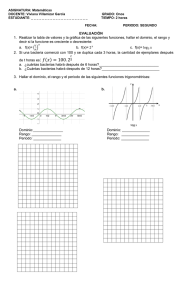
Descargar
Anuncio

1 E L M U N D O D E L O S M I C R O B I O S Autor: GEORGES DREYFUS CORTÉS http://bibliotecadigital.ilce.edu.mx/sites/ciencia/menu.htm Compilación y armado: Sergio Pellizza dto. Apoyatura Académica I.S.E.S. COMITÉ DE SELECCIÓN EDICIONES DEDICATORIA AGRADECIMIENTOS PRÓLOGO I. LOS PRINCIPIOS DE LA MICROBIOLOGÍA II. METABOLISMO BACTERIANO III. LAS BACTERIAS Y LA HISTORIA DE LA TIERRA IV. LOS MICROBIOS AL SERVICIO DEL HOMBRE CONTRAPORTADA 2 C O M I T É D E S E L E C C I Ó N Comité de Selección: Dr. Antonio Alonso Dr. Juan Ramón de la Fuente Dr. Jorge Flores Dr. Leopoldo García-Colín Dr. Tomás Garza Dr. Gonzalo Halffter Dr. Guillermo Haro Dr. Jaime Martuscelli Dr. Héctor Nava Jaimes Dr. Manuel Peimbert Dr. Juan José Rivaud Dr. Emilio Rosenblueth Dr. José Sarukhán Dr. Guillermo Soberón Coordinadora Fundadora: Física Alejandra Jaidar Coordinadora: María del Carmen Farías E D I C I O N E S Primera edición, 1987 Quinta reimpresión, 1996 La Ciencia desde México es proyecto y propiedad del Fondo de Cultura Económica, al que pertenecen también sus derechos. Se publica con los auspicios de la Subsecretaría de Educación Superior e Investigación Científica de la SEP y del Consejo Nacional de Ciencia y Tecnología. 3 D. R. © 1987, FONDO DE CULTURA ECONÓMICA, S. A. DE C. V. D. R. © 1995, FONDO DE CULTURA ECONÓMICA Carretera Picacho-Ajusco 227; 14200 México, D.F. ISBN 968-16-2703-2 Impreso en México D E D I C A T O R I A Si de todos los organismos creados por Dios los más pequeños y aparentemente menos útiles fueran suprimidos, la vida se tornaría imposible, ya que el regreso a la atmósfera y al reino mineral de todo lo que dejó de vivir sería bruscamente suprimido. LOUIS PASTEUR A G R A D E C I M I E N T O S Quiero agradecer la paciente ayuda de Nora Vázquez Laslop, Francisco Mariscal y María del Carmen Farías en la preparación del texto. También agradezco a la señora Guadalupe Ramírez, quien mecanografió una y otra vez el manuscrito, y al personal de mi laboratorio, por su constante interés. Las microfotografías se deben al doctor Alfonso Cárabez y al señor Jorge Sepúlveda de la Unidad de Microscopía Electrónica de este Instituto. P R Ó L O G O La idea básica de este libro es la de describir en términos sencillos el desarrollo de algunas áreas del conocimiento sobre los microorganismos. No ha sido mi intención abarcar todos los aspectos que un tema como éste presenta, sino sólo aquellos que, en mi opinión, interesan a un lector ajeno a la microbiología y, en general, a la ciencia. He tratado que su lectura sea amena y fluida por eso evité entrar en detalles sólo interesantes para un especialista. Es innegable que la vida en nuestro planeta está sustentada en un frágil equilibrio ecológico. La materia que da origen a todas las 4 formas de vida es utilizada a través de complicadas cadenas alimenticias. Dentro de este panorama tienen un papel muy importante los microbios, pequeñísimos seres que poseen las más diversas capacidades adaptativas y que ejercen una influencia determinante en el mantenimiento del equilibrio ecológico. Así pues, este libro empieza describiendo cómo la humanidad se dio cuenta de la existencia de los microorganismos, lo que le permitió explicar la causa de algunas enfermedades, hasta el grado en que se creó una ciencia encargada de su estudio. Estos pequeños organismos han facilitado el desarrollo del conocimiento sobre los aspectos bioquímicos de la vida. Veremos en las páginas siguientes cómo los microorganismos se adaptaron a diversos ambientes, tan extremos algunos que semejan, quizá, las condiciones que prevalecían en nuestro planeta hace millones de años; y cómo han constituido una ayuda invaluable en el estudio de los orígenes de la vida en nuestro planeta. Gran parte de los microorganismos no es nociva para el hombre, al contrario, es prometedora para la industria en el campo de la obtención de diversas sustancias y en la conservación del medio ambiente, mediante el control de la contaminación de la tierra y las aguas. La curiosidad científica, que aparentemente no conduce a nada, ha sido sin duda la que ha permitido al género humano poseer el conocimiento actual sobre la naturaleza. Este conocimiento, generado por el quehacer de la ciencia, ha permitido obtener resultados que han modificado la historia y el curso de la humanidad. Ciudad Universitaria, México D. F., mayo de 1987 L O S P R I N C I P I O S D E M I C R O B I O L O G Í A L A EL MUNDO de los microbios o microorganismos es quizá tan vasto como el Universo y, para dar un pequeño ejemplo, un puñado de tierra contiene tantos microorganismos como seres humanos el planeta entero. Esto representó un gran obstáculo para su estudio, ya que existía la dificultad de tener que separar un microorganismo de los demás. Y, de cualquier forma, si se lograra separar dicho microorganismo, ¿cómo se mantendría aislado de sus similares? La labor y el esfuerzo combinados de muchos hombres y mujeres de ciencia, a lo largo de unos 200 años de historia, dio como resultado el surgimiento de la microbiología, la ciencia que estudia estos pequeños seres de la naturaleza. Los métodos mediante los cuales ha sido posible separar a los microorganismos de sus alrededores, pueden ser descritos en orden de aparición, ya que los problemas 5 que surgen durante el desarrollo de una ciencia experimental sólo pueden ser comprendidos a la luz del conocimiento que prevalecía en su tiempo. I.1 LAS PRIMERAS IDEAS La teoría de la generación espontanea. La creencia de que existían seres tan pequeños que eran invisibles se remonta a tiempos tan lejanos como antes de la Era Cristiana. Doscientos años antes de ella, Varro ya proponía la posibilidad del contagio de ciertas enfermedades debido a criaturas invisibles suspendidas en el aire, y esta idea ya era compartida por los antiguos médicos latinos y árabes. Figura 1. Foto de una bacteria, la Escherichia coli Lucrecio fue conocido por su punto de vista peculiar sobre la materia, ya que pensaba que las cosas surgían de una especie de átomo o semilla. En su obra De Rerum Natura (75 a. C.) sugiere que las plagas eran causadas por una especie de átomo. Y en el libro VI dice: "Así como hay semillas benéficas para nuestra vida, seguramente existen otras que causan enfermedad y muerte," Este punto de vista puede ser considerado como un claro ejemplo de que se intuía ya que la naturaleza de la infección era microbiana. Sin embargo, estas semillas no se consideraban como organismos vivientes. Por otra parte, prevalecía la idea de que las cosas emergían de semillas y que sólo la combinación de varias de ellas podría dar origen a un organismo completo. Así pues, existía la duda sobre el 6 origen de la primera semilla. De alguna forma esta primera semilla se había generado espontáneamente, es decir que podían aparecer organismos vivientes a partir de materia no viviente. A esta corriente de pensamiento se le conoció como la teoría de la generación espontánea y persistió por más de 1 500 años. El contagio de las enfermedades. La epidemiología es la rama de la medicina que estudia la propagación de las enfermedades y se inició mucho antes de que se aplicara el término "enfermedad contagiosa" por los aún desconocidos agentes causantes de las infecciones. En la Antigüedad, los hebreos consideraban a las enfermedades infecciosas como un castigo proveniente del Señor. Sin embargo, los escritos bíblicos ya indicaban que las personas con lepra debían ser aisladas y los artículos y materiales que se utilizaban durante su enfermedad, enterrados o quemados. Más tarde, Lucrecio y Boccaccio reconocieron la naturaleza contagiosa de estos brotes epidémicos y, por fin, en 1546, Fracastorius de Verona presentó su obra titulada De Contagione. Este libro fundó la ciencia de la epidemiología propiamente dicha. Después de estudiar cuidadosamente la epidemiología de varias enfermedades, incluyendo la plaga y la sífilis, Fracastorius concluyó que la transmisión se producía de persona a persona, directamente o bien por medio de objetos de uso común. Más tarde otros, y él mismo, mezclaban las teorías racionales con otras ideas basadas en la superstición. Por ejemplo, pensaban que las plagas o epidemias eran gobernadas por fuerzas sobrenaturales. Como resultado de esta confusión, casi un siglo después médicos prominentes como William Harvey no tomaban en serio las ideas de Fracastorius y se adhirieron a las ideas de Hipócrates y Galeno que consideraban que las epidemias o vapores venenosos eran generados por conjunciones planetarias o por alteraciones en la propia Tierra. El panorama era aún más complicado si se toma en cuenta que las enfermedades pueden ser transmitidas de distintas maneras, como puede ser por el agua, la comida o los insectos y que en muchas ocasiones estas formas de transmisión no son tan obvias. Las ideas de Lucrecio sobre la naturaleza de la materia fueron resucitadas por Giordano Bruno,al final del siglo XVI, e influenciaron profundamente a Robert Boyle. Sin embargo, el simple razonamiento sobre la existencia de los microbios no constituyó la prueba de su verdadera existencia. Ésta sólo pudo ser demostrada gracias al descubrimiento de una lente de aumento. I.2 LAS PRIMERAS OBSERVACIONES El microscopio abrió las puertas de otro universo. El holandés Antonie van Leeuwenhoek (1632-1723) fabricó la primera lente lo suficientemente poderosa como para observar a los organismos unicelulares. Utilizó una lente convexa y colocó al objeto dentro de la distancia focal (Figura 2). Este arreglo produce una imagen virtual o aparente muy amplificada; con este rudimentario pero 7 ingenioso microscopio logró observar muchos "pequeños animalillos". Éstos incluían protozoarios, tanto de vida libre como parásitos de las vísceras de algunos animales; también logró observar hongos filamentosos y corpúsculos globulares que ahora conocemos como levaduras. Hizo importantes observaciones sobre la estructura de las plantas y descubrió los espermatozoides de algunos animales. Fue hasta 1676 que pudo observar organismos aún más pequeños, como las bacterias. Figura 2. Diagrama del microscopio utilizado por Leewenhoeck. El objeto "O" se coloca dentro de la distancia focal "F" y esto da por resultado una imagen virtual "T" muy amplificada. Por otra parte, el microscopio compuesto, capaz de ampliar la imagen con mayor eficiencia y más conveniente para su uso, fue inventado en 1590 por Zacharías Jensen y usado en 1630 por F. Stellerti para estudiar la miel de abeja y los escarabajos. Sin embargo, todos los modelos producidos hasta antes de 1800 no poseían la resolución y la calidad de los microscopios de un solo lente. Las observaciones de Leeuwenhoek llamaron mucho la atención, pero eso fue todo. Nadie en esta época trató de repetir o extender sus observaciones, ya que la alta calidad de sus lentes y su gran agudeza visual fueron factores determinantes para desalentar a cualquier competidor. A esto se debió el gran retraso que sufrió el desarrollo de microscopios más potentes. I.3 LA CONTROVERSIA DE LA GENERACIÓN ESPONTÁNEA ¿La vida proviene de la vida o existe la fuerza vital? La comprensión verdadera de la importancia de los microorganismos en el mundo 8 comenzó como resultado de la controversia sobre la generación de materia viviente a partir de la materia muerta. En un principio existían dos escuelas bien definidas de pensamiento: aquélla que tomando a Lucrecio al pie de la letra apoyaba la idea de que se podrían generar animales a partir de materia muerta gracias a la existencia de una "fuerza vital" (generación espontánea) y la que decía que la vida sólo se genera a partir de vida (en latín omne vivum ex vivo). Los antiguos que creían en la generación espontánea daban recetas para preparar "ratones" a partir de comida en putrefacción. Opuestas a este punto de vista eran las ideas de Redi, quien en 1668 mostró que la aparente generación espontánea de larvas en la carne provenía de la visita de las moscas que ponían huevecillos sobre ella. Sin embargo, Needham, otro investigador, hirvió extracto de carne en un frasco, lo tapó y encontró que después de algunos días aparecían criaturas que se movían. Esto, aunado a la idea de que los organismos vivientes morían al ser hervidos, llevó a pensar que dichos organismos eran realmente producto de la generación espontánea. Más tarde, Spallanzani llevó a cabo experimentos más cuidadosos con los que demostró que los organismos grandes eran destruidos al ser hervidos durante 30 segundos, pero los microorganismos sobrevivían y se desarrollaban aunque los frascos estuvieran herméticamente cerrados. Después de muchos ensayos, encontró que si hervía los frascos parcialmente cerrados durante 45 minutos, el contenido se mantenía sin contaminarse casi indefinidamente, y sólo si se permitía la entrada de aire, el contenido entraba en putrefacción rápidamente. Estos resultados llevaron a pensar que, al hervir el contenido, el aire del frasco se hacía inadecuado para la existencia de vida en su interior y esto era interpretado como la ausencia de la "fuerza vital". Un cocinero francés, llamado Francois Appert, a principios del siglo XIX desarrolló el arte de preservar comida en frascos sellados: lo lograba hirviendo el contenido dentro del frasco y cerrándolo sin permitir la entrada de aire fresco. Observó que así el contenido se mantenía libre de microorganismos por tiempo indefinido. Este hallazgo lo llevó no solamente a fundar una importante industria, la de las conservas, sino a ser inmensamente rico. La objeción a los experimentos de Spallanzani de que al hervir el contenido de los frascos se terminaba con la "fuerza vital" fue descartada por un experimento de Schultze en 1836. Éste consistió en que el recipiente que contenía extracto de carne fue conectado a otros dos recipientes, uno de los cuales contenía ácido sulfúrico y el otro potasa; a través de éstos se hizo pasar lentamente aire fresco todos los días durante tres meses y el extracto de carne no se contaminó. La clave del éxito de este experimento se debió a que las conexiones con ambos recipientes se hicieron inmediatamente 9 después de hervir el extracto contaminación del extracto. de carne, lo cual evitó la Theodor Schwann, en 1837, llevó a cabo un experimento similar, pero la diferencia consistió en que el aire fresco se hacía pasar por un recipiente que contenía un metal fundido en ebullición y de esta forma cualquier materia orgánica se mantenía estéril en el interior. Sin embargo, cuando se dejaba pasar aire fresco sin entrar en contacto con el metal fundido, el contenido se contaminaba invariablemente, con bacterias para el caso del extracto de carne y con levaduras para una solución con azúcar. La interpretación que dio Schwann a sus resultados fue la siguiente: "Los microorganismos que deben estar presentes en el aire son destruidos al hacer pasar el aire por un líquido incandescente. Por lo tanto, la putrefacción sin duda se debe al hecho de que estos gérmenes, al nutrirse y desarrollarse a costa de esta sustancia, la descomponen y sobreviene la putrefacción." Más tarde, la técnica de estos experimentos fue simplificada y en 1853 Schroeder y Von Dush descubrieron que, después de hervir el recipiente, bastaba con cerrar el extremo abierto con un tapón de algodón. De esta manera, el aire que entraba a través del algodón se filtraba al pasar por los múltiples y tortuosos caminos dentro del mismo. Estos experimentos abrieron el camino sobre el cual se basaron los métodos para cultivar bacterias. Dos postulados importantes se derivan de estos resultados: el primero consiste en hacer un medio de cultivo adecuado que esté libre de microorganismos y esto se logra hirviéndolo; el segundo consiste en mantener el medio estéril por largo tiempo y esto se logra evitando la entrada de los microorganismos que están suspendidos en el aire. A pesar de estos avances, los resultados de Spallanzani, Schultze y Schwann no fueron aceptados por la mayoría del público científico de la época. Hoy en día sabemos que muchos microorganismos resisten temperaturas hasta de l00°C durante varias horas. Esto ayudó a los que pensaban que la generación espontánea era un hecho, ya que un solo experimento fallido era utilizado como evidencia para la existencia de esta teoría. Koch, más tarde, llevó a cabo estudios sobre el bacilo del ántrax y encontró que las esporas de algunas bacterias eran altamente resistentes al calor y que sólo se destruían a l20°C o más, y eso al cabo de 20 minutos. Pasteur fue quien desechó la teoría de la generación espontánea. Fue el genio de Louis Pasteur (1822-1895) quien finalmente persuadió al pensamiento científico sobre lo verdadero de la generalización tan claramente formulada por Schwann en 1837. Pasteur se valió de una combinación de habilidad científica, persuasión y un sentido muy particular del drama. Su experimento consistió básicamente en hervir infusiones en un aparato tal que, cuando el contenido se enfriaba, el aire que entraba era calentado y luego vuelto a enfriar al pasar por una camisa enfriada con agua. Bajo estas condiciones la infusión se conservaba libre de 10 contaminación. La figura 3 ilustra el aparato tal como fue diseñado por Pasteur. Sin embargo, el experimento más sorprendente fue aquel en el que Pasteur mostró cómo un medio nutritivo permanecía estéril aun cuando estuviera comunicado con el exterior. Para esto diseñó unos frascos especiales (Figura 4) en los que, como se puede ver, se permite la entrada de aire, pero no se contaminan porque en el primer doblez del cuello se retienen los posibles microbios contaminantes. Estos frascos se encuentran hoy en día tal y como los dejó este notable hombre de ciencia. Figura 3. Experimento diseñado por Pasteur para demostrar que si el aire que entra al recipiente cuyo contenido hirvió es previamente calentado, el contenido no se contamina. El aire caliente se enfría con un trapo mojado antes de que éste llegue al caldo nutritivo. Pasteur confirmó también los experimentos de Schroeder y Von Dusch, quienes hirvieron una infusión de carne y cerraron los frascos con tapones de algodón. Este experimento lo amplió Pasteur utilizando tapones de nitrato de celulosa en lugar de algodón, para después disolver el tapón en alcohol, de tal forma que los gérmenes atrapados en el tapón fueran liberados; después examinó el contenido del tapón bajo el microscopio y observó partículas globulares que semejaban esporas de hongos y bacterias. A pesar del éxito de los experimentos de Pasteur no siempre se lograba reproducirlos, y esto se debía principalmente a la resistencia de ciertas esporas o microbios a las altas temperaturas. 11 Sin embargo, tal dificultad no pudo opacar el concepto general que Pasteur logró aclarar. Figura 4. Botellas de cuello de ganso que Pasteur dejó como prueba de que la putrefacción proviene del crecimiento de microorganismos y no de la generación espontánea. El físico Tindall estaba interesado en los fenómenos de la dispersión de la luz en el agua y en el aire (fenómeno que ahora se conoce como efecto Tindall) y sus observaciones se apoyaron en los experimentos de Pasteur. Tindall encontró que un rayo de luz puede observarse mejor cuando el humo de un cigarrillo pasa a través de él. Lo mismo ocurre en un cámara en cuyo interior hay aire normal. Al pasar un rayo de luz se observarán pequeñas partículas que flotan y que, él pensó, podían ser atribuidas a la presencia de los microorganismos responsables del fenómeno de putrefacción. Tindall mostró que el aire normal contenido en una cámara hermética presenta una serie de partículas diminutas que se hacen aparentes al incidir un rayo de luz en la cámara. Sin embargo, después de algunas semanas las partículas se depositan en el fondo y el rayo de luz ya no es dispersado. A esto Tindall lo llamó cámara ópticamente vacía. En 1869 este físico inglés demostró que al llenarse una cámara con aire que se forzaba a pasar a través de un algodón, se lograba obtener aire ópticamente vacío. La relación de sus experimentos con los de Schwann y Pasteur fue obvia para él: las pequeñas partículas no eran otra cosa que microorganismos. Demostró que el aire ópticamente vacío no iniciaba el proceso de putrefacción. En 1876 diseñó una cámara (Figura 5) que permitía observar tanto la presencia de partículas contenidas en el aire como la putrefacción de cualquier sustancia depositada en el interior de la cámara. Esta cámara tenía además una entrada para depositar la muestra (B) y una ventilación que servía también para equilibrar las presiones (A) 12 y consistía en un tubo con varios dobleces. Para determinar la presencia de partículas en el aire, el rayo de luz (E) se hacía pasar como se muestra en la figura. El 10 de septiembre de 1875 se cerró la caja de Tindall y, al pasar el rayo de luz, éste se hizo aparente por la presencia de las partículas suspendidas en el aire. Para el 13 de septiembre, el aire en su interior estaba ópticamente vacío. El experimento se llevó a cabo utilizando un caldo de cultivo que fue depositado en los tubos de ensayo del interior. El caldo hirvió durante 5 minutos al ser sumergidos los tubos en un baño hirviente de salmuera. Por otra parte, otros tubos que también fueron hervidos se dejaron fuera de dicha caja. Para el cuarto día, los tubos que quedaron fuera de la caja estaban ya turbios mientras que los que estaban en su interior se mantuvieron trasparentes durante 4 meses. Al abrir la caja el contenido de los tubos se volvió turbio en 4 días. Estos experimentos se repitieron en varias ocasiones con otro tipo de infusiones siempre con el mismo resultado y éxito. La inferencia era clara, el aire ordinario contiene normalmente cantidades variables de microbios y son a éstos precisamente a los que se debe la mal llamada "generación espontánea". Figura 5. Cámara de Tindall. La fermentación debe estar asociada a un microbio. A pesar de que Pasteur logró demostrar que la contaminación de un caldo de cultivo se debe a la presencia de microorganismos, seguía siendo importante saber cómo ocurrían los procesos de putrefacción de la materia. Así fue como otra serie casi innumerable de sucesos ocurrieron y permitieron descartar definitivamente las ideas sobre la generación espontánea. En estudios posteriores sobre la 13 fermentación butírica y la producción de vinagre, Pasteur encontró que estos procesos se debían al desarrollo de microorganismos específicos como son el Vibrion butiricum y el Mycoderma aceti. A pesar de esto, Liebig consideraba que la putrefacción y la fermentación eran producidas por "fermentos" (del latín fermentum, que viene de fervere hervir y se refiere a la evolución del gas) que, según él, consistían de materia orgánica en descomposición. Se consideraban fermentos, por ejemplo, las partículas de queso viejo que al mezclarse con un poco de leche hervida hacían que ésta se acidificara rápidamente y, según Liebig, tal reacción se debía a la existencia de fermentos. La existencia de levaduras, que para Pasteur explicaba la fermentación, para Liebig sólo significaba que estos microorganismos, al morir, se descomponían y, por lo tanto, actuaban también como fermento. Sin embargo, Pasteur finalmente demostró que las levaduras originan el proceso de fermentación mientras crecen. En resumen, Pasteur aclaró los conceptos sobre la fermentación y definió que ésta es el producto de una reacción que lleva a cabo un microorganismo y que cada microorganismo tiene su propio tipo característico de fermentación. Las levaduras producen alcohol, las bacterias lácticas ácido láctico, el vibrión butírico ácido butírico, etcétera. Otro investigador de la época, Gay-Lussac, proponía que la fermentación era el resultado de una serie de reacciones químicas. Sin embargo, sus resultados eran poco refinados como para que se sostuvieran ante las evidencias de Pasteur. Edward Buchner, más adelante, logró demostrar en forma accidental que la fermentación ocurría en un extracto de levaduras, y comenzó a pensar en la fermentación como una cadena de reacciones químicas que podían ocurrir aun en ausencia del microorganismo. I.4 CÓMO SON LAS BACTERIAS Es importante recordar que las bacterias no poseen todas las estructuras que contienen en su interior las células de los organismos que aquí llamaremos superiores y éstas pueden ser desde las levaduras hasta las células de cualquier animal. La figura 6 muestra estas diferencias estructurales. Las células bacterianas presentan dos características principales. La primera es que no tienen un núcleo que contenga al ácido desoxirribonucleico (ADN), el cual es el material genético, sino que éste se encuentra libre en su interior (citoplasma). El tamaño de esta molécula de ADN es varios cientos de veces más grande que la bacteria misma y contiene toda la información hereditaria necesaria para asegurar la superviviencia de la pequeña célula. La segunda es que el citoplasma contiene, además, moléculas de ARN, llamadas ribosomas, cuya función es la de ensamblar proteínas que tendrán, a su vez, diversas funciones. Estos ribosomas son más pequeños que los de las células de los animales superiores. 14 Las bacterias tienen también envolturas celulares diferentes a las de las células de los animales superiores. Todas presentan una pared formada de un componente químico específico llamado peptidoglicano, que es el responsable de dar a la célula una envoltura resistente. La presencia de la llamada pared celular ha permitido catalogar a las bacterias en gram positivas y gram negativas. Las bacterias que se tiñen con un colorante violeta son las positivas para la tinción de gram y no tienen pared celular. Otras no se tiñen con el colorante y se les conoce como gram negativas. 15 Figura 6. Diferencias estructurales entre la células superiores y las bacteias. Las bacterias pueden tener organelos que les permitan moverse. Los más comunes son los flagelos, que se proyectan de su superficie en uno de los extremos de la célula. Las bacterias pueden ser de muchas formas: esféricas, en forma de bastón y hasta ramificadas. En general, su tamaño es muy inferior al de una célula de un organismo superior y su multiplicación es por división asexual. Durante su ciclo reproductivo se forma una pared divisoria después de que su cromosoma, formado de ADN que contiene la información genética, se ha duplicado. De esta forma las dos porciones de la célula se separan, conteniendo cada una de las nuevas células su propio cromosoma (Figura 7). Algunas bacterias tienen estructuras conocidas como endosporas, las cuales pueden resistir el paso del tiempo y aun agresiones tales como altas temperaturas y productos químicos tóxicos que acaban normalmente con una bacteria. Estas esporas permanecen en estado latente y, bajo condiciones adecuadas, pueden dar lugar a una nueva bacteria. Las bacterias están íntimamente ligadas a la existencia de la vida sobre la Tierra. Son causantes de muchas enfermedades, pero también en muchos casos son las responsables de la continuidad de la vida. Los microbios y las enfermedades. Sin duda una de las cargas que más ha pesado sobre la humanidad ha sido las enfermedades causadas por los microorganismos. No solamente eran la causa más frecuente de muerte sino, además, de las enfermedades que atacaban a los menores de edad. No en pocas ocasiones a causa de la naturaleza epidémica de las infecciones, ejércitos y naciones enteras fueron exterminados. Una de las grandes hazañas de las ciencias médicas y biológicas ha sido el descubrimiento de la causa y el control de estas epidemias devastadoras. Hoy en día es fácil dar por hecho estos logros tan importantes y preocuparse por los problemas derivados de los avances tecnológicos. Sin embargo, Pasteur y Koch llegaron a ser considerados héroes por haber rescatado a la humanidad de una de las más grandes amenazas con la que tuviera que enfrentarse: la contaminación microbiana. Es indudable que el avance de la microbiología como una ciencia es inseparable de su función como pilar fundamental de la medicina moderna. 16 Figura 7. Ciclo reproductivo de una bacteria. La reproducción se lleva a cabo asexualmente por división celular. El diagrama muestra las siete etapas del ciclo de vida de la bacteria Escherichia coli que por lo común se encuentra poblando el intestino grueso de los humanos. 1) El único cromosoma circular de ADN ya se empieza a replicar. 2, 3) A los 20 minutos el nuevo cromosoma está completo y se ha fijado a un sitio en la célula. 4) A los 25 minutos los dos cromosomas se han empezado a duplicar. 5) Aparece una división en el centro de la célula. 6) A los 38 minutos la división es ya una pared. 7) A los 45 minutos la división se ha completado. Los microbios del suelo. Al final de la era encabezada por Pasteur, otros investigadores, como lo fueron Winogradsky en Rusia y Beijerinck en Holanda, comenzaron a explorar la microbiología del suelo y descubrieron para su sorpresa una gran variedad de microorganismos en los diferentes tipos de suelo que estudiaron. Con el desarrollo de la "microbiología del suelo" se hizo aparente que el principal papel que desempeñan los microbios en la naturaleza es geoquímico. Gracias a la presencia de bacterias en la tierra, los compuestos orgánicos como el carbono, el nitrógeno y el azufre pueden ser usados cíclicamente por las plantas y los animales, en vez de que dichos elementos se encuentren como materia inorgánica muerta. 17 Un hecho interesante es que la "fertilidad" de la tierra se acrecienta si en ésta hay bacterias que sean capaces de convertir nitrógeno atmosférico o amoniaco proveniente de la descomposición de material orgánico en nitrato (que viene a ser la forma no volátil de estos compuestos), forma en la cual es aprovechado por las plantas que en ella crecen. Por otra parte las algas y, en una menor proporción, las bacterias fotosintéticas llevan a cabo la otra mitad del ciclo geoquímico, la formación de materia orgánica a partir de bióxido de carbono (C02) por medio de la fotosíntesis. Es importante enfatizar el papel benéfico de ciertos microbios, ya que la conexión histórica entre las enfermedades infecciosas y la microbiología ha dado lugar a la imagen popular de que el mundo de los microbios es maligno y hostil. Sin embargo, la fracción de microbios nocivos para el hombre es mínima con respecto al del total de la masa microbiana sobre la Tierra. La mayoría de los microbios ataca la materia orgánica muerta cuando está bajo tierra y constituye un factor determinante en la transformación de la materia orgánica. I I . M E T A B O L I S M O B A C T E R I A N O TODOS los seres vivos llevan a cabo el procesamiento de los nutrientes que los mantienen vivos. A este conjunto de procesos, se le conoce como metabolismo y consiste de un gran número de reacciones químicas destinadas a transformar las moléculas nutritivas en elementos que posteriormente serán utilizados para la síntesis de los componentes estructurales; como pueden ser las proteínas. Otra parte importante del metabolismo es la de transformar y conservar la energía que está contenida en una reacción química en algún proceso que requiera de energía, como puede ser el trabajo o el movimiento. Es evidente que los nutrientes son transformados cuando entran en un organismo, ya que en ningún caso el alimento contiene todas las moléculas que una célula requiere. Esto se vio con claridad al observar el crecimiento normal de levaduras en un medio de cultivo que sólo contenía glucosa como única fuente de energía. Así pues, se pensó que la síntesis de todos los componentes celulares se llevaba a cabo en el interior de las levaduras. Hoy sabemos que las transformaciones que sufre la glucosa no ocurren en un solo paso, sino que, por el contrario, se forman varios productos intermedios que en muchas ocasiones no tienen una función específica a no ser la de formar parte de lo que se conoce como vía metabólica. La transformación de los nutrientes en compuestos útiles para la subsistencia de un organismo se lleva a cabo por medio de las reacciones químicas que realizan unas proteínas conocidas como enzimas. De tal forma que no podemos hablar del metabolismo si no describimos brevemente qué son y cómo funcionan las enzimas. 18 II.1 LAS ENZIMAS SON EFICIENCIA Y RAPIDEZ La enzimología es una parte fundamental de la bioquímica que se ha desarrollado rápidamente en los últimos cinco decenios. Ciertas manifestaciones de la acción enzimática eran conocidas desde mucho tiempo atrás y si bien sus causas eran ignoradas, ya se conocían varios procesos como la fermentación, la digestión y la respiración o consumo de oxígeno. Las enzimas son proteínas especializadas capaces de transformar químicamente una molécula; son sin duda las moléculas biológicas más notables ya que solamente transforman a una molécula y a ninguna otra y esto lo hacen varios cientos de veces por segundo. Estas características las hacen mucho mejores que cualquier catalizador químico hecho por el hombre. Figura 8. (a) Cadena lineal de aminoáidos. (b) Cadena tridimensional de aminoácidos. Pasteur pensaba que la fermentación era llevada a cabo por enzimas y que su acción estaba íntimamente ligada a la estructura y a la vida de las levaduras. Un hecho de gran importancia que marcó el inicio de la enzimología y de la bioquímica ocurrió cuando, en 1877, Edward Buchner logró que un extracto libre de levaduras enteras llevara a cabo la fermentación alcohólica. Esto implicaba que tales enzimas, de gran importancia para la célula viva, eran capaces de llevar a cabo su función con independencia de la integridad de la estructura celular. En 1926, el notable bioquímico J. B. Sumner aisló y cristalizó por primera vez una enzima, la ureasa de frijol, y comprobó que era una proteína, lo que era contrario a la 19 opinión general en aquel tiempo. Tuvieron que pasar 10 años, hasta 1926 que fue cuando Northrop cristalizó otras tres enzimas, para establecer finalmente que éstas son proteínas especializadas en una reacción. Hoy en día se conocen quizá más de 2 000 enzimas diferentes y, aunque se sabe mucho acerca de ellas, quedan aún grandes dudas sobre su control genético y su modo de acción. Figura 9. (a) Enzima con uno o dos sitios activos y una cadena de aminoácidos. (b) Enzima con varias cadenas de aminoácidos y varios sitios activos. Estas proteínas actúan sobre una sola molécula, es decir, son como una llave y una cerradura: la llave es la molécula y la cerradura es la enzima. La posibilidad de que una llave abra otra cerradura que no sea la que le corresponde es muy remota. Pues bien, lo mismo ocurre con las enzimas: la posibilidad de que una enzima actué sobre otra molécula que no sea la que le corresponde es muy baja. Las enzimas, en su calidad de proteínas, se hallan formadas por cadenas de aminoácidos, las cuales se arreglan espacialmente en formas variadas. En otras palabras, las cadenas de aminoácidos constituyen estructuras tridimensionales (Figura 8). Así, estas complicadas estructuras forman un espacio o hueco en donde se une la molécula que va a ser transformada; a este hueco se le denomina sitio activo. Las enzimas tienen entonces un sitio activo y una estructura tridimensional que las hace ser únicas. Sin embargo, pueden existir enzimas con dos o más sitios activos y con una estructura integrada por una o varias cadenas de aminoácidos (Figura 9). 20 La velocidad a la que dichas enzimas trabajan es muy alta y esto las hace muy eficientes. Su actividad está en función directa de la cantidad de moléculas que va a ser transformada y su acción es regulada en parte por la aparición en el medio del producto final de la reacción, que de hecho inhibe su actividad. Su acción puede ser concertada es decir, que una enzima se encuentra dentro de un proceso más o menos largo de degradación o de síntesis y de esta forma el producto de dicha enzima será el sustrato de la siguiente enzima y así sucesivamente (Figura 13). Figura 10. Simplificación de la relación que guardan los organismos autótrofos y heterótrofos. El metabolismo se podría definir como el conjunto de reacciones químicas que ocurren en la célula y que tienen por objeto generar la energía y los componentes necesarios para que ésta lleve a cabo sus funciones. Esta definición estaría incompleta si no aclaramos antes que todas estas reacciones están altamente coordinadas y dependen del perfecto funcionamiento de los sistemas multienzimáticos. II.2 UNOS SERES VIVOS DEPENDEN DE OTROS 21 Organismos autosuficientes y organismos dependientes. Para poder llevar a cabo todas sus reacciones metabólicas, los seres vivos necesitan una fuente de alimento y energía. De acuerdo con su fuente de obtención de nutrientes, los organismos vivos se pueden dividir en dos grandes grupos. Por una parte, los autosuficientes (autótrofos), que utilizan al bióxido de carbono (CO2) y al agua (H20) como única fuente de alimento y a partir de estos compuestos producen todas las moléculas necesarias para su subsistencia, crecimiento y proliferación. Por otra parte, los que dependen de otros compuestos o de sus productos, se llaman heterótrofos. Estos no pueden utilizar el bióxido de carbono como tal y por lo tanto deben obtener el carbono que necesitan a partir de otros compuestos que se encuentran en el medio que los rodea, como por ejemplo la glucosa, que es una molécula mucho más compleja que el C02. Lo cual quiere decir que unas formas de vida requieren de otras y así se establece una cadena alimenticia. Ya que unos organismos son relativamente autosuficientes y otros requieren de fuentes de carbono más complejas, su interrelación es necesaria y vital. Algunos ejemplos de células autosuficientes son las de las plantas, las bacterias fotosintéticas y algunas bacterias no fotosintéticas. Sin embargo, la mayoría de los organismos no son autosuficientes y por lo tanto requieren siempre de una asociación benéfica con un organismo que sí lo sea. El ciclo del carbono y del oxígeno. En la figura 10 podemos ver cómo se relacionan los organismos fotosintéticos y los que dependen de ellos. Los primeros, por medio de la energía luminosa del Sol, más el agua y el bióxido de carbono (C02) del suelo, llevan a cabo la fotosíntesis, produciendo así glucosa y oxígeno. A partir de estos compuestos básicos que se convierten en materia orgánica, los organismos dependientes obtienen a su vez los nutrientes que, al ser utilizados, son transformados y liberados al medio en forma de agua y C02. Estas moléculas a su vez serán nuevamente utilizadas por los organismos fotosínteticos. Así pues, la interrelación entre ambos tipos de organismos crea a su vez un ciclo importante para la conservación de la vida en la Tierra. A éste se le conoce como ciclo del carbono y del oxígeno. El ciclo del nitrógeno. El nitrógeno es sin duda un elemento muy importante para el metabolismo, ya que forma parte de las proteínas, los ácidos nucleicos y de muchas moléculas de gran importancia biológica. De manera similar a lo que ocurre con el carbono, el nitrógeno forma parte de un ciclo que hace a los diferentes tipos de organismos vivos depender entre sí. Esto se debe básicamente a que, a pesar de que el nitrógeno se encuentra distribuido en cantidades enormes en la atmósfera, es un elemento poco reactivo y debido a eso no puede ser aprovechado por la mayoría de las formas de vida. El nitrógeno debe ser asimilado por las plantas bajo la forma de nitrato o amoniaco, o bien por los animales bajo la forma de animoácidos, los cuales forman parte estructural de las proteínas. 22 La asimilación del nitrógeno en los ciclos de vida depende de varios factores. Las plantas toman el nitrógeno de la tierra en la forma de nitrato y lo utilizan para llevar a cabo las funciones metabólicas que dependen de este elemento. Por otra parte, los organismos dependientes utilizan las proteínas vegetales como nutrientes para aprovechar el nitrógeno contenido en éstas y lo devuelven a la tierra en forma de amoniaco como producto de desecho, o bien como producto de descomposición después de su muerte como se muestra en la figura 11. En la tierra, una especie de bacterias transforma al amoniaco en nitrato, el cual puede nuevamente ser utilizado por las plantas. Figura 11. El ciclo del nitrógeno. Este ciclo mantiene el balance entre las dos vastas reservas de compuestos nitrogenados: la atmósfera y la corteza terrestres. Existen, sin embargo, algunas especies de bacterias muy peculiares, conocidas como fijadoras de nitrógeno, que son capaces de procesar el nitrógeno atmosférico que por lo general no se asimila. Estas bacterias lo transforman en un producto útil para los organismos que dependen de él, como las plantas. Las bacterias fijadoras de nitrógeno atmosférico se asocian íntimamente con las raíces de algunas plantas. De esta forma, algunas, como el frijol, no requieren de ningún tipo de abono o fertilizante, ya que su asociación con la bacteria les permite utilizar en forma directa el nitrógeno atmosférico. Este tipo de simbiosis se antoja atractiva para que otras plantas, que requieren de fertilizantes, sean capaces también de asociarse con estas bacterias. Un gran número de 23 investigadores en México y otras partes del mundo se encuentra interesado en este problema tan importante para la agronomía. Así, se están estudiando las posibilidades de establecer este tipo de interacción bacteria-planta en plantas que normalmente no lo hacen. Hasta la fecha sólo se conoce un tipo de microorganismo que sea totalmente autosuficiente y que no dependa de ningún otro ser vivo, ya que utiliza la luz del Sol como fuente de energía, obtiene el carbono a partir del bióxido de carbono y el nitrógeno de la atmósfera. Estos organismos son las algas verdeazules, las cuales, se piensa, fueron los primeros pobladores de la Tierra cuando se establecieron las condiciones climáticas adecuadas. II.3 LA ACTIVIDAD METABÓLICA AUMENTA O DISMINUYE El metabolismo en los seres vivos es muy flexible y se puede ajustar a la cantidad y tipo de nutrientes disponibles en el medio que rodea a un microorganismo. Un ejemplo lo podemos obtener de las levaduras, las cuales se adaptan a las condiciones en que se les mantenga, pues poseen todo el conjunto de enzimas necesario para sintetizar sus elementos básicos como los aminoácidos y todos los demás componentes celulares, de tal manera que si crecen en un medio con glucosa como única fuente de energía, pueden sintetizar todas las moléculas que necesitan, aunque esto implica un gran gasto en energía. La levadura puede evitarlo si le proporcionamos los aminoácidos y nutrientes requeridos, ya que economiza el aparato enzimático responsable de la síntesis de todos los compuestos que necesita para vivir. Actualmente es posible tener en el laboratorio cultivos controlados de bacterias en los cuales el investigador modifica a voluntad la composición de los nutrientes. Esto ha permitido conocer y entender cómo los microorganismos modifican su metabolismo en función de la calidad nutritiva del medio en el cual se les hace crecer. Si, por ejemplo, la concentración de nutrientes ricos en nitrógeno baja, el microorganismo iniciará inmediatamente la síntesis de las enzimas necesarias para elaborar sus propios ácidos nucleicos y aminoácidos; todo está finalmente regulado y enfocado hacia la máxima economía metabólica y, en última instancia, al ahorro de energía. II.4 PARTES DEL METABOLISMO El metabolismo se puede dividir en dos grandes partes: el catabolismo y el anabolismo. La fase del metabolismo que descompone las moléculas grandes en pequeñas es el catabolismo. Las moléculas grandes como las proteínas, las grasas (lípidos) o los azúcares (carbohidratos), que provienen de nutrientes del medio ambiente, se descomponen enzimáticamente. El producto de esta descomposición o degradación lo forman las moléculas más simples y pequeñas como, 24 por ejemplo, el ácido láctico, el ácido acético, el bióxido de carbono, el amoniaco y la urea. De estas reacciones químicas de degradación se obtiene la energía química contenida en las estructuras de las grandes moléculas. Esta energía se conserva en forma de molécula conocida como adenosíntrifosfato (ATP), la cual es de vital importancia en el metabolismo de cualquier organismo vivo. Por otra parte, el anabolismo es la fase del metabolismo durante la cual se sintetizan de nuevo las moléculas que la bacteria o célula utiliza para regenerarse, mantenerse o dividirse, como son: las grasas, las proteínas, los azúcares o carbohidratos y los ácidos nucleicos (ADN y ARN), que forman parte funcional o estructural de los organismos. Esto lo lleva a cabo el microorganismo o la célula a partir de los constituyentes primarios que se obtienen de los nutrientes. Sin embargo, tales procesos de síntesis requieren de energía y ésta la proporciona el ATP que fue generado durante el catabolismo. Así, el anabolismo y el catabolismo se llevan a cabo simultáneamente y cada uno está regulado en forma muy precisa, ya que ambos procesos son interdependientes (Figura 12). Las enzimas que llevan a cabo las diferentes reacciones en el metabolismo funcionan en forma secuencial, lo que quiere decir que el producto de una reacción enzimática será a su vez degradado por la siguiente enzima de la cadena de reacciones de alguna vía metabólica, proceso al que se conoce como de sistemas multienzimáticos. La razón de este tipo secuencial de reacciones se debe, aparentemente, a los múltiples puntos de control que ofrece, ya que un solo paso para la conversión de una sustancia presentaría serios riesgos para un organismo; probablemente por esta razón tal estrategia no ha sido adoptada por la naturaleza. 25 Figura 12. El anabolismo y el catabolismo. Todos los procesos que ocurren en la célula o bacteria requieren de energía. Esta energía está almacenada como moléculas de ATP, que se forma a partir de ADP y fosfato inorgánico. La degradación de los nutrientes: Las proteínas, los lípidos y los carbohidratos son degradados mediante reacciones enzimáticas que ocurren una después de la otra. Este proceso de degradación está dividido en tres etapas que a grandes rasgos son las siguientes: La primera consiste en la conversión de moléculas complejas a moléculas más simples; esto quiere decir que una proteína, que no es más que una cadena de aminoácidos, tiene que ser convertida de nuevo en sus constituyentes, que son los aminoácidos, para que éstos puedan ser debidamente aprovechados en la segunda etapa de degradación. Estos procesos degradativos no requieren de energía y ocurren en el interior del microorganismo. La segunda etapa consiste en la conversión de estas moléculas, ya bastante simples, en una aún más simple y común, independientemente del origen de las moléculas. Es decir, que tanto los carbohidratos como las proteínas o los lípidos son convertidos en una molécula mucho más simple llamada "acetil coenzima A". Es a partir de esta molécula que la tercera etapa del metabolismo se lleva a cabo y consiste en generar la energía que necesita la célula para realizar procesos vitales, como desplazarse o dividirse, entre otros. En esta etapa ocurre uno de los ciclos metabólicos más importantes de la biología: el ciclo de los ácidos tricarboxílicos o ciclo de Krebs. Así pues, todas las moléculas que sirven a un organismo como fuente de subsistencia son llevadas, por medio de diversos caminos de degradación, a un camino metabólico común. Esta vía metabólica común degrada una sola molécula en una serie de pasos y como resultado se obtiene energía principalmente en forma de ATP. Los caminos metabólicos están finamente controlados. Estos mecanismos de control consisten en que los niveles de algunos productos regulan la actividad de algunas enzimas, de tal forma que su actividad se incrementa o disminuye dependiendo de los niveles del producto. Por otra parte, las concentraciones de ATP, que como ya vimos es una molécula muy importante en el metabolismo de la célula, regulan también la actividad de ciertas enzimas y esto lo hacen por medio de la unión del ATP a las enzimas susceptibles de ser reguladas por esta molécula. La regulación del metabolismo es vital, y de ésta depende que un un organismo produzca solamente la cantidad necesaria de cada una de las moléculas que requiere para subsistir y por lo tanto que el desperdicio de energía sea mínimo. 26 Tanto las bacterias como las células de los seres superiores generan la energía necesaria durante el catabolismo y la almacenan en forma de ATP. Así, éstas pueden realizar funciones vitales como el movimiento, el transporte de nutrientes a su interior y la síntesis de las moléculas que forman parte de su estructura o que tienen funciones específicas y deben ser sintetizadas en el interior. La síntesis de las moléculas es continua e implica un recambio constante entre las moléculas que se degradan y las que se sintetizan. De hecho, los procesos de degradación se conocen, aunque las señales que los gobiernan son aún tema de intensas investigaciones. El estudio del metabolismo se ha apoyado en el uso de microorganismos a los cuales se les induce un cambio genético o mutación. Este tipo de enfoque ha permitido entender la mayoría de las vías metabólicas. Así, por ejemplo, el hongo Neurospora crassa puede crecer en un medio simple que contenga glucosa como única fuente de carbono y amoniaco como fuente de nitrógeno. Sin embargo, si se expone este hongo a rayos X, se obtiene una neurospora que ya no crece en el medio simple. Esta mutante solamente puede crecer en un medio de cultivo al cual se le ha añadido el compuesto que ya no sintetiza dicho microorganismo. Un ejemplo de esto son las mutantes del hongo que ya no crecen a menos que se añada al medio un aminoácido conocido como arginina (como todos los aminoácidos contiene nitrógeno). Lo cual quiere decir que la síntesis de este aminoácido está alterada y el hongo, por lo tanto, no puede utilizar el nitrógeno del medio para sintetizarlo. Esta mutante no crecerá a menos que se le adicione dicho aminoácido en el medio de cultivo. Existen toda otra serie de mutantes similares que difieren en los pasos en que el metabolismo se encuentra alterado; así se han podido conocer las diferentes etapas que forman parte de una vía metabólica. II.5 LA MAQUINARIA DEL METABOLISMO El metabolismo es, entonces, un sistema complejo de reacciones químicas llevadas a cabo por las enzimas que son las responsables de la transformación de los nutrientes en moléculas útiles para la bacteria. 27 Figura 13. Sistemas multienzimáticos. Estos pueden ser tanto solubles como membranales, de su disposición depende su función. La complejidad de los sistemas multienzimáticos que lleva a cabo el metabolismo es variable. En algunos casos las enzimas se encuentran en el interior de la célula y las moléculas que van a ser degradadas interaccionan libremente. Estos sistemas funcionan con base en la difusión libre de los productos del metabolismo (metabolitos) en el interior de la bacteria o célula. En otras palabras, la probabilidad de que una molécula interaccione con la enzima adecuada es alta y la reacción ocurre por lo tanto a una gran velocidad. Por otra parte, existen sistemas multienzimáticos organizados espacialmente en la membrana celular; esto permite que las reacciones ocurran de una manera organizada, funcionando juntos como una perfecta maquinaria engranada. Este tipo de disposición de las enzimas en una membrana puede llegar a ser muy complicada. De hecho, su funcionamiento es aún un problema biológico que atrae la atención de varios grupos de científicos en el 28 mundo. Estos complejos enzimáticos altamente organizados forman parte indispensable de los organismos vivos desde las bacterias hasta los mamíferos. La interacción íntima entre sus componentes facilita la difusión de los metabolitos, y por lo tanto la velocidad a la que ocurren las reacciones es también muy alta. La figura 13 ilustra los tipos de sistemas enzimáticos y su disposición en el interior de la bacteria o de su membrana. El metabolismo puede tener dirección y tamaño. Como ya vimos, existen sistemas multienzimáticos asociados a una membrana celular. Estos sistema generalmente aprovechan la división entre uno y otro lado de la membrana, es decir entre el interior y el exterior de la bacteria. Esta disposición permite dividir un compartimiento de otro, ya que las membranas celulares son impermeables. La idea de que el metabolismo en compartimientos puede tener dirección y tamaño parece complicado o sin sentido, y así fue como lo tomaron los bioquímicos en los años sesenta. En 1961, Peter Mitchell, un microbiólogo inglés, propuso por primera vez que la síntesis de la molécula conocida como ATP, se debía a que ciertas reacciones metabólicas tenían como resultado el movimiento de una especie química cargada o ión, en este caso un protón (H+). Este protón, entonces, era el responsable de que el ATP fuera sintetizado. Figura 14. Algunas reacciones metabólicas tienen dirección y magnitud; estas reacciones se llevan a cabo en una membrana que funciona como barrera y que permite acumular una especie química, que al equilibrarse genera trabajo o energía. La forma como funciona el complejo enzimático que sintetiza el ATP puede expresarse así: logra que un protón que se encuentra en el 29 interior sea sacado al exterior. La figura 14 muestra en forma esquemática este proceso. Sumergidas en la membrana de la bacteria se encuentran las enzimas que componen la cadena respiratoria (así se les conoce, ya que al funcionar consumen oxígeno). La cadena respiratoria utiliza los compuestos que se producen durante el catabolismo en el ciclo de Krebs; éstos tienen la propiedad de ceder un electrón que con un protón formará un hidrógeno que ya no tiene carga. Este hidrógeno es transportado al lado opuesto de la membrana en donde pierde nuevamente el electrón y se libera el protón resultante al medio. Así, al cabo de varios ciclos se logra acumular una gran cantidad de protones en el exterior de la bacteria y esto tiene como consecuencia que se establezca una diferencia en la concentración de protones, que tiene la tendencia natural a equilibrarse. Un ejemplo podría ser la situación imaginaria de que un grupo grande de personas se encuentre en un pequeño cuarto comunicado con otro por una puerta que sólo se mueve en el sentido del cuarto vacío. Al estar tan apretadas empujarán dicha puerta y se empezará a llenar el cuarto vacío, y pasado un tiempo el número de personas será igual en ambos cuartos. Volviendo a la membrana de una bacteria, lo que ocurre es que la energía que se libera cuando se reequilibria la concentración de los protones en ambos lados de la membrana, permite que se sintetice el ATP. Lo anterior, que hoy en día se ve claro, fue muy difícil de aceptar por la comunidad científica de la época y pasaron 16 años para que esta teoría fuera completamente aceptada. Su aceptación culminó con la entrega del premio Nobel a Peter Mitchell, ya que su teoría permitió explicar una gran cantidad de fenómenos hasta entonces oscuros. Por ejemplo, si estas reacciones con dirección producen una diferencia en la concentración de protones se tendrá también una diferencia de cargas (+) (afuera una gran carga positiva). Esto puede ser usado por algunas especies químicas para entrar, por ejemplo, si su carga es positiva (A+), o para salir si es negativa (B). Como estos procesos metabólicos tienen dirección, magnitud y sentido, se dice que son "vectoriales"; además, son muy complicados y todavía no los comprendemos en su totalidad, pero baste decir que constituyen un pilar fundamental de los procesos energéticos celulares, ya que explican reacciones vitales como la síntesis del ATP o el transporte de nutrientes. Hasta ahora hemos visto a las bacterias que necesitan obtener su energía a partir de los nutrientes que se encuentran en el medio. Sin embargo, otro grupo de bacterias muy importantes son aquéllas cuya fuente principal de energía es la luz que proviene del Sol y de ellas nos ocuparemos a continuación. II.6 BACTERIAS FOTOSINTÉTICAS Menos familiares para nosotros, pero igualmente importantes, son las bacterias que tienen la facultad de utilizar la luz como fuente de energía. Éstas desempeñan un papel fundamental en la ecología y han servido para el estudio del proceso molecular de la fotosíntesis 30 en numerosos laboratorios. Cuando se toman muestras de las profundidades de un lago, cualquiera se sorprendería al notar que el agua huele muy mal; además, hay que agregar el color verdoso o rojizo del agua. Si la analizamos con más detalle, veremos que hay producción de gas y que no existe oxígeno. Este ambiente tan hostil alberga a las poco conocidas bacterias fotosintéticas, responsables de una gran parte de la producción de materia orgánica en los lagos y parte importante de la cadena biológica. Estos organismos son muy antiguos y se piensa que son los descendientes directos de las primeras células fotosintéticas que existieron en la Tierra, entre las que se pueden contar las algas verdeazules y las bacterias fotosintéticas verdes y púrpuras. Las algas verdeazules se pueden encontrar como células únicas o en colonias en la tierra, los ríos, los lagos y los océanos. Estos organismos pueden vivir utilizando CO2como única fuente de carbono y algunos son capaces de fijar el nitrógeno atmosférico. Las bacterias fotosintéticas verdes que producen azufre (sulfurosas), como Chlorobium, viven en ambientes carentes de oxígeno, como en el fondo de algunos lagos ricos en materia orgánica. Estos organismos le dan el color verde característico a estas aguas. Así encontramos también bacterias púrpuras sulfurosas, como Chromatium, que viven en ambientes también carentes de oxígeno y que utilizan compuestos con azufre en su metabolismo. Éstas se encuentran también en algunos lagos y en manantiales de aguas termales sulfurosas. Finalmente tenemos a las bacterias púrpuras no sulfurosas, que tienen la peculiaridad de utilizar la luz en ausencia de oxígeno, como las bacterias verdes y las púrpuras sulfurosas, pero además pueden utilizar al oxígeno durante los periodos de oscuridad. Estas bacterias poseen en su interior una complicada organización de moléculas encargadas de captar la luz solar y conservar la energía proporcionada por esta forma de energía química. La luz que es aprovechada es la luz visible y gran parte de la cual es empleada también por las plantas verdes, de tal forma que los pigmentos encargados de captar la luz lo hacen en las regiones del espectro de la luz visible que no son utilizadas por las plantas. Estas moléculas se conocen como pigmentos antena y, dado que la intensidad de la luz que llega a las profundidades donde se encuentran estas bacterias es muy baja, su captación debe ser muy eficiente para así aprovechar la energía al máximo. Estos microorganismos cuentan con estructuras internas que contienen las moléculas que captan la luz de tan baja intensidad. Estas estructuras se conocen como cromatóforos y contienen moléculas de clorofila típica de estas bacterias, que son las que captan la luz y retienen la energía que ésta proporciona (Figura 15). Las bacterias fotosintéticas realizan una parte muy importante de la transformación y generación de la materia orgánica de lagos y ríos. Su existencia es determinante en el frágil equilibrio energético de este planeta. Estudiarlas es de gran importancia. En México existen 31 ya algunos grupos interesados en estos microorganismos y en sus mecanismos de transformación y conservación de la energía solar. La fotosíntesis. Éste es sin duda uno de los procesos más importantes de captura de energía en nuestro planeta y lo podemos definir como la forma de utilización de energía solar que han desarrollado algunos microorganismos fotosintéticos y las plantas, mediante la cual llevan a cabo la síntesis de todos sus componentes celulares. Este proceso es fundamental para todos los organismos vivos, ya que es la piedra angular del ciclo de utilización de la materia en la Tierra. La luz solar es la fuente primordial de energía en la Tierra, tanto para los organismos fotosintéticos como para casi todos los organismos heterótrofos. Se utiliza a través de una complicada serie de procesos que van desde las cadenas alimenticias hasta complejos procesos industriales. Por ejemplo, el hombre utiliza la energía generada por la fotosíntesis de organismos que existieron hace millones de años y cuyos productos de descomposición se emplean en forma de gasolinas y derivados del petróleo, que proporcionan los combustibles para casi todas las máquinas construidas por el hombre. Figura 15. Micrografía electrónica de una bacteria fotosintética no sulfurosa, Rhodospirillum rubrum. En su interior se pueden observar pequeñas estructuras llamadas cromatóforos. El descubrimiento de la fotosíntesis. El desarrollo histórico de la fotosíntesis se inicia entre 1770 y 1777 cuando Joseph Priestley, uno de los descubridores del oxígeno, llevó a cabo importantes 32 experimentos sobre el intercambio de materia que ocurre durante la fotosíntesis. El experimento de Priestley consistió básicamente en consumir el oxígeno de un recipiente cerrado, lo que logró con una vela que ardía en el interior. Al consumirse el oxígeno, observó que la vela se apagaba. Por otra parte, si en dicho recipiente con aire se introducía además de la vela un ratón, al poco tiempo de consumirse el oxígeno de la cámara el animal moría. Ahora bien, si dentro del recipiente se incluía una planta, tanto la vela como el ratón se mantenían sin problema alguno. Priestley concluyó que las plantas verdes llevan a cabo la producción del oxígeno. Al parecer, este proceso era la inversa de la respiración en los animales, en la cual se consume oxígeno. A pesar de tan acertados experimentos, Priestley no logró determinar que este proceso estaba ligado a la luz. Fue Ingenhousz, un médico holandés, quien lo descubrió algunos años después, identificando que la porción verde de las plantas era la responsable de la producción del oxígeno. Posteriormente, en el siglo XIX se concretó otra serie de importantes descubrimientos que condujeron finalmente a dilucidar que las responsables de la fotosíntesis en las plantas son las hojas y que hay producción de materia orgánica durante este proceso que se puede esquematizar como sigue: 6CO2 + 6H2O + energía luminosa - C6H12O6 (glucosa) + 6 O2 La capacidad de llevar a cabo el proceso de fotosíntesis se encuentra en una gran variedad de organismos, tanto en bacterias como en plantas y algas. I I I . L A S B A C T E R I A S Y L A H I S T O R I A D E L A T I E R R A EN LA Antigüedad los sabios sostenían que la vida en la Tierra consistía fundamentalmente en dos formas de vida: las plantas y los animales. Más adelante, cuando los microbios fueron descubiertos, la división se hizo de una forma similar; los organismos más grandes y que se movían se consideraban como animales, y los que aparentemente no se movían, incluyendo a las bacterias, se consideraban como plantas. Conforme avanzó el conocimiento sobre el mundo microscópico, se llegó a la conclusión de que la clasificación original era insuficiente y se propusieron categorías adicionales tales como hongos, protozoarios y bacterias. Sin embargo, posteriormente una nueva simplificación se llevó a cabo, ya que se pensó que los seres vivos podrían ser divididos en dos nuevamente, sólo que esta división sería a un nivel más profundo, es decir, sobre la estructura de la célula viviente. Aparentemente todas las células vistas en un microscopio pertenecían a una de dos categorías: las que tienen un núcleo bien definido (eucariontes o eucariótico, que significa de núcleo 33 verdadero) y las células sin núcleo (procarionte, que quiere decir literalmente antes del núcleo, figura 6) Las plantas y los animales multicelulares, lo mismo que varios organismos unicelulares y microscópicos como las levaduras y los protozoarios, tienen un núcleo bien definido y son eucariontes. Por otro lado están las bacterias que no tienen núcleo. Hasta aquí no parecía haber dificultad alguna en la clasificación, sin embargo, muy recientemente un grupo de investigadores ha revisado la clasificación que hasta ahora prevalecía y llegó a interesantes conclusiones: se encontró que entre las bacterias existe un grupo de organismos que no parece estar relacionado ni con los organismos superiores ni con los más simples. Estos nuevos organismos tampoco contienen núcleo, como las bacterias, y se parecen mucho a éstas en su morfología vistas en un microscopio. Sin embargo, en su composición química y en la estructura de algunos de sus componentes moleculares son tan distintos de las bacterias (procariontes) como lo son de los organismos superiores (eucariontes). Estos microbios forman por sí mismos un nuevo grupo, que se caracteriza por tener una forma de vida completamente diferente a lo conocido hasta ahora, al que se ha dado el nombre de arqueobacterias (arqueo: viejo o primitivo) y este curioso nombre hace evidentes algunas conjeturas sobre su origen aún no comprobadas. Hay indicios de que este grupo de organismos es por lo menos tan antiguo como los otros dos, pero como algunas clases de arqueobacterias tienen formas de metabolismo que se adecuan bien a las condiciones climáticas que prevalecían probablemente en los orígenes de la historia de la Tierra, posiblemente las arqueobacterias son el grupo más antiguo de los tres. III.2 BACTERIAS FÓSILES Un hecho muy importante es que no solamente los animales y las plantas han dejado restos de su existencia como fósiles, sino que las bacterias, aunque pequeñas, también imprimieron la huella de su pasado en la Tierra, ya que, como vimos, estos pequeños organismos son capaces de modificar el medio ambiente. Los microfósiles, es decir los fósiles de las bacterias, se encuentran en sedimentos de todas las edades geológicas e incluso en las rocas sedimentarias que tienen 3 500 millones de años y que son las más antiguas que se conocen. Las bacterias o microbios existieron en un periodo de la historia de la Tierra en el que no había otras formas de vida. La Era de las bacterias fue muy importante, ya que en ella se produjo toda una serie de eventos evolutivos y geológicos. Hasta hace muy poco no se sabía gran cosa sobre la Era de los microorganismos y, por otra parte, los microfósiles aislados no dan mucha información. Sin embargo, se han descubierto otras estructuras fósiles de bacterias que forman aglomerados llamados estromatolitos y que son aparentemente colonias de bacterias mezcladas con minerales. Hoy en día se sabe que tales tipos de estructuras constituyen efectivamente aglomerados peculiares de 34 bacterias que quizá eran fotosintéticas. Se piensa que es así porque los estromatolitos de bacterias fotosintéticas que se han fosilizado recientemente se asemejan a los antiguos, a tal grado que es razonable pensar que las estructuras antiguas también fueron formadas por bacterias fotosintéticas. Ésta es la única evidencia que se tiene sobre el origen de la evolución de las bacterias, el cual es tan oscuro como lo es el origen de las formas de vida superiores (Figura 16). Figura 16. ¿Cuál es el antecesor común de todas las formas de vida? Ésta es sin duda una incógnita de gran importancia, ya que dentro de este esquema seforma el tronco común, de donde emergieron los organismos superiores primitivos (eucariontes), las bacterias y las arqueobacterias. III.3 LA BIOQUÍMICA Y LA EVOLUCIÓN La biología se ha servido de la bioquímica para tratar de descifrar el pasado de una célula o bacteria. Para tal fin se han utilizado 35 moléculas propias de la célula como lo son las proteínas y los ácidos nucleicos (ácido ribonucleico, ARN, y ácido desoxirribonucleico, ADN). La idea en que se apoya este proceso se basa en que los organismos vivientes son mucho más ricos en información que los fósiles, información que se extiende más allá de los fósiles más antiguos. Sin embargo, para poder descifrar los antecedentes biológicos ha sido necesario desarrollar la tecnología para determinar la estructura del ácido desoxirribonucleico (ADN) que conforma un gene, la del ARN y la del producto de un gene, como son las proteínas. Para el caso de las proteínas (del griego proteios, primero), desde hace aproximadamente 25 años se llevan a cabo las determinaciones de las secuencias de aminoácidos que constituyen a varias de ellas. Sin embargo, la tecnología para la determinación de la secuencia de bases de ADN o ARN que forman los ácidos nucleicos ha sido perfeccionada muy recientemente (premio Nobel de Química 1980). Esta nueva tecnología permitirá sin duda descubrir rápidamente más acerca de la historia de la vida sobre la Tierra. Ha sido con estas técnicas, que un grupo de científicos ha logrado reconocer recientemente a las arqueobacterias como una tercera forma de vida. Para apreciar la verdadera diferencia entre las arqueobacterias y las otras dos formas de vida, debemos recordar que se han definido criterios para distinguirlas. Recordemos que las células de los organismos superiores son relativamente grandes y están rodeadas por una membrana dentro de la cual se encuentra una serie de estructuras que a su vez están rodeadas de otras membranas. Estas estructuras subcelulares se llaman organelos. Por otra parte, las bacterias son muy distintas. En primer lugar, son más pequeñas, y en segundo lugar, ninguna de las estructuras que mencionamos antes se encuentra en ellas. Las bacterias están rodeadas por una sola membrana y en la mayoría de los casos están rodeadas, además, de una pared celular rígida. Toda la información genética de estos organismos está contenida en unos 2 000 a 3 000 genes, información que es muy pequeña comparada con la contenida en la célula de un animal superior. La cantidad de información genética en una célula eucarionte es varios cientos de miles de veces mayor que la de una bacteria. La distinción entre seres superiores y bacterias se definió inicialmente en términos de las pequeñas estructuras subcelulares que se podían observar con un microscopio de luz. A ese nivel de definición, las células parecían grandes y complejas o pequeñas y simples. Sin embargo, esta distinción se ha tratado de llevar a niveles biológicos básicos, o sea a las moléculas que conforman dicha célula. Ambos tipos de célula tienen procesos bioquímicos comunes, como, por ejemplo, los mecanismos por medio de los cuales se transforma la información genética que contiene el ADN en proteínas; sin 36 embargo, algunos detalles sobre los mecanismos de estos procesos son, o bien típicamente de los organismos superiores, o bien típicamente bacterianos. Estas diferencias y similitudes dieron la pauta para que pareciera evidente que el árbol de la vida tenía sólo dos ramas, la de las bacterias y la de los organismos superiores. Recientemente se han acumulado una serie de evidencias que indican que la relación evolutiva entre ambos reinos es más complicada de lo que se pensaba. Por ejemplo, dos organelos subcelulares, como las mitocondrias y los cloroplastos, contienen su propio ADN, aparte del que se encuentra en el núcleo. Ambos tipos de organelos subcelulares son del tamaño de una bacteria y el aparato que contienen, destinado al procesamiento de material genético, es muy similar al de las bacterias. Estas evidencias, entre otras, han llevado a pensar que tanto las mitocondrias como los cloroplastos descienden de los procariontes, y que quedaron atrapados dentro de una célula donde se desarrollaron, según propone la teoría endosimbiótica de la evolución (endo=interior, simbiosis=convivencia o relación). Esta teoría propone que la mitocondria era una bacteria respiratoria y el cloroplasto una bacteria fotosintética. Esta conjetura, que prevaleció durante más de cien años, fue comprobada para el caso de los cloroplastos y se logró comparando las secuencias de bases de una molécula de ARN (ácido ribonucleico), que resultaron estar íntimamente relacionadas con las de las bacterias fotosintéticas. Esto implica que por lo menos dos líneas descendientes de las bacterias están representadas en la célula de los organismos superiores. III.4 UN MODO DE VIDA DIFERENTE Con estos antecedentes podemos regresar al tema de las arqueobacterias. Cabe aclarar que hoy en día se conoce poco sobre las leyes biológicas que gobiernan a estos peculiares microbios y que se trata de un campo en pleno desarrollo. El grupo de las arqueobacterias incluye tres tipos de bacterias: metanogénicas, las que producen metano; halófilas extremas, las que viven en medios salinos extremos, y termoacidófilas, las que subsisten en ambientes calientes y ácidos. De los tres tipos las que predominan son las metanogénicas. En 1776 Alessandro Volta descubrió la existencia de lo que él llamó aire combustible, que se forma en las aguas estancadas de riachuelos y lagos, que tienen gran cantidad de sedimentos ricos en vegetación en proceso de descomposición. Sin embargo, el hecho de que un microorganismo fuera el responsable de la producción de metano (el gas combustible), se descubrió mucho más tarde. Las bacterias metanogénicas están ampliamente distribuidas en la naturaleza en sitios carentes de oxígeno y por esta razón es imposible encontrarlas a cielo abierto, ya que el oxígeno es altamente tóxico para estos organismos. 37 Es posible que las bacterias metanogénicas existieran en casi cualquier sitio cuando la Tierra era aún joven, ya que probablemente las condiciones atmosféricas eran más adecuadas para ellas. Hoy sólo se encuentran en sitios donde el oxígeno está ausente, y presente el hidrógeno y el bióxido de carbono (C02). También es frecuente encontrar a estas bacterias asociadas a otras como las del género Clostridium, que metabolizan la materia orgánica en descomposición y liberan al medio hidrógeno como producto de desecho. Este tipo de arqueobacterias, como ya mencionábamos, se encuentra en lugares donde hay agua estancada en putrefacción o en las plantas para el tratamiento de aguas negras. En la actualidad, el hombre ya las está aprovechando y se han podido obtener volúmenes suficientes de gas metano para ser utilizados industrialmente como combustible, hecho que atrae la atención de muchos científicos, ya que propone una alternativa al agotamiento de los energéticos no renovables. Las metanógenas se encuentran también en uno de los estómagos de los rumiantes, en el que se degrada la celulosa, y en el tracto digestivo de la mayoría de los animales. También se pueden obtener del fondo de los océanos o de los manantiales de aguas termales, lo que demuestra que, a pesar de su intolerancia al oxígeno, se encuentran ampliamente distribuidas sobre la Tierra. Actualmente estas bacterias se mantienen, en condiciones adecuadas, en varios laboratorios del mundo y son utilizadas en las investigaciones sobre sus procesos metabólicos. Otro tipo de arqueobacterias son las halófilas extremas, que requieren concentraciones muy altas de sal para vivir. Su localización más común es en aguas que contienen cantidades saturantes de cloruro de sodio (sal). Se encuentran más comúnmente a lo largo de las costas y en aguas saturadas de sal como los grandes lagos salados o el Mar Muerto. Las halófilas extremas dan un color rojizo a la sal en los pozos de evaporación y son capaces de descomponer el pescado salado. Estas bacterias han llamado la atención de los microbiólogos básicamente por dos razones. La primera es que presentan mecanismos para mantener diferencias enormes en la concentración de iones, como el sodio o el cloro, entre el interior y el exterior de la célula, y utilizan estas diferencias de concentración para el transporte de sustancias hacia el interior o el exterior de la célula. En segundo lugar, estas bacterias tienen un sistema fotosintético relativamente simple, ya que no se basa en la presencia de clorofila como en la molécula que capta la energía luminosa, sino en un pigmento presente en su membrana, llamado bacteriorrodopsina, que es muy similar a uno de los pigmentos que se encuentran en la retina del ojo. Dicho pigmento ha sido muy estudiado y actualmente se conocen su estructura y su función con gran detalle, lo que ha ayudado enormemente al avance del 38 conocimiento de las proteínas membranales y sobre todo de aquellas proteínas que transforman la luz en energía química. El tercer tipo de arqueobacteria que se conoce es el de las termoacidófilas, que también se caracterizan por habitar en un nicho ecológico peculiar. Sulfolobus, uno de los dos géneros de termoacidófilas, se encuentra en los manantiales de aguas sulfurosas, y crece y se desarrolla generalmente a temperaturas de aproximadamente 80°C, y algunas variedades lo hacen inclusive a temperaturas de 90°C. Otra característica particular de Sulfolobus es que el pH óptimo (la escala de pH se puede dividir en dos partes: 7.5 a 14, alcalino y de 7.5 a 0, ácido) para su crecimiento es muy ácido (generalmente pH = 2.0), si tomamos en cuenta que normalmente el pH adecuado para la vida es de 7.5. Así, se ve obligada a mantener en su interior un pH (el potencial hidrógeno = pH se refiere a la concentración de protones H+ en una solución) de aproximadamente 7.5. Pero no solamente puede sobrevivir con estas diferencias tan grandes de pH, sino que las aprovecha para importar nutrientes a su interior (Figura 14). Por algún tiempo se pensó que estos microorganismos tan peculiares se habían adaptado a nichos ecológicos distintos y extremos, pero hoy en día se sabe que integran un nuevo grupo de microorganismos. El descubrimiento de un grupo nuevo de organismos es de gran importancia, ya que ayudará a revelar la historia del origen de la vida. Cuando solamente se conocían dos líneas a partir de las cuales se originarían todos los organismos vivientes, se hacía muy difícil la interpretación de las diferencias que las separan. Ha sido a partir del descubrimiento de una tercera línea de vida que la interpretación de las propiedades ancestrales y las recientemente adquiridas se ha hecho más equilibrada, ya que estas tres líneas de desarrollo son equidistantes entre sí. El descubrimiento de las arqueobacterias ha permitido el acercamiento a dos problemas de gran importancia. Uno de ellos es la naturaleza del antecesor común a todas las formas de vida y el otro se refiere a la incógnita sobre la evolución de la célula de los organismos superiores. I V . L O S M I C R O B I O S A L D E L H O M B R E S E R V I C I O Los microbios útiles para el hombre constituyen solamente una proporción muy pequeña de la amplia variedad de especies que existe en la naturaleza. El papel que desempeñan algunos de ellos en la fabricación de la cerveza, el pan y el vino fue descubierto en forma accidental hace mucho tiempo. Por ejemplo, la transformación que sufre el jugo de uva y la masa para hacer el 39 pan, hoy en día se sabe que se debe a las levaduras. Microorganismos bien conocidos también lo son las bacterias que acidifican la leche o los hongos microscópicos que le confieren su carácter tan particular a los quesos. A estos tres grupos de microorganismos hay que agregar los actinomicetos, que son hongos filamentosos que viven en la tierra y que se han utilizado para la producción de antibióticos a partir del decenio de los años 40 del presente siglo. Es difícil clasificar a los microorganismos en útiles y no útiles, o en buenos y malos, ya que todos participan en el reciclaje de las moléculas del mundo orgánico y también en la conservación de una parte de la ecología; de tal forma que los microbios no sólo son útiles sino indispensables para la vida como la conocemos actualmente. Algunos son nocivos para los animales o plantas pero, en proporción, son muy pocos. Existe un buen número de microorganismos que está siendo utilizado en la industria. Esto se debe a que producen un compuesto de alto valor que no puede ser obtenido de una manera tan sencilla o tan económica por las técnicas químicas usuales. En algunos otros casos, los microorganismos son cultivados por su valor intrínseco, como es el caso de la levadura de panadería. Sin embargo, en la mayoría de los casos la sustancia buscada es un producto de su metabolismo, como es el caso del alcohol o algún antibiótico, por ejemplo. Figura 17 (a) Los hongos y las levaduras son microorganismos que forman estructuras visibles y en ocasiones muy coloridas. Varios de estos microorganismos producen sustancias de interés 40 en la industria, como la cerveza, el sake y hasta colorantes biológicos Figura 17 (b) Hongos: 1. Penicillium chrysogenum 2. Monascus purpurea 3. Penicillium notatum 4. Aspergillus niger 5. Aspergillus oryzae. Levaduras: 1. Saccharomyces cerevisiae 2. Candida utilis 3. Aureobasidium pullulans 4. Trichosporon cutaneum 5. Saccharomycopsis capsularis 6. Saccharomycopsis lipolytica 7. Hanseniaspora guilliermondii 8. Hansenula capsulata 9. Saccharomyces carlsbergensis 10. Saccharomyces rouxii 11. Rhodotorula rubra 12. Phaffia rhodozyma 13. Cryptococcus laurentii 14. Metschnikowia pulcherrima 15. Rhodotorula pallida. IV.1 LA VIDA CON O SIN OXÍGENO Los microorganismos se pueden dividir en tres grupos dependiendo del ambiente que necesitan para vivir. El primer grupo lo forman los organismos aeróbicos estrictos, que son aquellos que no pueden tener actividad metabólica ni crecer más que en presencia de oxígeno. El segundo grupo reúne a los organismos anaeróbicos estrictos, que llevan a cabo su actividad metabólica y crecimiento en ausencia total de oxígeno atmosférico. El tercer grupo está constituido por los organismos aeróbicos facultativos, los cuales presentan actividad metabólica en presencia de oxígeno (respiración) y en ausencia de oxígeno (fermentación) dependiendo del medio en el que se encuentren. 41 De los microorganismos aeróbicos estrictos podemos mencionar a los estreptomicetos, que son hongos microscópicos productores de antibiótico. También son aeróbicos la mayoría de los hongos filamentosos, como el Penicillum notatum, productor de la penicilina. Los organismos anaeróbicos estrictos están representados por las bacterias del género Clostridium, como Clostridium botulinum, fuente de la toxina productora de la enfermedad conocida como botulismo. Otro ejemplo es la Salmonella tryphi, causante de muchas infecciones intestinales en el ser humano. Las levaduras industriales son microorganismos facultativos que pueden respirar o fermentar de acuerdo con las condiciones en que se les cultive. El metabolismo anaeróbico, como la fermentación, es menos eficiente que la respiración, ya que la primera no aprovecha toda la energía de las moléculas como los azúcares. Algunos productos, como por ejemplo el alcohol etílico, son excretados por la levadura como producto de desecho, ya que en ausencia de oxígeno este producto no puede ser aprovechado en su totalidad. Las vías bioquímicas de la fermentación que conducen a la formación de productos útiles son diversas. Estas vías son de dos tipos: las homofermentarias, que son las que conducen a un producto principal, y las heterofermentarias que dan dos o más productos. Así pues, algunos bacilos son homofermentarios y producen ácido láctico a partir de azúcares; por otra parte, existen otros bacilos lácticos heterofermentarios que producen ácido láctico, alcohol etílico y gas carbónico por diferentes vías metabólicas. Otro organismo heterofermentario es el Clostridium aceto butylicum, que transforma la glucosa en una mezcla de acetona, alcohol etílico, isopropanol y butanol. Por otra parte, el crecimiento aeróbico que llevan a cabo algunos microorganismos es más eficiente para la producción de biomasa (número de organismos obtenidos a partir de una cantidad dada de nutrientes), ya que estos organismos degradan completamente las moléculas nutritivas y les extraen el máximo de energía. Esto quiere decir que si la meta de la producción industrial es mejorar los sistemas de obtención de grandes cantidades de organismos, como es el caso de la levadura destinada a la panadería, es ventajoso trabajar con el organismo en condiciones aeróbicas ya que de esta forma se utilizan los sustratos al máximo por respiración y por lo tanto se obtendrán muchas más levaduras por cantidad de nutrientes. IV.2 LA MANIPULACIÓN GENÉTICA Una forma de obtención de compuestos orgánicos a partir de organismos aeróbicos es mediante la manipulación genética, esto quiere decir que se pueden modificar, a través de cambios o mutaciones de los genes, los mecanismos por medio de los cuales los organismos transforman los nutrientes en las miles de moléculas 42 que constituyen una célula viviente. En condiciones metabólicas normales, cada uno de los constituyentes necesarios para la célula se fabrica en cantidades justas y suficientes. Esta fabricación controlada se lleva a cabo gracias a una serie de mecanismos de regulación genética que detienen la fabricación de intermediarios y de productos finales de una vía metabólica dada. Los microbiólogos han seleccionado colonias o cepas de bacterias mutantes en las que esta regulación tan fina se encuentre alterada, de tal forma que convenga para ciertos fines. Por ejemplo, algunos metabolitos primarios, que son necesarios para el crecimiento celular, son producidos en cantidades necesarias para la fabricación de moléculas utilizadas por la célula. Una mutante interesante es Corynebacterium glutamicum, defectuosa en su mecanismo de regulación para la producción de lisina, un aminoácido necesario para la síntesis de algunas proteínas, capaz de producir 50 gramos de lisina por litro de medio de cultivo. Este tipo de metabolito es considerado como primario y la mutante lo produce en exceso. Generalmente, las bacterias mutantes se separan de las silvestres haciéndolas crecer en un medio de cultivo en donde solamente dichas mutantes sobreviven. Para ello se incluyen en el medio antibióticos que impiden el crecimiento de las silvestres y no afectan a las mutantes. Son considerados metabolitos secundarios aquellos que no se requieren en la biosíntesis celular. Algunos microorganismos los sintetizan en la fase tardía de su crecimiento por razones hasta el momento oscuras. Uno de los casos más conocidos son los antibióticos; estos metabolitos secundarios no desempeñan un papel directo en el metabolismo energético de la célula o en su crecimiento. Por estas razones se piensa que estas sustancias contribuyen a la sobrevivencia del organismo al inhibir el crecimiento de posibles competidores que podrían ocupar el mismo nicho ecológico. Los organismos que producen este tipo de metabolitos secundarios son sensibles a sus propios antibióticos durante la fase inicial de crecimiento, sin embargo, durante la fase en la cual secretan el antibiótico, son insensibles a su presencia. Otra clase de sustancias importantes para la industria son las enzimas. Hasta la fecha no ha sido posible sintetizar una enzima en el laboratorio y su aislamiento puede ser un trabajo largo y de bajo rendimiento. Por otra parte, los microorganismos dependen de las enzimas para la degradación de las moléculas complejas. Las bacterias solamente producen la cantidad necesaria de enzimas, de la misma forma que otros productos del metabolismo. Sin embargo, por medio de las técnicas de ingeniería genética se pueden seleccionar microorganismos mutantes que sobreproduzcan alguna enzima en particular. Un método para incrementar la síntesis de una enzima es el de inducción. Cada enzima tiene su huella genética en la secuencia de una molécula de ADN. A esta secuencia se le denomina gene estructural y, en una bacteria, se encuentra en el único cromosoma 43 que ésta posee. Los genes estructurales con la información para la síntesis de una enzima están normalmente inactivos en ausencia del sustrato para dicha enzima. Por lo tanto, se dice que está reprimida la producción de una enzima y solamente cuando se agrega el sustrato requerido, o un análogo, el gene estructural es activado y dicha enzima es sintetizada, proceso al que se denomina desrepresión o inducción, y a las enzimas que responden se les llama inducibles, para distinguirlas de las que no son afectadas por la presencia del sustrato y que se denominan constitutivas. En algunos casos el inductor puede ser el producto de una reacción enzimática. Un ejemplo de esta situación es la enzima glucamilasa del hongo Aspergillus niger, la cual se encarga de romper la cadena de azúcares que componen al almidón en moléculas de glucosa. Sin embargo, para inducir la síntesis de glucamilasa no es necesario que el almidón se encuentre presente en el medio, ya que algunos análogos que son malos como sustratos son potentes inductores de la enzima. Gracias a estos métodos, se ha podido sobreproducir sustancias de gran importancia para la industria y para la medicina. IV.3 LA INDUSTRIA Y SUS PEQUEÑOS ALIADOS Las levaduras fueron explotadas durante miles de años para la elaboración de bebidas alcohólicas y de pan. Esto ocurrió antes de que se identificara a las levaduras como un microorganismo y de que se conociera la verdadera naturaleza de la fermentación. Antonie van Leeuwenhoek fue el primero en observar la presencia de las levaduras en la fermentación de la cerveza. Casi 200 años después, en 1876, Louis Pasteur presentó sus ideas sobre la fermentación en una obra clásica titulada Études sur la Bière ("Estudios sobre la cerveza"), en la cual postula que los microorganismos que viven en condiciones anaeróbicas son capaces de crecer y vivir sustituyendo al proceso de respiración por la fermentación. Este proceso convierte azúcares en alcohol y bióxido de carbono, y provee a las levaduras con la energía necesaria para su subsistencia. Este científico propuso además que cuando se cultivan las levaduras en presencia de oxígeno, el proceso de fermentación se detiene y es sustituido por la respiración o consumo de oxígeno, proceso que sí degrada el azúcar hasta CO2 y agua y no produce alcohol. La fermentación es muy importante socialmente, ya que mediante ella se producen las bebidas alcohólicas que han tenido un papel importante en la historia. El vino, por ejemplo, es el producto de la fermentación del jugo fresco de frutas, de las cuales tal vez la más popular sea la uva, aunque también se usa ocasionalmente la pera, el higo y algunos cereales. Estos jugos son generalmente ácidos (tienen un pH de aproximadamente 5), lo cual es favorable para que se lleve a cabo la llamada fermentación alcohólica. La "infección" del jugo ocurre inmediatamente. Pasteur fue el primero en mostrar que hay levaduras en la cáscara de la uva y sobre todo cuando está próxima a madurar. Dichas levaduras forman grandes colonias en los puntos donde se lesiona la fruta y hay salida de jugo. Por otra parte, los insectos son un factor muy importante en este proceso, ya que 44 diseminan las levaduras entre la fruta. Este hecho se conoció cuando Wortmann observó cómo un jugo estéril no se fermentaba, y no fue hasta que una avispa lo tocó que el jugo empezó a fermentarse. Figura 18. Las levaduras que son microorganismos con núcleo han sido aliados importantes para la alimentación del hombre a lo largo de su historia. En la superficie de las uvas se encuentra la levadura conocida como Saccharomyces, que es el principal género responsable de la fermentación. Durante la fermentación alcohólica las levaduras convierten en alcohol la dextrosa y la fructosa, dos azúcares que se encuentran en el jugo de la fruta. La concentración que alcanza el alcohol es de 14%, ya que a esta concentración tanto la fermentación como el crecimiento de la levadura se detiene. De 45 esta manera, el vino contiene una concentración limitada de alcohol; por otra parte, su sabor y aroma se deben principalmente al contenido de amil-alcohol, ácido acético, ácido láctico y acetato etílico, siendo todas estas sustancias importantes en la calidad del producto. Los azúcares más comúnmente utilizados por las levaduras durante su crecimiento son la glucosa y la fructosa, que contienen 6 átomos de carbono. Sin embargo, existen otros compuestos con mayor o menor número de átomos de carbono y que son de interés en algunas industrias. Estos azúcares diferentes pueden a su vez ser metabolizados por algunas levaduras. Por ejemplo, la levadura Candida utilis, la cual se emplea en la industria alimentaria, puede crecer metabolizando azúcares de cinco átomos de carbono (pentosas); esto permite que crezcan en un producto colateral de la industria del papel que es precisamente un azúcar de cinco átomos de carbono. Otra levadura, Saccharomycopsis lipolytica, puede romper cadenas lineales de hidrocarburos de 10 a 16 átomos de carbono; existe una instalación piloto que ha logrado hacer crecer esta levadura con buenos resultados en presencia de una fracción purificada de petróleo, el cual, como se sabe, es un hidrocarburo. Esta levadura, al crecer, produce ácido cítrico como producto de desecho, el que a su vez es utilizado en otra serie de procesos industriales. Asimismo, una variedad limitada de levaduras puede crecer en presencia de metanol (un alcohol con un solo átomo de carbono), esto es posible ya que tales levaduras llevan a cabo un proceso metabólico novedoso que involucra unos organelos especializados llamados microcuerpos. Lo anterior se ha aprovechado, ya que el cultivo utilizando metanol es muy barato y la producción de biomasa de las levaduras es tan abundante que se utiliza como complemento para la alimentación de animales. Otra levadura que ha sido útil desde tiempos remotos es la cepa denominada Saccharomyces cerevisiae. Esta ha sido utilizada en la elaboración de vino, sake, cerveza y, hoy en día, para la producción industrial de alcohol. Una aplicación interesante es la que se le está dando a otra especie, Phaffia rhodozyma, que produce un pigmento especial llamado astaxantina. Esta molécula está siendo probada para añadirla en forma complementaria al alimento de las truchas y salmones de criadero. Como estos peces pierden el color rosado que presenta su carne cuando crecen en su hábitat natural, la astaxantina da a la carne de estos peces nuevamente su color rosado. Otros tipos de productos metabólicos de los cuales se obtienen grandes volúmenes a partir de cultivos de microorganismos son el dextrán y la goma xántica. El dextrán es una cadena de moléculas de glucosa que se utiliza como complemento del plasma en enfermos graves o bien como mallas moleculares al entrecruzar sus cadenas para separar compuestos en el laboratorio con base en su tamaño. La goma xántica, que es adecuada para el consumo humano, se agrega en algunos productos alimenticios como 46 estabilizador. También tiene uso en la impresión de textiles, en la excavación de pozos, como aditivo y en la elaboración de cosméticos y productos farmacéuticos. Los hongos microscópicos tienen una gran importancia económica, tanto por sus beneficios como por el daño que pueden causar. Una gran parte de la materia orgánica de la tierra es destruida por los hongos. Esta actividad, por ejemplo, es benéfica, ya que completa el reciclaje de la materia viviente. Los hongos pueden, sin embargo, causar un gran número de enfermedades tanto en las plantas como en los animales: pueden, entre otras cosas, envenenar la comida. Por ejemplo, el hongo Claviceps purpurea produce una serie de sustancias venenosas (alcaloides) y parasita a las plantas que come el ganado. Cuando el ganado consume alimento contaminado se producen abortos en las hembras, enfermedad que se conoce como ergotismo. En el humano la toxina produce alucinaciones y en ocasiones la muerte. Otro tipo de toxinas son las aflatoxinas, que se producen al almacenar el heno o alimento a base de cacahuate en condiciones impropias; el hongo llamado Aspergillus flaws las produce como metabolito secundario y son altamente tóxicas. Los efectos nocivos de los hongos microscópicos están balanceados por su multitud de usos en la industria. Así se emplean en algunos procesos de fermentación, como por ejemplo el almidón del arroz que es degradado para producir sake, un licor elaborado principalmente en Japón; la fermentación combinada de frijol de soya, arroz y malta da lugar a algunos alimentos orientales como el miso, el shoyu y el tempeh. Algunas enzimas provenientes de ciertos hongos tienen usos comerciales importantes, como por ejemplo: las amilasas (degradan almidón), las proteasas (degradan proteínas) y las pectinasas (degradan pectina). Algunas especies de hongos producen agentes antibióticos, entre los cuales destaca la penicilina. Esta sustancia marcó una época importante para la humanidad, ya que permitió el control de las enfermedades infecciosas. 47 Figura 19. Bacillus brevis es un microbio muy útil, produce el antibiótico conocido como gramicidina S, comúnmente utilizado en el tratamiento de numerosas enfermedades infecciosas Figura 20. Los hongos también son productores de antibióticos. La foto muestra al hongo Cephalosporium acremonium que produce la cefalosporina. De gran importancia para la industria alimentaria son algunas levaduras y hongos. Existen varios ejemplos, como la salsa de soya, la cual se obtiene utilizando a la levadura Saccharomyces rouxii; la producción de cerveza utiliza la levadura Saccharomyces carlsbergensis y algunos otros tipos de levaduras son utilizadas para la producción de vino. Ejemplos de bacterias importantes en la industria son los que se dan a continuación: la industria alimentaria aprovecha la bacteria Lactobacillus bulgaricus (búlgaros) o el Streptococcus thermophilus para la elaboración de yoghurt; el vinagre puede ser producido por la bacteria Gluconobacter suboxidans; algunos saborizantes provienen de una bacteria conocida como Connebacterium glutamicum. La industria química produce acetona y butanol a partir de la bacteria Clostidium acetobutylicum, algunos polisacáridos (azúcares) a partir de la bacteria Xanthomonas campestris algunas vitaminas como por ejemplo la B12 es producida por la bacteria 48 Pseudomonas denitrificans o el Propionibacterium. La industria farmacéutica fabrica un buen número de antibióticos; Streptomyces, por ejemplo, produce la anfotericina B, la kanamicina, la neomicina, la estreptomicina, las tetraciclinas, entre otras y Bacillus brevis produce la gramicidina S. Otro caso interesante es el de la producción de insecticidas biológicos. Éstos, los producen dos bacterias, Bacillus thuringiensis y el Bacillus popilliae, y tienen la ventaja de tener menos efectos nocivos que los insecticidas sintéticos como el DDT. Pueden, entre otros usos, emplearse en los silos donde se almacenan granos. Otra serie de productos, como la insulina, la hormona de crecimiento y el interferón, pueden ser obtenidos mediante técnicas de ingeniería genética utilizando la bacteria Escherichia coli. Así pues, la industria microbiológica se encuentra, al inicio de una era importante en donde se sustituirán probablemente muchos procesos tradicionales por otros novedosos que serán sin duda más eficaces y menos costosos. Figura 21. Una levadura en proceso de división, vista al microscopio de barrido de ele L a m i c r o b i o l o g í a e n f o t o g r a f í a s 49 Figura 1. Antonie van Leeuwenhoek inventor del microscopio y el primer hombre que observó los microorganismos Figura 2. Lazzaro Spallanzani (1729-1799). Fisiólogo italiano, uno de los primeros investigadores de la vida microscópica. 50 Figura 3. Louis Pasteur es el creador de la microbiología. Estudió las fermentaciones, las enfermedades contagiosas, la asepsia y descubrió la vacuna contra la rabia Figura 4. Portada de uno de los trabajos básicos de Pasteur. 51 Figura 5. Cultivo de Mycoderna vini efectuado por Pasteur. Figura 6. El moho reacciona así al ser sometido a una atmósfera de aire enrarecido. 52 Figura 7. El médico alemán Robert Koch descubrió el bacilo de la tuberculosis. Figura 8. Ylia Metchnikoff, microbiólogo ruso que ganó el premio Nobel por su descubrimiento de los fagocitos. 53 Figura 9. Cultivo de bacterias fotosintéticas. Estas bacterias se mantienen en condiciones controladas de luz y temperatura y son capaces de reproducirse en grandes cantidades. En la fotografía aparecen un cultivo líquido y un cultivo sólido, en donde se mantienen las capas, microorganismos. libres de la contaminación de otros 54 Figura 10. De los microorganismos se obtienen diversas sustancias de utilidad variada. Su aislamiento se lleva a cabo por medio de técnicas bioquímicas convencionales. Figura 11. Los cultivos en gran escala deben siempre estar libres de posibles contaminaciones. Así, pues, se prueban medios de cultivo adecuados para obtener las mejores condiciones de crecimiento. 55 Figura 12. Hoy en día la tecnología es una herramienta muy útil en el desarrollo de la microbiología. Así, se han creado industrias dedicadas a apoyar la instrumentación necesaria para el estudio de los microorganismos Figura 13. Los avances se basan en las técnicas básicas que fueron desarrolladas por los pioneros de la microbiología. Hoy en día se siguen aplicando sus conocimientos sobre la esterilidad de un cultivo y cómo preservarla. 56 C O N T R A P O R T A D A El eminente científico francés Louis Pasteur escribió que si los organismos más pequeños y aparentemente menos útiles, esto es, los microorganismos, fueran suprimidos, la vida se volvería imposible ya que el regreso a la atmósfera y al reino mineral de todo lo que dejó de vivir —el proceso de biodegradación— sería bruscamente suprimido. El doctor Georges Dreyfus se aboca en el presente libro al desarrollo de algunas áreas del conocimiento de los microorganismos, destacando aquellos temas que interesan al lector ajeno a la microbiología. Logra un trabajo cuya lectura es amena y fluida sobre un tema en apariencia árido y de interés sólo para el especialista, provocando el interés por conocer un mundo que es tan vasto como el Universo. El camino recorrido por la microbiología es largo. Ya doscientos años antes de nuestra era el romano Varro consideraba la posibilidad de que el contagio de ciertas enfermedades se debía a "criaturas invisibles suspendidas en el aire" y, a lo largo de siglos, tal idea fue considerada por diversos estudiosos. Sin embargo, el simple razonamiento sobre la existencia de los microbios no probaba su existencia, la que sólo pudo ser demostrada gracias al descubrimiento del microscopio, "el instrumento que abrió las puertas de otro universo" y cuya invención se debe al holandés Antonie van Leeuwenhoek. Como es frecuente, el descubrimiento de los microorganismos, aunque provocó interés, no cambió los esquemas de pensamiento que favorecían entonces la teoría de la generación espontánea, según la cual se podían crear animales a partir de una "fuerza vital", teoría que fue finalmente aniquilada por Louis Pasteur. En nuestro tiempo los microorganismos son tomados muy en serio, al grado que se ha creado una ciencia encargada de su estudio. En principio, su adaptabilidad los ha hecho sobrevivir en condiciones en extremo diferentes de las que prevalecen actualmente, las que existían en la Tierra hace millones de año. Además, gran parte de los microorganismos no es nociva para el hombre, al contrario, como se verá en este libro, es prometedora para la industria y la conservación del medio ambiente. Georges Dreyfus es Investigador Nacional e investigador titular del Departamento de Bioenergética del Instituto de Fisiología Celular de la UNAM. Hizo su maestría y doctorado con los doctores René Drucker y Marietta Tuena. Realizó además su posdoctorado en Grenoble, Francia. Se dedica ahora al estudio de la estructura de la ATP sintetasa y la función de esta enzima en las mitocondrias y las bacterias fotosintéticas. 57 En la portada: Cultivo líquido arbuscula. Laboratorio del autor. del hongo Diseño: Carlos Haces/Fotografía: Carlos Franco Calcarisporium