un 60% de la energía. Esta vía está asociada a la producción de lactato en los músculos activos y su liberación y acumulo en la sangre. Este lactato irá; 1) a los capilares que rodean la fibra muscular y transportado en la sangre al hígado donde puede ser convertido en glucógeno (gluconeogénesis), o utilizado como fuente energética por otros órganos (Ej: el corazón). 2) a las fibras musculares cercanas en las que puede ser usado como sustrajo energético. Comentaremos a continuación sólo algunos aspectos relacionados con su regulación. La glucosa que hay en el torrente circulatorio (sobre todo después de las comidas) es transportada dentro de la célula muscular por un mecanismo de transporte facilitado, mediante una proteína insertada en la membrana plasmática, que hace de transportador de la molécula de glucosa. Estos transportadores de glucosa tienen un número limitado, por lo que la capacidad de transportar glucosa puede saturarse. La actividad de estos transportadores puede ser aumentada por la acción de hormonas como la Insulina, y por la actividad contráctil muscular. La acción de la Insulina es clave para que después de la ingestión de Hidratos de carbono se aumente el transporte de glucosa dentro del músculo, con lo que se consiguen dos objetivos; sacar el exceso de glucosa del torrente circulatorio y llenar los depósitos de glucógeno musculares, que serán necesarios para la futura actividad muscular. Este proceso de transporte no requiere energía pues la molécula de glucosa es transportada a favor de un gradiente de concentración, ya que dentro de la célula 55 Centro Olímpico de Estudios Superiores muscular las concentraciones de glucosa se mantienen bajas por la acción de la enzima Hexokinasa (ver Figura 1). FIGURA 1: Esquema simplificado de la glucolisis anaeróbica, resaltando sus puntos clave. PHOS=Glucógeno fosforilasa; GS=Glucógeno sintetasa; HK=Hexoquinasa; PGM=Fosfogluco mutasa; PFK=Fosfofructo quinasa; GPT=Transaminasa glutamico-piruvico; LDH=Lactato deshidrogenasa; PDH=Piruvato deshidrogenasa. En la contracción muscular, debido a que aumentan los niveles intracelulares de Ca2+, hay una mayor degradación de glucógeno a glucosa-1-fosfato (Glu-1-P), que entrará en la vía de la glucolisis (ver Figura 1.). Durante el ejercicio físico hay un gran aumento en la actividad de la glucógeno fosforilasa, en parte debido a los cambios en la concentración intracelular de Ca, pero parece ser que también está influenciado por la presencia de Pi, que proviene de la hidrólisis de la PCr. 56 Centro Olímpico de Estudios Superiores Durante la fatiga disminuye la actividad de la fosforilasa, se piensa que debido a que la caída en el pH intracelular puede inactivar esta enzima. La otra enzima de la que dependían las concentraciones de Glu-6-P es la PFK. Esta enzima es una de las principales reguladoras de la glucolisis. Su actividad se ve aumentada por la presencia de Pi, AMP y NH4, mientras que se inhibe con ATP, protones y Citrato. Siendo la presencia de grandes cantidades de ATP el principal inhibidor del flujo a través de esta enzima. Siguiendo la glucolisis (ver Figura 1.) la glucosa es metabolizada hasta convertirse en Piruvato. El Piruvato puede convertirse en acetil-CoA , reacción catalizada por la Piruvato deshidrogenasa (PDH).El Piruvato también puede pasar a lactato mediante la enzima Lactato deshidrogenasa (LDH). Así pues, en la glucolisis, la glucosa es metabolizada hasta Piruvato. A partir de ese punto puede continuar su metabolización de dos maneras: 1) Mediante la Lactato deshidrogenasa pasa a lactato, con un producto neto de 2 moles de ATP. 2) Mediante varias enzimas mitocondriales, ser oxidada completamente hasta CO2 + H2O con una producción neta de 36 mol ATP. BIBLIOGRAFÍA Peak Performance. Training and nutritional strategies for sport. Eds: Hawley J. and Burke L. Allen & Unwin. St. Leonards. Australia. 1998. 2.2.2 Glucolisis aeróbica y la Fosforilacion Oxidativa 57 Centro Olímpico de Estudios Superiores Los sistemas metabólicos que generan energía de la ruptura de substratos en presencia y utilizando el oxígeno, son los llamados “aeróbicos”: La glucolisis aeróbica (oxidación de hidratos de carbono) y la lipolisis aeróbica (oxidación de grasas). Para ejercicios intensos de larga duración (de 2 a 120 min. según autores), la vía metabólica que más energía aporta es la glucolisis aeróbica. Para ejercicios de más baja intensidad (y más larga duración) la oxidación de las grasas es la vía fundamental. Los factores que determinan el uso de un sustrato energético u otro (p.e.: grasa o carbohidratos) son fundamentalmente; la intensidad y la duración del ejercicio, la condición física de la persona y la dieta. La Fosforilacion Oxidativa es, cuantitativamente, el proceso más importante para aportar energía durante la actividad muscular. En esta vía metabólica, mediante varios complejos enzimáticos que están en la membrana interna de la mitocondria, se cataliza la transferencia de electrones desde cofactores reducidos (NADH, FADH) a oxígeno para formar agua. Del gradiente electroquímico de protones producido, se sintetizará ATP. Esta vía metabólica tiene como substrato metabolitos derivados de la degradación de; lípidos, hidratos de carbono y proteínas (Figura 2). La regulación de esta vía metabólica sigue siendo objeto de controversia, pero la hipótesis mas aceptada propone que la respiración mitocondrial es controlada por la disponibilidad de ADP en la mitocondria y que el paso limitante seria el intercambio de adenin- nucleotidos a través de la membrana mitocondrial. Siendo el parámetro fundamental el cociente ATP/ADP. La capacidad metabólica muscular puede definirse de una manera muy grosera como, la cantidad máxima de ATP que el músculo puede resintetizar (velocidad máxima a la que se puede resintetizar ATP), lo cual incluye; la actividad enzimática, la cantidad de enzima presente en la reacción, la concentración efectiva de substratos, de producto, de activadores y de inhibidores. 58 Centro Olímpico de Estudios Superiores FIGURA 2: Esquema simplificado, de la vía metabólica oxidativa intra-mitocondrial y sus substratos. El músculo se adapta de manera diferente según el tipo de demanda energética que tiene. Para aumentar la cantidad de enzima mitocondrial no aumenta la cantidad dentro de cada mitocondria sino que aumenta el número de mitocondrias. También se producen aumentos en la actividad enzimática de las enzimas que se utilizan, así como en la concentración del substrato utilizado y del producto obtenido. En resumen: 59 Centro Olímpico de Estudios Superiores La moneda básica de intercambio de energía en el metabolismo muscular son los fosfágenos de alta energía (ATP, PCr etc.). La cantidad que de estos compuestos dispone el músculo es limitada, por lo que cuando necesita energía para la contracción muscular, tiene que resintetizar rápidamente los fosfágenos (fundamentalmente ATP) que va utilizando. Los dos procesos principales para proveer la energía necesaria para la resíntesis de ATP son; la degradación de glucosa y glucógeno hasta Piruvato y/o Lactato y la oxidación aeróbica de Lípidos, Proteínas e Hidratos de Carbono. Si un músculo en particular necesita realizar contracciones muy intensas y/o a mucha velocidad, necesitará gran actividad de la ATPasa miosínica, para romper mucho ATP y obtener mucha energía, por lo que necesitará procesos metabólicos rápidos para resintetizar el ATP hidrolizado, como es el caso de la glucolisis anaeróbica. Si por el contrario necesita menos energía pero durante mas tiempo (para ejercicios de baja o moderada intensidad) se hidroliza menos ATP y se resintetiza fácilmente mediante la fosforilacion oxidativa. 2.3 METABOLISMO DE LOS HIDRATOS DE CARBONO 2.3.1 Características Los Hidratos de Carbono, también llamados Carbohidratos, están compuestos de unidades repetidas de CH2O de ahí el nombre de Hidratos de Carbono, pue s a cada molécula de Carbono le corresponde dos moléculas de Hidrógeno y una de Oxígeno (proporción similar a la de la molécula de agua, H2O). Los Carbohidratos mas simples, dentro de los que son metabolicamente importantes, son los azúcares (monosacáridos). Cuando las moléculas de Hidratos de carbono están agrupadas en grandes polímeros, son los llamados Polisacáridos. 60 Centro Olímpico de Estudios Superiores 2.3.2 Fuentes Los monosacáridos más importantes metabólicamente son; la Glucosa y la Fructosa, ambos presentes en muchos alimentos. Su fuente fundamental es la Sacarosa, la cual esta formada de Fructosa y Glucosa (por lo cual es un disacárido). Este disacárido se encuentra en grandes cantidades en el azúcar blanco y moreno y sus derivados. En el aparato digestivo, la Sacarosa se convierte en igual proporción de Glucosa y de Fructosa, si bien la cantidad de Glucosa que se absorbe en el intestino es casi el doble que de Fructosa. Gran parte de la Fructosa es metabolizada en el hígado. Tanto la Glucosa como la Fructosa pueden ser fosforiladas mediante la Hexoquinasa, pero hay que destacar que la afinidad de la Hexoquinasa del músculo por la Glucosa es 10 veces superior que por la Fructosa. La Glucosa es pues el monosacárido principal para el metabolismo del músculo en ejercicio. La fuente fundamental de Carbohidratos son los polisacáridos, principalmente los formados por polímeros de Glucosa, como son los almidones encontrados en los cereales, patatas etc. Estos alimentos son degradados en el estómago e intestino a Glucosa que se absorbe por la sangre y es transportada a los tejidos para su metabolización o su almacenamiento. 2.3.3 Almacenamiento La glucosa se almacena en el organismo principalmente en forma de Glucógeno, que es un polímero (agrupación) de moléculas de Glucosa, si el organismo absorbe mas Glucosa de la que puede utilizar o almacenar en forma de Glucógeno, entonces la convierte en grasa y es almacenada como triglicéridos en el adipocito. La cantidad total de Carbohidratos que se pueden almacenar en el organismo, en forma de Glucógeno es relativamente pequeña, situándose sobre todo en el músculo, en el hígado y algo en el líquido extracelular. Las cantidades en cada uno de estos tres compartimientos pueden oscilar entre 300 y 600 gramos en el músculo de un adulto no 61 Centro Olímpico de Estudios Superiores obeso de 70 Kg. de peso, de 0 a 90 gr. en el hígado y de 8 a 11 gr. en el líquido extracelular, dependiendo estas variaciones en las cantidades, sobre todo de la composición de su dieta, (alto o bajo porcentaje de Hidratos de Carbono) tal como veremos con detalle mas adelante. El Glucógeno es un polímero de Glucosa que se diferencia de los almidones en una mejor disponibilidad de la molécula de Glucosa para ser fosforilada y además un mayor número de puntos donde pueden añadirse moléculas de Glucosa durante la síntesis, poseyendo también una mayor solubilidad. La Glucosa que no se necesita para suplir las necesidades energéticas del organismo, se almacena prioritariamente en el músculo y después en el hígado. De hecho en la recuperación post-ejercicio, cuando el músculo ha consumido su glucógeno, el hígado no absorbe cantidades significativas de glucosa hasta que el músculo ha repleccionado al menos parcialmente sus depósitos de glucógeno. En períodos en los que la persona ingiera una dieta pobre en Carbohidratos, la poca Glucosa que se ingiere tiende a almacenarse en el músculo y no en el hígado. No se conoce con exactitud el mecanismo por el que la Glucosa llena preferentemente los depósitos de Glucógeno del músculo antes que los del hígado, ni la señal que transfiere la información de que los depósitos del músculo ya están suficientemente llenos, para que empiecen a llenarse los del hígado. En estado de reposo, el consumo de Glucosa por el músculo es controlado por acción de la Insulina. El músculo puede también usar lactato, junto con la Glucosa, para la formación de Glucógeno dentro del músculo, lo cual es de gran interés práctico después de un ejercicio muy intenso en el que se ha producido niveles altos de Ácido láctico. En el hígado, la formación y almacenamiento de Glucógeno tiene lugar a partir de otros precursores, además de la Glucosa, como son; el Glicerol, el Ácido Láctico y los aminoácicos. El pico de producción de Glucógeno en el hígado a partir de estos precursores (Gluconeogénesis) coincide con niveles bajos de Glucosa en el espacio extracelular, por ejemplo en ejercicios prolongados y en los períodos de ayuno. 62 Centro Olímpico de Estudios Superiores El contenido de Carbohidratos en la dieta, tiene una marcada influencia en el almacenamiento de los mismos en el organismo en forma de Glucógeno, tanto a nivel de hígado como de músculo. Cuando se consume una dieta muy rica en Hidratos de Carbono durante dos o tres días, que han estado precedidos de varios días de ejercicio muy intenso y/o varios días de una dieta pobre en Hidratos de Carbono, se produce el fenómeno llamado de "supercompensacion", pues los depósitos de Glucógeno en el organismo se duplican o triplican. En el músculo este fenómeno ocurre a nivel local exclusivamente, es decir, sólo en los músculos o fibras musculares que se han ejercitado intensamente. La utilización del fenómeno de "supercompensacion" de Glucógeno en el deporte de competición estuvo muy extendida desde que a finales de los años sesenta fisiólogos escandinavos lo descubrieran , realizándose la llamada "dieta disociada escandinava" que consistía en ingerir durante tres días una dieta muy pobre en Carbohidratos, a la vez que se entrenaba intensamente (con lo cual se vacían los depósitos de Glucógeno) seguidos de tres días de entrenamientos mas suaves y una die ta muy rica en Carbohidratos, tras los cuales se apreciaba en los depósitos de Glucógeno muscular (comprobado mediante biopsias musculares) los grandes aumentos mencionados antes. Actualmente, modificaciones de esta dieta disociada se siguen utilizando en deportes en los que el rendimiento en competición es dependiente de los niveles de Glucógeno almacenados en el organismo, como puede ser la maratón, pruebas ciclistas de un solo día, algunas distancias de natación etc. Las modificaciones de esta dieta disociada se deben a que en algunos casos una dieta estricta muy baja en Hidratos de Carbono, por consiguiente rica en proteínas y grasas, puede producir alteraciones digestivas y es poco agradable de consumir para un deportista. También se deben las modificaciones al hecho de conocer que el estímulo para que se produzca esta supercompensación es la deplección (vaciamiento) de los depósitos de Glucógeno, por lo que vaciando estos depósitos mediante el entrenamiento intensivo, y aumentando después durante varios días la ingesta de Carbohidratos y entrenando menos, se producen supercompensaciones parecidas. Los mecanismos que regulan este fenómeno han sido muy estudiados, pero no se conocen con exactitud, si bien se sabe que contenidos altos de Glucógeno en la célula, son un potente inhibidor de la enzima Glucógeno sintetasa, por lo que contenidos bajos puede que la estimulen mucho. Los músculos muy entrenados aeróbicamente, tienen mayor contenido de la enzima Glucógeno sintetasa, por lo que la supercompensación que se produce en ellos es mayor que en los músculos menos entrenados. 63 Centro Olímpico de Estudios Superiores En cuanto al almacenamiento de Glucógeno, hay que resaltar también que las fibras rápidas (tipo II) pueden almacenar algo mas de Glucógeno que las fibras lentas (tipo I). Cuando se almacena Glucógeno en la célula, se realiza con la asociación de moléculas de agua; en el hígado 1gr. de Glucógeno se almacena asociado a 2-3gr. de agua, en el músculo se cree que la asociación se realiza en cantidades parecidas. Por lo cual hay que asociar una ingesta abundante de líquidos a la dietas de "supercompensacion". El agua que se almacena en el músculo, asociada a las moléculas de glucógeno, puede dar en algunos momentos sensación de pesadez o de hinchazón. 2.3.4 Degradación de los Hidratos de Carbono La degradación completa de los Carbohidratos, se realiza en dos fases distintas; la primera podría denominarse "anaeróbica" y la segunda "aeróbica". En la primera fase se producen una serie de reacciones sin utilización de oxígeno. A este grupo de reacciones se las conoce como "vía de Embden-Meyerhof", también denominada "glucolisis anaeróbica" y a veces simplemente "glucolisis", si bien esta última denominación es demasiado extensa ya que puede abarcar, tanto a la fase anaeróbica como a la aeróbica. (Figura 1). Nos referiremos ahora a la primera fase (glucolisis anaeróbica) de la degradación de los Carbohidratos en el músculo esquelético. Cuando la fuente de Carbohidratos es el Glucógeno, los dos pasos iniciales de la degradación son, el paso a Glu-1-P y su posterior conversión a Glu-6-P. La Glu-6-P es un punto de conexión muy importante en esta vía, ya que también es la forma de entrada de la Glucosa libre proveniente de la sangre, por la acción de la Hexoquinasa. Es entonces en este punto donde se hacen comunes las dos formas de entrada de los Carbohidratos en la Glucolisis anaerobica,en forma de Glu-6-P. La degradación de la Glucosa a través de las diferentes reacciones de la vía de EmbdenMeyerhof , que ocurren en el citoplasma, llega hasta la producción de Piruvato (Fig.1). 64 Centro Olímpico de Estudios Superiores En condiciones normales , con la célula bien oxigenada, la mayoría del Piruvato formado es transportado a la mitocondria donde seguirá su oxidación hasta CO2 y agua. Pero hay dos situaciones en las que el Piruvato, es reducido a Lactato , estas dos situaciones son : -Aporte de Oxígeno insuficiente. -Niveles de Glucogenolisis muy altos. Aunque el aporte de Oxígeno sea suficiente, puede darse el caso de que se produzca mucho Piruvato , entonces el exceso de Piruvato , por la ley de acción de masas, activa la reacción mediatizada por la Lactato deshidrogenasa y se reducirá Piruvato a Lactato. Cuando la intensidad del ejercicio es muy alta, la producción de Lactato aumenta , bien por falta relativa de oxígeno o bien por la necesidad de producir más, y más rápido, energía de la que se produce en los procesos oxidativos de la mitocondria. 2.3.5 Regulacion de la vía de Embden-Meyerhof Todas las vías metabólicas necesitan regular la cantidad de substrato que pase a través de ellas de acuerdo con la cantidad de producto que necesitan. En el caso de la Glucolisis anaeróbica durante el ejercicio parece claro que tendrá que ajustar su funcionamiento a la cantidad de ATP que tiene que reciclar. En la vía de Embden-Meyerhof los dos enzimas claves en su regulacion parece que son la Glucógeno fosforilasa (PHOS) y la Fosfo- fructo quinasa (PFK). La PHOS tal como se explico anteriormente, inicia la degradación del Glucógeno , rompiéndolo en unidades de Glu-1-P. Esta enzima tiene dos subformas; PHOSa y PHOSb. 65 Centro Olímpico de Estudios Superiores La activación de la PHOS puede realizarse de dos maneras, una mediante la elevación de los niveles de Calcio libre (con lo cual la activación estará relacionada con la contracción muscular) y otra por los cambios en las concentraciones de hormonas (fundamentalmente Epinefrina e Insulina) que se producen con el ejercicio.Con lo que podríamos decir que hay una activación "interna" y otra "externa ". En algunos tipos de ejercicio predominara un tipo de activación y en otros predominara otro tipo. Así por ejemplo en ejercicio muy cortos o de tipo isométrico, en los que el flujo de sangre al músculo esta restringido, predominará la activación mediada por la liberación de Calcio por el Retículo Sarcoplásmico .En circunstancias de gran tensión emocional (precompetición) y en ejercicios de larga duración la activación hormonal es crucial. Otros factores pueden aumentar la actividad de la PHOS , como son los aumentos en la concentración de AMP y IMP y posiblemente los de Pi. La PFK es la otra enzima clave en la regulación la esta vía y para muchos autores (debido al hecho de que son muchos los factores que la pueden influenciar), es la principal reguladora. Entre esos factores están; AMP, ATP, ADP, Citrato, Amoníaco y Fructosa difosfato. Las concentraciones altas de ATP inhiben la afinidad de la enzima por la Fructosa-6fosfato , por lo que cuando baja el nivel de ATP, se pierde esa inhibición y aumenta el flujo de substrato a través de la enzima. El aumento de ADP, por el contrario ,libera la inhibición antes mencionada. La combinación de; concentraciones bajas de ATP con concentraciones altas de ADP, es decir una disminución de cociente ATP/ADP elevará la actividad de la PFK. El Citrato parece que actúa sobre la actividad de la PFK aumentando la inhibición que produce el ATP, con lo que elevaciones en las concentraciones de Citrato disminuirán la actividad de la PFK. El Citrato citoplásmico aumenta cuando hay un gran nivel de oxidación de grasas y parte del citrato producido pasa al citoplasma, donde tiene una acción inhibidora de la glucolisis anaeróbica. 66 Centro Olímpico de Estudios Superiores 2.3.6 Oxidación en la mitocondria Cuando la degradación de la Glucosa llega a Piruvato, generalmente este Piruvato en vez de acumularse en el citoplasma, pasa a la mitocondria donde continua su degradación mediante procesos oxidativos en el ciclo de Krebs y la cadena respiratoria, hasta llegar a dióxido de carbono y agua. El paso inicial de este proceso es el transporte del Piruvato dentro de la mitocondria, esto se realiza por medio de una Translocasa situada en la membrana interna de la mitocondria. El Piruvato es oxidado a Acetil Coenzima A por la Piruvato deshidrogenasa, esta enzima está influenciada por varios factores entre los que destaca el cociente intramitocondrial de ATP/ADP, el cociente entre la forma reducida y la oxidada de la Nicotinamida adenina dinucleotido (NADH/NAD) las cantidades de Acetil CoA. Está pues influenciada por la demanda energética en los elementos contráctiles. Avanzando en esta vía metabólica hay otra enzima de gran importancia; la Citrato sintetasa, que sintetiza citrato a partir de Acetil CoA, alimentando de esta manera el ciclo de Krebs. Esta enzima es un importante punto de regulación del ciclo y esta inhibida por la presencia de niveles altos de ATP. De una manera global se podría decir que el control de las diferentes reacciones de esta vía esta mediatizado por la demanda de energía o lo que es lo mismo, por la mayor o menor hidrólisis de ATP durante la contracción muscular. De la misma manera que varias enzimas ven inhibida su actividad por la presencia de concentraciones altas de ATP, las concentraciones altas de ADP aumentan la actividad de esas enzimas. Por ello muchos autores consideran que el principal parámetro regulador es el cociente ATP/ADP o el cociente ATP/ADP-Pi. 67 Centro Olímpico de Estudios Superiores Resumen: La visión clásica de las fuentes energéticas, nos dice que son: 1) Fosfágenos, para unos segundos o para unas contracciones de fuerza máxima. 2) Glucolisis anaeróbica para ejercicios de alta intensidad de hasta 1 min aproximadamente de duración. 3) Metabolismo aeróbico oxidando carbohidratos (CH) –glucógeno, glucosa- lípidos – TG: triglicéridos y FFA: acidos grasos libres, - y en menor proporción y en ciertas circunstancias; proteínas, cuerpos cetónicos y acetato. Una visión más actual de las tres vías de optación de energía durante el ejercicio debe ser completada con una división en dos del (3) metabolismo aeróbico: 3,1) Glucolisis aeróbica, en la que se oxidan CH para ejercicios de resistencia de alta intensidad. Una buena medida de este sistema sería teóricamente el VO2max. y parámetros cardiorespiratorios (cambios en el cociente respiratorio) combinados con valores de baja producción de lactato podrían indicarnos la utilización de este metabolismo energético. 3, 2) Lipolisis aeróbica (lo de aeróbica sobraría...), en la que se oxidan lípidos para obtener energía en ejercicios de media y baja intensidad y de larga y muy larga duración. La medición de la potencia lipolítica es difícil, sin embargo podría utilizarse el cociente respiratorio en tests submáximos. 2.3.7 Utilización La mayoría de los ejercicios incluyen una combinación de metabolismo de grasas y de carbohidratos para aportar ATP a través del metabolismo aeróbico. Sin embargo conviene recordar que el rango máximo de síntesis de ATP a partir de grasas, sólo puede proveer ATP para mantener un ejercicio a intensidades entre el 55-75% del VO2max, dependiendo del nivel de entrenamiento aeróbico del individuo. El metabolismo oxidativo de los carbohidratos podría mantener un ejercicio a intensidades del 100% del VO2max, tanto en personas entrenadas como no entrenadas. 68 Centro Olímpico de Estudios Superiores La producción ANAERÓBICA de ATP, se utiliza cuando la fosforilación oxidativa (metabolismo aeróbico) no puede proveer el suficiente ATP para los requerimientos de la intensidad del ejercicio. Esto ocurre a intensidades de ejercicio del 85-100% del VO2max y por supuesto en trabajos superiores al 100% del VO2max. Las fuentes de energía principales para la síntesis anaeróbica de ATP son; la degradación de fosfocreatina (PCr) y la glucolisis anaeróbica con producción de lactato e H+. La ventaja de vía anaeróbica es su velocidad de producción de ATP (5-6 veces más alta que la vía aeróbica), pero su mayor limitación es el corto periodo de tiempo que se puede mantener funcionando, debido a la deplección de PCr y a la acumulación de metabolitos de la glucolisis anaeróbica. Así pues, es importante recordar que los CH son un substrato energético tanto para la obtención aeróbica de ATP como para la anaeróbica (metabolización aeróbica de 1 mmol de glucosa=38-39mmol de ATP, metabolización anaeróbica de 1mmmol glucosa=3mmol de ATP por mmol de glucosa derivada del glucógeno). Por lo que en estas situaciones anaeróbicas se obtiene poca, pero rápida energía del glucógeno y en proporción se gasta mucho más glucógeno. METABOLISMO AERÓBICO de los CARBOHIDRATOS. Revisaremos los puntos fundamentales A- Transporte de glucosa al músculo. Este proceso es facilitado mediante transportadores. Por otro lado, el transporte está limitado por la propia utilización de la glucosa. En el músculo los transportadores son el 1 (GLUT1) y el 4 (GLUT4). El GLUT1 (presente en la membrana plasmática del músculo) está sobre todo relacionado con la entrada de glucosa en reposo y se cree que aumentan con el entrenamiento. El GLUT4 por el contrario es intramuscular, y responden a la insulina y a la contracción muscular. El entrenamiento de resistencia aumenta el contenido muscular de GLUT4 y la respuesta a la insulina y a la contracción mus cular. Las personas entrenadas son más sensibles a la insulina, “translocan” más GLUT4 a la superficie de la célula muscular en respuesta a la misma dosis de insulina. Este aumento en la respuesta debida al entrenamiento, permite a las personas entrenadas el tener una mayor entrada (en valores absolutos) de glucosa en el músculo. B- Glucolisis. La primera enzima importante en esta vía metabólica es la hexoquinasa (HEX) que se estimula por la insulina y por la concentración de glucosa libre. Su actividad permite mayor utilización de la glucosa cuando el transporte de glucosa hacia el interior del músculo es alto. Así mismo, la actividad de la HEX aumenta con el entrenamiento físico. Cuando ya se ha fosforilado la glucosa, la principal enzima 69 Centro Olímpico de Estudios Superiores reguladora es la fosfofructoquinasa (PFK). El principal regulador de esta enzima es el ATP. Concentraciones altas de ATP inhiben la PFK en reposo y en ejercicio. La presencia de Citrato y H+ potencia la inhibición realizada por la presencia de ATP. Cuando se degrada mucho ATP (durante ejercicio) el aumento de ADP, Pi y AMP estimulan la actividad de la PFK y también activan la glucógeno fosforilasa. Así pues la caída de energía interna en la célula muscular causada por las contracciones del ejercicio aumentan la glucolisis mediante una activación de la glucógeno fosforilasa y quitando la inhibición de la PFK. C- Glucogenolisis. El primer paso de la glucogenolisis es la fosforilación de una molécula final de glucosa mediante la glucógeno fosforilasa (PHOS), que tiene dos subformas; la b es menos activa y la a es más activa. La transformación de la forma b en la a ocurre en ejercicio por la estimulación de Ca2+ y en menor intensidad de la epinefrina. En los inicio de ejercicio aunque sea aeróbico lipolitico, se activa la PHOS por la presencia inicial, hasta que ya funciona “bien” la via aeróbica, de ADP AMP y Pi. D- Piruvato Deshidrogenasa. La entrada de CH en la mitocondria y su posterior oxidación metabólica, está regulada primariamente por la enzima mitocondrial, Piruvato Deshidrogenasa (PDH), que es un complejo enzimático compuesto de varias enzimas. Está regulada sobre todo por la presencia de sustrato, de piruvato, de Ca2+ y de ADP. REGULACIÓN COORDINADA del METABOLISMO de las GRASAS y de los CARBOHIDRATOS. La grasa y los CH se combinan para aportar energía al músculo durante la mayoría de las intensidades de ejercicio aeróbico. La proporción relativa en la que lo hacen las grasas o los CH depende de la intensidad del ejercicio, su duración, el nivel de entrenamiento aeróbico de la persona, sus hábitos dietéticos (CH, grasas, cafeína), la ingesta de CH previa y durante el ejercicio, y posiblemente la edad y el sexo. La interacción entre el metabolismo de las grasas y el de los CH durante ejercicio físico se cree que depende de la disponibilidad de substrato y del balance entre las actividades de las enzimas llave. 70 Centro Olímpico de Estudios Superiores Utilización en ejercicio El porcentaje de Hidratos de carbono que se utiliza durante las diferentes intensidades de ejercicio físico puede ser determinada, conociendo el cociente respiratorio (relación entre el volumen de CO2 espirado y el de O2 consumido, VCO2/VO2), si bien hay que tener en cuenta que este criterio es válido siempre que la dinámica ventilatoria no este alterada, y que para la cantidad de CO2 que se elimina sea el producto del metabolismo celular. Por lo cual hay diversas situaciones en las que no se cumple este criterio. Las dos mas importantes son: Hiperventilación y Acidosis metabólica. Con la Hiperventilación pulmonar se baja la presión alveolar de CO2 , con lo que no se descarga CO2 de la sangre. Con la Acidosis metabólica (como en el caso de gran producción de Ácido Láctico) se aumentara la liberación de CO2. En las condiciones standard que se mencionaron anteriormente, el cociente respiratorio es fiel valoración del metabolismo de los tejidos, esto ha sido ampliamente demostrado en estudios en los que se comparaban estos valores, con los obtenidos de muestras de sangre arterial y venosa de extremidades en ejercicio , con los obtenidos de biopsias musculares y con los obtenidos utilizando isótopos marcados. 2.3.7.1 Ejercicio ligero. Consideraremos ejercicio ligero, el realizado a una intensidad entre el 30 y 40 % del consumo máximo de oxígeno de la persona . Con esta intensidad de ejercicio el cociente respiratorio oscilará en valores cercanos a 0.80 - 0.85, bajando incluso a valores inferiores, si el ejercicio continúa durante mucho tiempo. En condiciones normales, una persona no llega al agotamiento con esta intensidad de ejercicio, ni hay una gran utilización del Glucógeno muscular. Los niveles de Ac. Láctico en sangre no aumentan. Los niveles de Glucosa en sangre permanecen estables, lo cual quiere decir que el consumo de Glucosa por el músculo, esta equilibrado con la liberació n de Glucosa por el hígado. Esta intensidad de ejercicio puede ser mantenida durante mucho tiempo (horas). 71 Centro Olímpico de Estudios Superiores Durante ejercicios ligeros de muy larga duración (240 minutos) la cantidad de Glucosa sintetizada en el hígado mediante la Gluconeogénesis y liberada a la sangre para su utilización por el músculo, parece que no es superior a un 8% de la cantidad total utilizada. Siendo todavía menor en ejercicios mas intensos. 2.3.7.2 Ejercicio moderado. Consideraremos ejercicio moderado el realizado a una intensidad entre el 60 y 70% del consumo máximo de oxígeno de la persona. El cociente respiratorio estará cercano a 0.90 y no bajará durante el tiempo de ejercicio. Con esta intensidad, después de varios minutos se empieza a notar la sensación de fatiga, que irá en aumento y dependiendo de la motivación de la persona, después de 1 a 3 horas se llega al agotamiento. La utilización del Glucógeno muscular es muy rápida en los primeros 20 ó 30 minutos de ejercicio, luego es algo mas lenta hasta que se llega al vacia miento total de los depósitos de Glucógeno. Al inicio del ejercicio son las fibras lentas (tipo I) las que se vacían de Glucógeno principalmente. Cuando una parte grande de fibras lentas se han vaciado de Glucógeno, se reclutan más fibras rápidas y ambas, lentas y rápidas, pierden Glucógeno rápidamente. En los primeros 5-10 minutos de este ejercicio se encuentran los niveles mas altos de Lactato en sangre, pero cuando las fibras rápidas son reclutadas los niveles de Lactato disminuyen. Con esta intensidad de ejercicio, los niveles de Glucosa en sangre, permanecen bastante estables hasta que las fibras se van vaciando de Glucógeno y tienen que utilizar más las fuentes extramusculares de energía y el hígado es incapaz de liberar suficiente Glucosa. 72 Centro Olímpico de Estudios Superiores El agotamiento suele coincidir con el vaciamiento total de los depósitos de Glucógeno en hígado y músculo. 2.3.7.3 Ejercicio intenso. Consideraremos intenso, el ejercicio realizado aproximadamente al 90% del consumo máximo de oxígeno de la persona. Con pulsaciones cercanas a la máxima frecuencia cardíaca. Con esta intensidad, la persona puede llegar al agotamiento a los 5 minutos de ejercicio, aunque dependiendo de los niveles de motivación y de entrenamiento, este agotamiento puede retrasarse incluso hasta los 60 minutos. Los valores de cociente respiratorio están alrededor de 0.95 o más. La deplección de Glucógeno es rápida, si bien no suelen vaciarse totalmente todas las fibras, cuando se llega al agotamiento. Tanto las fibras lentas como las rápidas se vacían, pero las rápidas lo hacen más, posiblemente debido a que tienen una mayor capacidad glucogenolítica. Las concentraciones de Ac.láctico en sangre llegan a valores por encima de 5mmol/L. a los pocos minutos. A diferencia de las intensidades mencionadas anteriormente, los niveles de Glucosa en sangre durante ejercicios de elevada intensidad aumentan algo y los niveles de Insulina están disminuidos. 73 Centro Olímpico de Estudios Superiores En ejercicio intenso, la fuente primaria de energía son los Hidratos de Carbono si bien no se llega al vaciamiento total de los depósitos de Glucógeno, por la menor duración del ejercicio. Las fibras que más se depleccionan de Glucógeno, son las fibras rápidas. 2.3.7.4 Ejercicio estático. Las contracciones estáticas, tienen la particularidad de que a partir de una intensidad superior al 20% de una contracción voluntaria máxima (MVC), la presión intramuscular que se desarrolla, disminuye el flujo de sangre en el músculo, con lo que limita las fuentes de energía a las exclusivamente intramusculares y minimiza el efecto de las hormonas y metabolitos sanguíneos. En contracciones estáticas intensas, la cantidad de Glucógeno muscular degradado por la vía glucolítica anaeróbica (el aporte de oxígeno también esta disminuido al estarlo el flujo de sangre) es muy alta, pero la cantidad de Lactato total liberado a la sangre es pequeña, por la poca duración del ejercicio. Si la contracción estática no es muy intensa (50% de MVC) los valores de Lactato serán mayores, pues la contracción durará mas tiempo. Cuando las contracciones se realizan de una manera intermitente, el músculo podrá utilizar sustratos energéticos extramusculares, pues en los momentos de relajación la tensión intramuscular disminuye y la sangre puede fluir dentro del músculo. 2.3.8 Regulacion de la utilización de Carbohidratos. La utilización de las fuentes de Carbohidratos intra o extramusculares durante el ejercicio esta regulada por diferentes estímulos.El transporte a través de la membrana sarcoplásmica de la Glucosa extramuscular, aumenta con la contracción muscular , independientemente de los niveles de Insulina, de hecho, el, músculo puede aumentar su transporte y consumo de Glucosa durante la contracción, en ausencia de Insulina. En el músculo tanto la Insulina como la contracción, pueden aumentar el transporte de Glucosa y parece que el efecto de ambos es aditivo. Se especula en la actualidad, que este transporta pueda estar regulado por las concentraciones intracelulares de Calcio. El consumo de Glucosa extracelular por el músculo, aumenta conforme se van acabando los depósitos de Glucógeno.Puede que la Glucosa extramuscular, no entre 74 Centro Olímpico de Estudios Superiores antes porque la Glucosa procedente de la degradación del Glucógeno muscular haga menor el gradiente entre la Glucosa extra e intramuscular.En este transporte la Insulina tiene un papel menor. La utilización de Glucosa sanguínea por el músculo conlleva una producción de Glucosa por el hígado ,que se ajusta muy bien a las cantidades consumidas por el músculo. Este equilibrio esta regulado por los niveles de Insulina y Glucagon , estimulados por los niveles de Glucosa en sangre. 2.3.9 Metabolismo del Ácido Láctico Tal como se mencionó anteriormente, la degradación de la molécula de Glucosa con producción de ATP, se puede llevar a cabo sin la presencia de oxígeno, con la consiguiente producción de Lactato. Esta parte del metabolismo de los Carbohidratos es de gran importancia, pues permite al músculo obtener energía de una manera muy rápida y sin depender de los mecanismos de transporte de oxígeno, sin embargo la cantidad total de energía que produce es menor que cuando hay una oxidación completa y el acúmulo de Lactato en el músculo será un mecanismo inductor de fatiga. Los factores que controlan la producción de Lactato en el músculo en ejercicio están siendo debatidos continuamente. Clásicamente se pensaba que cuando el músculo se contraía sin la presencia de oxígeno era obligatoria la producción de Lactato y que cuando el músculo tenía oxígeno suficiente, no produciría Lactato. Pero hay varias evidencias que cont radicen esta idea: Primera; se ha podido comprobar que en algunos ejercicio hay acúmulo de Lactato en el músculo bastante antes que empiece a ser insuficiente el oxígeno. Segunda; se comprueba fácilmente que después de un período de entrenamiento, cualquier persona realizando una misma carga de trabajo que antes del período de entrenamiento , producirá menos Lactato , a pesar de que para una misma carga de trabajo el flujo de sangre es el mismo , las diferencias arterio-venosas y el consumo de oxígeno iguales. Tercera; con estudios en los que se ha medido la tensión de oxígeno en el músculo se ha podido comprobar que el Lactato se puede producir en condiciones totalmente aeróbicas, si bien cuando el aporte de oxígeno es insuficiente, la formación de Lactato se acelera. 75 Centro Olímpico de Estudios Superiores Así pues, la disponibilidad de oxígeno por el músculo, no es la única causa responsable del aumento en la producción de Lactato muscular. Considerando que el Lactato se puede producir en condiciones aeróbicas, en algunas situaciones su producción será debida a un disbalance entre la producción de Piruvato y el Piruvato que se necesita (o se puede utilizar) para los procesos oxidativos intramitocondriales. En este caso el exceso de Piruvato, simplemente por efecto de la ley de acción de masas, se reducirá a Lactato, por la disponibilidad grande de Piruvato y de NADH, ya que la Lactato deshidrogenasa actúa siempre que disponga de los substratos necesarios para la reacción. También puede darse el caso, de que aunque las condiciones de ejercicio sean aeróbicas, la mitocondria no pueda disponer de suficiente oxígeno, por lo que a nivel mitocondrial, tendríamos una situación de hipoxia que aceleraría la producción de Lactato. Es importante destacar que la cantidad y la velocidad de producción de Lactato, dependerá de la mayor o menor concentración de enzimas de la vía de EmbdenMeyerhof (lo cual estará en relación con el tipo de fibra muscular). Para que funcione totalmente esta vía se necesita un tiempo mínimo para activar todas sus enzimas, esta es una de las causas por las que ejercicios intensos de muy corta duración (sprint) aumentan muy poco la concentración de Lactato. El Lactato producido en el músculo y eliminado a la sangre, puede ser utilizado por el músculo como fuente energética. Esto se comprueba al observar que en ejercicio de moderada intensidad y larga duración, al inicio hay un aumento en las concentraciones plasmáticas de Lactato, que después van gradualmente disminuyendo a pesar de mantener la misma intensidad de ejercicio. También se ha podido comprobar utilizando Lactato marcado con C14 y observando como es tomado del plasma y oxidado en el músculo. Es difícil evaluar la cantidad de Lactato que obtenido de la sangre es oxidado por el músculo, pero algunos autores consideran que será, aproximadamente del 2 al 8% del metabolismo muscular total. Siempre tratándose del músculo esquelético, pues el músculo cardíaco es diferente, por su gran capacidad de obtener energía de sustratos muy variados. 76 Centro Olímpico de Estudios Superiores 2.3.10 Hidratos de Carbono y Fatiga Son muchos los mecanismos y factores que pueden ser responsables del fenómeno de la fatiga, (tal como se verá mas adelante en el Módulo dedicado a "Fatiga, sobrecarga y sobreentrenamiento") de los que están relacionados con el metabolismo de los Carbohidratos destacaremos; a) la disminución en el pH producida por el acúmulo de Ácido Láctico y b) la falta de substratos energéticos. a) Muchas enzimas del metabolismo muscular son sensibles a la disminución de pH creada por la producción de Lactato, una de las que ve más disminuida su actividad por caídas de pH es la PFK, si bien, in vivo, no disminuye tanto como se pensaba hace unos años. La disminución del pH también tiene un efecto en las propiedades contráctiles del músculo, pues reduce la afinidad del Calc io por sus sitios de unión en las proteínas contractiles, por lo que disminuye la tensión desarrollada. Este efecto es más pronunciado en las fibras rápidas, en las que un pH de 6.5 reducirá la tensión en un 50% aproximadamente. b) Durante ejercicio de larga duración el Ácido Lactico no se acumula y no hay disminución del pH, sin embargo en este tipo de ejercicio lo que es crucial es el poder disponer de substrato energético suficiente. Por ello es muy importante el disponer de suficientes depósitos de Glucógeno, para retrasar la aparición de la fatiga. En muchos experimentos se ha comprobado que aumentando el contenido en Glucógeno muscular, se mejora claramente el rendimiento, en ejercicios de intensidad por encima del 5060% de consumo máximo de oxígeno. Un contenido alto en glucógeno, permitirá al músculo realizar ejercicios de más larga duración, pero no le servirá para desarrollar intensidades más altas. 77 Centro Olímpico de Estudios Superiores En resumen; en ejercicios de moderados e intensos, el momento de fatiga estará relacionado con el vaciamiento de los depósitos de Glucógeno en el músculo. En intensidades altas de ejercicio, el músculo utiliza preferencialmente sus reservas de Glucógeno: por su proximidad (al tenerlas dentro del mismo músculo y no tener que depender del transporte en la sangre y del paso de la membrana) y porque puede metabolizarlo tanto aeróbica como anaerobicamente (con lo cual no depende de disponer o no de suficiente oxígeno) 2.3.11 Entrenamiento Con el entrenamiento llamado de "resistencia" o de "larga duración" se depende menos de los Hidratos de Carbono como fuente energética, debido en parte al mayor número de mitocondrias en el músculo que ha estado entrenando. Con el aumento en el número de mitocondrias, así como en la actividad de las enzimas mitocondriales el aporte de energía al músculo se hace menos dependiente de la obtenida a través de la vía de Embden-Meyerhof lo cual también disminuirá la cantidad de Piruvato producida y las posibilidades de ese Piruvato de pasar a Lactato. Hay entonces un "ahorro de Glucógeno" pues el músculo al tener más mitocondrias trata de utilizar otras fuentes energéticas mediante procesos oxidativos, ya sea grasa o glucosa. Este "ahorro de Glucógeno" es muy importante para el mejor rendimiento deportivo, ya que entre dos deportistas compitiendo en resistencia, el que esté mejor entrenado utilizará más tiempo la oxidación de las grasas como fuente energética y tendrá más Glucógeno disponible para la última fase de la competición, que es cuando normalmente hay que subir la intensidad y es más dependiente de los mayores o menores depósitos de Glucógeno. Como en los ejercicios de moderada y alta intensidad, el momento del agotamiento esta relacionado con el vaciamiento de los depósitos de Glucógeno en el músculo, cualquier adaptación que conlleve a un menor uso en el músculo de sus depósitos de Glucógeno (o a un aumento de ellos), retrasará la aparición de la fatiga y mejorará el rendimiento deportivo. Ejemplo de ello sería el entrenamiento de "resistencia" (y las modificaciones en la dieta). El entrenamiento de "resistencia" produce adaptaciones locales en el 78 Centro Olímpico de Estudios Superiores músculo, por las que habrá una mayor utilización-oxidación de la grasa, para un mismo trabajo submaximo, con lo que hay una menor utilización (y un "ahorro") de Glucógeno. Con el entrenamiento de "fuerza", no se producen aumentos en el número de mitocondrias ni en el de capilares, con lo que el músculo dependerá más aún de las reservas energéticas que tenga dentro de la célula muscular es decir, de los depósitos de Glucógeno y de los Fosfágenos. Con el entrenamiento de "velocidad" se aumentan las concentraciones de las enzimas de la vía de Embden-Meyerhof (ya están con niveles altos en las personas sedentarias, pero aumentan aún más con entrenamiento anaeróbico) lo cua l hará que sea mayor y más veloz la degradación del Glucógeno a través de esta vía metabólica y por consiguiente sea mayor la producción y acúmulo de Lactato. 2.4 METABOLISMO DE LAS GRASAS El uso de la grasa como fuente energética para los ejercicios de muy larga duración, es conocido desde que estudios realizados en aves y peces migratorios, confirmaron que estos animales en sus largos viajes, tienen como principal fuente energética la grasa. También se conoce desde los primeros exploradores polares (que utilizaron en sus expediciones alimentación muy rica en grasa) que en el hombre, es también la fuente principal de energía para largos desplazamientos. Todo ello se confirmó en estudios que, valorando el cociente respiratorio, demostraron que la grasa era el substrato energético principal en reposo y en ejercicio de baja intensidad. 2.4.1 Características Dos características de la grasa la hacen la fuente ideal de energía, para los esfuerzos mantenidos durante largo tiempo, cuando el aporte dietetico es limitado. Estas dos 79 Centro Olímpico de Estudios Superiores características son; 1) la grasa contiene mas del doble de energía por unidad de peso que los Hidratos de carbono, 2) la grasa, a diferencia de los Hidratos de carbono, no se almacena hidratada en el organismo. Los compuestos que denominamos GRASAS son un grupo muy amplio de sustancias con el denominador común de ser insolubles en agua y solubles en disolventes orgánicos. Nos centraremos fundamentalmente en las grasas que son importantes como substrato energético para el músculo, estas son los Ácidos Grasos Libres (FFA) y su forma de almacenamiento que son los Trigliceridos (TG). Los FFA no están en forma libre en el plasma sino que están ligados a albúmina (0.01% están realmente en forma libre). Los lípidos “oxidables” como fuente energética son: FFA, Triglicéridos intramusculares (IMTG), Triglicéridos circulantes en el plasma (TG). Estos TG del plasma suelen estar incorporados en la lipoproteinas (quilomocrones, VLDL, LDL, IDL, HDL), que tienen diferentes funciones; VLDL, es la principal transportadora de triglicéridos del hígado al tejido adiposo y al músculo, mientras que HDL transporta colesterol de los tejidos periféricos al hígado. Por ello se cree que los quilomicrones (formados en la pared del epitelio intestinal, pasan a la sangre despues de pasar por el sistema limfático) y las VLDL juegan un papel mayor en el metabolismo energético. (Los FA provienen fundamentalmente de los TG del tejido adiposo, de los quilomicrones, las VLDL y los IMTG.). También otros compuestos derivados de las grasas, como los cuerpos cetónicos pueden servir de fuente energética mientras que la GLICERINA puede ser convertida en glucosa en la gluconeogénesis hepática y luego ser oxidada como glucosa. Los Ácidos Grasos tienen como fórmula general: CH3(C H2)nCOOH. siendo de mayor o menor longitud dependiendo del numero de unidades de CH2 que tengan. Los mas importantes metabólicamente son los de 12 a 18 carbonos. También pueden variar dependiendo de si su estructura esta saturada o insaturada. En este apartado tomaremos como ejemplo básico el Ácido Palmítico y su oxidación, pues representa el 75% del total de los Ácidos Grasos en el plasma humano. 80 Centro Olímpico de Estudios Superiores Una importante propiedad de las grasas comparadas con los Hidratos de carbono, es que los átomos de Carbono y de Hidrógeno son casi un 90% del total de la molécula del las grasas, mientras que en los Hidratos de carbono son un 50% aproximadamente. Esto es de interés por ser la oxidación del Carbono y el Hidrógeno de la molécula las responsables de la liberación de energía, es por ello que la cantidad de energía almacenada por unidad de peso es mayor en las grasas que en los Carbohidratos. Pero hay que destacar otra característica de los FFA y es el hecho de que a altas concentraciones, son tóxicos para la célula. Esta toxicidad se debe al efecto desestabilizador de membrana que tienen, por lo cual sus concentraciones dentro de la célula deben de mantenerse bajas de lo contrario dañarían la membrana mitocondrial. 2.4.2 Fuentes Las grasas que utilizamos para obtener energía o que almacenamos en nuestro organismo, provienen de la ingestión directa de grasas en la dieta y de la conversión (mediante la Lipogénesis) de los excesos de Hidratos de carbono y las Proteínas en grasa. Es decir, no es sólamente la grasa que ingerimos en la dieta la que se va a almacenar en el organismo si no se utiliza, sino que cuando se ingieren Hidratos de carbono o Proteínas en cantidades que no son utilizadas, se almacenan en forma de grasa, mediante un proceso de conversión denominado Lipogénesis, que se realiza fundamentalmente en el Hígado y en el tejido adiposo. La mayor o menor cantidad de Lipogénesis dependerá del total de calorías ingeridas y de las utilizadas. Las fuentes de grasa en la dieta son muy variadas, ya que gran variedad de alimentos contienen grasa en una mayor o menor proporción, si bien el tipo de grasa difiere de unos alimentos a otros. 2.4.3 Almacenamiento. La grasa se almacena principalmente en forma de Trigliceridos (TG) en el tejido adiposo (subcutáneo y visceral profundo) y en pequeñas cantidades en otros tejidos. El cuerpo humano tiene una gran capacidad para almacenar grasa, por ello el porcentaje de grasa que contiene nuestro organismo es relativamente alto, pudiendo oscilar entre valores del 5 al 50%. Los valores medios para hombres están entre el 15 y el 25%, 81 Centro Olímpico de Estudios Superiores mientras que para mujeres el rango es de 25 al 35%. Como se puede apreciar por estos valores medios, los porcentajes de grasa son normalmente mas bajos en hombres que en mujeres. Así pues una mujer de 60 Kg. de peso y un porcentaje de grasa corporal del 25% tendrá aproximadamente unos 15 Kg. de grasa en su organismo. Un hombre de 80 Kg. con porcentaje de grasa del 20%, tendrá unos 16 Kg. de grasa. Estos kilos de grasa implican un almacén de energía que oscila alrededor de 160.000 Kcal., mientras que el rango de calorías almacenadas en forma de Carbohidratos es de 2500 a 4500 Kcal. aproximadamente. Si sólo grasa o carbohidratos se utilizaran para un ejercicio continuo, con los depósitos normales de CH tendríamos energía para 95 minutos de una carrera de maratón, mientras que si se utilizaran los depósitos de grasa tendríamos para 119 horas de maratón continuo. La grasa se almacena principalmente en el tejido adiposo, sobre todo el situado a nivel subcutáneo. Este tejido es muy dinámico y tiene muchas variaciones. Las variaciones se deben fundamentalmente al balance o disbalance entre la ingesta y el gasto calórico, así pues, estados nutricionales negativos o mucha actividad física producirán una disminución del tamaño de los adipocitos y la cantidad de grasa disminuirá. El exceso en la ingesta calórica inducirá un agrandamiento de los adipocitos, debido a que absorben FFA y los almacenan en forma de Trigliceridos. Debido al hecho de que la mayor parte de la grasa se almacena en el tejido subcutáneo y a que las variaciones en la cantidad de este tejido están en relación directa con las fluctuaciones en el contenido en grasa en el cuerpo humano, se realizan mediciones antropométricas simples valorando en diferentes puntos del cuerpo este tejido, para calcular el porcentaje de grasa que tiene el cuerpo de la persona. Aunque la mayoría del tejido adiposo esta situado a nivel subcutáneo, también lo hay en otros puntos del organismo, es importante señalar que a nivel muscular también hay depósitos de tejido adiposo, situados alrededor de las fibras musculares. En algunos animales estos depósitos son tan grandes que se pueden identificar a simple vista. Dentro de la fibra muscular parece ser que también hay depósitos de lípidos en forma de Trigliceridos (IMTG), en una cantidad difícil de valorar, pero que es mayor en las fibras lentas (Tipo I). 82 Centro Olímpico de Estudios Superiores Los FFA (también llamados ahora Fatty Acid "FA") dan más ATP por molécula que la glucosa. Así, 1 molécula de Glucosa puede producir 38 ATP mientras que una de ácido esteárico 147, pero para esta oxidación, la molécula de grasa necesita mas oxígeno que la de glucosa (6 mol oxígeno , 26 mol oxígeno), y además por unidad de tiempo, se obtiene más ATP de una molécula de glucosa que de grasa. Es decir, es un proceso más lento. El porqué, no se sabe todavía con exactitud. 2.4.4 Regulacion del metabolismo de los Lípidos Por la acción de las hormonas adrenérgicas en el tejido adiposo se produce la hidrolisis de los Triglicéridos. Esta acción la realiza fundamentalmente la Norepinefrina (NE) que activa a través del AMPcíclico una quinasa proteínica en el adipocito que a su vez activará la Lipasa del adipocito, que realizará la hidrolisis de los Trigliceridos en Glicerol y FFA, que pasan a la sangre. Los FFA son transportados por la sangre (unidos a la Albúmina) a los tejidos periféricos, este transporte es constante. En los tejidos periféricos, por ejemplo el músculo, se produce un proceso inverso al ocurrido en el adipocito, ya que el complejo FFA-Albúmina se rompe y el FFA es transferido al interior de la célula muscular donde será utilizada para obtener energía inmediatamente o se almacenara provisionalmente en forma pequeñas gotas lipídicas. Estas gotitas de lípidos se sitúan generalmente cerca de las mitocondrias y su peso y cantidad son muy bajos. En reposo al ser las necesidades energéticas muy bajas, los FFA sobrantes se almacenan en la gotitas lipídicas, pero durante la actividad física se oxidan para obtener energía (ver Figura 6). La oxidación de los lípidos se inicia en la membrana externa de la mitocondria, donde por medio de la enzima Acyl CoA sintetasa, el FFA pasa a Acyl CoA. Como la oxidación se realiza en la matriz mitocondrial el Acyl CoA tiene que ser transferido al interior de la mitocondria, esto se realiza por medio del sistema Carnitina-AcylTransferasa situado en la membrana interna de la mitocondria, que une la Carnitina al Acyl CoA y lo transporta al interior de la mitocondria. Se especuló durante varios años, sobre la posibilidad de que este paso pudiera estar limitado por la mayor o menor cantidad de Carnitina en el organismo y que en personas muy entrenadas, con una gran capacidad de utilizar la grasa como fuente de energía, la cantidad de Carnitina fuera un factor limitante, por lo que se usó la ingestión oral de Carnitina como ayuda ergogénica en varios deportes de gran componente aeróbico (Sin que tenga ningún efecto demostrado en la oxidación de grasas, tal como se verá en el capítulo dedicado a las Ayudas Ergogénicas). 83 Centro Olímpico de Estudios Superiores Las unidades de Acetil-CoA siguen en la mitocondria los reacciones de la B-oxidación y entran luego en el ciclo de Krebs o del Ácido Cítrico a través de la reacción catalizada por la Citrato sintetasa, siguiendo el mismo proceso que los acetiles provenientes del Piruvato. La energía obtenida por la degradación total de un Mol de FFA es aproximadamente de 129 Mol de ATP (variando según el tipo). Tal como se mencionó en el apartado corespondiente al Metabolismo de los Hidratos de Carbono, la oxidación de las grasas, tanto a nivel de músculo cardíaco, como de músculo esquelético, inhibe la captación de glucosa por el músculo y la glucolisis, teniendo entonces un efecto positivo en el ahorro de glucógeno. Este efecto esta mediado por ; a)la acumulación de Citrato, que inhibe la FFK, lo cual produce un aumento de la Glucosa-6-fosfato y una inhibición de la Hexoquinasa. b) la inhibición de la Piruvato deshidrogenasa que se produce por el acumulo de productos de la oxidación de los Acidos Grasos, principalmente Acetil-CoA y NADH. 2.4.5 Movilización La movilización de las grasas (lipolisis) se produce fundamentalmente por acción hormonal; la Insulina lo disminuye, y lo aumentan Glucagón, Epinefrina, Norepinefrina, GH y Cortisol. El estímulo de las catecolaminas para la lipolisis es muy potente y la acción inhibidora de la insulina muy fuerte. Además del aumento de las catecolaminas durante ejercicio, otros factores explican el aumento en la lipolisis, como son; El flujo sanguíneo en el tejido adiposo, la concentración de lactato, los niveles de insulina, y el flujo en la via glucolítica. Asi pues, la movilización de FFA almacenados en el tejido adiposo (y posiblemente su oxidación) se realiza principalmente bajo el control del Sistema Nervioso Simpático, pero otras hormonas y factores también le influyen, como por ejemplo la Insulina y la Hormona de Crecimiento. Durante el ejercicio físico (ver revisiones de Galbo etc) las catecolaminas plamáticas y la actividad simpática aumenta exponencialmente con la intensidad del ejercicio, aunque como se mencionó antes otros factores influyen también (flujo sanguíneo en el tejido adiposo, concentraciones plamáticas de lactato, concentraciones de insulina, aumentos en el flujo de la via glucolítica). 84 Centro Olímpico de Estudios Superiores INTERACCIÓN con CH En reposo parece claro que un aumento en las concentaciones plasmáticas de FA, llevará a una mayor oxidación de grasa, y que este aumento en de la b-oxidación, inhibirá la PDH y el aumento en la concentración de citato a una inhibición de la PFK, (llamado por algunos autores ciclo de Randle y/o ciclo glucosa/FA)ambas inhibiciones disminuirian la glucolisis y la glucogenolisis , es decir el uso de CH. En ejercicio, existe más controversia sobre estos mecanismos, y también se postula la regulación a través de la malonil-CoA, en la Carnitina Acil Transferasa. 2.4.6 EFECTO DE LA INTENSIDAD DE EJERCICIO, en el metabolismo graso. El factor principal que determina el grado de oxidación de grasa o de CH durante ejercicio, es la INTENSIDAD. La intensidad del ejercicio es el factor fundamental en la elección de grasas o H de C como sustrato durante el ejercicio. A intensidades de 25% del VO2max, casi toda la energía proviene de la grasa (sobre todo de los FA del plasma). Al 65% la grasa provee un 50% de la energía (mayor en cifras absolutas que al 25%), pero a estas intensidades son los depósitos intramusculares (de IMTG y glucógeno) los más importantes, aunque los FA del plasma siguen aportando energía. A intensidades mayores de ejercicio (i.e. 85% del VO2max ), la contribución de las grasas en proporción a los H de C es mucho menor, aunque algo todavía se utilizan. El hecho de que a intensidades altas de ejercicio, (además de la posible inhibición de la lipolisis debida al aumento en la concentración plasmática de lactato y el menor flujo en el tejdo adiposo) aunque haya presencia alta de FA (aporte suficiente) no se utilicen, hace pensar en factores inhibitorios intraceluraes, como que la glucolisis inhibe el transporte de FA a la mitocondria. Además a esas intensidades se reclutan más fibras FT, menos oxidativas y más glucolíticas, además puede haber una competición entre los AcetilCoA derivados del Piruvato con los derivados de FA, para entrar en el TCA-cycle. El uso de grasa puede aumentar marcadamente cuando los depósitos de glucógeno se han vaciado, pero la intensidad alta no se puede realizar y se tienen que bajar la intensidad, ya que la velocidad de producción de ATP con la grasa es mucho menor. El aumento en la concentración plasmática de catecolaminas va seguida de un aumento en la liberación de FFA al plasma, es decir un aumento en la Lipolisis. La lipolisis 85 Centro Olímpico de Estudios Superiores aumenta por aumentos en la actividad de la Lipasa de los adipocitos, esta lipasa esta regulada hormonalmente, fundamentalmente por las catecolaminas y la insulina. Las primeras, acelerando su actividad y la segunda inhibiendola. FIGURA 3: Esquema de la movilización y transporte de Acidos grasos libres (FFA) del tejido adiposo al músculo esquelético. En el adipocito la Norepinefrina (NE), activará a través del AMPc una Lipasa que hidrolizará los Triglicéridos (TG) en FFA y Glicerol (Glic). Los FFA y el Glic, pasan a la sangre y de ahí al músculo. En la sangre, los FFA se transportan ligados a la Albúmina (ALB). Cuando entran en el músculo (desligados de la ALB), pueden pasar directamente a la mitocondria para su oxidación o almacenarse en forma de gotitas lipídicas (GL). Los fármacos B-bloqueantes disminuyen la Lipolisis mientras que los B-agonistas la aumentan. La mayor o menor concentración de Lactato en sangre también influye en la Lipolisis, pues el aumento en sangre de Lactato tiene un efecto inhibitorio en la liberación de FFA, tal como se ha podido comprobar en estudios en los que sujetos que realizaban un ejercicio de intensidad baja con las piernas e intercalaban repeticiones cortas pero muy intensas de ejercicio con los brazos, constatandose un aumento de Lactato en plasma y una disminución en la liberación de FFA. 86 Centro Olímpico de Estudios Superiores Otras manipulaciones pueden alterar la movilización de las grasas, como la utilización de Heparina o de Cafeína. La inyección de Heparina en ratas aumenta la actividad de la Lipoproteina-Lipasa, por lo que aumenta la concentración de FFA en plasma. La ingestión, en humanos, de Cafeína eleva las concentraciones de FFA en plasma, posiblemente debido a su efecto inhibidor de la Fosfodiesterasa (enzima que degrada el AMP-cíclico) por lo que prolonga la presencia de este activador cuando se activa el sistema de la Adenilato Ciclasa, asociada con la degradación de los TG. En varios estudios se ha demostrado una mejora en el rendimiento aeróbico, con el uso de dosis bajas (no dopantes) de Cafeína. Los Acidos Grasos provenientes de la lipolisis en el tejido adiposo son muy hidrófobos, por lo que tienen que unirse a una proteína para ser transportados en un medio acuoso. En el plasma se unen a la Albúmina para ser transportados. En los demás tejidos se unen a proteínas especificas para su transporte intracelular (Fatty Acid Binding Proteins, FABPs). Cuando entran en la celular muscular, los Acidos Grasos pueden almacenarse como Trigliceridos o unirse a las FABPs en el citoplasma, donde estarán disponibles para su conversión en Acyl-CoA y ser transportados al interior de la mitocondria para su oxidación. Utilización durante el ejercicio. Hay dos aspectos del metabolismo de las grasas que son de gran importancia respecto al ejercicio físico, estos son: 1-La grasa sólo puede utilizarse como fuente energética en condiciones aeróbicas. 87 Centro Olímpico de Estudios Superiores 2-Debido al hecho de que en la célula muscular sólo se almacenan pequeñas cantidades de grasa, esta debe de ser transportada hasta el músculo e introducida en él. La contribución de la grasa como fuente energética durante el reposo y el ejercicio ha sido muy estudiada, fundamentalmente mediante la valoración del Cociente Respiratorio (RQ), por el hecho de que los CH producen diferentes cantidades de CO2 y requiren diferentes cantidades de O2 que las grasas, cuando se oxidan. Este parámetro tiene unos valores en reposo (en personas que ingieran una dieta mixta) entre 0.80 y 0.85. Lo que indica que la grasa está contribuyendo en un 50% a la producción total de energía. Sin embargo será de aproximadamente 0.69-0.73 cuando se oxida sólo grasa, y 1 cuando se oxida sólo glucosa. Estudios recientes usando isótopos radioactivos están precisando aún más estos conceptos. Durante el ejercicio físico la utilización mayor o menor de la grasa dependerá fundamentalmente de la intensidad y duración del ejercicio, haciéndolo en muchos casos de una manera inversa a los Hidratos de carbono (Figura 4 ), como ya se demostró en los años treinta. 88 Centro Olímpico de Estudios Superiores FIGURA 4: Contribución (en % aproximado) al metabolismo energético total, de las grasas (o) y los Hidratos de carbono (.), durante un ejercicio prolongado. Modificado de Edwards y col. 1934. Cuando el ejercicio es muy intenso la fuente energética principal son los Hidratos de carbono y la aportación de las grasas es pequeña. Conforme el ejercicio va siendo de menor intensidad y de mayor duración la contribución de las grasas es mayor. Así por ejemplo, durante ejercicio de baja intensidad y muy larga duración hay un progresivo aumento en la utilización de la grasa como fuente energética, llegando a ser en una persona no-entrenada de un 50% del total. Hay estudios científicos clásicos que muestran en ejercicios de muy larga duración que la contribución de la grasa a la producción total de energía puede ser de un 80% (incluso un 90% cuando la dieta de la persona es alta en grasa). La energía obtenida de la grasa proviene de la oxidación de los FFA, tal como se explicó anteriormente. Pero recientes estudios parecen indicar que los TG almacenados en el músculo pueden ser otra fuente energética en ejercicios de larga duración. No está claro si los TG que se utilizan son los de los adipocitos situados entre las fibras musculares o los de las gotitas lipidicas de las fibras o ambos. 89 Centro Olímpico de Estudios Superiores Durante ejercicio la movilización de FFA también está controlada fundamentalmente por el sistema nervioso Simpático con influencias de otras hormonas (Hormona de Crecimiento). Existe una gran correlación entre el consumo de FFA por el músculo y la concentración en el plasma. En cuanto a los TG que circulan en la sangre en forma de quilomicrones y lipoproteinas, la mayoría de los autores están de acuerdo en que no son utilizados directamente como fuente energética y en caso de serlo, lo serían en un porcentaje muy pequeño del total de los sustratos lipídicos utilizados. Como los depósitos de grasas son mucho mayores que los de Hidratos de Carbono, seria muy beneficioso para el rendimiento, el poder utilizar sobre todo grasa como fuente energética. Es decir, si a la misma intensidad de ejercicio se oxidaran sobre todo grasas en vez de H de C, seria energéticamente muy económico. Pero no es asi y su oxidación está limitada. Esta limitación puede estar a diferentes niveles: 1. En la movilización de la grasa del tejido adiposo. 2. En el transporte de FA al músculo 3. En la entrada a la célula muscular. -de FA del plasma -de FA de las lipoproteinas circulantes. 4. En la movilización de FA del pool de IMTG 5. En el transporte de FA hacia dentro de la mitocondria. 6. En la oxidación en la mitocondria. 90 Centro Olímpico de Estudios Superiores LOS DOS PASOS QUE POSIBLEMENTE LIMITEN MÁS EL USO (OXIDACIÓN) DE GRASA SON EL 1 Y EL 5, JUN TO CON LA DENSIDAD MITOCONDRIAL Y LA CAPACIADAD OXIDATIVA DE ESAS MITOCONDRIAS. La intensidad del ejercicio es el factor fundamental en la elección de grasas o H de C como sustrato durante el ejercicio. A intensidades de 25% del VO2max, casi toda la energía proviene de la grasa (sobre todo de los FA del plasma). Al 65% la grasa provee un 50% de la energía, pero a estas intensidades son los depósitos intramusculares (de IMTG y glucógeno) los más importantes, aunque los FA del plasma siguen aportando energía. A intensidades mayores de ejercicio (i.e. 85% del VO2max ), la contribución de las grasas en proporción a los HdeC es mucho menor, aunque algo todavía se utilizan. El hecho de que a intensidades altas de ejercicio, aunque haya presencia alta de FA (aporte suficiente) no se utilicen, hace pensar en factores inhibitorios, como que la glucolisis inhibe el transporte de FA a la mitocondria. Además a esas intensidades se reclutan más fibras FT, menos oxidativas y más glucolíticas, además puede haber una competición entre los AcetilCoA derivados del Piruvato con los derivados de FA, para entrar en el TCA-cycle. El uso de grasa puede aumentar marcadamente cuando los depósitos de glucógeno se han vaciado, pero la intensidad alta no se puede realizar y se tienen que bajar la intensidad, ya que la velocidad de producción de ATP con la grasa es mucho menor. 2.4.7 Dieta SUPLEMENTACION DE GRASA Tal como se mencionó en el apartado relativo a los Hidratos de Carbono, cambios en la dieta pueden inducir cambios en la elección de la fuente energética. Así por ejemplo, cuando se consume una dieta muy rica en grasas y baja en hidratos de carbono, el Cociente Respiratorio en reposo es muy bajo y permanece bajo durante el ejercicio, indicando un mayor uso de grasa como fuente energética y un ahorro de glucógeno. Aunque el aumento de grasa en la dieta tiene el riesgo de aumentar el compartimento graso y el peso corporal. Triglicéridos de cadena larga y Ejercicio. La ingestión aumentada de Triglicéridos de cadena larga (LCTG), durante ejercicio, no parece recomendable por las siguientes razones; Enlentecen el vaciado gástrico, pasan muy lentamente a la circulación (3-5 horas), entran en la sangre en forma de 91 Centro Olímpico de Estudios Superiores quilomicrones que no parece que sirvan como fuente energética inmediata, pero que podrían servir para rellenar los depósitos de IMTG despues del ejercicio. Triglicéridos de cadena media y Ejercicio. Al contrario que los LCTF los TG de cadena corta y media (MCTG) son absorbidos por el intestino y no son transportados como quilomocrones sino que entran directamente a la sangre a traves de la vena porta. Por lo que entran en la sangre muy rápidamente. Por ello, se ha sugerido que los MCTG pueden ser una fuente exógena de energía importante y rápida durante el ejercicio, junto con los CH, y que podrían aumentar el rendimiento físico al aumnentar los niveles de FA en plasma ahorrando glucógeno (ya que se cree que un aumento en la disponibilidad de FA plasmáticos reducirá la glucogenolisis y retrasará la fatiga. (Vukovich MD, Costill DL, Hickey MS, Trappe SW, Cole KJ and Fink WJ. Effects of fat emulsion infusion and and fat feeding on muscle glycogen utilization during cycle exercise. J Appl Physiol, 75: 1513-1518, 1993). Sin embargo, la cantidad máxima que se puede ingerir de MCTG, sin tener transtornos gastrointestinales es relativamente pequeña (unos 30gr - 10g/h), lo cual limita la posible contribución al gasto energético total de los MCTG, a valores entre un 3 y un 8% (según autores). Así pues; MCTG se vacian del estómago, se absorben rápidamente, su oxidación es rápida y se ve potenciada por la co- ingestión de CH, pero la contribución de cantidades cercanas a 30 g contribuyen sólo en un 3-8% al metabolismo energético. La ingestión de cantidades mayores de MCTG durante el ejercicio, puede producir trastornos gastrointestinales. Por todo ello no parece que tengan el efecto positivo en el rendimiento que postulan algunas compañias farmaceuticas-nutricionistas. Dietas muy altas en grasa. Varios estudios han demostrado que el consumo de una dieta muy alta en grasa, durante un largo período de tiempo reduce el uso de Carbohidratos como fuente energética, aumentando la contribución de la grasa. Esto ocurre sin una disminución del rendimiento físico en ejercicios de moderada o baja intensidad, apreciándose incluso un ahorro de glucógeno, siempre que el ejercicio sea de baja o moderada intensidad. Estos datos indican que hay en el músculo una adaptación en respuesta a un consumo de una dieta muy alta en grasa. Esta adaptación parece ser una mejora en la capacidad de utilizar la grasa durante ejercicios prolongados de baja o moderada intensidad. Pero suelen producir una disminución en los depósitos de glucógeno muscular lo cual puede ser la causa de un menor rendimiento. Cuando estas dietas se utilizan durante un periodo muy prolongado de tiempo, si bien podrían producir una adaptación mayor al uso de grasa, pueden producir bajadas en el sistema inmune, alteraciones negativas en el perfil lipídico (factores de riesgo cardiovascular) y en resistencia a la insulina en hígado, 92 Centro Olímpico de Estudios Superiores aumento de panículo adiposo y su efecto en el rendimiento no está claro. Sin embargo un reciente estudio de Van Zyl y col. Eur J. Appl. Physiol., 1998, se observa la posibilidad de que en la preparación de deportistas de ultra-resistencia, se beneficiarían de una periodización nutricional consistente en : Dieta rica en CH durante la mayoria del año, pero 2 semanas antes de la ultra-competición dieta muy alta en Grasa durante 710 dias, seguida de 2-3 dias de dieta de recarga de CH. Con esta manipulación, asociada a la ingesta de CH+MCTG durante la competición, estos autores observaron mejoras significativas en el rendimiento en competiciones de larga duración. 2.4.8 Entrenamiento Desde hace muchos años se conoce que después de períodos largos de entrenamiento físico, aumenta la utilización de la grasa como fuente energética. Los estudios clásicos mostraban la diferencia entre los RQ de personas entrenadas y no entrenadas, así como la disminución en los valores de este parámetro después de períodos de entrenamiento, indicando un cambio en la elección del substrato energético, con una mayor utilización de las grasas. Este cambio en la elección del substrato, es importante por dos aspectos: Primero, al ser la grasa la reserva de energía mas grande que -en condiciones normales- se tiene en el organismo, es muy "lógico" por parte de nuestro cuerpo intentar utilizarla mas, pues además es el substrato que mas fácilmente se almacena en nuestro cuerpo. Segundo, la utilización de glucógeno está limitada, pues los depósitos se vacían relativamente pronto y tardan 24-48 horas en rellenarse. Teniendo en cuenta que el punto de agotamiento está relacionado con el vaciamiento de estos depósitos, será de mucha importancia el hecho de que se usen otras fuentes energéticas lo mas posible. Entre las muchas adaptaciones que se producen en el organismo de una persona que realiza entrenamiento físico de resistencia, dos son fundamentales para que se produzca la mayor utilización de grasa que se comentó con anterioridad, estas son: a) Aumento en el numero de mitocondrias en la célula muscular, con el consiguiente aumento en la concentración de enzimas oxidativas y en la capacidad metabólica oxidativa del músculo. Este aumento permite al músculo adaptarse mas y mejor a una mayor demanda energética, no sólo por poder oxidar mas grasa sino también por tener aumentado el potencial de transporte de FFA de fuera a dentro de la mitocondria a través del sistema de la Carnitina-Palmitil- Transferasa situado en la pared mitocondrial. 93 Centro Olímpico de Estudios Superiores b) Aumento en la capilarización del músculo, pues al ser mayor la superficie de contacto entre el lecho capilar y la célula muscular, será mayor la posibilidad de paso de sustratos energéticos (oxígeno y FFA). Al ser mayor el lecho capilar también aumentará el tiempo de paso de la sangre a través de los capilares del músculo, con lo que aumenta el tiempo en el que los FFA y el Oxígeno están en contacto con la membrana capilar y puedan pasar al músculo. Además contribuyen: Aumento en la oxidación de IMTG, aumento en la entrada de FA y cambios en la movilización de FA desde el tejido adiposo. El aumento en la concentración mitocondrial que se produce en el músculo, debida al entrenamiento de resistencia, conlleva, tal como se mencionó con anterioridad, una mayor capacidad para oxidar no sólo FA sino también carbohidratos (a partir del Piruvato), por lo que surge la duda de ¿porqué se oxida relativamente mas grasa que carbohidratos en el músculo de una persona entrenada que en el de un sedentario ?. La respuesta es que la oxidación de los FFA inhibe, al menos parcialmente, la captación de glucosa por el músculo, la glucolisis, la glucogenolisis y la oxidación del Piruvato. Este efecto inhibitorio ya fue descrito en el músculo cardíaco en 1963 por investigadores dirigidos por Newsholme y confirmado en el músculo esquelético por Rennie y Holloszy en 1977. En las estancias en altitud, debido al hecho de que la hipoxia aumenta las concentraciones plasmáticas de catecolaminas, se observa un aumento en la lipolisis del tejido adiposo y una mayor concentración (¿ y utilización?) de FFA en la sangre de los deportistas. Tal como se verá con mas detalle en el Módulo dedicado al Entrenamiento en Altitud. Es importante destacar que estudios recientes (ver revisión de Jeukendrup y col. 1998), indican que despues de un periodo de entrenamiento, aumentan los IMTG y su utilización (su oxidación) durante ejercicio. La producción de Cuerpos Cetónicos durante el ejercicio ya fue observada por fisiólogos del ejercicio hace muchos años , sobre todo en personas sedentarias y en las que tenían una dieta rica en Grasa y pobre en Hidratos de carbono. Por lo que se relacionó su producción y su aclaramiento no sólo con la dieta, el ayuno y la descompensación diabética, sino también con el nivel de entrenamiento. En el año 1969, Johnson y sus colaboradores demostraron claramente, que en deportistas entrenados, no aumentaba la concentración de Cuerpos Cetónicos en su sangre cuando realizaban un ejercicio de larga duración, mientras que en personas sedentarias que realizaban el 94 Centro Olímpico de Estudios Superiores mismo ejercicio, sus concentraciones en sangre de Cuerpos Cetónicos aumentaban en un 50%. Esto se interpretó como señal de que en los deportistas entrenados, había una mayor limpieza de Cuerpos Cetónicos que en personas sedentarias. Posteriormente se ha comprobado que el músculo entrenado tiene aumentada la capacidad de oxidar Cuerpos Cetónicos, por lo que podemos concluir que el entrenamiento induce también unas adaptaciones que permiten al músculo utilizar mas Cuerpos Cetónicos durante el ejercicio. Así pues con el entrenamiento de resistencia se producen una serie de adaptaciones (fundamentalmente el aumento en la concentración de mitocondrias y enzimas oxidativas y el aumento en densidad capilar en el músculo) que contribuyen a un mayor y mejor uso de la Grasa como fuente energética durante el ejercicio. Lo cual reducirá el uso de Glucógeno y aumentará el resistencia física. 2.5 METABOLISMO DE LAS PROTEÍNAS 2.5.1 Características Las proteínas están compuestas de aminoácidos, que están unidos formando largas cadenas. A diferencia de las Grasas y de los Hidratos de Carbono, las Proteínas tienen en su composición (además de Carbono, Oxígeno e Hidrógeno) Nitrógeno, lo cual les confiere unas características metabólicas y plásticas muy diferentes. Las Proteínas ingeridas en la dieta son hidrolizadas completamente en el estómago y en el intestino delgado, hasta quedar en forma de aminoácidos, que son absorbidos y pasan a la circulación. Las Proteínas son un substrato imprescindible para los procesos anabólicos o de formación del organismo, aunque en este capítulo sólo nos ocuparemos de su papel como substrato energético en el metabolismo humano. 95 Centro Olímpico de Estudios Superiores Hay que recordar que los aminoácidos se suelen clasificar en ; No esenciales y en Esenciales , según si pueden o no ser sintetizados por el organismo (los aminoácidos Esenciales al no poder ser sintetizados por el organismo es "esencial" que sean aportados por la dieta). Los aminoácidos Esenciales son: Fenilalanina, Isoleucina, Leucina, Lisina, Metionina, Treonina, Triptófano y Valina. Los No esenciales los restantes; Alanina, Ácido aspártico, Ácido glutámico, Arginina, Cisteína, Cistina, Citrulina, Glicina, Hidroxilina, Hidroxiprolina, Histidina, Prolina, Serina y Tirosina. 2.5.2 Fuentes Las principales fuentes de obtención de Proteínas para el hombre están en las carnes, pescados, huevos y la leche y sus derivados. Además son fuente de Proteínas; las legumbres, los cereales, los frutos secos y las semillas comestibles. Algunos autores las clasifican en; proteínas completas y proteínas incompletas. Las completas serían las que contienen en su composición los ocho aminoácidos esenciales y son fundamentalmente las de origen animal. Las incompletas son las que no contienen los aminoácidos esenciales, o lo hacen en poca cantidad. Estas son fundamentalmente las derivadas de los vegetales. 2.5.3 Almacenamiento Las Proteínas no se pueden almacenar en el organismo, salvo formando parte de estructuras, como por ejemplo el músculo, y en esas estructuras se están reciclando continuamente (el músculo se renueva en unos 160 días aproximadamente), por lo que hay que estar aportando sin cesar unas cantidades mínimas de Proteína al cuerpo. Cuando el aporte es excesivo, las Proteínas no se almacenan en el organismo como tales Proteínas, sino que se convierten en Grasa, en Glucosa o se eliminan. Como mencionamos anteriormente, las Proteínas tienen una función plástica formando parte de tejidos orgánicos, por lo que son muy importantes para el crecimiento físico y para el desarrollo de la masa corporal y tienen una función limitada como reserva 96 Centro Olímpico de Estudios Superiores energética en relación con el ejercicio físico. Sin embargo existe una gran relación entre el ejercicio físico y una correcta utilización de las Proteínas en el crecimiento corporal y en la formación de músculo. En estudios realizados con niños en período de crecimiento se ha demostrado que crecían más y mejor los niños que tenían una actividad física moderada comparados con los que tenían una vida más sedentaria, demostrando que el ejercicio físico moderado tiene un efecto positivo en el crecimiento ya que favorece una más correcta utilización de las proteínas aportadas por la dieta. En adultos también se ha observado este efecto, en el aumento de la masa corporal y en aumento de tamaño de ciertos músculos. Así pues, podemos resumir que el almacenamiento de las Proteínas en nuestro organismo es fundamentalmente formando parte del músculo, por lo que su contribución como fuente energé tica durante el ejercicio físico es muy limitada, ya que se necesitaría la ruptura de la estructura y de la funcionalidad de este importante tejido orgánico. Aunque, como veremos más adelante, los aminoácidos contenidos en las Proteínas musculares, pueden ser movilizados en situaciones especiales para ayudar en la Gluconeogénesis hepática (Figura 5), cuando los depósitos de Grasas y de Hidratos de Carbono se han acabado o se ha impedido su movilización. 2.5.4 Utilización de las Proteínas durante ejercicio. En general la mayoría de los fisiólogos opinan que la contribución de las Proteínas al metabolismo energético durante el ejercicio físico es limitada y puede oscilar entre un 1% y un 15% del total de la energía utilizada. Muchos han sido los intentos y los métodos para cuantificar el uso de las Proteínas como fuente energética, de estos métodos dos han sido los más usados: a)La valoración de la producción de Urea y b)La cuantificación del uso de determinados aminoácidos. De todos los estudios realizados sobre la utilización de las Proteínas durante el ejercicio, tres conceptos parecen claros: Primero; durante el ejercicio aumenta la oxidación en el músculo activo (con la consiguiente producción de energía) de Leucina y probablemente también de otros 97 Centro Olímpico de Estudios Superiores aminoácidos de cadena ramificada. Producto de esta utilización es el aumento en la eliminación de Urea que se produce después del ejercicio. Segundo; los aminoácidos pueden actuar en caso necesario como precursores en la Gluconeogénesis hepática, principalmente la Alanina que libera el músculo en ejercicio. Tercero; el porcentaje de energía que se puede obtener de las Proteínas durante el ejercicio físico, es pequeño. Hay que recordar también, que la Alanina actua como transportador no tóxico de aminas nitrogenadas del músculo al hígado, donde se formará Urea (ver Figura 5). 2.5.5 Efecto del ejercicio físico en la síntesis y degradación de Proteínas. Tal como se indicó en el apartado anterior, el ejercicio físico puede inducir la ruptura y utilización de Proteínas, que externamente se aprecia por el aumento en la eliminación de Urea. Sin embargo no es claro que el ejercicio produzca un efecto catabólico en el organismo, ya que en muchos casos el ejercicio produce un efecto anabólico, con un aumento en la formación de Proteínas en el músculo, lo cual pone en duda si la degradación de Proteínas observada indirectamente por el aumento en la eliminación de Urea, se produce en el músculo o en otro órgano. La dificultad metodológica que hay para estudiar con exactitud estos problemas, hace que todavía varios de ellos estén sin resolver. En los estudios en los que se valoró la síntesis de Proteínas durante ejercicio físico, en animales y en hombres, se encontraron resultados contradictorios, pues en algunos de ellos se apreciaba una disminución en la síntesis de Proteínas, mientras que en otros la síntesis estaba aumentada. Cuando se valoró la síntesis de Proteínas en el período posterior al ejercicio, la mayoría de los estudios encontraron un aumento. La variedad de modelos de ejercicio 98 Centro Olímpico de Estudios Superiores (intensidad, duración, tipo de ejercicio, tipo de estímulo, momentos de medida durante y después del ejercicio etc.) utilizados en estos experimentos hace imposible por ahora sacar unas conclusiones definitivas, aunque muchos autores coinciden en afirmar que en el período post-ejercicio hay un aumento en la síntesis de Proteínas, inducida por el propio ejercicio. La degradación proteínica durante el ejercicio, también ha sido muy estudiada y con resultados muy contradictorios, debido a los métodos de medida. En ratas parece claro que con el ejercicio aumenta la degradación de proteínas en el hígado, lo cual sería consistente con un aumento en la disponibilidad de aminoácidos para ser oxidados o utilizados en la gluconeogénesis. En resumen podríamos decir que la actividad física moderada tiene un efecto positivo en el crecimiento y desarrollo de la masa corporal, tanto en niños como en adultos. El ejercicio físico tiene un efecto beneficioso especialmente para la síntesis de las proteínas del músculo. Durante el ejercicio físico se producen cambios en la síntesis y en la degradación de proteínas (siendo esta degradación utilizada para la obtención de energía), que pueden continuar más o menos tiempo después de acabado el ejercicio. Las causas y mecanismos que regulan estos procesos están todavía sin definir totalmente. 99 Centro Olímpico de Estudios Superiores FIGURA 5: Representación esquemática del ciclo Glucosa-Alanina. 2.5.6 Dieta. Durante muchos años, el hecho demostrado de que las Proteínas eran una fuente de energía de poca importancia, comparadas con las Grasas y los Hidratos de Carbono, hizo que las recomendaciones dietéticas para los deportistas, enfatizaran en que las cantidades por kilo de peso y día de Proteínas (0.8 gr/kilo de peso/día), y los porcentajes de Proteinas en la dieta diaria (10-15% del total de la ingesta calórica), fueran muy bajos. Debido a los estudios que han demostrado que, durante los ejercicios de moderada o baja intensidad y de varias horas de duración, se aumentaba claramente la ut ilización y oxidación de Leucina (y posiblemente de otros aminoácidos) las recomendaciones dietéticas se están modificando, sobre todo para deportistas que realizen ejercicios y/o entrenamientos que duren varias horas y para niños en edad de crecimiento que realicen 100 Centro Olímpico de Estudios Superiores entrenamientos físicos de larga duración. En estos casos parece más aconsejable recomendar que el consumo de Proteínas sea del orden de 1.0 a 1.5 gr/kilo de peso/día. Ni siquiera en los casos de deportistas que realizan entrenamientos de fuerza y musculación, se deben de aumentar mucho las cantidades de Proteínas mencionadas anteriormente, sin pasar en ningún caso de 2.0 gr/kilo de peso/día. 101 Centro Olímpico de Estudios Superiores PREGUNTAS DE AUTOCONTROL DE LA ASIMILACION 1 Establece las diferencias entre la Glicolisis y la Fosforilación Oxidativa, y elabora un cuadro comparativo entre las características de una y otra. 2 Los Principios inmediatos tienen rasgos diferentes. A partir de los siguientes criterios; fuente, almacenamiento, utilización en el ejercicio, y alguno más que consideres oportuno, establece un cuadro comparativo. 102 Centro Olímpico de Estudios Superiores BIBLIOGRAFIA Ahlborg,G.,Felig,P.,Hagenfeld,L.,Hendler,R.y Wahren,J.1974.Subtrate turnover during prolonged exercise in man:Splachnic and leg metabolism of glucose , free fatty acids and amino acids. J.Clin.Invest.53:1080-1098. Andersen,P. y Saltin,B.1985.Maximal perfusion of skeletal muscle in man. J.Physiol. (London).366:233-249. Astrand,P-O.y Rodahl,K.1986. Textbook of Work Physiology.3 ed.New York:McgrawHill. Balaban,R.1990.Regularion of oxidative phosphorylation in the mammalian cell. AM.J.Physiol.(Cell Physiol.).258(27):C377-C389. Bang,O.1936.The lactate content of the blood during and after exercise in man. Skand.Arch.Physiol.74:51-82. Bergstrom,J.,Hermansen,L.,Hultman,E.y Saltin,B.1967.Diet,muscle glycogen and physical performance. Acta Physiol.Scand.71:140-150. Booth,F.W.y Watson,P.A.1985.Control of adaptations in protein levels in response to exercise. Fed.Proc.44:2293-2300. Cathcart,E.P.1925.Influence of muscle work on protein metabolism. Physiol. Rev. 5:225-243. 103 Centro Olímpico de Estudios Superiores Christensen,E.H.y Hansen,O.1939.Arbeitsfahigkeit und Ehrnahrung. Skand.Arch.Physiol.81:160-175. Cori,C.F.1926.The fate of sugar in the animal body.III.The rate of glycogen formation in the liver of normal and insulinized rats during the absortion of glucose,fructose and galactose. J.Biol.Chem.70:577-585. Costill,D.L.,Coyle,E.,Dalsky,G.,Evans,W.,Fink,W.y Hoopes,D.1977.Effects of elevated plasma FFA and insulin on muscle glycogen usage during exercise. J.Appl.Physiol.43:695-699. Costill,D.L.,Dalsky,G.y Fink,W.1978.Effects of caffeine ingestion on metabolism and exercise performance. Med.Sci.Sports Exerc.10:155-158. Dill,D.B.,Edwards,H.T.y Talbott,J.T.1932.Studies in muscular activity.VII.Factors limiting the capacity of work. J.Physiol.77:49-62. Dohm,G.L.,Kasperek,G.J.,Tapscott,E.B.y Barakat,H.A.1985.Protein metabolism during endurance exercise.Fed.Proc.44:348-352. Gaesser,G.A.y Brooks,G.A.1980.Glycogen repletion following continuous and intermittent exercise to exhaustion. J.Appl.Physiol.49:722-728. Gollnick,P.D.,Piehl,K.,Saubert,C.W.,Armstrong,R.B.y Saltin,B.1972.Diet,exercise and glycogen changes in human muscle fibers.J.Appl.Physiol.33:421-425. Goodman,M.N.y Ruderman,N.B.1982.Influence of muscle use on amino acid metabolism.En:R.L.Terjung,eds.Exercise and Sport Science Reviews. Philadelphia: Franklin Institute Press. 104 Centro Olímpico de Estudios Superiores Hatefi,Y.1985.The mitochondrial electron transport and oxidative phosphorylation system.Annu.Rev.Biochem.54:1015-1069. Hermansen,L.y Stensvold,I.1972.Production and removal of lactate during exercise in man.Acta Physiol.Scand.86:191-201. Hilty,M.,Groth,H.,Moore,R.y Musch,T. 1989.Determinants of VO2max in rats after high- intensity sprint training. J.Appl.Physiol.66:195-201. Hochachka,P. 1991.The biochemical limits of muscle work. En:A.W.Taylor,ed. Biochemistry of Exercise VII. Champain,IL.Human Kinetics. Hogan,M.,Bebout,D.y Wanger,P. 1991.Effects of hemoglobin concentration on maximal O2 uptake in canine gastrocnemius muscle in situ. J.Appl.Physiol. 70:11051112. Hoerr,R.A.,Young,V.R.y Evans,W.J.1982.Protein metabolism and exercise. En:P.L.White y T.Mondeika,eds. Diet and Exercise: Synergism in Health Maintenance.Chicago:American Medical Association. Holloszy,J.O. y Booth,F.W.1976.Biochemical adaptations to endurance exercise in muscle.Annu.Rev.Physiol.38:273-291. Hoppeler,H. 1990.The different relationship of VO2max to muscle mitochondria in humans and quadrupedal animals. Resp.Physiol.69:27-46. Jansson,E.1980.Diet and muscle metabolism in man. Acta Physiol. Scand. (Suppl.) 487. 105 Centro Olímpico de Estudios Superiores Jansson,E.1982.On the significance of the respiratory exchange ratio after different diets during exercise in man.Acta Physiol.Scand.114:103-110. Jansson,E.y Kaijser,L. 1987.Substrate utilization and enzymes in skeletal muscle of extremely endurance-trained men. J.Appl.Physiol.62:999-1005. Jeukendrup A.E., Saris W.H.M. and A.J.M. Wagenmakers. Fat metabolism during exercise: A review-Part I: Fatty acid mobilization and muscle metabolism. Int J. Sports Med., Vol.19, pp.231-244, 1998. Jeukendrup A.E., Saris W.H.M. and A.J.M. Wagenmakers. Fat metabolism during exercise: A review-Part II: Regulation of metabolism and the effects of training. Int J. Sports Med., Vol.19, pp.293-302, 1998. Jeukendrup A.E., Saris W.H.M. and A.J.M. Wagenmakers. Fat metabolism during exercise: A review-Part III: Effects of nutritional interventions. Int J. Sports Med., Vol.19, pp.371-379, 1998. Kiens,B.y Terrados,N.1985.Coupling between work performance and metabolic adaptation in a single muscle group in man. Clin.Physiol.5.Suppl.4.(Abstract). Koeslag,J.H.1982.Post-exercise ketosis and the hormone response to exercise:A review. Med.Sci.Sports Exerc.14:327-334. Krogh,A.y Lindhard,J.1920.The relative value of fat and carbohydrate as sources of muscular energy. Biochem.J.14:290-336. Kushmerick,M.J.1983.Energetics of muscle contractions. En: L.D.Peachy, R.H.Adrian y S.R.Geiger,eds. Handbook of Physiology. Sec 10:Skeletal Muscle. Bethesda,Md:American Physiological Society. 106 Centro Olímpico de Estudios Superiores Lipmann,F.1941.Metabolic generation and utilization of phosphate bond energy. Adv.Enzymol.1:99-162. Mitchell.J. 1990.Neural control of the circulation during exercise. Med.Sci.Sports Exerc.22:141-154. Nilsson,L.H.y Hultman,E.1973.Carbohydrate metabolism of the liver in normal man under varying dietary conditions. Scand.J.Clin.Lab.Invest.32:331-337. Sahlin,K.1978.Intracellular pH and energy metabolism in muscle of man. Acta Physiol.Scand.(Suppl).455:1-56. Saltin,B. 1985a.Hemodinamics adaptations to exercise. Am.J.Cardiol.55:42D-47D. Saltin,B. 1985b.Malleability of the sistem in overcoming limitations:functional elements. J.Exp.Biol.115:345-354. Saltin,B.1986.Physiological adaptations to physical conditioning:Old problems revisited. Acta Med.Scand. (Suppl.).711:11-24. Saltin,B.y Astrand,P.-O. 1967.Maximal oxigen uptake in athletes. J.Appl.Physiol. 23:353-358. Saltin,B.y Gollnick,P.D.1983.Skeletal muscle adaptability: Significance for metabolism and performance. En:L.D.Peachy,R.H.Adrian y S.R.Geiger,eds. Handbook of Physiology.Sec.10:Skeletal Muscle. Bethesda,Md.:American Physiological Society. Saltin,B.,Hartley,L.H.,Kilbom,A.y Astrand,I.1969.Physical training in seddentary middle-age men.II.Oxygen uptake,heart rate and blood lactate concentration at submaximal and maximal axercise. Scand.J.Clin.Lab.Invest.1969.24:323-334. 107 Centro Olímpico de Estudios Superiores Saltin,B.,Kiens,B.y Savard,G.1986.A quantitative approach to the evaluation of skeletal muscle substrate utilization in prolonged exercise. En: B.Benzi, L.Packer y N.Siliprandi,eds. Biochemical Aspects o fPhysical Exercise. Amsterdam: Elsevier Science Publishers. Segal,S.S.,Damon,D.N.y Duling,B.R. 1989.Propagation of vasomotor responses coordinates arteriolar resistences. Am.J.Physiol.(Heart Circ.Physiol.). 256(25): H832H837. Sweene y,T.E.y Sarelius,I. 1990.Spatial heterogeneity in striated muscle arteriolar tone, cell flow, and capillarity. Am.J.Physiol.259:H124-H136. Terrados,N.1988.Bases medicas y fisiologicas del calentamiento y el estiramiento como prevencion de lesiones deportivas. Med.Ejerc.(Uruguay). vol.2:23-26. Terrados,N.,Jansson,E.,Sylven,C.y Kaijser,L.1990.Is hypoxia a stimulus for synthesis of oxidative enzymes and myoglobin?. J.Appl.Physiol.68:2369-2372. Terrados,N.y Marin,B.1985.Lipoproteinas de alta densidad y ejercicio físico. Apunts Med.Dep.85:7-17. Terrados,N.,Melichna,J.,Jansson,E.y Kaijser,L. 1985.Entrenamiento en altitud.Su efecto en el rendimiento deportivo y en los enzimas musculares. Arch.Med.Dep. Vol.II.8:303309. Terrados,N.,Melichna,J.,Jansson,E.y Kaijser,L.1988.Effects of training at simulated altitude in performance and muscle metabolic capacity. Eur.J.Appl.Physiol. 57:203-209. Terrados,N.,Melichna,J.,Sylven,J.y Jansson,E. 1986.Decrease in muscle myoglobin with intensive training in man. Acta Physiol.Scand.128:651-652. 108 Centro Olímpico de Estudios Superiores Terrados,N.,Mizuno,M.y Andersen,H.1985a.Reduction in maximal oxigen uptake at low altitudes;role of training status and lung function. Clin.Physiol. 5.Suppl.3:75-79. Terrados,N.,Mizuno,M.y Andersen,H.1985b.Efecto de altitudes moderadas (900,1200 y 1500 m. sobre el nivel del mar) en el consumo maximo de oxigeno. Apunts Med.Dep.86:97-103. Von Pettenkoffer,M.y Voit,C.1866.Untersuchungen uber den Stoffverbrauch des normalen Menschen. Z.Biol.2:459-573. Wittenberg,B.y Wittenberg,J. 1989.Transport of oxygen in muscle. Ann.Rev.Physiol. 51:857-878. 109 Centro Olímpico de Estudios Superiores