Fecha: 15 de Junio de 2012 Nombre: Dra. Llanos Belmonte Andújar
Anuncio

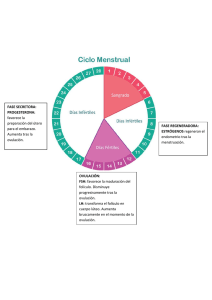
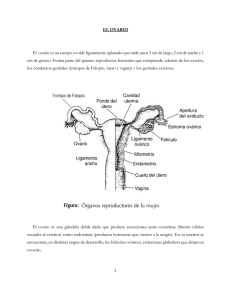
Fecha: 15 de Junio de 2012 Nombre: Dra. Llanos Belmonte Andújar R-4 Tipo de Sesión: Seminario HISTOLOGÍA DEL APARATO GENITAL FEMENINO 1. VAGINA. La vagina es un canal fibromuscular que consta de un epitelio escamoso estratificado, una capa de músculo liso y una capa externa adventicial. En estado de relajación, la pared de la vagina se colapsa para obliterar la luz y el epitelio se pliega. La densa lámina propia contiene muchas fibras elásticas, un rico plexo vascular de pequeñas venas y no posee glándulas. El moco cervical, procedente de la abundante red vascular de la lámina propia, lubrica la vagina junto al secretado por las glándulas de los labios menores de la vulva. Los haces de músculo liso de la capa muscular se disponen en una capa circular interna y una longitudinal externa mal definidas. La capa adventicial de la vagina no está recubierta por mesotelio, sino que se fusiona con las capas adventiciales de la vejiga por delante y del recto por detrás. La combinación de una capa muscular y una lámina propia muy elástica permite la gran distensión que se produce durante el parto. A la inversa, tras el coito, la contracción involuntaria de la capa de músculo liso asegura que una cantidad de semen permanezca en la región del cuello. Las gruesas fibras elásticas de la capa adventicial externa contribuyen también a estas funciones. Durante el ciclo menstrual, el epitelio escamoso estratificado de la vagina sufre cambios cíclicos de sus concentraciones de glucógeno. Durante todo el ciclo, las células superficiales producen glucógeno, que es metabolizado de forma anaeróbica por las bacterias comensales de la vagina para formar ácido láctico e inhibir así el crecimiento de los microorganismos patógenos. 2. ÚTERO El útero es un órgano con forma de pera aplanada que mide alrededor de 7 cm de longitud en estado de reposo. Su revestimiento mucoso, el endometrio, proporciona el entorno adecuado para el desarrollo del feto; la gruesa pared de músculo liso, el miometrio, se expande enormemente durante el embarazo, proporcionando protección al feto y un mecanismo para su expulsión en el momento del parto. ENDOMETRIO El revestimiento endometrial de la cavidad uterina consiste en un epitelio cilíndrico seudoestratificado ciliado sostenido por un estroma celular que aloja las numerosas glándulas tubulares simples. Bajo la influencia de los estrógenos y la progesterona secretados por los folículos durante el ciclo ovárico, el endometrio sufre cambios cíclicos regulares destinados a ofrecer un entorno adecuado para la implantación del óvulo fertilizado. Para que la implantación tenga éxito, el óvulo fecundado necesita de un tejido fácilmente penetrable y muy vascularizado, con un suministro abundante de glucógeno para su nutrición hasta que puedan establecerse conexiones vasculares con el medio ambiente materno. El ciclo de los cambios endometriales consta de dos fases distintas, proliferación y secreción; estos cambios afectan tanto al epitelio como al estroma de sostén. • Fase proliferativa: el estroma endometrial prolifera, haciéndose más grueso y ricamente vascularizado. Las glándulas tubulares simples (G) proliferan y forman numerosas glándulas de aspecto contorneado al final de esta fase, que comienzan a secretar coincidiendo con la ovulación. La fase proliferativa se inicia y mantiene hasta la ovulación gracias a la creciente producción de estrógenos por los folículos ováricos en desarrollo. • Fase secretora: la liberación se progesterona por el cuerpo amarillo después de la ovulación estimula la producción de una copiosa secreción rica en glucógeno en las glándulas endometriales (vacuolicación) (V). A menos que se produzca la implantación del óvulo fecundado, la producción continua de progesterona resulta inhibida por la hipófisis anterior gracias a un mecanismo de retroalimentación negativa, con supresión de la liberación de LH y consiguiente involución del cuerpo amarillo. En ausencia de progesterona, el endometrio no puede mantenerse, y durante el periodo de hemorragia conocido como menstruación, se descama en su mayor parte. La reactivación de la secreción de FSH inicia un nuevo ciclo de desarrollo folicular y de secreción de estrógenos; a su vez, ello inicia un nuevo ciclo de proliferación del endometrio a partir de los restos que quedan del ciclo anterior. Aunque el proceso de la menstruación representa el punto y final del ciclo de cambios endometriales, el primer día de ella también marca el comienzo de una nueva fase de proliferación. Además, como momento más fácilmente identificlable suele ser el que se toma para señalar el primer día del ciclo menstrual de 28 días. La duración media de la menstruación es de 5 días; la fase proliferativa continúa hasta el día 14, momento en el que se produce la ovulación y se inicia la fase secretora. Esta última culmina al comienzo de la menstruación, es decir, hacia el día 28. El endometrio se divide en tres capas histológica y funcionalmente distintas. La más profunda o basal, el estrato basal (B) adyacente al miometrio, apenas sufre cambios durante el ciclo menstrual y no se descama con la menstruación. La ancha capa intermedia se caracteriza por un estroma de aspecto esponjoso, por lo que se llama estrato esponjoso (E). La capa superficial, la más delgada, tiene un estroma de aspecto compacto, por lo que se conoce como estrato compacto (C). Las capas esponjosa y compacta son las que muestran los cambios más espectaculares durante el ciclo y las dos se descaman con la menstruación, por lo que en conjunto, también se llama estrato funcional. La organización de la irrigación arterial del endometrio influye de manera importante en el ciclo menstrual. Las remas de las arterias uterinas llegan a través del miometrio y se dividen inmediatamente en dos tipos distintos de arterias, las arterias rectas y las arterias espirales. Las primeras son cortas y penetran muy poco en el endometrio antes de bifurcarse para formar un plexo que irriga el estrato basal. Las arterias espirales son largas, contorneadas y de pared gruesa, y alcanzan la superficie del endometrio donde originan numerosas ramas que forman un plexo capilar alrededor de las glándulas y en el estrato compacto. A diferencia de las arterias rectas, las espirales responden a los cambios hormonales del ciclo menstrual, de forma que la interrupción de la secreción de progesterona al final del ciclo hace que se contraigan, provocando una fase isquémica que precede inmediatamente a la menstruación. MIOMETRIO. La mayor parte de la masa del útero consiste en músculo liso, formado por haces entrelazados de fibras largas y finas dispuestas en capas mal definidas. El músculo posee una rica red de arterias y venas sostenidas por un denso tejido colágeno. Durante el embarazo, el tamaño del miometrio aumenta mucho, especialmente por el incremento del tamaño celular (hipertrofia), aunque también por un cierto aumento del número de células (hiperplasia), ya que los miocitos pueden dividirse. CUELLO DEL ÚTERO. El canal endocervical (CE) está revestido por una sola capa de células cilíndricas altas productoras de moco. El revestimiento epitelial secretor de moco del canal endocervical forma profundos pliegues y túneles que adoptan el aspecto de glándulas tubulares ramificadas, de donde procede el nombre inexacto de glándulas endocervicales. Durante el ciclo menstrual, la actividad secretora de estas glándulas sufre cambios cíclicos. En la fase proliferativa, los niveles crecientes de estrógenos estimulan la secreción de un moco acuoso y fluido que permite el paso de los espermatozoides hacia el útero durante el período de la ovulación. Una vez pasada ésta, el moco cervical se hace muy viscoso y forma un tapón que impide la entrada de microorganismos y espermatozoides desde la vagina. En la parte del cuello expuesto al ambiente hostil de la vagina, el exocérvix, su revestimiento es un grueso epitelio escamoso estratificado, similar al del resto de la vagina. Las células del exocervix suelen tener citoplasmas claros debidos a su elevado contenido en glucógeno. La unión (U) entre el epitelio vaginal y el endocervical es muy brusca y normalmente se encuentra situada en el OCE, punto en el que el canal cervical se abre en la vagina (V). La mayor parte de la masa del cuello está formada por un fuerte tejido colágeno con una proporción relativamente escasa de músculo liso. Por debajo de la unión escamocilíndrica, el estroma cervical suele estar infiltrado por leucocitos que forman parte de las defensas contra la penetración de microorganismos. 3. TROMPAS DE FALOPIO. La trompas de Falopio (también llamadas trompas uterinas u oviductos) conducen los óvulos desde la superficie de los ovarios a la cavidad uterina y son también el lugar donde se produce la fertilización del óvulo por el espermatozoide. La forma de las trompas de Falopio es la de un embudo alargado que, anatómicamente, puede dividirse en cuatro partes (istmo, ampolla, infundíbulo y fimbrias). Las trompas tienen una estructura básica compuesta por una pared de músculo liso (M), un revestimiento mucoso interior y una capa externa de tejido de sostén laxo. El músculo liso a su vez está formado por dos capas, una de musculo liso longitudinal y otra de musculo liso circular. El movimiento peristáltico de ambas, facilita el movimiento del óvulo en su interior; a ello ayuda también una corriente de líquido impulsada por la acción del epitelio ciliado que reviste la trompa. La fertilización suele producirse en la ampolla. El revestimiento mucoso de la trompa se dispone de un laberinto de ramificaciones y pliegues longitudinales que proporcionan un medio ambiente adecuado para la fertilización. Esta característica alcanza su máxima expresión en la porción ampular. Las células cilíndricas de epitelio (E) son de tres tipos: ciliadas, secretoras no ciliadas y células intercalares. Las células no ciliadas producen una secreción que es impulsada hacia el útero por el movimiento en forma de ondas de los cilios de las células ciliadas, lo que ayuda al transporte del óvulo. Las células intercalares podrían ser una variación morfológica de las células secretoras. La proporción entre células ciliadas y no ciliadas y la altura de todas ellas sufren variaciones cíclicas debida a la influencia de las hormonas ováricas. En general las células ciliadas son más cortas que las secretoras, lo que hace que la superficie del epitelio sea algo irregular. 4. OVARIO. Los ovarios que en una mujer miden unos 3 a 5 cm de longitud, tienen forma oval aplanada. El cuerpo del ovario consiste en células fusiformes, finas fibras de colágeno y sustancia fundamental que, en conjunto, constituyen el estroma ovárico. Las células del estroma son parecidas a los fibroblastos, aunque algunas contienen gotitas de lípidos. También existen algunos haces de fibras musculares dispersos. En la zona periférica del estroma, conocida como corteza, se encuentran numerosos folículos (F) que contienen los gametos femeninos en distintos estadios de desarrollo. Además, también pueden identificarse folículos postovulatorios de varios tipos, en especial cuerpos amarillos (CA)(responsables de la producción de estrógenos y progesterona), antiguos cuerpos amarillos degenerados (D)(corpora albicantes) y folículos degenerados (atrésicos). La corteza superficial es más fibrosa que la corteza profunda y suele recibir el nombre de túnica albugínea (TA). Sin embargo, a diferencia del testículo, en el ovario no existe una cápsula anatómicamente separada. Sobre la superficie del órgano hay una cobertura epitelial, a la que erróneamente se le ha llamado epitelio germinal, que se continúa con el peritoneo. La zona central del estroma ovárico, la médula (M), está muy vascularizada y contiene células hiliares morfológicamente similares a las células de Leydig del testículo. La arteria ovárica (una rama de la aorta) y las ramas ováricas de la arteria uterina forman anastomosis en el mesoovario y en el ligamento ancho. Desde este plexo arterial, alrededor de 10 arterias contorneadas, las arterias helicoidales (H) penetran en el hilio ovárico. Otras ramas aún más pequeñas forman un plexo en la unión corticomedular, dando origen a las arteriolas corticales rectas que se irradian hacia la corteza. Allí se ramifican y anastomosan para formar arcos vasculares de los que surge una rica red de capilares que se disponen alrededor de los folículos. El drenaje venoso sigue el mismo trayecto que el sistema arterial, siendo las venas medulares especialmente grandes y tortuosas. Los linfáticos nacen en el estroma perifolicular y drenan en vasos de mayor calibre que se enrollan alrededor de las venas medulares. La inervación del ovario se hace a través de las fibras simpáticas que no sólo inervan a los vasos sanguíneos, sino también las células musculares lisas del estroma que rodea a los folículos, por lo que es posible que intervengan de alguna manera en la maduración del folículo y en la ovulación. En el ovario maduro, los folículos no desarrollados aparecen como folículos primordiales (FP), formados por un oocito primario rodeado de una sola capa de células foliculares aplanadas. El oocito primario tiene un núcleo grande con cromatina granular finamente dispersa, un nucléolo prominente y escaso citolplasma. En el ángulo inferior derecho de la imagen aparece un folículo estimulado que ha aumentado mucho de tamaño hasta formar un folículo primario; su oocito (O1) es mucho más grande y las células foliculares (F) se han multiplicado por mitosis y han adquirido una forma cúbica; ahora se las conoce como células de la granulosa. Entre el oocito y las células foliculares se desarrolla una gruesa capa homogénea de glucoproteínas y proteoglucanos ácidos, la zona pelúcida (ZP) a cuya formación contribuyen ambos tipos de células. En una fase más avanzada (ángulo inferior izquierdo de la imágen) del desarrollo folicular se observan un folículo de gran tamaño, en el que las células del estroma que lo rodean comienzan a producir una capa organizada alrededor del folículo, la llamada teca del folículo (TF), separada de las células de la granulosa por una membrana basal. Las células de la teca derivan de las células de tipo fibroblástico de estroma ovárico. El folículo primario continúa aumentando de tamaño y las células de la granulosa siguen proliferando hasta constituir una capa de varias células de grosor denominada zona granulosa (ZG). Los folículos primarios continúan desarrollándose hasta formar los folículos secundarios, en ese momento suelen encontrarse en una parte más profunda de la corteza ovárica. La zona de la granulosa (ZG) sigue proliferando y en ella aparecen pequeños espacios llenos de líquido que se fusionan para formar en antro folicular AF, en que se acumula el líquido folicular. En esta fase el oocito (O1) casi ha alcanzado su mayor tamaño y se adopta una posición excéntrica, en una zona engrosada de la granulosa denominada cúmulo ovígero (CO). En la periferia del folículo, la teca ha formado dos capas, la teca interna (TI), que consta de varias capas de células redondeadas, y la peor definida teca externa (TE), formada por células fusiformes que se confunden con el estroma adyacente. Las células de la teca interna muestran las características de las células secretoras de esteroides típicas y secretan precursores de los estrógenos y, en la fase preovulatoria, progesterona. En el ovario, suele decirse que estas células están luteinizadas. Las hormonas del folículo estimulas la proliferación del endometrio preparándolo para la implantación del óvulo fertilizado. La teca externa está formada por células del estroma aplanadas y no tiene funciones endocrinas. Las células de la granulosa también producen hormonas a partir del estadio de la formación del antro; elaboran estrógenos a partir de los precursores secretados por la teca interna y además producen pequeñas cantidades de FSH interfolicular y (en el momento de la ovulación) un inhibidor de la FSH, la inhibina F. Al acercarse el momento de la maduración, cesa el crecimiento del oocito y se completa la primera división meiótica inmediatamente antes de la ovulación. En este estadio el oocito se conoce como oocito secundario (folículo de Graaf) y comienza la segunda división meiótica. El primer cuerpo polar, que tiene un citoplasma muy escaso, permanece anodino en la zona pelúcida. El antro folicular aumenta mucho de tamaño y la zona de la granulosa forma una capa incluso más gruesa en torno a la periferia del folículo. El cúmulo ovígero disminuye, dejando al oocito (O2) rodeado por una capa de varias células de grosor, la corona radiada (CR), que permanece unida a la zona de la granulosa mediante unos puentes celulares. Antes de la ovulación, estos puentes se rompen y el oocito, en el seno de la corona radiada, flota libremente en el interior del folículo. En esta fase, el folículo mide entre 1,5 y 2,5 cm y protruye bajo la superficie del ovario. Las células epiteliales superficiales se aplanan y atrofian y el fino estroma interpuesto degenera y pierde los vasos. En el momento de la ovulación, el folículo maduro se rompe y el óvulo, formado por el huevo secundario, la zona pelúcida y la corona radiada, es expulsado a la cavidad abdominal cerca de la trompa de Falopio. La segunda división meiótica del oocito sólo se completa después de que un espermatozoide haya penetrado en el óvulo. Tras la ovulación, el folículo roto se colapsa y queda ocupado por un coágulo hemático, formándose el cuerpo amarillo de la menstruación, con una breve carrera como órgano endocrino. El tamaño de ese cuerpo amarillo es casi igual al de un folículo ovulatorio previo, es decir, 1,5 a 2,5 cm. Bajo la influencia de la hormona luteinizante (LH) secretada por el lóbulo anterior de la hipófisis, las células de la granulosa aumentan mucho de tamaño y comienzan a secretar progesterona. Su amplio citoplasma contiene gran cantidad de retículo endoplásmico liso, abundantes mitocondrias, gotitas de lípidos y cierta cantidad de lipofuscina, lo que en conjunto proporciona el color amarillo que lo caracteriza macroscópicamente. En ese momento, las células de la granulosa se conocen como células luteínicas de la granulosa. La progesterona estimula la secreción de las glándulas del endometrio que, para entonces han proliferado mucho gracias a la influencia de los estrógenos secretados por el folículo antes de la ovulación. Se logra así un ambiente propicio para la implantación de un óvulo fertilizado. Las células de la teca interna también aumentan algo de tamaño y adquieren unas características citoplasmáticas similares a las de las células de la granulosa luteinizadas. Aunque interrumpidas por la ovulación, estas células continúan secretando estrógenos, que son necesarios para mantener el grosor de la mucosa uterina. Estas células se llaman ahora células luteínicas de la teca. La producción de progesterona por el cuerpo amarillo depende de la LH secretada por la hipófisis anterior, pero la elevación de los niveles de progesterona inhibe la producción de LH. Sin el estímulo continuo de la LH, el cuerpo amarillo no puede mantenerse pasados 12ª 14 días de la ovulación y, por tanto, involuciona para dar lugar a un cuerpo albicans no funcionante. Cuando el cuerpo amarillo regresa, se interrumpe la secreción tanto de estrógenos como de progesterona. Sin estas hormonas el revestimiento endometrial se colapsa y comienza la menstruación. Sin embargo la implantación de un óvulo fertilizado en la pared uterina interrumpe los ciclos ovárico y endometrial integrados. Tras la implantación, la placenta en desarrollo comienza a secretar hacia la circulación materna la hormona HCG, de función análoga a la LH, por lo que mantiene la función del cuerpo amarillo en lo que se refiere a la secreción de estrógenos y progesterona hasta la semana 12. A partir de ese momento el cuerpo amarillo del embarazo comienza a regresar lentamente hasta formar un cuerpo albicans no funcionante, en tanto que la placenta se convierte en el lugar principal de secreción de estrógenos y progesterona hasta el momento del parto. 5. MAMA. Las mamas (glándulas mamarias) son glándulas sudoríparas apocrinas muy modificadas que en el embrión se desarrollan a lo largo de dos líneas, las líneas de la leche, que van desde las axilas a las ingles. En el hombre sólo se desarrolla una glándula a cada lado del tórax, aunque en cualquier lugar de la línea de la leche puede encontrarse tejido mamario accesorio. Este diagrama, muy esquemático, muestra la organización general de la mama. Cada una de ellas consta de 15 a 25 unidades glandulares independientes llamados lóbulos mamarios, constituidos por una glándula túbulo-acrinar compuesta. Los lóbulos están inmersos en una masa de tejido adiposo subdividido por tabiques colágenos. Hay tabiques fibrosos más robustos que separan el territorio de cada lóbulo. Los lóbulos se disponen de manera radial a distintas profundidades alrededor del pezón y un conducto grande y único llamado conducto galactóforo, drena a cada lóbulo a través de un orificio separado que se abre en la superficie del pezón. Inmediatamente antes de su apertura en la superficie, el conducto forma una dilatación llamado seno galactóforo. El pezón contiene bandas de músculo liso orientadas en paralelo con los conductos galactóforos y circulares cerca de la base; la contracción de ese músculo provoca la erección del pezón. En el interior de cada lóbulo mamario, el conducto principal se ramifica repetidamente para formar varios conductos terminales, cada uno de los cuales da lugar a un lobulillo compuesto por múltiples ácinos (revestidos por dos capas celulares, una capa luminal de células epiteliales y una capa basal de células mioepiteliales aplanadas). Cada conducto terminal y su lobulillo asociado recibe el nombre de unidad ducto-lobulillar terminal. Los lobulillos están separados por tejido interlobulillar colágeno de densidad moderada, mientras que el tejido de sostén intralobulillar que rodea a los conductos de cada lobulillo tiene menos colágeno y está más vascularizado. La piel que rodea al pezón, la areola, esta pigmentada y contiene glándulas sebáceas no asociadas a folículos pilosos.