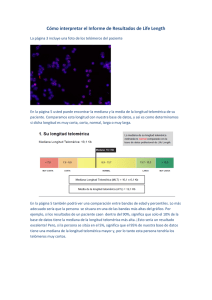
sumario • contents - Biblioteca Virtual en Salud de Cuba
Anuncio

SUMARIO • CONTENTS TRABAJOS DE REVISIÓN ESTRÉS OXIDATIVO Y ENVEJECIMIENTO Oxidative stress and aging 67 Karina Rodríguez Capote y Ela Céspedes Miranda LA ENFERMEDAD POR QUEMADURAS COMO MODELO DE RESPUESTA INFLAMATORIA SISTÉMICA Burn-related diseases as a model of systemic inflammatory response 77 Félix Broche Valle, Ela M. Céspedes Miranda, Alberto Saldaña Bernabeu y Arturo L. Cruz Pérez TÉCNICA LIPICID: SOFTWARE PARA LA DETECCIÓN, EVALUACIÓN Y TRATAMIENTO DE LAS DISLIPIDEMIAS LIPICID: Software for detection, assessment and treatment of dyslipidemia 86 Alfredo Nasiff Hadad y Eugenia G. Muñiz Lodos INFLUENCIA DE LA CONSERVACIÓN DE LA MUESTRA EN LA DETERMINACIÓN DE LA ENDOTELEMIA Effect of the preservation of samples in determining endothelmia 91 Ana María Quintela Pena TRABAJOS ORIGINALES COMPORTAMIENTO DE LA MORTALIDAD POR BRONQUITIS CRÓNICA. CUBA, 1980-1997 Situation of mortality from chronic bronchitis. Cuba, 1980-1987 95 Patricia Varona Pérez, Enrique Molina Esquivel, Joaquín Hechavarría Miyares, Vicente I. Prieto Díaz y Bárbara Taboada Fernández 67 CRECIMIENTO Y COMPOSICIÓN CORPORAL DE LAS CRÍAS DE 104 RATAS SOMETIDAS A RESTRICCIÓN ALIMENTARIA Body growth and composition of offspring of rats on diet restriction Daisy Valiente Bisset, Cristina Alfonso Zerquera, Manuela Gilda Bernardo Fuentes, José R. Molina García y Valentina T. González Cabrales DUPLO Y: ¿ESTIGMATIZACIÓN GENÉTICA? Duplex y: genetic stigmatization? 111 Héctor I. Pimentel Benítez, Jaime Fajardo Castellanos y Julia García Capote ALGUNAS CONSIDERACIONES GENERALES SOBRE EL DESARROLLO DE LA EVALUACIÓN DE PRODUCTOS FARMACÉUTICOS Y BIOTECNOLÓGICOS Y EQUIPOS MÉDICOS EN CUBA Y EL MUNDO Some general considerations on the development of the assessment of pharmaceuticals and biotechnological products and medical equipment in Cuba and the world 117 Martha María Fors López y Daniel Peña Amador TRABAJOS DE ACTUALIZACIÓN TELÓMEROS Y TELOMERASAS Telomeres and telomerases 121 Rolando A. Hernández Fernández ADYUVANTES INMUNOLÓGICOS Immunoadjuvants Humberto Joaquín Morris Quevedo, Clara Martínez Manrique, Roberto T. Abdala Díaz y Denia Campos Orama BOCIO DIFUSO EUTIROIDEO: 100 AÑOS DE TRATAMIENTO Diffuse euthyroid goiter: 100 years of treatment 130 138 Mayque Guzmán Cayado CARDIOPROTECCIÓN: UN TRIUNFO DE LA BIOMEDICINA DEL SIGLO XX Cardioprotection: an achievement of biomedicine in the 20th century Iliana Cabrera Rojo 68 146 Rev Cubana Invest Biomed 1999;18(2):67-76 TRABAJOS DE REVISIÓN Instituto de Ciencias Básicas y Preclínicas "Victoria de Girón" ESTRÉS OXIDATIVO Y ENVEJECIMIENTO Dra. Karina Rodríguez Capote y Dra. Ela Céspedes Miranda RESUMEN Se ha reportado un incremento del estrés oxidativo durante el envejecimiento y en las enfermedades asociadas a éste. Aunque no se conocen los mecanismos moleculares que condicionan el fenómeno del envejecimiento, se han formulado numerosas hipótesis al respecto. En el presente artículo se abordan aspectos bioquímicos relacionados con los radicales libres y el metabolismo oxidativo de las células como causa posible de senectud. Descriptores DeCS: ENVEJECIMIENTO/fisiología; RADICALES LIBRES/ /química; ESTRES OXIDATIVO. El aumento de la expectativa de vida ha originado un envejecimiento de la población y por consiguiente un incremento relativo de las enfermedades asociadas a éste. Aunque no se conocen los mecanismos moleculares que condicionan el fenómeno del envejecimiento, son numerosas las hipótesis formuladas al respecto.1,2 Éstas proponen perturbaciones en órganos, sistemas o en procesos bioquímicos vinculados con los radicales libres y el metabolismo oxidativo de las células, como causa fundamental de la senectud. La incuestionable ventaja evolutiva que supone desde el punto de vista energético la utilización del oxígeno molecular como último aceptor de electrones, haciendo posible la síntesis de ATP, se contrapone a su alta potencialidad citotóxica, pues de su reducción univalente se generan productos intermedios más reactivos conocidos como especies reactivas del oxígeno (ERO). Frecuentemente se ha empleado la expresión paradoja del oxígeno para referirse a estas funciones opuestas del O 2 en los organismos aerobios. El organismo desarrolla mecanismos de defensa para contrarrestar la acción de las especies reactivas del oxígeno. Estos mecanismos están constituidos por sistemas enzimáticos y sustancias de bajo peso molecular como tocoferoles, ascorbato, carotenos, ácido úrico y otros. En relación con la participación de los radicales libres en el envejecimiento es necesario considerar 3 factores fundamentales: la produc- 67 ción de oxidantes, el nivel de defensa antioxidante y la extensión del daño por oxidantes. TEORÍA DE LOS RADICALES LIBRES EN EL ENVEJECIMIENTO La hipótesis original de los radicales libres en el envejecimiento fue propuesta por Gerschman y Harman en los inicios de la década del 50, en un momento en que se conocía relativamente poco sobre los sitios celulares de generación de los radicales libres y sus subsecuentes reacciones moleculares.3,4 El dogma central de esta teoría radica en que durante el metabolismo aerobio se producen incidental e incontrolablemente especies radicálicas derivadas del oxígeno que, una vez generadas, promueven reacciones que dañan macromoléculas. Este daño irreversible se acumula con el tiempo resultando en una pérdida gradual de la capacidad funcional de la célula.1,2,4 PRODUCCIÓN DE OXIDANTES Aunque son varias las fuentes generadoras de ERO (tabla), se considera que la mitocondria es la más importante.5-8 TABLA. Producción endógena de ERO Enzima o sistema ERO CTE: Coenzima Q, cit b566 NADH deshidrogenasa SOD Monoaminooxidasa NADPH oxidasa (neutrófilos) Xantinooxidasa Reacción de Fentón Oxido nítrico sintetasa O -2 68 H 2O2 H 2O2 O-2 O -2 OH NO- El primer producto de la reducción parcial del oxígeno es el anión superóxido (O-2) que es producido en condiciones fisiológicas en la cadena transportadora de electrones (CTE) y es convertido en peróxido de hidrógeno (H2O2) espontáneamente o por acción de la enzima superóxido dismutasa (SOD). El radical hidroxilo (-OH), altamente reactivo, puede ser producido por reducción directa del H2O2 por el O-2, por transferencia directa de un electrón del O-2 al H2O2 catalizada por metales (reacción de Fenton) o por reacción directa de la ubisemiquinona reducida con el H2O2. La coenzima Q puede intervenir además en la formación de radicales hidroxilo, al reaccionar con el H2O2 en ausencia de iones metálicos como catalizadores y no requiere de protones que compensen la carga.6 El O-2 puede además reaccionar con el óxido nítrico (NO-) y generar peroxinitrito (ONOO-). El incremento en la formación de O-2 y H2O2 se justifica con el hallazgo de que en el envejecimiento se modifican las condiciones del flujo de electrones en la CTE y es más fácil a los electrones escapar de la secuencia normal del flujo.6 En estudios realizados en el músculo esquelético humano se vio un decrecimiento selectivo en los complejos I y IV durante el envejecimiento.9 Sohal reportó un incremento en la concentración de coenzima Q, en relación con otros componentes de la CTE, en insectos y en tejido murino de animales viejos.10,11 Villa y otros encontraron disminución en la síntesis del citocromo aa3 y de NADH deshidrogenasa y aumento en la familia de citocromo b, con la edad.12,13 Estos investigadores postulan que las ERO generadas pueden infligir daño, tanto a la membrana interna de la mitocondria como a los componentes de la CTE o al ADN mitocondrial, lo cual incrementa aún más la producción de ERO y consecuentemente más daño a la mitocondria e incremento del estrés oxidativo por aumentar la producción de oxidantes.4,6,10,13-16 El envejecimiento de la mitocondria determinaría también la duración de la vida. NIVEL DE DEFENSA ANTIOXIDANTE Numerosos estudios se han realizado para determinar si las defensas antioxidantes declinan con la edad. Estos estudios involucran mediciones de las enzimas y compuestos con funciones antioxidantes: actividad o expresión de SOD, catalasa (CAT), glutatión peroxidasa (GPx), glutatión reductasa (GRd), concentración de glutatión reducido (GSH), vitaminas, ácido úrico y otros. Diferentes autores han demostrado que las concentraciones de GSH en varios tejidos de origen murino disminuyen significativamente con la edad;6,11,17-21 similares resultados se han observado en insectos.11,22 Se ha encontrado una disminución en la actividad CAT en Drosophila y mosca doméstica durante el envejecimiento.11,21,23 En cambio, en mamíferos, el comportamiento de esta enzima con la edad es diferente según el tejido; disminuye en riñón e hígado y tiende a aumentar en corazón y cerebro, donde disminuye marcadamente en las últimas etapas de la vida.6,16,17,21,24-26 La actividad y expresión de GPx y glutatión-S-transferasa (GST) declinan durante el envejecimiento en todos los tejidos estudiados6,16,21,24,25 a diferencia de la GRd, que en mamíferos aumenta en cerebro y corazón pero disminuye en hígado y riñón6,17,27 y en insectos disminuye.11,21,23 Las marcadas diferencias entre los estudios referentes a la actividad o expresión SOD impiden arribar a una conclusión definitiva. Las referencias sobre el comportamiento de la actividad de esta enzima durante el envejecimiento son contradictorias. En numerosos estudios realizados no se encuentran cambios en la actividad o en la expresión con la edad 28-30 mientras que otros autores refieren disminución31,32 o aumento.33-35 Estas divergencias en los resultados en la determinación de la actividad SOD pueden ser causadas por diferencias entre especies, condiciones del hábitat del animal y por procedimientos técnicos empleados. Estudios en moscas Drosophila melanogaster que sobrexpresaban SOD-Cu/ Zn y CAT mostraron disminución del daño oxidativo molecular relacionado con la edad, disminución de la susceptibilidad al estrés oxidativo agudo y aumento en el tiempo de vida de las moscas. El tiempo de vida de mutantes que no expresan actividad SODCu/Zn fue significativamente más corto que el de su grupo control. 21,36 Estos estudios muestran que durante el envejecimiento hay un declinar en las defensas antioxidantes de la célula. El gen de la SOD-Cu/Zn se localiza en el cromosoma 21, por lo que varios autores han sugerido que el envejecimiento precoz asociado al síndrome de Down e incluso el retardo mental, pueda deberse a una sobrexpresión de esta enzima sin el aumento compensatorio de la CAT y la GPx.33,37,38 EXTENSIÓN DEL DAÑO POR OXIDANTES Existen evidencias del acúmulo de daño oxidativo con la edad: Daño al ADN • Se ha reportado incremento en la excreción urinaria de timidinglicol con la edad en ratón, ratas, monos y humanos. 5,39 • Se ha observado un incremento de 69 8 hidroxi-2-desoxiguanosina (8-OHdG) tanto en el ADN nuclear como el mitocondrial durante el envejecimiento.14,40 • Incremento progresivo, relacionado con la edad en el número de enlaces iónicos entre proteínas no histonas y grupos fosfatos del ADN y aumento en la concentración de aductos desoxiguanosinamalondialdehídos en el ADN con la edad.40 • Aumento en la condensación de la cromatina con la edad.4 Daño a proteínas • Se ha correlacionado con la edad el aumento en el contenido de grupos carbonilos por oxidación de proteínas.3,4,16,17 • Aparición de enlaces cruzados entre proteínas.2,4 Daño a lípidos • Estudios de fluorescencia, dienos conjugados y malondialdehídos han mostrado mayores niveles de peroxidación lipídica en tejidos de animales viejos respecto a los jóvenes.3,5,11,17,23,33,36,41 • Se han utilizado los niveles de exhalación de alcanos como indicador de la peroxidación lipídica in vivo en mamíferos e insectos (el n-pentano y el etano son productos del catabolismo de los ácidos grasos polinsaturados ω-3 y ω-6 respectivamente) y se ha observado que son mayores en animales envejecidos respecto a los jóvenes.3,15,20,23 • Con la edad aumentan las concentraciones de lipofucsina en la célula, el componente mayoritario de este pigmento del envejecimiento, es producto de la peroxidación lipídica.3,4,42 CRÍTICA A LA TEORÍA DE LOS RADICALES LIBRES EN EL ENVEJECIMIENTO El talón de Aquiles de esta teoría es que concibe al envejecimiento como un 70 fenómeno totalmente estocástico, pues postula que los daños que lo condicionan ocurren al azar, cuando en realidad durante el envejecimiento se produce una secuencia característica, progresiva, irreversible e incluso predecible, de cambios en todos los niveles de organización biológica que no pueden ser explicados a través de mecanismos azarosos. Otra objeción a esta hipótesis es que no explica por qué los cambios son graduales e irreversibles. Si el envejecimiento se debiera sólo al aumento de ERO y disminución relativa de antioxidantes endógenos, entonces con la administración de antioxidantes se podría enlentecer o acelerar el envejecimiento. Estudios experimentales de restricción dietética y suplemento antioxidante en ratón, rata e insectos no aumentaron el tiempo de vida máximo de estas especies; sólo se logró que más individuos lo alcanzaran y mejoró la calidad de la vida.3,43 La hipótesis del estrés oxidativo en el envejecimiento reconcilia y conceptualiza la información existente sobre este tema e intenta responder las interrogantes que se generan a partir de la teoría de los radicales libres. HIPÓTESIS DEL ESTRÉS OXIDATIVO EN EL ENVEJECIMIENTO El estrés oxidativo (EOx) se define como el desequilibrio entre las moléculas de alto potencial oxidante derivadas del oxígeno (conocidas como especies reactivas del oxígeno) y los sistemas antioxidantes; a favor de la generación de las ERO. La teoría del estrés oxidativo es una de las hipótesis que intenta explicar los cambios degenerativos y la pérdida neuronal que ocurren durante la senescencia. Esta hipótesis considera que el envejecimiento y el desarrollo no son fases distintas de la vida sino más bien que el envejecimiento es la etapa final del desarrollo y que aun cuando no es un fenómeno genéticamente programado ocurre por la influencia del EOx en el programa genético.3,4,16 Los postulados de esta hipótesis son:4 • El completamiento del programa genético que gobierna la secuencia y duración de varias fases ontogenéticas está ligado al gasto de una suma definida de energía. • El nivel de EOx depende de la velocidad de generación de oxidantes y de los niveles de defensa antioxidante, los cuales están genéticamente controlados; pero están influidos también por factores epigenéticos. • El EOx ejerce una influencia regulatoria en la expresión génica y es diferente en los distintos estadios del desarrollo. EL RELOJ METABÓLICO El concepto de índice basal metabólico como una determinante de la longevidad fue introducido por Rubner en 1908. Él encontró que la cantidad de energía metabolizada por gramo de peso, desde la madurez hasta la muerte, en 5 especies diferentes de mamíferos era relativamente similar. A partir de este hallazgo postuló que la materia viva gasta una cantidad definida de energía biológica durante la vida y entonces la duración de ésta se determina por el tiempo necesario para transformar dicha energía.21 Desde su formulación se han realizado numerosos experimentos dirigidos a verificar esta hipótesis. Es bien conocido que el tiempo de vida de los animales poiquilotérmicos puede ser alterado variando la temperatura ambien- tal, lo que afecta su índice metabólico. Estudios realizados por Sohal en la chinche Oncopeltus fasciatus mostraron que la vida de estos insectos fue cuatro veces mayor y con un consumo de oxígeno 4 veces menor a 18 EC que a 30 EC. Similares resultados fueron reportados en Drosophila melanogaster y mosca doméstica.21,23 La relación entre la temperatura, índice metabólico y longevidad en los mamíferos es más compleja a causa de que son homeotérmicos y a la necesidad fisiológica de actividad física para prevenir la atrofia muscular; no obstante, se ha podido demostrar que el índice de metabolismo basal en los mamíferos se correlaciona inversamente con el tiempo de vida de la especie.3,20 Estos hallazgos demostraron que si bien el tiempo de vida máximo podía ser alterado variando el índice metabólico, el total de energía gastado durante la vida (potencial metabólico) permanece constante y es característico de la especie. La existencia de un potencial metabólico para un genotipo dado sostiene el concepto de un reloj genéticamente controlado, el cual corre más en relación con el gasto de energía que con el tiempo.4 EL ESTRÉS OXIDATIVO ESTÁ BAJO CONTROL GENÉTICO Un mecanismo mediante el cual el índice metabólico influye en el desarrollo y el envejecimiento puede ser a través de modulaciones en el nivel de EOx. La célula tiende a generar oxidantes y antioxidantes en una forma inter-dependiente. Mientras que los oxidantes estimulan la producción endógena de antioxidantes;6,17-19,22 la administración de antioxidantes suprime varios componentes de las defensas endógenas.3,14 Estos hallagos sugieren que 71 el estado oxidativo de la célula es mantenido por mecanismos de retroalimentación. La administración de vitamina C (ácido ascórbico), vitamina E (tocoferol) o β-carotenos deprime las concentraciones de GSH y disminuye la actividad en moscas.44 La inactivación de CAT en ranas adultas indujo la síntesis de GRd45 y aumentó en el 20 % la concentración de GSH en el cerebro de las ratas.18,19 La transcripción de los genes de la SOD-Mn en E. coli fue suprimida ante la administración de GSH,46 mientras que al suministrarle diamida (agente oxidante de GSH) se vio inducción de la expresión SOD, CAT y GRd.47 Sohal encontró que existe relación entre el potencial de vida máximo y los niveles de defensa antioxidante en 6 especies diferentes de mamíferos y en insectos. 11,20,44 Los resultados de estos estudios indican que la generación de oxidantes, niveles de defensa antioxidante e índice metabólico son interactivos. EL ENVEJECIMIENTO ESTÁ ASOCIADO CON ALTERACIONES EN EL NIVEL DE EOX Y CAMBIOS EN LA EXPRESIÓN GÉNICA • Cambios en el nivel de EOx y, por tanto, en el estado redox de la célula, ejercen alteraciones en el balance iónico intracelular y en las propiedades estéricas de la cromatina, por formación de puentes disulfuro y enlaces iónicos entre las proteínas.21 • La expresión génica está fuertemente relacionada con el grado de condensación de la cromatina, por lo que se ha sugerido que la influencia del EOx en la expresión de los genes durante el desarrollo, se deba a los efectos que tiene en su configuración espacial.3,4,21 72 • El envejecimiento se caracteriza por alteraciones indicativas de un desgaste gradual del genoma tales como: disminución del número de receptores celulares, aparición de proteínas anormales, desrepresión de oncogenes, disminución de la transcripción, traducción y procesamiento del ARN. Este fenómeno fue descrito y denominado por Cutler como disdiferenciación.48 EL INCREMENTO DEL ESTRÉS OXIDATIVO ES UN FENÓMENO EPIGENÉTICO Aunque el nivel del estrés oxidativo está influido por mecanismos genéticos, el incremento en la formulación de ERO es un fenómeno epigenético.4 • La mitocondria es el principal sitio productor de ERO y también el primer blanco para el efecto deletéreo de estas moléculas, por lo que se genera un ciclo vicioso: formación de ERO → daño mitocondrial (ADN, membrana, trans- portadores) → alteraciones en la cadena transportadora de electrones →más formación de ERO. Alteraciones en la mitocondria pueden ser consecuencia y causa del incremento en la producción de ERO; pero este fenómeno es enteramente epigenético. • Durante el desarrollo, cambios en el nivel de EOx, programados genéticamente, inducen la expresión de nuevas proteínas o la supresión de determinados genes,48 por lo que es posible que en la fase adulta de la vida, modificaciones en el EOx por factores epigenéticos supriman también la expresión de determinados genes.4,21 • A partir de estas observaciones Sohal y Allen postularon que el envejecimiento no está gobernado por un programa genético per se, pero que ocurre por la influencia del EOx en el programa genético.3,4 INFLUENCIA DEL EOX EN LA EXPRESIÓN GÉNICA Existen evidencias que muestran que el EOx puede influir en la expresión génica en distintos niveles: transcripción, modificaciones postranscripcionales y traducción. Se ha sugerido que el efecto del EOx en la expresión génica sea a través de 2 mecanismos: 1. Por efecto directo sobre la producción y procesamiento del ARN. 2. Por cambios en la distribución iónica de la célula. Efectos en el nivel del control transcripcional: – En E. coli, Salmonella y Typhimurium una proteína conocida como Oxy R, cuando es oxidada por H2O2 actúa como un activador transcripcional que induce la expresión de CAT, SOD-Mn, GRd, alkilhidroperóxido reductasa, entre otras.49 – En células eucariotas todavía no está claro cómo se controla la expresión de los genes SOD, pero hay 2 mecanismos propuestos: • A través del factor transcripcional AP-1, que en su componente c-fos tiene un residuo de cisteína crítico muy sensible a la oxidación.33,49 • A través de la oxidación de la subunidad inhibitoria del NF-kB.49 – La SOD-Mn es específicamente inducida por el factor de necrosis tumoral (TNFα). Existen evidencias de que este factor es inducido por ERO, y éste a su vez activa al NF-kB por lo que se sugiere que existe una relación entre la activación oxidativa del NF-kB y la inducción de la SOD-Mn. 33 Efectos en el nivel de las modificaciones postranscripcionales: – Recientemente se ha observado que el procesamiento del ARN es alterado por la exposición de la célula a sistemas generadores de radicales del oxígeno.4 – Anticuerpos contra la enzima SOD incrementan la liberación de ARN inmaduro del núcleo al citoplasma, efecto que puede ser suprimido por la administración de SOD. – El hecho de que en organismos en desarrollo, en tejidos de individuos senescentes y en tejidos bombardeados con O2 se haya encontrado ARN inmaduro en el citoplasma sugiere que los cambios asociados al envejecimiento y al desarrollo puedan ser mediados por la generación y eliminación de ERO.4 EN RESUMEN LA HIPÓTESIS DEL ESTRÉS OXIDATIVO PLANTEA – Los cambios en la expresión génica que gobiernan eventos ontogenéticos se acompañan de cambios en el nivel de EOx y viceversa. – El daño que se acumula durante el envejecimiento es más bien un efecto secundario que una causa directa de la senescencia. – El EOx es uno de los factores que gobiernan los cambios en la expresión génica durante la diferenciación y el envejecimiento. Concluimos que el envejecimiento es un problema científico que en las ciencias 73 biomédicas contemporáneas se aborda con un enfoque multidisciplinario. En estos momentos se plantea, a pesar de las numerosas hipótesis que se han propuesto para tratar de explicar este fenómeno, que existe un proceso único modificable por facto- res genéticos y ambientales, responsable del envejecimiento, que también es determinado por el envejecimiento de la mitocondria y la producción de radicales libres como resultado del metabolismo en este organelo. SUMMARY An increased oxidative stress has been reported during aging and the diseases associated to it. Although the molecular mechanisms triggering the phenomenon of aging are unknown, a number of hypotheses have been franed on this subject. The present article deals with biochemical aspect of free radicals and oxidate cell metabolism as a possible causes of senescence. Subject headings: AGING/physiology; FREE RADICALS/chemistry. REFERENCIAS BIBLIOGRÁFICAS 1. Medveded Z. An attempt at a rational classification of theories of aging Biol Rev 1990;65:375-98. 2. Hayflick L. theories of bilogical aging. Exp Gerontol 1985;20:145-59. 3. Sohal RS. The free radical hypothesis of aging: an appraisal of the current status. Aging Clin Exp Res 1993;5:3-17. 4. Sohal RS, Allen RG. Oxidative stress as a causal factor in differentiation and aging; a unifying hipothesis. Exp Ger 1990;25:499-522 5. Floyd RA. Free radidals in molecular biology. Aging and disease. New York: Raven, 1984. 6. Benzi G, Moretti A. Age-and peroxidative stress-related modifications of the cerebral enzymatic activities linked to mitochondria and the Cclutatione System. Free Rad Biol Med 1995:19(1):77-101. 7. Storz G, Tartaglia LA, Ames BN. Transcriptional regulator of oxidative stress-inducible genes: direct activation by oxidation: Science 1990;248:189-94. 8. García JC, Clapés S, Céspedes EM, Simplice O, Morín MA. Radicales libres sueños moleculares o realidades clínicas. Rev Cubana Invest Biomed 1994;13(1-2):12-8. 9. Cooper JM, Mann UM, Schapira AHV. Analysis of mitochondrial respiratory chain function and mitochondrial DNA deletionin in human skeletal muscle: effect of aging. J Neurol Sci 1992;113:91-8. 10. Farmer KJ, Sohal RS. Relationship beween superoxide anion radical generation and aging in the housefly, Musca Domestica. Free Rad Biol Med 1989;7:23-9. 11. Sohal RS, Arnold LA, Sohal BH. Aged-related changes in antioxidants enzymes and pro-oxidant generation in tissues of the rat with special reference to parameters in two insect species. Free Rad Biol Med 1990;10:495500. 12. Villa RF, Gorini A. Action of L-acetylcarnitine on different cerebral mitochondrial populations from hippocampus and striatumm during aging. Neurochem Res 1991;16:1125-32. 13. Sohal RS, Sohal BH, Brunk UT. Relationship between antioxidant defenses and longevity in diferent mammalian species. Mech Ag Dev 1990;53: 217-27. 14. Beal FM. Aging, energy and oxidative stress in neurodegenerative diseases. Ann Neurol 1995;38:(3):357-66. 15. Sohal RS, Brunk UT. Mitochondrial production of pro-oxidants and cellular senescence. Mut Res 1992;275:295304. 74 16. Sohal RS, Agarwal S, Sohal BH. Oxidative stress and aging in the Mongolian Gerbil.Mech Ag Dev 1995;81(1):15-25. 17. Mo JQ, Hom DG, Andersen JK. Decreases in protective enzymes correlates with increased oxidative damage in the aging mouse brain. Mech Ag Dev 1995;81:73-82. 18. Benzi G, Pastorris S. Age-related effect induced by oxidative stress on the cerebral Glutathione System. Neurochem Res 1989;14(5):473-81. 19. Benzi G, Marzatico F, Pastoris O. Influence of oxidative stress on the age-linked alterations of the cerebral Glutathione System. J Neurosci Res 1990;26:120-8. 20. Villa RF, Gorini A. Effect of CDP-choline treatment on mitochondrial and synaptosomal protein composition in different brain regions during aging. Int J Dev Neurosci 1993;11:83-93. 21. Sohal RS, Allen RG. Relationship between oxygen metabolism, aging and development. Adv Free Rad Biol Med 1986;2:117-60. 22. Shull S, Heints NH. Differential regulation of antioxidant enzymes in response to oxidants. J Biol Chem 1991;266:24398-403. 23. Sohal RS, Donato H, Biehl ER. Effect of age and metabolic rate on lipid peroxidation in the housefly, Musca Domestica. Mech Ag Dev 1981;16:159-67. 24. Nisticò G, iriolo MR, Fiskin K. NGF restores decrease in catalase activity and inreases superoxide dismutase and glutathione peroxidase activity in the brain of aged rats. Free Rad Biol Med 1992;12:177-81. 25. Nagy ZS. The horizons of an interdisciplinary synthesis in experimental gerontology. Arch Gerontol Geriatr 1991;12:329-49. 26. Gutteridge JMC. Oxigen radicals, transition metals and ageing. Advances in age pigments research. ed. York: Pergamon, 1987:1-22. 27. Iantomasi SM, Favilli F. Glutathione metabolism in heart and liver of the aging rat. Biochem Cell Biol1994;72(1-2):58-61. 28. Danh HC, Strolin BM, Dostert P. Differential changes in superoxide dismutase activity in brain and liver of old rats and mice. J Neurochem 1986;46:1344-52. 29. Rao G, Semsei I, Richardson A. Expression of superoxide dismutase and catalase in rat brain as a function of age. Mech Ag Dev 1992;58:13-9. 30. Rao G, Xia E, Richardson A. Effect of age on the expression of antioxidant enzymes in male Fisher F344 rats. Mech Ag Dev 1990;53:49-60 31. Gupta A, Hasan M, Chander R. Age related elevation of lipid peroxidation products: diminution of superoxide dismutase activity in the CNS of rats. Gerontology 1991;37:305-9. 32. Vanella A, Geremia E, D’Urso G. superoxide dismutase activities in aging brain. Gerontology 1982;28:108-13. 33. Warner HR. Superoxide dismutase, aging and degenerative diseases. Free Rad Biol Med 1994;7(3):249-58. 34. Scarpa M, Rigo A, Viglinio P. Age dependence of the level of the enzymes involved in the protection against active oxygen species in the rat brain. Proc Soc Exp Biol Med 1987;185:129-33. 35. Ross BM, Kim DK, Bonventre JV. Characterization of a novel phospholipase A2 activity in human brain. J Neurochem 1995;64(5):2213-21. 36. Shi I, Sawada M. Alterations in free radicals activity in aging Drosophila. Exp Gerontol 1994;29(5):574-84. 37. Ciriolo MR, Fiskin K, Martino A. Age-related changes in Cu/Zn SOD, Se-dependent and independent Glutathione Peroxidase and Catalase activities in specific areas of rat brain. Mech Ag Dev 1991;61:287-97. 38. Oenzil F, Kishikawa M, Mizuno T. Age-related acumulation of lipofuscin in three different regions of rat brain. Mech Ag Dev 1994;76(2):156-63. 39. De Haan JB, Newman JD. Cu/Zn superoxide dismutase mRNA and anzyme activity, and susceptibility to lipid peroxidation, increases with aging in murine brains. Mol Brain Res 1992;13:179-87. 75 40. Ames BN. Endogenous oxidative DNA damage, aging and cancer. Free Rad Res Comms 1989;7:121-8. 41. Nakano M, Oenzil F, Mizuno T Gotoh S. Aged related changes in the lipofuscin acumulation of brain and heart Int J Exp Clin Gerontol 1995;41(2):69-79. 42. Oenzil F, Kishikawa M, Mizuno T. Age-related acumulation of lipofucsin in three different regions of rat brain. Mech Ag Dev 1994;76(2):156-63. 43. Harman D. Aging: a theory based on free radical and radiation chemstry. J Gerontol 1956;11:298-300. 44. Sohal RS, Toy PL, Allen RG. Ralationship between life expectancy, endogenous antioxidants and products of oxygen free radical reactions in the housefly. Mech Ag Dev 1986;36:71-7. 45. Pérez-Campo R, López-Torres M, Rojas C. Lung Glutathione reductase induction in aging catalase-depleted frogs correlates with early survival throughout the life span. Mech Ag Dev 1993;67:115-27. 46. Gardner PR, Fridovich I. Controls of the biosynthesis of Mn-SOD of E coli. J Biol Chem 1981;262:17591-5. 47. Privalle CT, Fridovich I. Biosynthesis of the manganese containing superoxide dismutase in E. Coli: anaerobic induction of active Mn-SOD by diamide. Free Rad Biol Med 1990;9:1-5. 48. Cutler RG. Antioxidants, aging and longevity. Free Rad Biol 1984;6:371-428. 49. Schroder HC, Messer R, Bachmann M. Superoxide radical induced loss of nuclear restriction of inmature mRNA: A posible cause of aging. Med Ag Dev 1987;41:251-6. Recibido: 21 de mayo de 1999. Aprobado: 19 de julio de 1999. Dra. Karina Rodríguez Capote. Instituto de Ciencias Básicas y Preclínicas " Victoria de Girón" . Calle 146 No. 3102. Playa, Ciudad de La Habana, Cuba. 76 Rev Cubana Invest Biomed 1999;18(2):77-85 Instituto de Ciencias Básicas y Preclínicas "Victoria de Girón" LA ENFERMEDAD POR QUEMADURAS COMO MODELO DE RESPUESTA INFLAMATORIA SISTÉMICA Dr. Félix Broche Valle, Dra. Ela M. Céspedes Miranda, Dr. Alberto Saldaña Bernabeu y Dr. Arturo L. Cruz Pérez RESUMEN La quemadura corporal es una violenta agresión que modifica todos los mecanismos de la homeostasis orgánica y por su connotación clínica y social es un problema que enfrentan los servicios médicos en la sociedad contemporánea. Desde el punto de vista fisiopatológico, en el paciente quemado se desarrolla un Síndrome de Respuesta Inflamatoria Sistémica (SRIS) caracterizado por la hiperactivación de todos los mecanismos de defensa. La disregulación de estos mecanismos conduce al daño de los tejidos propios cuyas consecuencias se expresan en alteraciones morfofuncionales de todos los sistemas. Con independencia de la etiología, evolución inicial, manejo terapéutico y respuesta individual, la sepsis generalmente complica la evolución del gran quemado. En este trabajo, se presentan elementos clínicos e histopatológicos de la enfermedad por quemadura y se comentan los mecanismos moleculares de la respuesta inflamatoria sistémica destacando el papel de los mediadores de la comunicación intercelular. Descriptores DeCS: QUEMADURAS/patología; SINDROME SEPTICO/ /fisiopatología. Aldrich en 1894 definió que la quemadura es una pérdida de sustancia de la superficie corporal por destrucción de la piel y el tejido subcutáneo ocasionada por alteraciones térmicas que comprenden el calor, el frío, agentes químicos, la electricidad y las radiaciones.1 Todos los tipos de quemaduras poseen un común denominador: la producción de alteraciones histológicas de la piel y la aparición de un síndrome de respuesta inflamatoria sistémica (SRIS).2 La piel es el tejido más extenso del organismo y diana primaria en la quema- dura corporal. Fue considerada durante mucho tiempo como una estructura sólo de protección, pero se ha demostrado su carácter inmunocompetente.1-3 Los queratinocitos (cerca del 95 % de las células epidérmicas) pueden actuar como células presentadoras de antígeno y sintetizar mediadores de la comunicación intercelular que intervienen en el proceso inflamatorio. Por otra parte, las células de Langerhans (3-5 % de las células epidérmicas) funcionan como fagocitos tisulares así como en la presentación de antígenos a los linfocitos T CD4+.2 Los polimor- 77 fonucleares neutrófilos (PMN) circulantes son atraídos hacia la piel afectada y su hiperactivación contribuye a potenciar el daño generado por el trauma inicial. 4 Cuando la extensión de la quemadura rebasa ciertos límites deja de ser un trastorno local para convertirse en la "enfermedad por quemadura"1 que requiere de un tratamiento intensivo. En este contexto, nuestro trabajo se orienta en función de profundizar en el estudio de los mecanismos moleculares involucrados en la fisiopatología de la enfermedad por quemaduras con la óptica del enfoque multidisciplinario que la complejidad del problema nos impone. CONSIDERACIONES GENERALES SOBRE LA ENFERMEDAD POR QUEMADURAS Al analizar la evolución del quemado debe distinguirse entre el efecto agudo de la quemadura que conduce al shock hipovolémico en las primeras 48-72 h y un efecto de evolución progresiva que se convierte en la "enfermedad del quemado".2 La teoría del choque nervioso de Dupuyttren es la más antigua, se le atribuye al choque neurógeno por efecto del dolor el origen de todos los trastornos en el gran quemado. Eppinger sostuvo que las lesiones producidas por la quemadura debían atribuirse a trastornos de la permeabilidad capilar, en tanto Roplleer demostró en el suero de los quemados graves la existencia de una sustancia con tal efecto.1 Lorthioir señaló la liberación de enzimas proteolíticas intracelulares como causa de agresión generalizada a los tejidos. Otros autores identifican como causa fundamental de las alteraciones viscerales del gran quemado a la hipoxia hística.1,5 78 Hayde y Vogt sostuvieron el origen tóxico de los trastornos humorales, aunque sin precisar la naturaleza de las toxinas.1 Las toxinas del quemado pueden generarse a partir de la coagulación y destrucción hística producidas por el accidente, por la actividad metabólica de las células vecinas, en particular los fagocitos tisulares, el endotelio vascular y los fagocitos circulantes o los microorganismos que colonizan la piel o alcanzan la circulación sistémica. Duval atribuyó la muerte de los quemados graves a una toxina polipeptídica producida por la propia quemadura.1 Se ha identificado un principio tóxico que puede ser separado por precipitación salina y es de naturaleza lipoproteica, lo cual sugiere su probable procedencia de las membranas celulares.2,6 Esta toxina está constituida por 6 subunidades asociadas a proteínas del shock térmico, pesa entre 4-16 kDa y el 40 % del peso total de la par-tícula corresponde a lípidos.2,7,8 Los fagocitos pueden activarse tras su exposición a este complejo lipoproteico (CLP). Sin embargo, aunque este complejo induce una activación inicial, algunos días después de la quemadura disminuye significativamente el tiempo de vida media y la capacidad de respuesta de todas las células fagocíticas.2 El CLP genera una respuesta inmune específica.2,9 En estudios realizados con inóculos de Pseudomona sp. en ratones se obtuvo el 20 % de mortalidad en tanto tras la administración conjunta con una dosis subletal del CLP se obtuvo el 80 % de mortalidad. Múltiples estudios demuestran que el conjunto de las diversas toxinas identificadas hasta el momento en pacientes quemados ejerce un efecto citotóxico mucho menos pronunciado que el CLP, especialmente sobre las células del sistema reticuloendotelial.2 Kosaka K y otros, identificaron un epóxido de linoleato sintetizado por los polimorfonucleares neutrófilos (PMN) cuya concentración exhibe una cinética bifásica con máximos durante y después de la primera semana que sigue a la quemadura. Se obtuvo una correlación positiva entre los niveles de esta leucotoxina y la superficie corporal dañada, la tasa de mortalidad y la concentración plasmática de proteína C reactiva.10 La producción de neopterina por los fagocitos también se incrementa, de inicio como indicador de la respuesta de fase aguda al trauma tisular, pero posteriormente es inducida por las endotoxinas bacterianas. 11 En pacientes quemados con más del 20 % de superficie corporal comprometida, se elevan las concentraciones plasmáticas de endotelina 1 y trombomodulina, proporcionalmente con la concentración plasmática de factor de necrosis tumoral (TNF), la severidad del cuadro inflamatorio sistémico y la presencia de sepsis.12 En todos los quemados graves se incrementa la lipolisis y por tanto, la concentración de ácidos grasos libres, proceso estimulado por los glucocorticoides, interleuquina 1 (IL-1) y TNF. Sin embargo, IL-1 y TNF también estimulan la síntesis hepática de lipoproteínas de muy baja densidad; éstas pueden fijar e inactivar las endotoxinas bacterianas. Estos pacientes presentan también hipocetonemia.13 Desde el punto de vista histopatológico, en las quemaduras superficiales se produce vasodilatación cutánea con área de espongiosis en el epitelio, acantólisis secundaria que da lugar a la formación de flictenas y ampollas dermoepidérmicas con desprendimiento total de la epidermis y edema intenso de la dermis. En las quemaduras profundas en el nivel de la piel, es característica la escara constituida por los tejidos necróticos.1 Poco tiempo después de la quemadura, aparece vasodilatación y edema cerebral marcados con degeneración neuronal difusa. Las manifestaciones neurológicas iniciales del quemado incluyen inquietud y ligera desorientación. En los casos complicados aparecen progresivamente excitación, delirio y coma.1 En el miocardio aparecen áreas de necrosis, hemorragias focales, pericarditis fibrinosas y miolisis que clínicamente se expresan por arritmias e insuficiencia cardíaca.1,14 Las alteraciones de la integridad y permeabilidad vascular son evidentes en el nivel del foco de quemadura. En los casos más graves pueden aparecer focos de necrosis en la pared de los grandes vasos así como microtrombosis generalizada. El sistema hemolinfopoyético muestra alteraciones morfofuncionales en todos sus componentes en el nivel medular y periférico.1 En los túbulos renales aparecen depósitos de hemoglobina con degeneración parenquimatosa difusa y signos de fallo renal que puede llegar a la anuria. En los casos más graves aparece necrosis cortical bilateral, nefritis intersticial, abscesos múltiples e infartos difusos.1 Durante las primeras 48-72 h aparece edema pulmonar intenso con áreas de enfisema y neumonía reticulohiperplástica. Clínicamente estos pacientes presentan un cuadro de insuficiencia respiratoria progresiva. La congestión intensa, las hemorragias focales y las ulceraciones son manifestaciones frecuentes en la mucosa del tubo digestivo. Algunos casos presentan un síndrome ictérico.1 Los microorganismos causantes de la infección del quemado proceden al menos, de la piel, la mucosa intestinal1,5,15 y el medio ambiente.16,17 A menudo existen aso- 79 ciaciones de microorganismos multirresistentes. Las bacterias gramnegativas son la causa principal de la sepsis sistémica en los pacientes quemados (aproximadamente el 65 % de los casos) seguidas por las grampositivas (20 %), hongos, rickettsia y virus.18,19 En el nivel de la piel quemada es mucho mayor la incidencia de infecciones con respecto a otras lesiones traumáticas a causa de la pérdida de continuidad más o menos extensa de la barrera cutánea, primera línea de defensa del organismo, y a la presencia de alteraciones cuantitativas y cualitativas en la respuesta inmune humoral y celular de estos pacientes.20 Las quemaduras pueden presentar signos de infección local como: secreción verdosa fétida o escaras aparentemente secas por debajo de las cuales se acumula pus, en tanto en el nivel sistémico los signos clínicos de la infección local son la fiebre, leucocitosis, anemia progresiva, retraso o detención del proceso cicatrizal.1 La infección del quemado debe enfocarse como generalizada y tratarse local y generalmente. No obstante, la asociación microbiana por incorporación de nuevos gérmenes por vía externa o por la resistencia bacteriana a la antibioticoterapia complica notablemente el tratamiento.1 El paciente quemado pudo presentar: Bacteriemia y Endotoxemia (se refieren a la presencia de hemocultivos positivos y de endotoxina bacteriana en sangre respectivamente; Sepsis que implica la evidencia clínica de infección acompañada de reacción sistémica (taquipnea, taquicardia, leucocitosis, fiebre);18,21 Síndrome séptico que incluye sepsis con alteraciones en la perfusión de 1 o varios órganos y Shock séptico que requiere reposición de volumen y tratamiento vasopresor.18 Con el incremento de los niveles plasmáticos de endotoxinas bacterianas 80 aumenta la incidencia de insuficiencia multiorgánica y la tasa de mortalidad; de hecho, la endotoxemia persistente es siempre un indicador de mal pronóstico para el paciente quemado.22 Las estrategias más recientes basadas en la atención integral del paciente, eliminación temprana de los tejidos necróticos, uso de poliquimioterapia antimicrobiana local y sistémica y la hospitalización en condiciones de aislamiento han mejorado el pronóstico general del gran quemado23 pero aún son muchos los problemas que requieren consideración y solución adecuada tanto en el orden de la investigación básica como en el campo de la práctica asistencial. EL SÍNDROME DE RESPUESTA INFLAMATORIA SISTÉMICA EN LA FISIOPATOLOGÍA DE LA ENFERMEDAD POR QUEMADURA Aunque en la reacción inflamatoria participan factores celulares y humorales en el nivel local y sistémico, el SRIS se reconoce como el conjunto de alteraciones morfofuncionales que se derivan de la hiperactivación prolongada de dichos factores con independencia de la fuente de daño primario.24 Bone C. definió el SRIS como la reacción inflamatoria masiva resultante de la liberación de mediadores sistémicos que conducen a la disfunción multiorgánica.25 Los criterios de Bone para el diagnóstico de la respuesta inflamatoria sistémica de origen séptico son los siguientes: – Fuente primaria de infección confirmada. – Temperatura axilar < 36 o >38 E. – Frecuencia cardíaca > 90 latidos/min. – Frecuencia respiratoria > 20 resp./min. – Leucograma: > 12 000/mm3 , < 4 000/mm3 o > 10 % de células inmaduras. El origen del SRIS puede ser infeccioso y no infeccioso.24 En ambos casos, puede evolucionar hacia formas más graves que incluyen el shock, síndrome de insuficiencia multiorgánica y la muerte. En todo paciente quemado grave se presenta un SRIS de origen traumático, inicialmente no infeccioso y al cual se sobreañade la sepsis en el transcurso de su evolución2,26 (fig.). La hiperactividad de los leucocitos, en particular los PMN y las células endoteliales constituye el elemento fisiopatológico esencial en el SRIS.24,27.28 La quimiotaxis, adherencia, migración a través del endotelio y actividad fagocítica son las principales funciones biológicas que expresan las células activadas en el curso de la reacción inflamatoria aguda. Los macrófagos tisulares al actuar como células presentadoras de antígeno liberan factor de necrosis tumoral a (TNF a) e Microorganismos IL-1 que a su vez inducen por un mecanismo autocrino la síntesis de IL-6 e IL-8. Esta úlitma se comporta como potente activador de los PMN. 29 Otros estímulos como el complemento activado (C5a), el factor activador plaquetario, leucotrieno B4, peróxidos lipídicos, inmunocomplejos y restos celulares inducen también la activación de los PMN. 30 En todos los eventos del proceso inflamatorio interviene un complejo sistema de "señalización" integrado por medidoras de la comunicación intercelular. En 1974 Cohen utilizó el término "citoquinas" para identificar un grupo de polipéptidos con acciones locales y sistémicas, que participan en el control de diversas poblaciones celulares y que no derivan de los linfocitos. Actualmente, se define como citoquinas un grupo de proteínas inducibles que modulan la actividad (metabolismo, síntesis y secreción de otras proteínas, etc), proliferación y diferenciación celular.31 Según Bone C., en el desarrollo del SRIS pueden distinguirse 3 etapas de acuer- Toxinas endógenas y exógenas Traumatismos Síndrome de respuesta inflamatoria sistémica Manifestaciones focales Manisfestaciones generales Sepsis, shock séptico Enfermedad por quemaduras Distrés respiratorio Coagulación intravascular diseminada Fig. El síndrome de respuesta inflamatoria sistémica en la patología clínica. 81 do con la actividad de las citoquinas involucradas en la respuesta: Primera etapa que se caracteriza por la liberación local de citoquinas como respuesta al trauma y la infección. Segunda etapa, donde se incrementa la concentración de citoquinas en la circulación sistémica. Tercera etapa que se define por la reacción generalizada en la que la liberación masiva y prolongada de citoquinas conduce al daño vascular y de órganos diana.25 Entre las citoquinas proinflamatorias que desempeñan una función esencial en la respuesta aguda a la infección y el trauma tisular se incluyen, entre otras, IL-1, IL-6, IL-8 y TNF.26,31,32 Como IL-1 se conocen 2 proteínas (IL1 a e IL-1 b) sintetizadas por monocitos, macrófagos, linfocitos, queratinocitos, células de Kupffer, hapatocitos, fibroblastos y células de la glia entre otras.33 Se conocen 2 tipos de receptores y sólo el tipo I (en linfocitos T, fibroblastos, queratinocitos y células endoteliales) puede iniciar el mecanismo de transducción de señales. Sus principales funciones incluyen la estimulación de su propia síntesis e inducción de la síntesis de TNF e IL-6 (por los monocitos y macrófagos) y de IL-2, IL-4 y factor estimulante de colonias de granulocitos/monocitos (por los linfocitos T). Estimula la proliferación y actividad de los linfocitos B, induce la síntesis de prostaglandinas en células endoteliales y del músculo liso, aumenta la síntesis de las proteínas de fase aguda en los hepatocitos e induce la síntesis de colagenasa en fibroblastos y mesotelio sinovial. También induce la fiebre.33 La IL-8 es sintetizada por monocitos/ /macrófagos, eosinófilos, linfocitos, neutrófilos, fibroblastos, células endoteliales, queratinocitos, hepatocitos, astrocitos y condrocitos. Sus principales 82 acciones biológicas son la quimiotaxis, activación de leucocitos y expresión de varias moléculas de adhesión intercelular (MAI). Es además un factor angiogénico. La IL-8 actúa por unión a 3 receptores asociados a proteínas G en la membrana citoplasmática.34 El TNF α (caquectina) y el TNF β (linfotoxina) son 2 proteínas con el 30 % de identidad secuencial, que se incluyen entre las citoquinas proinflamatorias por su papel central en la iniciación de la respuesta inmune frente a infecciones y algunos tipos de tumores. El TNF α es sintetizado por PMN, linfocitos activados, macrófagos, astrocitos, células endoteliales, células musculares lisas y algunas células transformadas. El TNF β es sintetizado sólo por linfocitos.35 Virtualmente todas las poblaciones celulares poseen al menos 1 de los 2 tipos de receptores para TNF. Al parecer el receptor tipo I está vinculado con la actividad lítica en tanto el tipo II regula la proliferación y activación linfocítica. Se han identificado proteínas solubles que forman complejos inactivos con el TNF.35 La concentración plasmática de TNF se eleva significativamente y de forma persistente en pacientes quemados que fallecen por síndrome séptico.36-38 Las MAI son proteínas de membrana que intervienen en la interacción y unión de células sanguíneas a sus similares, al endotelio y a la matriz subendotelial.39,40 La mayoría de las MAI conocidas se incluyen dentro de 4 superfamilias: inmunoglobulinas, integrinas, selectinas y cadherinas.40 La superfamilia de inmunoglobulinas incluye las MAI (ICAM-1, ICAM-2, ICAM3, VCAM-1, MadCAM-1) que reconocen a las integrinas presentes en la membrana de los leucocitos y promueven la marginación y posterior extravasación de éstos a través del endotelio.39,40 La molécula de adhesión intercelular 1 (ICAM-1) es una licoproteína de membrana que se expresa en monocitos, células endoteliales, linfocitos, células dendríticas y epiteliales. Su función principal es la de intervenir como mediador de la adhesión leucocitaria al endotelio vascular activado.41 Las selectinas son una familia de glicoproteínas que se activan en las fases iniciales de la extravasación leucocitaria durante el proceso inflamatorio. 39,40 En particular, la L-selectina interviene en el "reclutamiento" de los leucocitos circulantes, su adherencia al endotelio y extravasación 39,42 hacia los sitios de inflamaciones aguda y crónica. La L-selectina (constitutiva de los PMN) y las β2 integrinas (inducibles durante el proceso inflamatorio) facilitan la adherencia de estas células a los endoteliocitos a través del reconocimiento de sus receptores (E-selectina para la Lselectina, ICAM-1 e ICAM-2 para las β2 integrinas). Las células endoteliales activadas sintetizan también IL-8.43 Además de la hiperproducción de estas citoquinas, se ha reportado también un aumento asociado de la producción de IL-6 por los macrófagos, lo cual podría ser una de las causas de la respuesta celular deficiente en estos pacientes, aumentando su susceptibilidad a las infecciones. 44 Las concentraciones plasmáticas de TNF, IL-6 e IL-8 alcanzan valores más altos en los quemados que desarrollan sepsis y en los que fallecen en insuficiencia multiorgánica.45 Los fagocitos y el endotelio son los principales puntos de generación de ERO en el SRIS. En pacientes críticos se han encontrado altos niveles de lipoperóxidos y disminución de varios sistemas antioxidantes. 46 La instalación de la insufiencia multiorgánica va precedida generalmente por un incremento en la producción de NO y ERO en el nivel de monocitos y macrófagos;47 evidencias que sugieren el papel del estrés oxidativo en la fisiopatología del gran quemado. En términos generales, la magnitud de la lesión, rapidez de la terapia de reposición hidroelectrolítica y corrección de los desequilibrios acidobásicos, estado de salud anterior, procederes para la eliminación del tejido necrótico, el tratamiento local y sistémico así como la respuesta individual de cada organismo, influyen en la evolución del SRIS en el gran quemado. Pero indudablemente la sepsis es uno de los factores más importantes, pues impone la necesidad de hospitalización prolongada en unidades cerradas, a lo cual se asocia también el fenómeno de la resistencia a los antibióticos, entre otros factores. 48 La consideración de los elementos comentados supone que los criterios de pronóstico, evolución y manejo de la conducta terapéutica pudieran complementarse con la inclusión de indicadores bioquímicos cuya base analítica se sustenta en la participación de los mediadores de la comunicación intercelular y el estrés oxidativo como eventos metabólicos de la respuesta inflamatoria sistémica del paciente quemado. En una serie de artículos posteriores se abordarán en detalle los aspectos relacionados con la biología molecular de la respuesta inflamatoria sistémica y su aplicación al modelo de la enfermedad por quemaduras. 83 SUMMARY A body burn is a violent agression that modifies all the mechanisms of organic homeostasis and is also a problem facing the medical services in the contemporary society because of its clinical and social connotation. From the physiopathological viewpoint, a burned patient develops a systemic inflammatory response characterizaed by hyperactivation of all its defense mechanisms. The de-regulation of such mechanisms leads to damage of tissues which is expressed as morpho-functional modifications of all systems. Regardless of the ethiology, initial evolution, therapeutic management and individual response, sepsis generally complicates the evolution of burn injuries. This paper sets forth the clinical and histopathological elements of burn-related diseases and comments on the molecular mechanisms of systemic inflammatory response, underlining the role of the intercellular signalling mediators. Subject headings: BURN/pathologic; SEPSIS SYNDROME/physiopathology. REFERENCIAS BIBLIOGRÁFICAS 1. Kirschbaum MS. Quemaduras y cirugía plástica de sus secuelas. La Habana: Editorial Científico Técnica, 1987;4:129. 2. Allgower M, Schoenenberger GA, Sparkes BG. Burning the largest immune organ burns 1995;21(Supp 1):537-47. 3. Pejnovic N, Lilic D, Zunic G, Colic M, Kataranovski M, Dujic A. Aberrant levels of cytokines within wound after burn injury. Arch Surg 1995;130(9):999-1006. 4. Schoenenberger GA, Baven UR, Cueni LB. Isolation and characterization of a cutaneous lipoprotein with lethal effects produced by thermal energy in mouse skin. Biochem Biophus Res Commun 1971;42:975-82. 5. Garner WL, Rodríguez JL, Miller CG, Till GO, Rees RS, Smith DJ, Remick DG. Acute skin injury releases neutrophil chemoattractans. Surgery 1994;116(1):42-8. 6. Schoenenberger GA, Cueni LB, Baven U. Isolation and characterization of a toxic lipid-protein complex formed in mouse skin by controlled thermal energy. Biochem Biophys Acta 1972;263:49-163. 7. Baron P, Traber LD, Traber DL, Nguyen T, Hollyoak M, Heggers JP, Herndon DN. Gut failure and translocation following burn and sepsis. J Surg Res 1994;57(1):197-204. 8. Allgower M, Cueni LB, Stadtler K. Burn toxin in mouse skin. J Trauma 1973;13:95-111. 9. Teodorczyk-Injeyan J, Sparken BG, Mills GB. Impairment of T cell activation in Burned Patients: A possible mechanism of Thermal injury-induced immunosuppression. Clin Exp. Immunol 1986;65:570-87. 10. Miles JM. Lipid fuel metabolism in health and disease. Curr Opin Gen Surg 1993;78:84. 11. Varriale P, Ramaprasad S. Septic cardiomyopathy as a cause of long QT syndrome. J Electrocardiol 1995;28(4):327-29. 12. Yang HM, Guo ZR, Sheno ZY. Delayed fluid resuscitation-induced bacterial translocation after lethal thermal injury. Role of oxygen free radical injury of intestinal mucosa. Chung Tlua I Hsueh Tsa Chih 1994; 74(9):552-84. 13. Kosaka K, Suzuki K, Hayakawa M, Sugiyama S, Ozawa T. Leukotoxin, a linoleate epoxide: its implication in the late death of patients with extensive burns. Mol Cell Biohem 1997;139(2):141-8. 14. Epstein MD. The Role of the gastrointestinal tract in the development of burn sepsis. Plast Reconstr Surg 1992;90:524-31. 15. Yao YM, Yu Y, Wang YP, Tian HM, Sheng ZY. Elevated serum neopterin level: its relation to endotoxemia and sepsis in patients with major burns. Eur J Clin Invest 1996;26(3):224-30. 16. Schiller HJ. Tissue perfusion in critical illnesses antioxidant Suppl):S92-102. 17. Nakae H, Endo S, Inada K, Yamada Y, Takakuwa T, Yoshida M. Plasma levels of endothelin 1 and thrombomodulin in burn patients. Burns 1996;22(8):594-7. 18. Páramo J, Rocha E. Nuevos conceptos sobre el papel de la hemostasia en la patogenia y tratamiento de la sepsis. Med Clin 1994;103:260-3. 19. Thomas RM, Letulzo Y. Clinical and etiological diagnosis of septic states. Rev Prat 1993;43(1):35-40. 20. Sanyal SC, Mokaddas EM, Gang RK, Bang RL. Microbiology of septicaemia in burn patients. Ann Burns & Fire Dis 1998;Vol XI(1):19-22. 21. Natanson C. Selected treatment strategies for septic shock based on proposed mechanisms of pathogenesis. Ann Int Med 1994;Vol. 120(9):771-83. 84 22. Weber JM, Tompkins DM, Improving Survival, infection control and burns. AACN-Clin Issues Crit Care Nurs 1993;4(2):414-23. 23. Simms H, D’Amico R. Polymorphonuclear leukocyte dysregulation during the Systemic Inflammatory Response Syndrome. Blood 1994;Vol. 83(5):1398-407. 24. Bone RC. Toward a theory regarding the pathogenesis of the Systemic Inflammatory Response Syndrome: What we do and what we do not know about cytokine regulation Crit Care Med 1996;24(1):163-72. 25. Koichiro T, Shigeki M. Early events and stimulants triggering oxidative metabolism in neutrophils. En: Knox Van D, Castianova V, eds. Cellular Chemiluminiscence CRC Boca Ratón: Press Inc., 1982;1:113-27. 26. R & D Systems. 1995 Catalog. cytokines and related reagents. Folleto comercial y promocional. R & D Systems Inc. minneapolis. USA, pág 3-5. 27. Eduards S. Cell Signalling by integrins and immunoglobulin Receptor in Primed Meutrophils. trends in Biochem Sci 1995;20(9):362-7. 28. Yao m, Sheng ZY, Tian HM, Yu Y, Wang YP, Yang HM. et al. The Association of circulating endotoxemia with the development of multiple organ failure in burned patients. burns 1995;21(4):255-8.o 29. Henderson B, Pool S. Modulation of cutokine function: therapeutic applications. En: August V, Anders M, Murad F, eds. Advances in Pharmacology. San Diego: Academic Press, 1994;25:53-96. 30. Yentis SM. Cytokines and Sepsis: Time for Reappraisal. Br J Anaesth 1995;74(2):119-20. 31. Harris BH, Gelfand JA. The immune response to trauma. Semin Pediatr Surg 1995;4(2):77-82. 32. Clancy KD, Lorenz K, Hahn E, Christiansen B, Hofmann C, Gamelli RL. Down regulation of tissue specific tumor necrosis factor alpha in the liver and lung after burn injury and endotoxemia. J Trauma 1997; 42(2):169-76. 33. R & D Systems. 1995 Catalog. Cytokines and ralated reagents. Folleto comercial y promocional. R & D Systems Inc. Minneapolis. USA, pág 49-50. 34. Mileski WJ, rothlien R, Lipsky P. Interference with the function of leukocyte adhesion molecules by monoclonal antibodies: a New approach to burn injury. Eur J Ped Surg 1994;4(4):225-30. 35. R & D Systems. 1995 Catalog. Folleto comercial y promocional. Cytokines and related reagents. R & D Systems Inc. Inneapolis. USA, pág. 187-9. 36. Yeh FL, Lin WL, Shen HD, Fang RH. Changes in serum tumor necrosis factor alpha in burned patients. Burns 1997;23(1):6-10. 37. Liu XS, Yang ZC, Luo ZH, Huang WH, Li A. A Preliminary exploration of the relationship between Tumor Necrosis Factor and monocytic in vitro production of interleukin (IL-1) and internal organ dysfunction in severely burned Patients. Burns 1995;21(1):29-33. 38. Liu XS, Luo ZH, Yang ZC, Huang WH, Li An. The Significance of changes in serum tumor necrosis factor (TNF) activity in severely burned Patients. Burns 1994;20(1):40-4. 39. R & D Systems. 1996 Catalog. Cytokines and related reagents. Folleto comercial y promocional. R & D Systems Inc. Minneapolis. USA, pág. 14. 40. Serotec 1996 Product Guide. Folleto comercial y promocional. Serotec Systems Inc Nmmeapolis. USA pág. 21-2. 41. Monden k, Arii S, Ishiguro S. Involvement of ICAM-1 expression on sinusoidal endothelial cell and neutrophil adherence in the reperfusion injury of cold preserved liver. Transplant Proc 1995;27(1):759-61. 42. Ahmed NA, Yee J, Giannias B, Kapadia B, Christou NV. Expression of human neutrophil L-Selectin during the systemic Inflammatory Response Syndrome is Partly mediated by Tumor Necrosis Factor Alpha. Arch Surg 1996;131(1):31-5. 43. Parke A, Parke DV. The Pathogenesis of the inflammatory disease: surgical shock and multiple systemic organ failure. Inflammopharmacology 1995;3/2:149-68. 44. Zhou DH. Inhibitory effect of albumin or crystalloid resuscitation on bacterial translocation and endotoxin absorption following experimental burn injury. J Surg Res 1992;52:161-6. 45. Yamada Y, Endo S, Inada K. Plasma cytokine levels in patients with severe burn injury with reference to the relationship between infection and prognosis. Burns 1996;22(8):587-93. 46. Kettle A, Winterbourn C. Assays for the chlorination activity of myeloperoxidase. Meth Enzymnol 1994;233:50212. 47. Tanjoh K, Shima A, Aida M, Tomita R, Kurosu Y. Nitric oxide and active oxygen species in severe sepsis and surgically stressed patients. Surg Today 1995;25(9):774-7. 48. Parrillo J. Septic shock in humans. Ann Intern Med 1990;113(3):227-42. Recibido: 29 de septiembre de 1998. Aprobado: 19 de julio de 1999. Dr. Félix Broche Valle. Instituto de Ciencias Básicas y Preclínicas " Victoria de Girón". Calle 146 No. 3102, Playa, Ciudad de La Habana, Cuba. 85 Rev Cubana Invest Biomed 1999;18(2):86-90 TÉCNICAS Hospital Clinicoquirúrgico " Hermanos Ameijeiras" LIPICID: SOFTWARE PARA LA DETECCIÓN, EVALUACIÓN Y TRATAMIENTO DE LAS DISLIPIDEMIAS Dr. Alfredo Nasiff Hadad y Lic. Eugenia G. Muñiz Lodos RESUMEN Por la importancia patogénica de las lipoproteínas plasmáticas, en el desarrollo de la aterosclerosis y por la alta prevalencia de cualquiera de las formas clínicas de esta última, un grupo de especialistas del Hospital " Hermanos Ameijeiras" y del Instituto Central de Investigación Digital de La Habana, Cuba, se dieron a la tarea de crear un sistema computadorizado que permite diagnosticar y tratar de manera uniforme a todo paciente dislipidémico, además promueve un adecuado estilo de vida y el conocimiento de los factores de riesgo así como las medidas para actuar sobre los factores modificables. El sistema se basa en la experiencia de la Clínica de Lípidos del Hospital "Hermanos Ameijeiras", que utiliza, entre otros, los criterios del Segundo Panel de Expertos del NCEP de EE.UU. El sistema denominado LIPICID es una aplicación Windows y la información sobre los pacientes se almacena en bases de datos tipos FoxPro, además una aplicación DOS denominada LIPINFO analiza dicha información. LIPICID detecta la dislipidemia a través del lipidograma y realiza una encuesta para detectar antecedentes personales y familiares, refleja los elementos positivos al examen físico tales como santomas, arco lipoideo, índice de masa corporal, presión arterial; recomienda los exámenes complementarios para el despistaje de causas secundarias de dislipidemias y determina la existencia de aterosclerosis preclínica. Finalmente recomienda el tratamiento no farmacológico y en una etapa posterior el tratamiento farmacológico. LIPICID es un instrumento de fácil manejo por cualquier médico, que tiene un cuestionable valor docente y constituye una útil herramienta de gran valor práctico. Descriptores DeCS: PROGRAMAS DE COMPUTACION; HIPERLIPIDEMIA/ diagnóstico; HIPERLIPIDEMIA/terapia. Las dislipidemias son un grupo de enfermedades causadas por diversos trastornos del metabolismo de las lipoproteínas del plasma y que se expresan por la elevación del colesterol y los triglicéridos.1 86 Numerosos estudios epidemiológicos han demostrado que aumenta la morbimortalidad cardiovascular en la misma medida que aumenta el valor plasmático de colesterol (Col), las lipoproteínas de baja densidad (LDL) y los triglicéridos (Tg) y disminuye el valor de las lipoproteínas de alta densidad (HDL).2,3 Las alteraciones del metabolismo de las lipoproteínas constituyen la base bioquímica en el desarrollo de la aterosclerosis, potencializada por la asociación con otros factores de riesgo.4 Por los estudios de intervención realizados se conoce que si se modifican favorablemente los valores de las lipoproteínas plasmáticas 5 mediante acciones no farmacológicas6 o farmacológicas, 7-11 utilizando estrategias poblacionales o individuales, se puede disminuir la prevalencia de cardiopatía isquémica, enfermedades cerebrovasculares y enfermedad vascular de miembros inferiores, pudiéndose alcanzar, según opinión de algunos, no sólo la prevención sino incluso la regresión de este proceso. 12-15 Debe destacarse que la cardiopatía isquémica y las enfermedades cerebrovasculares están entre las primeras causas de muerte en Cuba y en otros muchos países. Se estima que entre el 20 y el 25 % de la población cubana tiene hipercolesterolemia. 16 No existen datos acerca de los Tg séricos en la población cubana. El tratamiento de las dislipidemias tiene un carácter multidisciplinario.13 Mediante la participación de clínicos, nutriólogos, dietistas, psicólogos y enfermeras se influye educativamente sobre el paciente con el objetivo de modificar su estilo de vida.13 La variedad de situaciones posibles a las que se enfrentan los médicos en la atención a pacientes dislipidémicos conlleva soluciones distintas para cada paciente.17 La existencia de numerosos factores de riesgo de la aterosclerosis, unos ya conocidos y otros nuevos, recién categorizados, complican esta situación.18,19 Por otra parte, la existencia de dislipidemias de causas secundarias y otras de origen familiar exigen una cuidadosa evaluación del enfermo que incluye la realización de investigaciones que permitan identificar a cuál grupo pertenece,20 así como determinar la magnitud de la aterosclerosis en los 3 territorios vasculares más frecuentemente afectados. El orden necesario para dar cumplimiento a este plan, que se realiza por etapas, y que persigue la normalización de los niveles de lipoproteínas séricas y la prevención tanto primaria como secundaria de la aterosclerosis clínica, resultaría más sencillo si contara con un sistema computadorizado que permita uniformar criterios según las recomendaciones internacionales establecidas para estos casos.12,21 El Instituto Central de Investigación Digital (ICID) y el Hospital "Hermanos Ameijeiras", ambos de La Habana, Cuba, se dieron a la tarea de crear un sistema computadorizado que permitiera diagnosticar y tratar de manera uniforme a todo paciente dislipidémico, además de promover la educación sanitaria de pacientes y población en general sobre hábitos nutricionales y estilo de vida. El sistema también aporta conocimientos sobre los factores de riesgo de la aterosclerosis y la atención de aquéllos que son modificables. El sistema computadorizado desarrollado se denominó LIPICID. DESCRIPCIÓN DE LIPICID LIPICID es una aplicación Windows para el diagnóstico, evaluación y tratamiento de las dislipidemias. Toda la información sobre los pacientes manipulada por LIPICID se almacena en bases de datos tipo FoxPro. Se adiciona una aplicación DOS denominada LIPINFO mediante la cual puede consultarse y analizarse la información de dichas bases de datos. LIPICID para su trabajo se guía por la experiencia tomada de la consulta de Dislipidemias y Aterosclerosis del Hospi- 87 tal "Hermanos Ameijeiras" que utiliza las normas propuestas por el II Reporte del Panel de Expertos en Detención, Evaluación y Tratamiento del Colesterol Elevado en Adultos del Programa Nacional para a Educación del Colesterol (NCEP) de EE.UU. Por tanto, LIPICID identifica las LDL como la meta que hay que alcanzar en el tratamiento para reducir el riesgo aterogénico, tanto en la prevención primaria como secundaria y de acuerdo con el número de factores de riesgo identificados; igualmente considera los niveles bajos de HDL como un factor de riesgo idenpendiente y promueve la reducción ponderal en obesos y la actividad física como elementos básicos del tratamiento no farmacológico de las dislipidemias. LIPICID concebido para ser utilizado por el médico general, el de familia o cualquier especialista brinda las opciones siguientes: creatinina y transaminasa séricos, electrocardiograma de reposo y dosificación de hormonas tiroideas sólo en casos de sospecha clínica. Para determinar la existencia de aterosclerosis, el sistema puede indicar pruebas adicionales como el ecodoppler modo B de arterias carótidas e hilio femorales y el electrocardiograma de esfuerzo. Dado que la dieta y los ejercicios físicos constituyen la terapia inicial, LIPIDIC sólo indica fármacos cuando las condiciones individuales del paciente o la respuesta inadecuada a la dieta lo hacen necesario. La "agresividad" de la terapia depende del tipo de prevención (primaria o secundaria) y de los factores de riesgo de aterosclerosis presentes en el paciente. LIPIDIC indica tratamiento y da seguimiento al paciente permitiendo su modificación si ello fuera necesario. DIAGNÓSTICO, EVALUACIÓN Y TRATAMIENTO DE LAS DISLIPIDEMIAS A partir de un lipidograma y de un cuestionario, se determinan los factores de riesgo ateroscleróticos de un paciente y se emiten recomendaciones para actuar sobre aquellos que son modificables. Se detecta la dislipidemia a partir de un lipidograma o de un Col. El diagnóstico positivo siempre se realiza a partir de un lipidograma. Para determinar las posibles causas de una dislipidemia el sistema recomienda la realización de una encuesta donde aparecen reflejados antecedentes personales y familiares de primera línea vinculados con la aterosclerosis así como elementos del examen físico tales como xantelasmas, xantomas del tendón de Aquiles, xantomas tuberosos y de los pliegues palmares y arco lipoideo corneal. Igualmente la talla, el peso corporal y la tensión arterial. Finalmente recomienda la realización de exámenes hemoquímicos: prueba de tolerancia a la glucosa con insulinemia de 2 h, amilasa, uratos, 88 ANÁLISIS DE LOS FACTORES DE RIESGO ATEROSCLERÓTICOS INICIACIÓN DEL SISTEMA Incluye varias subopciones que permiten cambiar la unidad de medida en que se expresan los niveles de los lípidos séricos, los valores de parámetros consultados por el sistema, así como especificar la impresora y características relacionadas con el proceso de impresión. AYUDA GENERAL Suministra información sobre la forma de trabajo del sistema y explica con- ceptos propios de la especialidad médica. Incluye amplia información sobre las drogas hipolipemiantes que el sistema puede utilizar. Adicionalmente, el programa LIPINFO permite acceder a toda la información de las bases de datos (BD) generadas por LIPICID brindando facilidades que complementan sus funciones. Incorpora funciones de mantenimiento sobre la BD y facilita el análisis global de la información sobre los pacientes. ESTADO ACTUAL DEL SISTEMA LIPICID pasó satisfactoriamente las pruebas de validación del software y en estos momentos está concluyendo un ensayo clínico como requisito necesario para obtener el registro médico según la metodología orientada por el Centro de Control de Equipos Médicos (CCEM) del Ministerio de Salud Pública de la República de Cuba. Por lo que concluimos que con LIPICID cualquier médico, no necesariamente un especialista, tiene en sus manos una herramienta con buen desempeño profesional e incuestionable valor docente, que lo apoya en la detección y tratamiento de pacientes dislipidémicos. El sistema chequea la evolución del paciente y en caso de no ser satisfactoria, le recomienda modificaciones que debe realizar sobre el tratamiento del paciente. En cualquier momento, el médico puede consultar la historia clínica del paciente e imprimirla, así como obtener un resumen de ella. A través de LIPINFO el médico puede realizar sencillos análisis sobre las características y evolución de sus pacientes. El sistema constituye un instrumento de ayuda médica que tiene gran valor práctico. SUMMARY Due to the pathogenic importance of plasma lipoproteins in the development of atherosclerosis and to the high prevalence of any clinical form of this disease, a group of experts from "Hermanos Ameijeiras" Hospital and the Central Digital Research Institute in Havana endeavored to create a computarized systems for diagnosing and treating every dislipidemia patient in a uniform way in addition stimulating an adequate lifestyle and promoting the knowledge of risk factors measures to act upon the modifiable factors. The systems is based on the experience of the Lipid Clinics of "Hermanos Ameijeiras" Hospital which uses, among others, the criteria of the Second National Cholesterol Education Expert Panel in USA. The system called LIPIDIC is a Windows application, FoxPro-type databases store information on patients and a DOS application called LIPINFO analyzes such information. LIPICID detects dyslipidemia by using lipidograms and makes a survey to detect individual and family background; reflects the positive elements when making a physical examination, such as santoma, lipoid arch, body mass index, blood pressure; recommends the supplementary test for secondary causes of dyslipidemias and determines the presence of preclinical atherosclerosis. Finally LIPICID recommends the non-pharmacological treatment and in a further stage, the pharmacological treatment. LIPICID is an instrument that may be easily operated by any physician, has a questionable teaching value and is a useful tool of high practical value. Subject headings: SOFTWARE, HYPERLIPIDEMIA/diagnosis; HYPERLIPIDEMIA/therapy. REFERENCIAS BIBLIOGRÁFICAS 1. Fredrickson DS. Dyslipoproteinemia. From phenotypes to genotypes... A remarkable quarter century. Circulation 1993;87(4 Suppl):III 1-III 59. 2. Jacobs D, Blackburn, Higgins M, Reed D, Iso H, Mc Millan G, et al. Report of the conference on low blood cholestero: mortality association. Circulation 1992;86:1046-60. 89 3. Sacks FM. Desirable serum total cholesterol with low HDL cholesterol levels. An undesirable situation in coronary heart disease. Circulation 1992;84(4):1341-4. 4. Nasiff-Hadad A. Los lípidos plasmáticos. Rev Cubana Med Int 1987;26(12):1344-63. 5. ruibins HB. Cholesterol in patients with coronary heart disease. How low should we go? J Intern Med 1995;10:464-71. 6. Gotto AM Jr. Hyperlipidemia. En: Conn’s current therapy 1998. Phyladelphia: Saunders, 567-74. 7. Davignon J. Indications for lipid-lowering drugs. Eur Clin Pharmacol 1991;40(Suppl):S3-S10. 8. Di Perri T, Gevini M. Estudio comparativo de benfluorex y de clofibrato en las hiperlipoproteinemias del tipo IIa, IIb, IV. Acta Terap 1986;7:25-31. 9. Grundy SM. HMG-CoA reductase inhibitors for treatment of hypercholesterolemia. N Engl J Med 1988;319:24-32. 10. Schwartz CJ. Introduction of the probucol experience: a review of the past and a look at the future. am Cardiol 1988;62:1B-5B. 11. Havel R. Simvastatin: once a day treatment for hypercholesterolemia. Am J Med 1989;87 (Suppl 4A):1S-59S. 12. National Cholesterol Education Program. Report of the Expert Panel on Population Strategies for Blood Cholesterol Reduction: Executive Summary. Arch Intern Med 1991;151:5-19. 13. Bush TL, Riedel D. Screening for total cholesterol. Do the National Cholesterol Education Program’s recomendations detec individuals at high risk of coronary heart disease? Circulation 1991;83:1287-93. 14. Report of the National Cholesterol Education Program Expert Panel. Adut Treatment Panel II. High Blood Cholesterol. Circulation 1994;89(3):1336-431. 15. Caggiula AW, Watson JE, Milas NC, Olson MB, Kuller LH, Orchard TJ. Evaluating the efficacy of the National Cholesteol Education Program Adult Treatment Guidelines: cholesterol lowering intervention program. Preventive Med 1995;24:485-91. 16. Nasiff-Hadad A, Klaindor SB, Jiménez PR, Baldor NF. Prevalencia de hipercolesterolemia en la población adulta del municipio Habana Vieja. Rev Cubana Med Gen Integr 1992;8(4):293-306. 17. Yeshurun D, gotto AM. Hyperlipidemia; perspectives in diagnosis and treatment South Med J 1995; 88(4):379-91. 18. Sacks FM. Is there anything to add to our lipid risk actors for coronary heart disease? Am J Cardiol 1995;75:1189-95. 19. Vogel RA. Risk factor intervention and coronary artery disease: clinical strategies. Coron Artery Dis 1995;6:466-71. 20. Sierra AID. Metabolismo de los lípidos y su importancia clínica. Libro de Texto. 21. Recomendaciones del International Lipid Information bureau (ILIB) para el diagnóstico de las dislipidemias en latinoamerica. Cardiov Risk Factors 1994;3(1):6-27. Recibido: 13 de mayo de 1998. Aprobado: 19 de julio de 1999. Dr. Alfredo Nasiff Hadad. Hospital Clinicoquirúrgico " Hermanos Ameijeiras", San Lázaro No. 701, esquina a Belascoaín, Ciudad de La Habana, Cuba. 90 Rev Cubana Invest Biomed 1999;18(2):91-4 Instituto Nacional de Angiología y Cirugía Vascular INFLUENCIA DE LA CONSERVACIÓN DE LA MUESTRA EN LA DETERMINACIÓN DE LA ENDOTELEMIA Ana María Quintela Pena RESUMEN Se estudiaron 60 muestras de sangre de donantes voluntarios a las cuales se les realizó el conteo de células endoteliales en plasma el mismo día de la extracción sanguínea y se repitió el procedimiento después de conservado el plasma 24 h a 40 EC. Se encontraron diferencias significativas (p < 0,001) entre el número de células contadas en plasma fresco y el conservado en frío. Una vez leída la muestra original al microscopio, la suspensión celular es conservada en iguales condiciones y se repite sólo el conteo a las 24 h, encontrándose de igual forma valores significativamente inferiores (p < 0,05) en la muestra conservada, por lo que no se puede realizar esta determinación si no es el mismo día de la extracción de sangre. Descriptores DeCS: CONSERVACION DE LA SANGRE; DONADORES DE SANGRE; ENDOTELIO VASCULAR. El endotelio vascular tiene un papel importante en la regulación de la permeabilidad selectiva vascular, en el control de la hemostasia y en la interacción con los diferentes componentes de la sangre,1 ya que ocupa una posición crucial en la interfase sangre-tejido y está involucrado en numerosos mecanismos hemostáticos como lo es el mantenimiento de una superficie no trombogénica. La alteración de las propiedades endotelilales o el daño de la pared vascular provoca cambios protrombóticos y procoagulantes que se manifiestan en patologías agudas o crónicas como es el caso del infarto del miocardio, trombosis o aterosclerosis entre otras.2,3 Desde la década de los años 70, Bouvier 4 y Hladovec5 demostraron la presencia en sangre circulante de células no hematológicas que se identificaron como endoteliales a través de métodos de leuococoncentración; éstas se modificaron con posterioridad con el objetivo de facilitar su identificación y conteo. En la actualidad se cuenta con métodos que permiten medir estas células en sangre total, pero son de un elevado costo.6 Se ha observado que cuando ocurre una lesión endotelial ocurre una decamación celular al torrente circulatorio, lo que representa un índice7 o un marcador de dicha lesión, utilizándose para su 91 identificación la morfología típica de dichas células y/o algunos marcadores específicos.6 Por todo lo anterior, se puede considerar al endotelio vascular como un órgano sistemáticamente diseminado, por su compleja estructura y por la variabilidad de las funciones que realizan las células endoteliales (CE) como su componente fundamental.8 El propósito de nuestro trabajo es conocer la influencia que puede tener la conservación de la muestra en la determinación de la endotelemia. vistas a eliminar el posible error entre observadores. Como se muestra en la tabla 1, se encontró una diferencia significativa (p < 0,001) entre el número de CE circulantes en plasma al realizar la determinación el mismo día de la extracción y el número de CE contados cuando el plasma se conserva 24 h en frío y se monta la técnica al día siguiente de la toma de la muestra, con valores de 203 ± 7 y 125 ± 4 cel/mL PRP respectivamente, valor este último que resulta inferior. MÉTODOS TABLA 1. Número de CE el día de la toma de la muestra y en el plasma conservado 24 h en frío. Valores de la media y desviación estándar Se estudiaron 60 muestras de sangre de donantes voluntarios del Banco de Sangre del Hospital "Salvador Allende". Tomamos 10 mL de sangre colectada en un tubo de centrífuga plástico con 1 mL de citrato trisódico al 3,8 % como anticoagulante, en proporción 1:9, procediéndose según Hladovec y otros6 para el conteo de CE circulantes. Del plasma rico en plaquetas (PRP) sobrante se guardó 1 mL en frío durante 24 h en que se repitió el proceder. Una vez concluida la lectura de la muestra original, el pellet de células se conservó en iguales condiciones y se repitió la lectura a las 24 h, comparándose los resultados en ambos casos con los obtenidos en la muestra sin conservación previa. El análisis estadístico de los resultados se realizó mediante la prueba t de Student. RESULTADOS El conteo de CE circulantes es un valor individual que se eleva en plasma tras una lesión del endotelio. Es bueno señalar que en todos los casos se realizó la lectura por duplicado y por la misma persona, con 92 N 60 Día 203 ± 7 CE / mL PRP X ± DS 24 h 125 ± 4 * N: Número de muestras. PRP: Plasma rico en plaquetas. * p < 0,001. Al realizar el conteo nuevamente con el pellet celular que se conservó 24 h en frío, encontramos, como se presenta en la tabla 2, niveles inferiores de endotelemia en la muestra que ha sido conservada en relación con los niveles reales, con valores respectivos de 140 ± 6 cel y 218 ± 8 con diferencias estadísticamente significativas (p < 0,05). TABLA 2. Número de CE el día de la toma de la muestra y en el pellet conservado en frío por 24 h. Valores de la media y desviación estándar N 60 CE / mL PRP X ± DS Día 218 ± 8 N: Número de muestras. PRP: Plasma rico en plaquetas. * p < 0,05. 24 h 140 ± 6* DISCUSIÓN La disfunción de las células endoteliales como componente fundamental del endotelio puede provocar cambios en la capacidad de estas células de participar adecuadamente en los mecanismos de coagulación y fibrinolisis, predisponiéndose a la trombosis, por lo que la predicción de estas alteraciones endoteliales tiene un gran valor clínico.9 Existen diferentes métodos para medir las células endoteliales en sangre circulante, ya que su valor diagnóstico es muy importante, pues ha sido comprobado que el número de estas células circulantes se encuentra aumentado en pacientes con enfermedades malignas, trombóticas y en estadios posoperatorios.10 Algunos de estos métodos requieren de marcadores específicos y equipos de alta resolución, como el microscopio electrónico, con el que no siempre se dispone. El método de cuantificación celular utilizado por nosotros, a pesar de haber sido diseñado desde hace algún tiempo, permite dar un resultado confiable en poco tiempo por su sencilla ejecución, dependiendo fundamentalmente de la experiencia del observador al identificar las células. No obstante, en ocasiones, algunos técnicos dejan el método sin concluir completamente por determinados motivos sin conocer que no siempre es posible hacerlo. Como podemos ver en nuestros resultados, el hecho de conservar la muestra sin procesar o de detener el procedimiento al final para realizar la lectura al día siguiente de la toma de la muestra, conduce a errores en los valores de conteo, por lo tanto, el valor pronóstico y/o diagnóstico de este método se pierde si procedemos de esta manera. Por tal motivo, si en alguna ocasión no resulta posible la ejecución completa del procedimiento se deberá desechar la muestra, pues los resultados no serían utilizables por el facultativo teniendo el técnico la responsabilidad en este sentido para obtener un dato confiable. Por todo lo anterior concluimos que es indispensable realizar el conteo de CE el mismo día en que se realiza la extracción sanguínea para garantizar el valor diagnóstico de la prueba. AGRADECIMIENTOS Agradezco la colaboración prestada para la realización del presente trabajo a la Téc. Esperanza Reyes Pineda. SUMMARY Sixty blood samples from voluntary donors were studied. Plasma endothelial cells were measured in these samples the same day of blood extraction and this count was repeated after the preservation of plasma at 40 EC for 24 h. Significant differences were found (p < 0,001) between the number of counted cells in fresh plasma and that of the cold preserved plasma. Once the original sample is observed on the microscople, the cell suspension is preserved under similar conditions and only the count is repeated after 24 h. Similary, significant low values are found in the preserved samples (p < 0,05) therefore, this count must be made on the same day of blood extraction. Subject headings: BLOOD PRESERVATION; BLOOD DONORS; ENDOTHELIUM, VASCULAR. 93 REFERENCIAS BIBLIOGRÁFICAS 1. Rau L. Hypertension, endothelium and cardiovascular risk factors. Am J Med 1991;90(Suppl 2A):13S. 2. Pearson JD. Vessel wall interaction regulating thrombosis. Br Bull 1994;50(4):776. 3. Ross R. The pathogenesis of atherosclerosis: a perspective for the 1990s. Nature 1993;362:801. 4. Bouvier CA, Gaynor E, Cintron JR, Bernhardt B, Speat TH. Circulating endothelium as an indication of vas ular injury. Diathes Haemorr 1970;Suppl 40:163. 5. Hladovec J, Rossmann P. Circulating endothelial cells isolated together with platelets and the experimental modification of their counts in rats. Thromb Res 1973;3:665. 6. Hladovec J, Prerovsky I, Stanek V, Fabian J. Circulating endothelial cells in acute infarction and angina pectoris. Klin Wochenschr 1978;56:1033. 7. Takahashi H, Harker LA. Measurement of human endothelial cells in whole blood. Thromb Res 1983;31:1. 8. Augustin HG, Kozian DH, Johnson RC. Differentiation of endothelial cells: analysis of the constitutive and activated endothelial cell phenotypes. BioEssays 1994;16(12):901. 9. Blann AD. A reliable marker of endothelial cell dysfunction: does it exist. British J Hematol 1995;90:244-8. 10. Takahashi H, Harker LA. Measurement of human endothelial cells in whole blood. Thromb Res 1983;31:1-12. Recibido: 25 de noviembre de 1996. Aprobado: 19 de julio de 1999. Lic. Ana María Quintela Pena. Instituto Nacional de Angiología y Cirugía Vascular, Calzada del Cerro No. 1551. Ciudad de La Habana, Cuba. 94 Rev Cubana Invest Biomed 1999;18(2):95-103 TRABAJOS ORIGINALES Instituto Nacional de Higiene, Epidemiología y Microbiología Centro Colaborador de la OMS COMPORTAMIENTO DE LA MORTALIDAD POR BRONQUITIS CRÓNICA. CUBA, 1980-1997 Dra. Patricia Varona Pérez, Dr. Enrique Molina Esquivel, Dr. Joaquín Hechavarría Miyares, Lic Vicente I. Prieto Díaz y Dra. Bárbara Taboada Fernández RESUMEN La bronquitis crónica es una afección cuya morbilidad y mortalidad, pueden ser prevenidas. Con el objetivo de determinar el comportamiento de la mortalidad en Cuba durante el período 1980-1997, se realizó un estudio descriptivo longitudinal, que incluyó a todos los fallecidos, cuya causa básica de muerte correspondiera al código 491 (9na. Clasificación Internacional de Enfermedades). Se consideraron las variables edad, sexo, provincia y área de residencia (urbana, rural), sitio de ocurrencia de la muerte, fecha de defunción y realización de necropsia. Se utilizaron como estadígrafos las proporciones, razones, tasas crudas y específicas por grupos de edad y sexo. La tasa de mortalidad promedio del período fue 2,1 x 100 000 habitantes, mostrando una tendencia decreciente. El riesgo de morir fue mayor en los hombres y resultó mayor en los de 75 años y más. Las áreas urbanas se asociaron a un mayor riesgo que las rurales. La provincia Guantánamo fue el territorio de mayor notificación, con una proporción de necropsias que superó la media nacional. Los meses de mayor registro de fallecidos fueron julio, agosto y septiembre, correspondiente a los más cálidos en Cuba. Se hace necesario profundizar en el comportamiento de la mortalidad en el nivel local, de forma diferenciada, lo que permitirá aplicar acciones de salud para su adecuado control y prevención. Descriptores DeCS: BRONQUITIS/mortalidad; CUBA. La bronquitis crónica integra el grupo de las afecciones pulmonares obstructivas crónicas (EPOC). De acuerdo con estudios epidemiológicos realizados,1,2 la prevalencia de EPOC en general está entre el 2,5 y 3,5 % de la población adulta, ascen- diendo al 19 % en los mayores de 65 años, en los que corresponde el mayor número de casos a la bronquitis crónica. Se ha referido que las estadísticas disponibles presentan dificultades que se derivan entre otras razones, de las diferentes 95 formas de recolectar la información, desacuerdo en las definiciones y falta de adecuada información para su identificación y diagnóstico como causa de muerte.3 Definida la bronquitis crónica, como hipersecresión bronquial crónica es un proceso frecuente que afecta entre el 10 y el 15 % de la población adulta, incrementándose a 40-60 % en los mayores de 40 años fumadores de más de 20 cigarrillos al día.1 Se ha reportado que en España la bronquitis crónica, se sitúa en la cuarta causa de mortalidad, con una tasa de 33 por 100 000 habitantes, elevándose progresivamente en los grupos de edades más avanzadas (mayores de 75 años).4 En Estados Unidos, la bronquitis crónica y el enfisema pulmonar se consideran después de las cardiopatías y las esquizofrenias, como una de las causas principales de incapacidad en individuos de edades más avanzadas.5 Según estadísticas en América Latina, entre 1990 y 1992, los países con mayor número de defunciones por esta entidad fueron México(8 297), Argentina(1 897), Colombia(1 788), Ecuador (1 489) y Chile(1 318). En todos los países, el rango de edad donde se observó la mayor cantidad de defunciones, fue el de mayor que 75 años, predominando el sexo masculino excepto en el Salvador y Nicaragua.6 En Cuba, se ha estudiado la problemática de la bronquitis crónica, enfisema y asma como grupo (rubro EPOC) y éstas se ubican entre las 10 primeras causas de mortalidad general, desde finales de la década del 60,7,8 mas la entidad privilegiada en su abordaje ha sido el asma, lo que justifica esfuerzos para realizar estudios que faciliten la aplicación de acciones de forma diferenciada en nuestro medio. Por la importancia que reviste la bronquitis crónica como problema de salud, incluidas las repercusiones económicas y sociales que de ella se derivan, unido a las posibilidades de ejercer acciones efectivas de prevención y control, nos proponemos describir el comportamiento de la mortalidad por esta causa, 96 según variables seleccionadas, durante el período 1980-1997. MÉTODOS Se realizó un estudio descriptivo longitudinal que incluyó a todos los fallecidos, cuya causa básica de muerte reflejada en el certificado de defunción fuera bronquitis crónica, atendiendo a la 9na. Clasificación de Enfermedades (código 491), durante el período de 1980-1997. En todos los casos, el certificado fue expedido por un médico y se tuvo en cuenta que la cobertura de la información no se encontrara afectada en alguna zona o área geográfica. Las variables que se consideraron fueron: edad, sexo, provincia y área de residencia (urbana, rural), sitio de ocurrencia de la muerte, fecha de la defunción y realización de necropsia. Los estadígrafos utilizados fueron, proporciones, razones, tasas crudas, específicas por grupos de edad y sexo. RESULTADOS Entre el total de casos fallecidos por EPOC en Cuba, en el período comprendido entre 1980-1997, 1 de cada 4 correspondió a bronquitis crónica, proporción que disminuyó a lo largo del período, de forma más evidente a partir de 1988 (fig. 1). El riesgo promedio nacional de morir por bronquitis crónica fue de 2,1 x 100 000 habitantes, una variación porcentual del 28 % (1,8 en 1980; 1,3 en 1997) y una tendencia decreciente en los últimos 10 años. Fue en los años 1987-1988 en que se produjo la mayor variación: 111 %. Respecto a la razón de tasas por sexo masculino/femenino (fig. 2), ésta varió entre 1,1 (año 1986) y 1,9 (1992). La tasa de mortalidad se incrementó con la edad y se destacó la magnitud del incremento del riesgo en los mayores de 75 años (fig. 3) que representaron el 63 % del total de muertes. 97 1980 1981 1982 Bronquitis crónica 1983 1984 1985 1986 1987 1988 1989 EPOC (bronquitis, enfisema y asma) 1990 1991 Fig. 1. Enfermedades pulmonares obstructivas crónicas. Mortalidad proporcional por bronquitis crónica. Cuba, 1980-1997. Fuente: Dirección Nacional de Estadísticas. Minis terio de Salud Pública. 0 20 40 60 80 100 1992 1993 1994 1995 1996 1997 Años 98 1980 1986 1987 1988 Lineal (femenino) 1985 Lineal (masculino) 1984 Femenino 1983 Masculino 1982 Fuente: Dirección Nacional de Es t adísticas. Minis terio de Salud Pública. 1981 Fig. 2. Bronquitis crónica. Tasas anuales y tendencias de la mortalidad global y por sexo. Cuba, 1980-1997. 0 1 2 3 4 Tasa x 100 000 habitantes 5 1989 1990 1992 Lineal (global) Global 1991 1993 1994 1995 1996 Años 1997 99 1993 * 1994 * Fig. 3. Riesgos de morir por bronquitis crónica según grupos de edad. Cuba, 1993-1997. Fuente: Dirección Nacional de Estadísticas. Ministerio de Salud Pública. 0 5 10 15 20 25 30 35 40 Tasas x 100 000 habitantes 65-69 35-49 1995 * * 50-64 Grupos de edad (años) 75 y más 1996 * 70-74 1997 Años * 100 Ene Fallecidos Feb Abr Mas culino Lineal (masculino) Mar May Jun Fig. 4. Promedios mensuales globales y por sexos de muertes por bronquitis crónica. Cuba, 1980-1994. Fuente: Dirección Nacional de Estadísticas. Minis terio de Salud Pública. 0 5 10 15 20 25 Femenino Lineal (femenino) Jul Ago Sep Oct Dic Meses Global Lineal (global) Nov La proporción promedio de necropsias realizadas, durante el período 1990-1995, fue del 25 %, variando entre el 9 % (1990) y el 40 % (1993). En todos los años se realizaron más necropsias a hombres que a mujeres. La mortalidad por provincias en el período 1993-1997, mostró que Guantánamo fue el territorio de mayor notificación de bronquitis crónica; y la proporción promedio de necropsias confirmada en ella fue del 36,2 %. En las áreas urbanas fue mayor el riesgo de morir por esta causa que en las rurales. Otra de las variables descritas fue el sitio de ocurrencia de la muerte, que mostró que a partir del año 1989, aproximadamente el 30 % de ellas ocurrió fuera de una institución de salud. Respecto a la mortalidad por meses los valores mínimos se observaron en diciembre y los máximos en enero, ubicándose por encima de la línea de tendencia los registros en los meses de mayo, julio, agosto y septiembre (fig. 4). DISCUSIÓN El riesgo de morir por esta afección en Cuba, se consideró bajo, si se tiene en cuenta que el principal factor de riesgo lo constituye el tabaquismo, cuya prevalencia ha sido históricamente elevada en la población de ambos sexos, de 17 años y más (mayor que el 37 %) durante todo el período analizado, según reportes anteriores de Encuestas Nacionales del Instituto Cubano de Investigaciones de Orientación de la Demanda Interna (1988 y 1990), incluida la Primera Encuesta Nacional de Factores de Riesgo y Actividades preventivas de enfermedades crónicas no transmisibles (ENFR) realizada en el año 1995, a lo que se añade que tanto en las viviendas como en los centros laborales, la exposición al humo de tabaco ambiental es alta y los efectos negativos sobre la salud, resultado de ambas exposiciones, activa y pasiva pueden ser similares, 9 aun teniendo en cuenta la susceptibilidad individual, la edad, la predisposición genética y la historia de trastornos respiratorios como el asma10 entre otros factores. Durante el período analizado no se produjeron cambios en los criterios diagnósticos de la enfermedad, ni en la clasificación internacional de enfermedades utilizada (9na. CIE), ni variaciones sustanciales en la población que expliquen el supuesto incremento notificado en los años 1988 y 1990, que se ubicó fuera del comportamiento esperado. La baja proporción de necropsias realizadas entre los fallecidos por esta causa, según datos de la Dirección Nacional de Estadísticas del Ministerio de Salud Pública de Cuba, no permite asegurar que las cifras sean completamente reales, y que otras muertes, aun causadas por bronquitis crónica se notifiquen con otros diagnósticos, como asma bronquial, principalmente en personas de edades avanzadas. Una situación similar ha sido reportada en Estados Unidos de Norteamérica durante el período 1960-1995.11 Cabe señalar que durante el período, fue justamente la provincia de Guantánamo, la que superó la media nacional de necropsias realizadas y la que notificó mayor tasa de mortalidad. Ha sido reportado en otras publicaciones,12,13 mayor riesgo de morir en los hombres a causa de mayor prevalencia de tabaquismo entre ellos y mayor exposición laboral, lo que concuerda con estudios realizados de prevalencia de síntomas en 8 zonas de la ciudad de La Habana.14 El comportamiento de la mortalidad según grupos de edad, resultó consecuente 101 con otros reportes15 y se explica por la presencia tanto de factores intrínsecos como el envejecimiento tisular, como de factores extrínsecos, entre los que se incluyen la exposición más prolongada a elementos exógenos. Resultó contrastante encontrar las mayores tasas de mortalidad en las provincias con menor prevalencia de tabaquismo según la ENFR, sin que por ello pretendamos establecer relaciones causales, al tratarse de un proceso patológico con prolongado período de latencia. La mayor proporción de muertes en áreas urbanas se ha asociado a la mayor prevalencia de tabaquismo activo y pasivo en éstas, así como a la mayor exposición a la contaminación atmosférica entre otros factores. Este exceso de riesgo de morir, ha sido también observado en no fumadores.16 Los registros de la mortalidad según meses, hace sospechar posibles retardos en las notificaciones a fines de año, las que pasarían a computarse en el mes de enero. Una vez excluido este mes, se aprecia un aumento de la mortalidad en los meses de julio, agosto y septiembre, correspondiente a los meses más cálidos en nuestro país,17 a diferencia de lo que ocurre en los países templados y fríos en los que durante los meses de otoño e invierno se elevan las muertes por bronquitis crónica, relacionado con la mayor frecuencia de infecciones respiratorias condicionadas por la mayor exposición a la contaminación biológica y química del aire en interiores. Consideramos que aunque esta afección tiene características comunes con otras como el enfisema pulmonar, muestra diferencias que la particularizan a escala local, lo que requiere profundizar en su estudio, en atención a la posibilidad de modificar su tendencia, fundamentalmente a través de la aplicación de acciones de salud que permitan su adecuado control y prevención. SUMMARY Chronic bronchitis is a disease which mortality and morbidity can be prevented. With the objective of determining the situation of mortality in Cuba from 1980-1997, a longitudinal descriptive study was performed, which covered all those deceased whose basic cause of death was included in Code 491 (Ninth International Classification of Diseases). Variables such as age, sex, province and area of residence (urban, rural), site of occurrence of death, date os death and necropsy were considered. Proportions, ratios, crude & specific rates by age groups and sex were used. The average mortality rate of this period was 2. 1 x 100 000 population, showing a decreasing trend. The risk to death was higher in males, mainly in those aged 75 years and over. The urban areas were more linked to a high death risk than rural areas. Guantanamo province was the most affected territory with a necropsy rate exceeding the national average figure. The highest number of deaths occured in July, August and September which are the warmest months in Cuba. It is required to go deeper into the situation of mortality at a local level, in a differentiated way, so as to put health actions into practive for adequate control and prevention. Subject headings: BRONCHITIS/mortality; Cuba. REFERENCIAS BIBLIOGRÁFICAS 1. Hodkin J. Chronic obstructive pulmonary disease. Clin Chest Medicine 1994;11:363-571. 2. ERS Consensus Statement. Optimal asessment and management of chronic obstructive pulmonary disease. Eur Repir J 1995;8:1398-420. 102 3. Colle J. Chronic non-specific lung diseases (CNSLD) and asthma. epidemiology of diseases. Blackwell Scientific Publications 1982; ISBN 0-632-00686-2. 4. Sobredillo V. Enfermedad Pulmonar Obstructiva Crónica: Enfoque actual. En EPOC: perspectivas actuales. Editor: J Castillo. Madrid 1995;3-17. 5. Anthonisen NR. Hypoxemia and Oxigenotherapy in the COPD. Am Rev Respir Dis 1994;126:729. 6. OPS. Estadísticas de Salud en las Américas. Washington 1995. 7. MINSAP. Informe de Balance del MINSAP 1997. Marzo 1998. Cuba. 8. MINSAP. Anuario Estadístico 1997. Ciudad Habana. Cuba. 9. EPA (Environmental Politic Agency). Respiratory health effects of passive smoking: Lung cancer and other disorders. 600/6-90/006F. December 1992. Office of Health and Enviromental Assesment or Research and Development. U.S. Enviromental Protection Agency. Washington D.C. 10. Buist AS. Asthma as a risk factor for chronic airways disease. Chest 1989;96:314S-5S. 11. Guidotti TL, Jhangri G. Mortality from airways disorders in Alberta, 1927-1987: An expanding epidemic of COPD, but asthma shows little change. J Asthma 1994;31:277-90. 12. Guidotti TL. Trends in mortality from chronic obstructive pulmonary disease in Alberta; Back to the future? Can Resp J 1995;2:97-103. 13. Thom TJ. International comparisons in COPD mortality. Am Rev Resp Dis 1989;140:S27-34. 14. Molina E, Pita G, Monterrey P. Contaminación atmosférica, otros factores ambientales y morbilidad respiratoria en la tercera edad. En: Memorias XIV Congreso Interamericano de Ingeniaría Sanitaria y Ambiental. Buenos Aires, 1994. 15. Guidotti TL. Mortality from airflow disorders in Alberta: Studies on asthma and chronic obstructive pulmonary disease. Alberta studies in Occupational Health, No. 18. Septiembre 1996. 16. Guidotti TL. Ambient air quality and human health: currents concepts. Part 2 Can Respir J 1996;3(1):29-39. 17. Instituto Cubano de Geodesia y Cartografía. Atlas climático de Cuba. La Habana: Instituto cubano de Geodesia y Cartografía; 1987. Recibido: 1ro. de junio de 1999. Aprobado: 19 de julio de 1999. Dra. Patricia Varona Pérez. Instituto Nacional de Higiene, Epidemiología y Microbiología, Infanta No. 1158 Ciudad de La Habana, Cuba. CP 10300. 103 Rev Cubana Invest Biomed 1999;18(2):104-10 Instituto de Ciencias Básicas y Preclínicas "Victoria de Girón" CRECIMIENTO Y COMPOSICIÓN CORPORAL DE LAS CRÍAS DE RATAS SOMETIDAS A RESTRICCIÓN ALIMENTARIA Dra. Daisy Valiente Bisset, Dra. Cristina Alfonso Zerquera, Dra. Manuela Gilda Bernardo Fuentes, Dr. José R. Molina García y Dra. Valentina T. González Cabrales RESUMEN Nos propusimos en el presente trabajo, estudiar la influencia de la restricción alimentaria de las ratas gestantes sobre el crecimiento y el contenido de grasas y agua corporal de la prole durante los primeros 21 días de vida. El experimento se realizó estudiando 40 ratas Sprague Dawly hembras, gestadas y separadas en grupo control y restringidas; estimamos el contenido de agua y de grasa de las crías desde el nacimiento hasta el destete, mediante la desecación de la muestra y la extracción de solventes orgánicos de la grasa. El peso y la longitud corporal de los hijos de madres restringidas son menores desde el destete que los de los controles. El contenido de grasa es menor al nacimiento de las crías del grupo restringido, lo cual se recupera al final de la primera semana. El contenido de agua no difiere en ambos grupos estudiados. La cantidad de grasa corporal en las crías de madres restringidas depende más del peso que de la edad. Descriptores DeCS: AGUA CORPORAL; PESOS Y MEDIDAS CORPORALES; EMBARAZO. Los efectos adversos de la desnutrición materna durante el embarazo en la madres y sus descendientes constituyen un gran problema de la salud pública, especialmente entre los países en desarrollo, con variado resultado en el crecimiento fetal, morbilidad, mortalidad y en el desarrollo inmediato y a largo plazo de los niños.1 Durante mucho tiempo se pensó que el feto tenía la capacidad para obtener de la madre todas las capacidades dietéticas con entera independencia de su estado nutricional. Estudios experimentales en 104 animales de laboratorio, sobre todo empleando ratas, han tratado de comprobar esa hipótesis.[Peña R. de la. El ratón y la rata como animales de laboratorio. ISCM-H:1982.]2,3 Estos estudios indican que la madre puede movilizar nutrientes almacenados durante la fase anabólica de la preñez. La movilización de esos nutrientes no incrementa significativamente el crecimiento fetal; sin embargo, se sostiene la hipótesis de que la madre es capaz de compartimentar los nutrientes disponibles durante la malnutrición y prevenir la depleción severa de sus reservas por el feto.3,4 Algunos autores se oponen a la hipótesis de la compartimentalización, planteando que durante la restricción de alimentos el feto no puede utilizar las reservas de nutrientes maternos para prevenir el retardo en el crecimiento.5,6 En relación con las investigaciones en animales de experimentación, se ha encontrado que grandes trastornos metabólicos afectan el desarrollo y la composición corporal.3,5,7-9 Estudiando la desnutrición inducida en ratas durante la gestación se reportan disminuciones en el peso corporal, en la talla,2,10-12 y sobre el contenido de grasa de la camada.3,13 El presente trabajo tiene como finalidad el estudio del efecto de la restricción alimentaria de la rata gestante sobre el crecimiento, el contenido de grasa y agua corporal de la prole durante los primeros 21 días de vida. MÉTODOS Se emplearon 40 ratas Sprague Dawley, hembras vírgenes cuyos pesos oscilaron entre 180 y 220 g y 8 machos de la misma línea; ambos procedentes del Bioterio del ISCM-H. El diseño de esta investigación consistió en el estudio del peso, la longitud corporal total, el contenido de agua y de grasa corporal de la prole de ratas que recibieron dieta libre, y de ratas restringidas durante toda la gestación, desde el nacimiento hasta el destete, mediante la desecación de la muestra y la extracción de grasa por lavado con un solvente orgánico. Los animales fueron apareados durante la noche y el inicio de la gestación fue determinado por la presencia de espermatozoides en el "smear" vaginal a la mañana siguiente, considerándose ese día como día "0" de la gestación. De las 40 ratas gestadas, 20 fueron escogidas al azar pasando al grupo control, que recibió dieta libre durante todo el embarazo, y las 20 restantes, fueron restringidas al 60 % del consumo promedio de los controles durante todo el embarazo. Durante la gestación fue controlado solamente el peso de la rata gestante. En el primer día, posterior al parto, fue registrado el número de la camada, el peso corporal y la longitud corporal total. Se dejaron sólo 8 crías (4 hembras y 4 machos), por cada camada y se repitieron las mediciones a los 7, 14 y 21 días (destete), después del nacimiento de la progenie de ambos grupos, control y experimental. Durante el período posnatal de experimento, ambos grupos de ratas recibieron alimentación ad libitum. El alimento utilizado en todo el experimento fue el pienso especial para ratas (ratonina), de composición conocida y que aportaba un promedio de 22 % de proteína y 4,3 % de grasa. El estudio del contenido de grasa fue igualmente realizado al nacimiento, a los 7, 14 y 21 días de edad para lo cual 20 madres de cada grupo fueron subdivididas en 4 subgrupos, constituido por 5 madres cada uno (40 crías), las que fueron sacrificadas a las edades señaladas. Una vez sacrificados los animales en una atmósfera de cloroformo, fueron pesados, triturados y transferidos a una estufa convencional a 60 EC hasta su total desecación, la cual se consideró completa cuando no hubo diferencia entre 2 pesadas de control sucesiva. La diferencia entre el peso inicial de la camada y el peso seco representa el contenido de agua. La determinación de la grasa corporal se realizó mediante extracción con un sistema de solventes orgánicos dietil éter-éter de petróleo 1:1, efectuándose dicha extracción en un equipo SOXHLET, según el mé- 105 todo descrito por la Asociación Oficial de Químicos Agrícolas en 1980.9 El contenido de grasa se expresa como porcentaje de peso corporal inicial (peso húmedo). Análisis estadístico de los datos: Se calcularon los estadígrafos descriptivos del peso, la longitud corporal (LC), el porcentaje de grasa corporal, la masa seca (%) y el contenido de agua, en los 2 grupos de estudio, en los 4 momentos del experimento. Se realizaron comparaciones binarias para cada variable dentro de cada tiempo del experimento entre los grupos de estudio. Se ajustó un modelo lineal general para el contenido de grasa en el que se analiza el efecto de la restricción alimentaria de la madre, la edad y el peso corporal. El procesamiento de los datos se autorizó con el auxilio de la mi-crocomputadora IBM-PC y se utilizó el paquete estadístico comercial Systat. RESULTADOS En la tabla 1 se presentan los valores medio y la desviación estándar del peso, la longitud corporal y el contenido de grasa de los animales de ambos grupos (control y restringido), en los 4 tiempos del experimento, así como la comparación binaria entre grupos en cada momento del estudio. Se observa un crecimiento casi lineal del peso y la longitud desde el nacimiento hasta los 21 días, tanto en el grupo control como en el restringido; los valores promedio son siempre mayores en el primer grupo que en el segundo. Con respecto al contenido de grasa se observa poco incremento durante la primera semana, tanto para el grupo control como para los restringidos, aunque estos últimos muestran una recuperación evidente, pues de valores significativamente menores al nacimiento, alcanzan valores casi idénticos a los 7 días. Entre los 7 y 14 días el contenido de grasa aumenta más de una vez y TABLA 1. Valores medio (DS) y comparación estadística en los distintos tiempos de las variables estudiadas en ambos grupos Variables Grupo Nacimiento X(DS) 7 días X(DS) 14 días X(DS) 21 días X (DS) Peso n = 40+ Control Restr. t (p) Control restr. t (p) Control restr. t (p) 7,03 (0,51) 5,68 (0,57) 3,94 (0,004)* 61,12 (3,97) 60,84 (3,19) 0,12 (NS) 0,47 (0,11) 0,28 (0,07) 3,07 (0,015)+ 14,72 (1,43) 12,99 (1,77) 1,71 (NS) 101,24 (10,28) 94,57 (4,84) 1,31 (NS) 0,79 (0,37) 0,78 (0,28) 0,048 (NS) 27,02 (0,66) 25,63 (1,99) 1,48 (NS) 148,06 (3,18) 134,20 (4,97) 5,25 (0,001)* 1,93 (0,48) 2,14 (0,47) 0,69 (NS) 44,61 (4,59) 37,04 (4,45) 2,65 (0,029)* 185,64 (16,58) 166,92 (9,17) 2,22 (0,058)* 2,96 (0,98) 2,37 (0,63) 1,13 (NS) Longitud corporal n = 40+ Contenido de grasa % n= 160++ Las mediciones del peso y la longitud corporales fueron realizadas en los mismos 40 animales en los 4 tiempos del estudio. La determinación del contenido de grasa, obviamente, fue realizada en 4 grupos diferentes de 40 animales, cada uno. * Significación estadística p < 0,05. NS: No significación. + ++ 106 media en ambos grupos. En la tercera semana disminuye el ritmo de acumulación de grasa en los restringidos en tanto que en los controles su crecimiento es lineal. Es interesante el hecho de que la diferencia de la longitud corporal entre controles y restringidos no es significativa al nacer, pero se acentúa, alcanzando significación a los 14 días. La diferencia de peso es pequeña hasta los 14 días pero aumenta hacia el destete a favor de los controles. La tabla 2 corresponde a los valores medio y la desviación estándar de la masa seca y el contenido de agua de ambos grupos en los 4 tiempos del experimento, así como la comparación binaria entre grupos de cada momento del experimento. Encontramos con respecto a la masa seca, diferencias significativas al nacimiento entre ambos grupos, manteniendo valores superiores en el grupo control hasta los 21 días, aunque no existen significativas entre controles y restringidos. Al analizar los valores medio del contenido de agua observamos valores ligeramente superiores desde el nacimiento hasta el destete en el grupo control, no obstante desde el punto de vista estadístico no existen diferencias significativas entre ambos grupos. A continuación se presentan los resultados del modelo general para el contenido de grasa en el que se analizan el efecto del tiempo y la restricción, así como el peso corporal. (n= 40) Fuente de variación F Grupo (Restringido vs Control) 4,45 Edad 6,01 Peso 42,74 P 0,042 0,002 0,000 Se pone de manifiesto que el mayor factor adiposidad es el peso corporal, se observa también un efecto importante de la edad y una discreta influencia de la restricción alimentaria de la madre durante el embarazo. TABLA 2. Valores medio (DS) y comparación estadística de la masa seca y el contenido de agua entre los 2 grupos estudiados Variables Grupo Nacimiento X(DS) Masa seca Control. restr. t (p) Control restr. t (p) 3,73 (0,34) 2,15 (0,23) 8,44 (0,14)* 3,77 (1,39) 3,53 (0,42) 0,36 (NS) Contenido de agua 7 días X(DS) 4,97 (1,14) 3,80 (0,87) 1,82 (NS)* 9,73 (0,45) 9,18 (1,02) 1,15 (NS) 14 días X(DS) 9,23 7,55 1,23 17,9 18,1 1,31 (2,54) (1,67) (NS) (2,59) (0,93) (NS) 21 días X (DS) 16,29 11,91 1,62 28,3 25,1 1,85 (5,74) (1,87) (NS) (2,55) (2,83) (NS) Significación estadística p < 0,05. NS: No significación. * 107 DISCUSIÓN Los efectos adversos de una alimentación inadecuada no admiten ninguna clase de dudas. Aunque la magnitud del daño es muy variable en diferencia de numerosos factores adicionales, la reducción del peso al nacer, el acortamiento de la gestación y el incremento materno y fetal son consecuencias invariables de la desnutrición.1,8 Nuestros resultados indican objetivamente que la restricción alimentaria influye negativamente sobre el peso, pues muestra una reducción significativamente del orden del 20 % en el momento del nacimiento, lo cual concuerda con datos de otros autores,2,4,8,12,14 mientras que la longitud corporal apenas afecta, a diferencia del contenido de grasa que muestra valores significativamente menores en una proporción del 40 % con respecto a los animales del grupo control. Aunque observamos que existe una disminución marcada del peso y la grasa corporal no podemos interpretar este hecho como la simple relación peso-adiposidad, dada porque la reducción del peso sea a expensas de la reducción de grasa, pues en esta etapa de la vida la grasa significa una fracción insignificante del peso corporal, aunque de hecho en esta relación pesoadiposidad puede influir la reducción de otros nutrientes no grasos lo cual se ve evidentemente en la reducción de masa seca en aproximadamente 1 g de porcentaje en los animales del grupo restringido en relación con el grupo control. Después del nacimiento y hasta los 21 días de edad, nosotros encontramos que los valores medio del peso son más bajos en las crías de ratas restringidas que en las del grupo control, por lo que existe significación estadística al momento del destete; pensamos que ésta puede estar relacionada con el hecho de que la composición 108 corporal de la madre, los patrones de alimentación y sus propios ajustes metabólicos son por supuesto diferentes en controles y restringidos. En efecto, es evidente que las madres gestantes sometidas a restricción alimentaria, a las cuales luego del parto y durante la lactancia se les realiza un cambio en su alimentación con respecto a la cantidad de alimento suministrado, puede conducir a cambios metabólicos en éstas que provocarían modificaciones notables en cuanto a composición y variabilidad de la leche materna en un período de vida en el cual la lactancia natural es la vía principal de administración de nutrientes a sus hijos, razón que parece influir directamente en cuanto a la menor ganancia de peso de los hijos de madres restringidas durante la gestación. Es importante advertir que en lo que se refiere a la longitud corporal, el grupo experimental no presentó diferencias significativas con respecto al grupo control en el nacimiento, lo cual difiere de los resultados encontrados por otros autores. 5,8,14 No obstante, observamos que a pesar de que existe un incremento progresivo con el tiempo, las diferencias comienzan a evidenciarse ya a los 7 días después del nacimiento, haciéndose significativas a los 14 y 21 días posnatales, resultados que coinciden con los de otros autores,2,8,12,14 que refieren menor longitud corporal al destete en los hijos de madres restringidas, por lo que hemos considerado que durante el período de lactancia, el cambio de alimentación de las madres parece producir notables cambios en la composición corporal con grandes transformaciones metabólicas que repercuten directamente sobre las dimensiones corporales de las crías de ratas restringidas que reciben como alimentación la secreción láctea de sus madres. Con respecto al contenido de gra- sas no hemos tenido referencias en relación con esta variable como indicador de adiposidad; solamente algunos autores13 han reportado un estudio durante las primeras horas, de la grasa corporal después del nacimiento en las crías de ratas de varios grupos de animales, dentro de ellos subnutridos durante la gestación, los cuales presentaron reducción del contenido de grasa en las primeras horas con respecto a los controles, resultados que son comparables con los nuestros al momento del nacimiento. Al analizar el comportamiento del contenido de grasa observamos incrementos muy pequeños durante la primera semana de vida, lo cual podría estar en relación con los profundos ajustes metabólicos que acompaña la transición de la vida intrauterina a la extrauterina, así como a las grandes demandas de calor en una etapa de ajustes fisiológicos en los mecanismos de termorregulación, en el cual juega un importante papel el tejido adiposo multilocular que predomina en esta etapa de la vida [Castillo MM. Desarrollo prenatal del tejido adiposo. Estudio descriptivo y significado de su distribución en el humano, la rata y el ratón. Tesis de Especialización ISCM-H. La Habana; 1988.] Debemos observar que el ritmo de crecimiento de la grasa corporal es mucho mayor en los hijos de madres restringidas, lo que evidencia claramente un mecanismo de regulación nutricional, ampliamente reconocido en la literatura médica. Transcurrida la etapa crítica de la primera semana de vida hay un rápido cre- cimiento de la grasa corporal, cuya proporción al momento del destete casi sextuplica los valores del nacimiento y cuadruplica los valores encontrados a los 7 días. En cuanto al contenido de agua nuestros resultados muestran diferencias significativas entre los grupos estudiados, lo cual es comparable con los resultados de otros autores.15 Pensamos que este hecho se deba a que las madres del grupo experimental tenían restricción de alimentos pero no de ingestión de agua. El análisis multifactorial de la influencia de la restricción, la edad y el peso corporal, pone de manifiesto que el mayor efecto corresponde por mucho al peso corporal, lo cual significa que un animal cualquiera en cualquiera de los grupos y con una edad determinada, a mayor peso le corresponde mayor grasa corporal; igualmente dentro de los grupos, animales de igual peso les corresponde mayor cantidad de grasa corporal a los de mayor edad, resultados que coinciden con los de otros autores.3,16 El efecto de la restricción alimentaria materna apenas alcanza significación estadística en el modelo. Obviamente es de esperar que así ocurra, ya que estamos analizando el efecto de la restricción crónica de la madre durante toda la gestación sobre el crecimiento y el desarrollo posnatal. Hecho que puede deberse en gran medida a los mecanismos de reajustes metabólicos en la madre depletando sus reservas de nutrientes, mientras que el crecimiento fetal se afecta poco; estos resultados coinciden con los reportes de otros autores. 2,14,17 SUMMARY In the present paper, we intend to study the impact of dietary restriction of pregnant rats on the growth, body fat and water content of their offspring during their first 21 days of life. The experiment was performed with 40 Sprague Dawly female pregnant rats which were included in a control group and in a dietary restricted group; we estimated the fat and water contents of the offspring since they were born until they were weaned by means of 109 sample dissecation and extraction of organic fat solvents. Body weight and lenght of pups of mothers on dietary restriction were lower than those of the control group at weaning. Fat content was lower when the offspring of the restricted food group was born but this is balanced at the end of the first week. Water content did not differ in both studied groups. Body fat volume of the progeny of mothers on food restriction is more weight than age-dependent. Subject headings: BODY WATER; BODY WEIGHTS AND MEASURES; PREGNANCY. REFERENCIAS BIBLIOGRÁFICAS 1. World Health Organization. The effects of nutrition during pregnancy on the offspring. Draft synopsis of the IPA/WHO workshop, Barcelona, 1980. 2. Alfonso C, Melo M, González MC, Valiente D. Effectos adversos de la restricción alimentaria gestacional en ratas sobre el crecimiento de las crías. Rev Cubana Invest Biomed 1995;14(2):131-9. 3. Fiorotto ML, Davis TA, Schoknecht P, Mersmann HJ, Pond WG. Both maternal over and undernutrition during gestation increase the adiposity of young adult progeny in rats. Obes Res 1995;57(3):469-76. 4. Anderson GA, Ahokas RA, Lipschitz J, Diltz PV. Effects of maternal dietary restriction during pregnancy on maternal weight gain and fetal birth weight in the rat. J Nutr 1980;110(5):883-90. 5. Lederman SA, Rosse P. Effects of obesity. Food restriction and pregnancy on fetal and maternal weight and on body composition in rats. J Nutr 1981;111(12):2162-71. 6. Boxwell J, Ayson P, Remenofsky M. Growth and metabolic parameters in pups of undernourished lactating rats. Physiol Behav 1995;57(3):469-75. 7. Azain MJ, Hausman DB, Kasser TR, Martin RJ. Effect of somatotropin and feed restriction on body composition and adipose metabolism in obiso Zucker rats. Am J Physiol 1995;269. (1Pt 1):E 137-44. 8. Barbosa L, Santiago S de. Efecto de la restricción en el consumo de alimento de la rata adulta sobre el crecimiento y la composición tisular de la cría lactante. Arch latinoam Nutr 1984;44(2):98-104. 9. Park CS, Baik MG, Keller WL, Slanger WD. Dietary energy restriction mediated growth and mammary development in rats. J Anim Sci 1984;72(9):2319-24. 10. Anokas RA, Lahaye EB, Anderson GD, Lipschitz J. Effect of maternal dietarty restriction on fetal growth an placental transfer amino isobutyric acid in rats. J Nutr 1981;11(12):2052-8. 11. Zartarian GN, Galler JR, Munro HN. Marginal protein deficiency in pregnant rats. I changes in maternal body composition. J Nutr 1980;110(7):1291-7. 12. Pond WG, Jung FW. Nature body weight and life span of male female progeny of primaporous rats fed at low protein or adequate diet throughout pregnancy. J Nutr 1981 111(11):1949-54. 13. Chung JA, Cha M, Oh W. Growth and fatty acid metabolism in experimental growth retardation: effect of posnatal nutrition in rat. J Nutr 1986;116(6):1080-7. 14. González JC, Alfonso C, Melo M, Valiente D, Payne S. Variaciones del peso y la talla de la prole de ratas sometidas a restricción alimentaria moderada en distintos momentos de la gestación. Rev Cubana Cienc Vet 1989;20(2):219-28. 15. Rucklidge GJ. Diferances in body compositions, growth and food intakes between mice which have been selected for small and large body size. Br J Nutr 1981;46(3):441-50. 16. Shemmel R, Mickelson O, Tolgany Z. dietary obesity in rats: influence of diet, weight, age and sex on body composition. A J Physiol 1969;216(2):373-9. 17. Alfonso C, González MC, Melo M, Bernardo MG, Payne S. Variaciones de los diámetros craneanos en la prole de ratas sometidas a restricción alimentaria moderada en distintos momentos de la gestación. Rev Cubana Cienc 1989;20(2):211-8. Recibido: 13 de mayo de 1998. Aprobado: 19 de julio de 1999. Dra. Daisy Valiente Bisset. Instituto de Ciencias Básicas y Preclínicas "Victoria de Girón", 146 esq. Ave 31, Playa, Ciudad de La Habana, Cuba. 110 Rev Cubana Invest Biomed 1999;18(2):111-6 Instituto de Ciencias Básicas y Preclínicas "Victoria de Girón" DUPLO Y: ¿ESTIGMATIZACIÓN GENÉTICA? MC. Héctor I. Pimentel Benítez, Dr. Jaime Fajardo Castellanos y Lic. Julia García Capote RESUMEN La estigmatización genética constituye un hecho que cobra fuerza con los avances de la biología molecular, y el descubrimiento de cada nuevo gen, que aportan las investigaciones del proyecto Genoma Humano, pero existen síndromes como el XYY, que desde su primera descripción se asocian a las formas de conducta de violencia. En el presente trabajo, realizamos una valoración de las principales ideas que acerca de este síndrome y su relación con la criminalidad, aparecen en los reportes hallados en la base de información MEDLINE, en el período de 1990 a mayo de 1998; donde se destacan la insuficiencia de datos personales de los sujetos descritos; éstos son de vital importancia para poder apreciar el papel que el medio social (ambiente) desempeña en la aparición de este fenotipo. Realizamos además, nuestra valoración sobre la ubicación de esta línea problémica entre lo biológico y lo social, así como algunas sugerencias y/o consideraciones; teniendo en cuenta los ilimitados campos, sobre todo desde el punto de vista ético que tiene el síndrome XYY. Descriptores DeCS: CARIOTIPO XYY; VIOLENCIA; ESTEREOTIPO. En el proceso de edificación de la sociedad cubana, concierne a toda la ciudadanía hacer retroceder paso a paso la criminalidad y sus causas, investigar con profundidad la estructura y otras causas de este negativo fenómeno social, y como tal la investigan, en diferentes aspectos, las ciencias sociales, la medicina y otras ciencias.1 En la literatura médica se consigna con marcado y repetido hincapié, la relación existente entre el cariotipo XYY y las formas de conducta violenta, extremo éste por el cual se le ha dado en llamar, "síndrome de la criminalidad". Pese a la amplia bibliografía existente al respecto, es innegable el desconocimien- to de este síndrome genético en nuestro medio, que evidentemente trae consigo un problema, que ha surgido de un conocimiento preexistente, constituido por datos, generalizaciones empíricas, teorías y técnicas, que le dan el carácter científico, y como tal parte de una demanda social, que implica una búsqueda teórica y metodológica, para dar respuesta a una incógnita no resuelta por la ciencia. El presente estudio encamina sus objetivos a dar a conocer las principales ideas que en la bibliografía consultada hemos encontrado y que relacionan la presencia de este síndrome cromosómico con las formas de conducta violenta, haciendo nuestra valoración del lugar que debe ocupar 111 este tema en la problematización científica entre lo biológico y lo social, así como ofrecer algunas consideraciones éticas (estigmatización genética del síndrome) respecto a él. XYY, CLÍNICA Y ÉTICA El síndrome XYY es conocido desde que Jacob y otros realizaron un muestreo entre pacientes que eran mentalmente subnormales, y que habían sido internados en instituciones especializadas a causa de su propensión a la violencia y a la criminalidad, además se ha comprobado su gran frecuencia entre los reclusos de cárceles para individuos peligrosos, donde alrededor del 3 % de estos sujetos son XYY.2 Clínicamente estos individuos corresponden a varones de elevada talla, (mayor que 1,80 m), con inteligencia algo inferior a lo normal, temperamento agresivo, trastornos de la personalidad, inestabilidad, irresponsabilidad; algunos autores plantean que desde los 13 años se les suele encontrar en los tribunales de menores, como delincuentes juveniles.2-5 La mayor frecuencia con que aparecen en las poblaciones penales y hospitales psiquiátricos de seguridad puede ser indicio de su relación con su comportamiento agresivo, psicopático, agresivo y antisocial.6,7 Esta relación ha despertado gran interés entre el público. Se calcula que estos varones presentan 6 veces más probabilidades de ser encarcelados que los sanos (XY). Cuando en las revisiones en el período neonatal, o incluso prenatal, se reconocen niños XYY es difícil de decidir el modo de enfocar la situación, a causa de la estigmatización genética de este síndrome. Los varones XYY se originan por falta de disyunción paterna en la II división 112 meiótica, que produce espermatozoides cromosómicamente YY. Este proceso no es familiar, constituye un accidente biológico y son mutaciones frescas. Las aberraciones cromosómicas, en especial las que involucran al cromosoma Y, dan un modelo que nos muestra cómo la variabilidad genética y el ambiente pueden interactuar en la producción de un fenotipo sicológico, en el cual intervienen variables que pueden ser consideradas anormalidades en la fisiología del cerebro o en el sistema endocrino.8 Esta breve descripción genética del síndrome XYY no nos hace partidarios de atribuirle a la agresividad, el calificativo de congénita en nuestra especie, pero dado que en ocasiones se han deducido conclusiones con poca base científica acerca de las implicaciones sociales que el cariotipo XYY presenta, se ha originado una controversia muchas veces extra-científica sobre esta problemática. En realidad, como señala Hook,7 la cuestión debe plantearse en los términos siguientes: ¿Es más frecuente la ocurrencia de personas XYY en las instituciones penales y/o de enfermedades mentales que en la población general? En caso afirmativo podríamos preguntarnos: ¿Que relación causa-efecto existe entre la constitución XYY y el comportamiento por el que dichas personas han tenido que ser internadas en tales instituciones? En nuestro caso, cualquier duda relacionada con las interrogantes que anteceden, debe ser entendida bajo la tesis de que por su "naturaleza" cromosómica, el hombre no es, a pesar de todo, un animal sanguinario, agresivo, que busca la violencia por todos los medios, inunda al hombre contemporáneo en la vida, los medios de comunicación masiva, el cine y la literatura. Unos países la padecen y sufren menos, otros más, pero ninguno puede sentirse libre de la violencia. La problematización sobre esta alteración, que tiene un defecto genético marcado, pero en la que el medio social impone prácticamente la última palabra, tiene ilimitados campos, sobre todo desde el punto de vista ético. Los que hemos estudiado algo sobre este síndrome, revalidamos que se ha asociado a las formas de conducta violentas, sobre todo a la criminalidad, pero sólo eso; son referencias de estudios en otros países, los que han seguido cierta estrategia metodológica en su investigación y de esa forma han establecido diferentes parámetros, como es el conocer su frecuencia de aparición, diagnóstico, pronóstico y tratamiento; este último es motivo de controversia, e incluye estimulación precoz; sin olvidar la vertiente ética que un diagnóstico prenatal de este síndrome puede proponer; nos preguntamos por ejemplo: ¿Qué riesgo hay de que un niño recién nacido, XYY, manifieste el comportamiento agresivo antisocial en comparación con otro niño cualquiera, XY, nacido y criado en condiciones sociales semejantes? ¿En qué medida cada caso de conducta agresiva individual, está relacionada con las condiciones externas de vida, con la educación, la forma ideopolítica, la actividad, etc. del hombre en cuestión, y en qué medida obedece a su mundo interno, a su naturaleza? ¿Por qué personas que están expuestas a la influencia de los mismos factores sociales y criminógenos, NO comenten delitos? ¿MITO O REALIDAD? Desde la antigüedad hasta nuestros días, la cuestión de los móviles internos de la conducta humana ha interesado a científicos, filósofos, abogados, biólogos, genetistas, psiquiatras y fisiólogos, lo que ha provocado la aparición de numerosas y disímiles hipótesis. La insuficiencia de investigaciones con verdadero rigor científico y falta de claridad teórica en los fundamentos en torno al problema en cuestión han ocasionado las dudas existentes, y es esto precisamente lo que ocurre respecto a esta línea problemática que se investiga y que da título a este capítulo. En la bibliografía consultada sólo se expone con franca claridad el aspecto clínico del fenómeno, haciéndose referencia a revisiones puramente médicas, permitiendo enmarcar el síndrome hasta el más sencillo detalle anatómico, descrito siempre con la asociación al comportamiento agresivo, antisocial y criminal. De un total de 115 resúmenes de investigaciones consultadas en la base de datos MEDLINE, comprendidos desde 1990 hasta mayo de 1998, sólo 10 de ellos nos aportan nuevos elementos esenciales de lo genético o de lo social del fenómeno en cuestión; que no relata la literatura especializada; entre los cuales se destacan los siguientes: • Suiza (1993). Aspectos biológicos de la delincuencia y la agresión. Prueba que existe un factor hereditario fuerte en la génesis de la criminalidad en la delincuencia.3 • Irlanda (1992). Genotipo XYY y crimen. Describe a 2 individuos XYY que cometieron varios crímenes. Señala además la importancia del conocimiento sobre este genotipo para no errar en la forma en que deben ser analizados estos individuos en la corte.9 • Rusia (1991). Describen a 5 pacientes 47, XYY que manifestaban signos de desviación del comportamiento sexual 113 normal, específicamente, con predominio en la estructura de la libido de "objetivos sádicos", lo que contribuyó a mostrar su comportamiento sexual agresivo.10 • Filipinas (1992). Un niño XYY, experiencias de su madre. Basadas en notas del diario de la madre del propósito y comparables con la de otros padres con hijos con retraso mental. Plantean que la realización de un estudio genético selectivo para el ingreso a escuelas de niños con retraso mental, el presentar un fenotipo poco conspicuo, el diagnóstico tardío de este síndrome y la falta de ayuda profesional ocasionaron complicaciones, al adquirir modelos de conducta mucho más agresivos a los ya expresos por él, dada su condición de XYY.5 • Suecia (1995). XYY estado mental y funcionamiento sicosocial. En un estudio de 1968-1993 se reportan 75 varones con un cromosoma Y extra. Del total de individuos estudiados el 80 % estuvo caracterizado por un comportamiento agresivo y problemas siquiátricos, contra el 24 % que era mentalmente normal.11 • Inglaterra (1997). 47, XYY y 47, XYYY. Indicación para el diagnóstico posnatal con implicaciones para el diagnóstico prenatal y el consejo genético: Afirma el riesgo del comportamiento agresivo que puede aparecer en los niños XYY y su relación con las leyes judiciales.12 A propósito alegamos que sólo incluimos referencias de la literatura extranjera, pues en la cubana no ha sido tratado el tema, no obstante, en el proceso formativo de los profesionales de la salud, éstos reciben información teórica sobre él. Generalizando, la literatura consultada, no muestra testimonios de los individuos XYY como particular, ya que adole- 114 cen de datos personales, procedencia social, micromedio, nacionalidad verdadera o aspectos sociodemográficos, aunque son muy importantes para conocer y demostrar la influencia del medio social sobre este desorden genético; aun cuando es sabido que este factor es determinante en las formas de aparición de conductas agresivas y violentas. En nuestra posición, la simple presencia de un cromosoma Y extra, no es total ni cierta explicación al sometimiento de este síndrome ante la estigmatización de criminalidad, pues todo genotipo se expresa mediante su interacción con el ambiente, por lo que a nuestra consideración lo social determina en la aparición del fenotipo sicológico que se presenta en algunos casos de individuos XYY. Muchos países han creado, y no con muy buenas intenciones, centros especializados para el estudio de la violencia, y este hecho, analizando en el contexto de la tendencia a la globalización de la economía, encamina su utilización en contra de los pueblos latinoamericanos, planteando que somos inferiores, no capaces de desarrollarnos por nosotros mismos, al carecer de inteligencia para ello, enervándonos moralmente, siendo tratados como personalidades marginales, con etiqueta de desviados, bajo cualquier marcador genético. Autores como el profesor Pinatel, relacionan su concepción de la personalidad criminal con la sociedad global, y los estímulos que proceden de esta sociedad, con los factores del medio individual, los cuales, combinándose con los factores biológicos juegan un papel determinante en la formación no sólo de la personalidad criminal, sino también de las situaciones en las cuales ella se encontrará confrontada.13 Por su parte Vogel,8 apoyándose en el modelo que muestra cómo la variabilidad genética y el ambiente pueden interactuar en la presentación de un fenotipo psicológico, nos indica de modo sugerente una estrategia de investigación, consistente en identificar la variante genotípica al explorar su influencia en el fenotipo, y considerar concomitantes las diferencias intra e interindividuales en un ambiente determinado, lo que es opuesto a los objetivos usuales, que comienzan por el fenotipo; por lo que los disturbios provocados por aberraciones cromosómicas en el desarrollo embrionario y fisiológico, son pobremente entendidos. Lo hasta aquí expuesto, resume lo hallado en la literatura consultada; si partimos tanto del ángulo social, como de la tendencia estigmatizadora de lo biológico (genético) y valoramos su interacción, no recibimos una respuesta concluyente a la interrogante de este apartado ¿mito o realidad?, pero sí nos permite sugerir: 1. En la publicación de síndromes con fenotipos psicológicos, como en el caso del XYY, el ofrecer datos (verdadera procedencia social, nacionalidad del paciente, micromedio, aspectos sociodemográficos, etc.), permite hacer una valoración más justa y científica sobre la influencia del ambiente en su aparición. 2. Establecer la frecuencia de aparición de esta anomalía cromosómica en nuestro medio (aunque no constituya un problema de salud), sólo por las implicaciones bioéticas que de él se deriven, producto de la estigmatización a la que está sometido. 3. Instrumentar como requisito indispensable para la incorporación a escuelas especiales o instituciones para niños con retraso mental, la realización de un estudio genético selectivo, para evitar un diagnóstico tardío de este síndrome y poder ofrecer un tratamiento especial a estos niños, lo que redundará en bienestar individual, a la familia, y a la sociedad. SUMMARY Genetic stigmatization is an event that takes on significance with the advances in molecular biology and the discovery of every new gene contributed by the Human Genome research project; however, there are still syndromes like the XYY one which has been associated to violent behaviours since it was described for the first time. In this paper, we made an assessment of the basic ideas on this syndrome and its relationship with delinquency based on the reports found in MEDLINE database from 1990 to may, 1998. The unsatisfactory collection of the described subjects’ personnal data, so important for the analysis of the role of the social environment on the occurence of this fenotype, was stressed. We also made our own assessment of the location of this problem line between the biological field and the social field. As well as some suggestions and/or considerations, taking unlimited scope of XYY syndrome mainly from the ethical viewpoint into account. Subject headings: XYY KARYO TYPE; VIOLENCE; STEREOTIPING. REFERENCIAS BIBLIOGRÁFICAS 1. Olmo R del. El surgimiento de la criminología. Divulgación Jurídica, MINJUS 1985;3(12):25-6. 2. Roldan EJ. Síndromes pediátricos dismorfogénicos. Madrid: 1982:430-1. 3. Knech T. Biological-psychiatric aspect of delinquency and agression. Kantonale Psychaitrische Klinik. Munsterlingen. Schweiz Med Wochenschr 1993;123(22):1165-75. 4. Gelisio P. WYY syndrome. Report of a case. Venezia Mastre. Minerva Endocrinol 1991;161(4):199-201. 5. Gontard V. A child with WYY syndrome as experienced by his mother. Klinik für kinder - Und Jugend psychiatrie. 1994;123(24):140-3. 115 6. Marinello MJ. A study of the WYY syndrome in tall men and juvenile delinquents. J Am Med 1969; 208:321-2. 7. Hook EB. Behavioral implications of the human XYY genotype Science 1973;179:139-40. 8. Vogel F, Motulsk A. Human genetics. Springer Verlag, Frankfurt 1982:478-505. 9. Freyre A, O’Connor A. XYY genotype and crime: cases. Dublin Ireland. Med Sci Law 1992;32(3):261-3. 10. Shostakowich BV, Smirnova LV. The clinico-biochemical characteristics of persons with paraphilias and chromosome anomay (47, XYY). Neuropathol Psikiatr 1991;91(3):24-8. 11. Fryns JP. XYY. Psics and social funtioning of mental state Genet Couns 1995;6(3):197-206. 12. Abransky L. 47 XYY, Pre and postboon diagnostic. Genetical advaisoary implications Pre enant. Diagnostic Preenat Diagn 1997;17(4):363-88. 13. Grandes tendencias de la criminología contemporánea. Divulgación Jurídica: MINJUS 1986;14(13):19-22. Recibido: 9 de febrero de 1999. Aprobado: 19 de julio de 1999. Lic. Héctor I. Pimentel Benítez. Instituto de Ciencias Básicas y Preclínicas " Victoria de Girón" , Ciudad de La Habana, Cuba. 116 Rev Cubana Invest Biomed 1999;18(2):117-20 Centro Nacional Coordinador de Ensayos Clínicos ALGUNAS CONSIDERACIONES GENERALES SOBRE EL DESARROLLO DE LA EVALUACIÓN DE PRODUCTOS FARMACÉUTICOS Y BIOTECNOLÓGICOS Y EQUIPOS MÉDICOS EN CUBA Y EL MUNDO Dra. Martha María Fors López y Dr. Daniel Peña Amador RESUMEN Se pone en conocimiento de los profesionales de la salud el proceso mediante el cual llegan al mercado los medicamentos y equipos médicos que utilizan diariamente en la práctica médica. Se explican brevemente las etapas por la que atraviesan hasta alcanzar el registro sanitario. Se exponen las tendencias actuales referentes a este proceso. Descriptores DeCS: EQUIPOS Y SUMINISTROS; EVALUACION DE MEDICAMENTOS; EVALUACION DE TECNOLOGIA BIOMEDICA; ENSAYOS CLINICOS; VIGILANCIA EPIDEMIOLOGICA. Para muchos de nuestros profesionales de la salud es desconocido el proceso mediante el cual los medicamentos o equipos médicos que utilizan, llegan al mercado. Este proceso es complejo y largo, necesitándose la cooperación de numerosos investigadores médicos para su realización. El desarrollo de los agentes terapéuticos comenzó a principios del siglo XIX y en este siglo la mayoría eran de origen natural. Con el avance de las ciencias químicas y de la fisiología se impulsó la introducción de nuevas drogas y de tecnologías sanitarias y hasta la actualidad el número de éstas es cada vez mayor.1 Hoy en día la Biotecnología moderna comprende técnicas tan distintas como la de cultivos celulares y tisulares, la micropropagación y la fermentación y cons- tituye un pilar fundamental en la producción de los productos biotecnológicos para el diagnóstico y tratamiento de numerosas enfermedades. Esta rama de la ciencia se basa en diversas disciplinas científicas, en particular la biología molecular y celular, la bioquímica, la genética, la inmunología y la informática entre otras. Cada producto utilizado por la medicina atraviesa por diferentes etapas: • Estudios preclínicos • Estudios clínicos • Estudios poscomercialización ESTUDIOS PRECLÍNICOS Esta etapa comprende desde que se descubre una nueva molécula hasta que está lista para poder ser evaluada en humanos. 117 Se realizan estudios químico-farmacéuticos y toxicológicos, determinándose algunos elementos en modelos de animales experimentales que sugieran su utilización con cierta seguridad en seres humanos. ESTUDIOS CLÍNICOS Esta etapa comprende la realización de los llamados ensayos clínicos, los cuales atraviesan por 4 fases en las cuales se determinan la seguridad del producto, su efecto terapéutico y su eficacia. El producto es sometido a un monitoreo estrecho y sólo es administrado a centenas de pacientes. Una vez concluidos estos estudios, se somete el expediente del producto con la información generada a la consideración de la Agencia regulatoria del país del que procede, el cual lo revisa y determina si el producto debe ser puesto en el mercado o no. ESTUDIOS DE POSCOMERCIALIZACIÓN Esta etapa comprende la realización de estudios de Farmacovigilancia, donde se identifican los efectos del uso de tratamientos farmacológicos en el conjunto de la población.2 Cuba, a pesar de ser un país del llamado tercer mundo, cuenta con una naciente industria médico-farmacéutica y biotecnológica, la cual produce ya agentes novedosos que requieren ser registrados en el país y fuera de él, con el fin de ser comercializados y aplicados en la práctica médica a la mayor velocidad y con la mayor seguridad y protección de la población. La Biotecnología en Cuba, comienza su andar de manera acertada y más de una institución goza de reconocido prestigio 118 internacional, por lo que se hacía necesario la creación de otros centros que la apoyaran en el desarrollo de sus productos. En 1989 se crea, mediante una Resolución Ministerial, el Centro para el Control Estatal de la Calidad de los Medicamentos (CECMED), el cual es la autoridad competente para la autorización del registro de medicamentos de uso humano, nacionales y extranjeros para toda Cuba. En 1991, también por Resolución Ministerial, se crea el Centro Nacional Coordinador de Ensayos Clínicos, cuyos objetivos son garantizar la evaluación clínica que se requiere para el registro y la comercialización de productos farmacéuticos y biotecnológicos así como de equipos médicos realizados en Cuba y en otros países. Este centro se considera como un CRO (Contract Research Organization) el cual es resultado del desarrollo acelerado en nuestro país de la industria biotecnológica y médico-farmacéutica. La creación de este centro favorece el trabajo que deben realizar las compañías productoras de medicamentos en la evaluación de sus productos, que en ocasiones excede sus posibilidades para enfrentarse a tales procesos. Es importante saber que muchos profesionales de nuestro Sistema de Salud participan junto a este centro en la evaluación de los productos fabricados en nuestro país y que para ello nos guiamos por las llamadas Buenas Prácticas Clínicas (BPC), publicadas por el CECMED. Este documento se corresponde con la guía de la Conferencia Internacional de Armonización (ICH) de BPC, la cual es una norma internacional de calidad científica y ética dirigida al diseño, conducción y reporte de ensayos clínicos.3 Países como EE.UU., Japón y la Comunidad europea se han reunido en la ICH con el objetivo de hablar un lenguaje co- mún referente al desarrollo de los medicamentos y equipos médicos. Durante 1997, se revisaron y reactivaron las guías que habían sido propuestas por la ICH. Desde 1991 el comité directriz ha estado de acuerdo con 34 guías y otras 11 se encuentran actualmente en revisión. Se están revisando 2 guías adicionales: una de terminología médica y otra de estándares electrónicos. El nuevo reto de la ICH es la armonización de un documento técnico común (CTD) con el que se espera perfilar la preparación y la revisión de las solicitudes internacionales. En estos momentos no es posible una solicitud completamente armonizada, sin embargo, se han definido los elementos esenciales por donde se desarrollará esta guía. Este documento incorporará resúmenes escritos, datos tabulados y documentación de soporte, no se incluirán documentación administrativa, informes de expertos y datos originales.4 En los países desarrollados, se está produciendo un fenómeno alrededor del proceso para el registro de los productos de la Industria Médico-Farmacéutica y Biotecnología (IMFB), condicionado por diversos factores y que ha conducido a la revisión y análisis de los requerimientos para dicho registro, con el fin de hacerlo más eficiente sin alterar el equilibrio que este proceso requiere. Nuestro país también está enfrascado en adoptar las medidas que sean posibles para alcanzar el mismo nivel del primer mundo. La tendencia actual es compartir información sobre nuevas tecnologías y productos, en reuniones realizadas entre las industrias farmacéuticas, los CRO y las agencias regulatorias para beneficiarse de la sabiduría colectiva de oficiales claves gubernamentales, de personas consultoras de renombre y leaders de la industria farmaceútica. Es cada vez más importante que cada profesional busque, descubra e integre la riqueza del conocimiento disponible. Dentro de los 5 años siguientes existirá un aumento de las nuevas tecnologías lo que requerirá un pensamiento innovador de cómo regular este amplio espectro de productos. La industria farmacéutica necesitará trabajar estrechamente con las agencias regulatorias y pensar sobre cómo regular nuevas tecnologías.5 Dentro del campo de los productos biológicos y de otros tipos de medicamentos se ha observado en los últimos 2 años una colaboración mayor entre ellos. En general, la forma de regular los productos tradicionales no debe ser la misma de regular los no tradicionales. El espíritu de colaboración existente entre las agencias regulatorias y los patrocinadores de ensayos clínicos es imperecedera. Se ha conformado un equipo bien integrado, los procesos administrativos de las agencias se han vuelto más transparentes facilitando al productor ver exactamente qué es lo que tiene que hacer para lograr un registro de su producto. Esto permite establecer el contacto para determinar el tipo de evidencia científica necesaria para demostrar la eficacia de una droga o equipo médico antes de comenzar el ensayo clínico. Existe un proceso de revisión colaborativo en el cual la agencia se reúne con el productor para discutir las deficiencias y decidir qué información adicional se necesitaría para corregirlas. Las mejoras regulatorias han perfeccionado ostensiblemente la calidad de las solicitudes. 119 SUMMARY Health professionals are informed of the processes by which daily used drugs and medical equipment reach the market. The different stages of this process until the sanitary register is obtained are briefly explained. The present trends of this process are set forth. Subject headings: EQUIPMENT AND SUPPLIES; DRUG EVALUATION; TECHNOLOGY ASSESSMENT BIOMEDICAL, CLINICAL TRIALS: EPIDEMIOLOGY SURVEILLANCE; POSTMARKETING. REFERENCIAS BIBLIOGRÁFICAS 1. Laporte J. Principios básicos de investigación clínica. Madrid: Ediciones Ergon, 1993. 2. Laporte J, Tognoni G. Principios de epidemiología del medicamento. Barcelona: Masson-Salvat Medicina, 1993. 3. Guía ICH tripartita y armonizada para la buena práctica clínica. El Medicamento. Barcelona: Biomedical Systems, 1997. 4. Leary F. ICH update. Regul Affairs Focus 1997;2(12):16-9. 5. Bass J. Regulatory cooperation over the next five years. Regul Affairs Focus 1997;2(12):8-9. Recibido: 21 de octubre de 1998. Aprobado: 19 de julio de 1999. Dra. Martha María Fors López. Centro Nacional Coordinador de Ensayos Clínicos, Ciudad de La Habana, Cuba. 120 Rev Cubana Invest Biomed 1999;18(2):121-9 TRABAJOS DE ACTUALIZACIÓN Instituto de Ciencias Básicas y Preclínicas "Victoria de Girón" TELÓMEROS Y TELOMERASAS Dr. Rolando A. Hernández Fernández RESUMEN Los telómeros son estructuras cromatínicas especializadas que se encuentran localizadas en los extremos de los cromosomas eucariontes. Tanto el ADN como las proteínas que los constituyen presentan características singulares que los diferencian del resto de los cromosomas. Parecen estar implicados en numerosas funciones celulares, especialmente las relacionadas con el control de la duración de la vida de diferentes estirpes celulares. Estas estructuras se replican durante el ciclo celular gracias a la acción de enzimas denominadas telomerasas que están formadas por proteínas y ARN y presentan un mecanismo peculiar. Recientemente se ha estudiado el comportamiento de las telomerasas en las células cancerosas y sus posibles aplicaciones diagnósticas y terapéuticas. En este trabajo se presenta un resumen de los principales hallazgos más recientes sobre la estructura y funciones de los telómeros y la acción de las telomerasas. Descriptores DeCS: TELOMERASA/fisiología; TELOMERO/fisiología; CICLO CELULAR. Los telómeros constituyen estructuras especializadas que forman los extremos de los cromosomas eucariontes que participan en funciones celulares tan importantes como la mitosis, la estabilidad cromosómica y el tiempo de vida de las estirpes celulares. Recientemente se ha demostrado su relación con algunas enfermedades, especialmente con el cáncer. Durante las últimas 2 décadas mucho se ha avanzado en el conocimiento de su estructura y dinámica. En este trabajo se resumen los hallazgos recientes más importantes. ESTRUCTURA DE LOS TELÓMEROS Los telómeros fueron identificados por H.J. Muller durante la década de los años 30. Desde entonces, se ha profundizado extraordinariamente en el conocimiento de estas estructuras, gracias a la introducción de la moderna tecnología de la Genética Molecular. Un resumen de los principales trabajos realizados en los primeros años de aplicación de estas técnicas fue publicado en 1984 por Blackburn y Szostack.1 Los estudios se han realizado principalmente en ciliados, cuyos macronúcleos 121 pueden contener de 40 000 a 1 000 000 de telómeros según la especie, por lo que constituyen una excelente fuente de componentes teloméricos y de las enzimas que participan en su replicación. Posteriormente, se ha comprobado que los aspectos descritos en ciliados están presentes en otros organismos. En los ciliados y en S. cerevisiae los telómeros se encuentran en una forma de estructura cromatínica particular no nucleosómica que ha sido denominada telosoma. Los nucleosomas adyacentes presentan histonas hipoacetiladas características de la cromatina que no se transcribe activamente. En mamíferos, donde son mucho más largos, se presentan formados por nucleosomas pero hacia la zona más extrema aparecen como telosomas. Esto evidencia que al menos en parte hay conservación estructural de los telómeros. También muestran diferencias interespecies algo sorprendentes. cantidad de ADNt por cromosoma también fluctúa. En algunos organismos la longitud promedio de los telómeros responde a cambios genéticos o nutricionales. Las secuencias de ADNt mejor caracterizadas se muestran en la tabla 1 confeccionada a partir de la excelente revisión de V.A. Zakian.2 La mayoría de las secuencias son cortas y precisas, como en Tetrahymena que es de 6 pb (TTGGGG). Sin embargo, en S. cerevisiae es heterogénea, y muy larga en K. lactis donde presenta 25 pb. Especies muy distantes pueden presentar la misma secuencia, como los vertebrados, y especies muy cercanas como Tetrahymena y Oxytricha ser diferentes aunque muy relacionadas. A TTAGGGTTAGGGTTAGGGTTAGGGTTA AATCCCAATCCC T A EL ADN TELOMÉRICO En casi todos los eucariontes estudiados, el ADN telomérico (ADNt) consiste en repeticiones en tanden de pequeñas secuencias nucleotídicas con una distribución asimétrica de los pares G:C, pues las G se acumulan en una de las hebras (hebra G) y se encuentran agrupadas. La hebra G está orientada de 5' a 3' hacia el extremo del telómero y forma el extremo 3’del ADN cromosómico. En la zona más extrema no está apareada formando un segmento final monofibrilar con una longitud que varía según la especie. Las principales características estructurales de los telómeros se resumen en la figura 1. La longitud del telómero es variable. En Oxytricha y Euplotes es apenas de 38 pb mientras en ratones alcanza 150 kb. Cada organismo posee una longitud media característica. La 122 B TTAGGGTTAGGG-T-T-A AATCCCAATCCC A G-G G-G G-G TTA T A C TTAGGGTTAGGG-T-T-A AATCCCAATCCC A G-G G-G G-G TTA Fig. 1. Representación esquemática de la estructura de los telómeros. (A) El ADN telomérico consta de pequeñas secuencias repetidas ricas en bases de guanina con un extremo monofibrilar. (B) El extremo monofibrilar puede formar estructuras de tipo secundario por apareamiento de las bases de guanina. (C) Estas repeticiones sirven para la unión a proteínas tanto en la doble hebra como en la zona monofibrilar. TABLA 1. Secuencias de ADN telomérico mejor caracterizadas Organismo Secuencia Tetrahymena Oxytricha Giardia Physarum Kluyveromyces Neurospora Caenorphabditis Vertebrados TTGGGG TTTTGGGG TAGGG TTAGGG TCGGATTTGATTAGGTATGTGGTGT TTAGGG TTAGGG TTAGGG Mientras más secuencias teloméricas se conocen, más difícil resulta encontrar una secuencia consenso. La existencia de múltiples secuencias teloméricas sugiere que las funciones de los telómeros no requieren de una secuencia única. El hallazgo de secuencias teloméricas en sitios internos de los cromosomas demuestra que ellas por sí mismas no hacen los telómeros. In vitro, las hebras G pueden aparearse de formas diversas. Una de ellas es la de hélices tetrafibrilares o cuartetos G, que se mantienen por múltiples pares G:G. Si estas estructuras son importantes para la función de los telómeros se explicaría el agrupamiento de las G que se observa en las secuencias. Dos excepciones notables aparecen entre los telómeros. En Parascaris no se observa la distribución asimétrica de las G, éstas ni siquiera están agrupadas pues su secuencia es TTGCA. En Drosophila presentan una estructura totalmente diferente representada por repeticiones de elementos de transposición. LAS PROTEÍNAS TELOMÉRICAS Las proteínas teloméricas (PT) pueden presentarse asociadas al extremo monofibrilar o a la zona adyacente de doble hebra. Entre las primeras se ha iden- tificado una proteína de Oxytricha que no está relacionada estructuralmente con ninguna otra proteína. In vitro, esta proteína facilita la formación de cuartetos G, lo cual sugiere la existencia de estas estructuras in vivo. No se le ha detectado unión a zonas internas del cromosoma. Una proteína de unión a la hebra G monofibrilar en levaduras es el producto del gen EST1. La unión de EST1p es específica para las repeticiones (TG1!3)n de una sola hebra y requiere de un extremo 3’ libre.3 La principal PT de unión a la zona de doble hebra en S. cerevisiae es el producto del gen RAP1. Su unión a las repeticiones (TG 1!3)n de hebra simple se realiza con menos afinidad que a las de doble hebra. Se ha demostrado que RAP1p contiene 2 motivos estructurales de unión al ADN que son homólogos a los del oncogen myb. A partir de células HeLa se purificó una proteína de 60 kDa que se une a los telómeros humanos y se le denominó hTRF. La proteína tiene 439 aminoácidos para una masa molecular de 50 341 dalton y su expresión in vitro da lugar a una proteína de tamaño similar a la identificada en los extractos nucleares de células HeLa.4 Alrededor de la posición 350 hTRF contiene 2 secuencias de localización nuclear, la zona N terminal es rica en aspártico y glutámico y en la zona C terminal presenta fuerte homología con la región de unión al ADN del oncogen myb. Ver la referencia5 para una comparación con otros TRF de mamíferos. A partir del hTRF se pudo localizar una proteína telomérica con motivos del oncogen myb en Schizosaccharomyces pombe.6 Es de esperar que en los próximos años se puedan identificar otras proteínas implicadas en la estructura y función de los telómeros. 123 LAS TELOMERASAS El descubrimiento de las telomerasas resolvió en principio el problema de la replicación de los estremos de moléculas lineales de ADN. La actividad de telomerasa fue detectada por primera vez en Tetrahymena y después en otros eucariontes. En la referencia7 aparece una revisión de los principales trabajos sobre esta enzima. La enzima es una ribonucleoproteína y para su actividad son esenciales tanto el componente proteínico como el ARN. En Tetrahymena el ARN telomerásico (ARNtl) tiene una longitud de 159 nucleótidos (nt) y contiene 9 repeticiones del molde 5' CAACCCCAA3'. Esta característica que se observa también en todos los otros ARNtl identificados, permite el apareamiento del telómero en crecimiento con el ARNtl y que aún quede una región que pueda servir de molde. Las telomerasas difieren de todas las polimerasas en que utilizan un molde interno en vez de uno externo, lo cual impone limitaciones estéricas específicas para la elongación del iniciador y la catálisis. La telomerasa alarga el ADN iniciador por la adición uno a uno de los desoxinucleósidos trifosfatados y así genera las repeticiones en tanden de los telómeros. La de Tetrehymena puede alargar ADN iniciadores Oxytricha, humanos, plantas y levaduras. Esto sugiere que la enzima tiene afinidad general por las secuencias ricas en G más que por un motivo específico como sucede con otras proteínas de unión al ADN. Pero en todos los casos, la secuencia añadida se corresponde con el molde de ARN de la enzima. Actualmente, se propone la existencia de 2 sitios enzimáticos independientes de interacción con el ADN iniciador. Uno contiene el molde de ARN y alínea el ex- 124 tremo 3' del iniciador para su elongación en el centro catalítico. El otro se une al ADN iniciador hacia el lado 5’del molde y proporciona una vía de salida para la hebra en crecimiento. Este modelo explica la adición de varias repeticiones sin que la enzima se disocie del ADN iniciador. Este sitio catalítico único debe moverse en relación con el ARN molde. Experimentos en Euplotes apoyan este modelo de los 2 sitios.8 La enzima posee también actividad endonucleolítica que pudiera estar relacionada con una función de corrección (fig. 2). 1 TTAGGGTTAGGGTTA AATCCC CAAUCCCAAUC 5 3 2 TTAGGGTTAGGGTTA AATCCC CAAUCCCAAUC 3 5 TTAGGGTTAGGGTTAGGGTTAG 3 AATCCC CAAUCCCAAUC 3 4 5 TTAGGGTTAGGGTTAGGGTTAG AATCCC CAAUCCCAAUC 3 5 Fig. 2. Mecanismo de acción de la telomerasa. (1) La enzima contiene el ARN telomerásico con un sector complementario a la secuencia de los telómeros. (2) La unión del ADN telomérico con el ARN telomerásico se produce por apareamiento de bases complementarias. (3) La enzima alarga el telómero usando como molde el ARN. (4) Al completar un alargamiento la enzima se desplaza y comienza un nuevo ciclo de alargamiento y así sucesivamente. EL ARN TELOMERÁSICO PROTEÍNAS TELOMERÁSICAS El ARNtl ha sido aislado de numerosas especies. La longitud de éste varía desde 160 a 190 nt en los ciliados hasta 1 300 en levaduras y cada uno de ellos contiene una secuencia molde apropiada para la síntesis de las repeticiones teloméricas. Los ARNtl presentan poca homología de secuencia, sin embargo, al menos en ciliados, la estructura secundaria está muy conservada. En Tetrahymena solamente 159 nt son suficientes para un funcionamiento adecuado. Se ignora por qué en otras especies es mucho más largo. Algunas características de estos ARN se recogen en la tabla 2. Los estudios con mutantes de ARNtl en Tetrahymena han mostrado que cambios en algunos de los 6 nt del extremo 5' (CAACCCCAA) producen secuencias repetidas alteradas, sin embargo los 3 últimos no son utilizados como molde y más bien parecen participar en alinear el iniciador. En el mutante (AAACCCCAA) se produce una disociación prematura de la enzima antes de arribar al nucleótido cambiado, tal vez por efectos estéricos de los nucleótidos alterados en el molde o porque el ARN desempeñe algún papel en la catálisis u otra función.9 La telomerasa de Tetrahymena consta de 2 polipéptidos, p80 y p95, el menor se une al ARNtl y el mayor al ADN iniciador. La enzima contiene una copia de cada una de las proteínas y una del ARNtl para una masa molecular aparente de 250 kDa. Las secuencias de p80 y p95 no guardan una extensa homología con otras proteínas, lo que indica que se trata de proteínas nuevas. No obstante, se ha encontrado una homología limitada entre p95 y las ARN polimerasas dependientes de ARN (transcriptasas inversas, T1) así como con motivos estructurales de algunas ADN polimerasas. Este motivo de T1 también se encuentra en la p123 de E. aediculatus y en el producto del gen EST2 de S. cerevisiae cuya deleción produce defectos teloméricos. Los cambios de un aminoácido por otro en el motivo de T1 de EST2p produce acortamiento de los telómeros y senescencia en levaduras, lo que indica su importancia para el alargamiento de los telómeros in vivo.10 También se han identificado en mamíferos 2 proteínas con homología secuencial con p80. Una de ellas designada TP1 interactúa con el ADNt. 11 La otra obtenida de ratas se ha denominado TLP1 y su ADNc TABLA 2. Características de algunos ARN telomerásicos Organismo Tetrahymena Euplotes Oxytricha Humanos Ratón S. cerevisiae K. lactis Molde CAACCCCAA CAAAACCCCAAAACC CAAAACCCCAAAACC CUAACCCUAAC CCUAACCCU CACCACACCCACACAC UCAAAUCCGUACACCAAUACCUAAUCAAA Longitud (nts) 159 190 190 450 450 ~1300 ~1300 125 codifica una proteína de 2 629 aminoácidos para una masa molecular deducida de 240 kDa (p240).12 Esta p240 es modificada in vivo y convertida en p230, tal vez como un mecanismo postraduccional de regulación de la actividad de la telomerasa. GENÉTICA DE LA TELOMERASA Al introducir cambios en la secuencia molde del ARNtl en Tetrahymena las células se hacen muy grandes con problemas en la división nuclear y pierden viabilidad después de varias generaciones. Existe una alteración en los mecanismos que regulan la longitud de los telómeros, tal vez porque se inhibe la unión de las PT al cambiar la secuencia. En S. cerevisiae la deleción de EST1 produce un acortamiento progresivo de los telómeros y al cabo de unas 50 generaciones las células se hacen muy grandes y muchas de ellas mueren. La deleción del gen TLC1, que codifica el ARNtl y de EST2, EST3 y EST4 produce fenotipos similares que no se agravan en los dobles mutantes, lo que sugiere que estos genes están en epistasis.13 REPLICACIÓN DE LOS TELÓMEROS En la estructura de los telómeros predomina la zona de repeticiones en la doble hebra y solamente el extremo terminal presenta la estructura monofibrilar. Aunque la telomerasa es necesaria para mantener la longitud de los telómeros, esta enzima sólo alarga la hebra G. La replicación de la hebra C debe hacerse por el sistema convencional de las polimerasas. En levaduras mutantes para la polimerasa a y la proteína replicativa C (RFC) tienen alteraciones en 126 la síntesis de los telómeros. Tanto en levaduras, en humanos, como en otros organismos los telómeros se replican al final de la fase S.14 La replicación de los telómeros es sólo necesaria para compensar la pequeña y lenta pérdida de ADN que resulta de la replicación incompleta, por lo tanto debe ser considerada como una función de reparación. En este proceso, el ADN telomérico no tiene exactamente la función de molde. In vitro la telomerasa de Tetrahymena puede añadir hasta 500 nt antes de separarse del ADN. La síntesis de la hebra C por el sistema convencional de las polimerasas generaría una larga molécula bifibrilar con un extremo monofibrilar de 8 a 12 nucleótidos, que es un buen sustrato para la telomerasa. El estado dinámico de los telómeros depende de varios factores, entre ellos, la procesividad de las telomerasas, la frecuencia de su acción sobre cada telómero particular y la velocidad de degradación del ADNt.15 Las PT como el hTRF1 también pueden regular su longitud.16 Se ha visto que en levaduras, un mutante (TEL1) deficiente en la regulación de los telómeros es homólogo de 2 genes implicados en la regulación del ciclo celular: el MEC1 en levaduras y el de la ataxia telangiectásica (ATM) en humanos, lo cual apunta a un vínculo entre la replicación de los telómeros y el ciclo celular. FUNCIONES DE LOS TELÓMEROS Los telómeros representan estructuras esenciales para las células, pues evitan la fusión cromosómica, desempeñan un importante papel en mantener la estabilidad cromosómica 17 y participan tanto en la meiosis como en la mitosis.18 Sin embargo, su función más notoria es la de servir como un reloj mitótico que mide y regula el número de las divisiones celulares.19 Los telómeros se acortan con cada división celular y el número de divisiones que la célula puede experimentar se correlaciona con la longitud de los telómeros.20 Este acortamiento pudiera eliminar genes indispensables para la vida o silenciar genes cercanos por el efecto de posición del telómero. Una longitud crítica pudiera ser la señal para la entrada en la senescencia celular. Sin embargo, vale tener presente que no está relacionada con la edad del organismo. La actividad de la telomerasa varía en diferentes etapas de la vida. Se ha detectado en ovarios y testículos en fetos, recién nacidos y adultos, pero no en óvulos ni espermatozoides maduros. El blastocisto y la mayoría de los tejidos somáticos de 16 a 20 semanas de desarrollo exhiben un alto nivel de telomerasa que desaparece después del nacimiento.21 También es alta en tejidos adultos con una intensa proliferación celular como en las células endoteliales22 y el endometrio. 23 En otras células puede ser inducida en determinadas etapas de la vida como ocurre con la activación de los linfocitos T24 y los B.25 Esta función de los telómeros los relaciona de inmediato con la transformación cancerosa. En uno de los primeros trabajos sobre el tema se encontró que en células cultivadas de 18 diferentes tejidos humanos 98 de cada 100 inmortales y ninguna de 22 mortales mostraban actividad de telomerasa. Asimismo 90 de 101 biopsias de 12 tipos de tumores y ninguna de 50 de tejidos somáticos normales poseían actividad de la enzima.26 Estudios posteriores han confirmado una actividad incrementada de telomerasa en cáncer de mama,27 próstata,28 astrocitomas29 y otros. Estos hallazgos pudieran tener importantes implicaciones clínicas. La determinación de la actividad de telomerasa pudiera ser utilizada para el diagnóstico precoz del cáncer en pruebas no invasivas 30 y los inhibidores de la enzima pudieran ser usados como agentes antitumorales con un alto grado de selectividad para las células transformadas. PERSPECTIVAS El descubrimiento de la singular estructura nucleoproteínica de los telómeros y su conservación filogenética estructural y funcional demuestran el carácter esencial de esas estructuras para la vida de la célula. La existencia de las telomerasas soluciona el viejo problema sobre la replicación de los extremos de moléculas lineales de ADN. Sin embargo, estos hallazgos plantean nuevos problemas. Entre ellos está determinar la función del ARNtl en la zona que no funciona como molde y su posible participación en la catálisis enzimática. Los factores que regulan la actividad de la enzima por una parte y la longitud de los telómeros por otra. Dilucidar los mecanismos moleculares que vinculan los telómeros con la regulación de la proliferación celular. También será interesante y trascendental la posible aplicación de estos conocimientos para el diagnóstico y tratamiento de enfermedades proliferativas, especialmente el cáncer. Para dar respuesta a estos y otros problemas deben encaminarse las futuras investigaciones. 127 SUMMARY Telomeres are specialized chromatin structures located in the extremes of eukaryotic chromosomes. Both their DNA and proteins present singular characteristics that differentiate them from the rest of chromosomes. They seem to be involved in a member of cell functions, especially those related to the control of life span of different cell species. These structures replicate in the cell cycle thanks to the activity of telomerases. which are enzimes composed by proteins and RNA with a peculiar mechanism. Recent research works have studied the behaviour of telomerases in cancer cells and their possible diagnostic and therapeutical uses. This paper shows a summary of the main and most recent findings on the structure and functions of telomeres and the action by telomerases. Subject headings: TELOMERASE/physiology; TELOMERE/physiology; CELL CYCLE. REFERENCIAS BIBLIOGRÁFICAS 1. Blackburn EH, Szostak JW. Molecular structure of centromeres and telomeres. Ann Rev Biochem 1984;53:16394. 2. Zakian VA. Telomeres beginning to understand the end. Science 1995;270:1601-7. 3. Viria Peariman V, Morris DK, Lundblad V. Est 1 has the properties of a single-stranded telomere endbinding protein. Genes Dev 1996;10:3094-104. 4. Chong L, Stensel B van, Broccoli D, Erdjument-Bromage H, Hanish J, Tempst P, et al. A human telomeric protein. Science 1995;270:1663-7. 5. Smith S, Lange T de. TRF1, a mammalian telomeric protein. Trends Genet 1997;13:21-6. 6. Cooper JP, Nimmo ER, Alishire RC, Cech TR. Regulation of telomeric lenght and function by a mybdomain protein in fission yeast Nature 1997;385:744-7. 7. Collins K. Structure and function of telomerase. Curr Opin Cell Biol 1996;8:374-80. 8. Hammond PW, Live TN, Cech TR. The anchor site of telomerase from Euplotes aediculatus revealed by photo-cross-linling to single- and double-stranded DNA primers. Mol Cell Biol 1997;17:296-308. 9. Battachay A, Blackburn EH. A functional telomerase RNA swap in vivo reveals the importance of nontemplate RNA domains. Proc Natl Acad Sci USA 1997;94:2823-7. 10. Lingner J, Hughes TR, Shevchenko A, Mann M, Lundblad V, Cecch TR. Reverse transcriptase motifs in the catalytic subunit of telomerase. Science 1997;276:561-7. 11. Harrington L, McPhail T, Mar V, Zhou W, Oulton R, Bass MB, et al. A mammalian telomerase-associated protein. Science 1997;275:973-7. 12. Nakayama J, Saito M, Nakamura H, Matsuura A, Ishikawa F. TLP1: a gene encoding a protein component of mammalian telomerase is a novel member of WD repeats family. Cell 1997;88:875-84. 13. Lendva TS, Morris DK, Sah J, Balasubramanian B, Lundblad V. Senescence mutants of Saccharomyces cerevisiae with a defect in telomere replication identify three additional EST genes. Genetics 1996;144:1399-412. 14. Shimada Y, Nakano M, Kanda N, Murakami-Murofushi K, Kim JK, Ide T, et al. Cycle-dependent activation of telomerase in naturally synchronized culture of a true slime mold, Physarum polycephalum. Biochem Biophys Res Commun 1997;232:4921-6. 15. Greider C. Telomere length regulation. Ann Rev Biochem 1996;65:337-5. 16. Steensel B van, Lange T de. Control of telomere length by the human telomeric protein. Nature 1997; 385:740-3. 17. Pommier JP, Lebeau J, Ducray C, Sabatier L. Chromosomal instability and alteration of telomere repeat sequence. Biochimie 1995;77:817-25. 18. Kirk KE, Harmon BP, Reichardt IK, Sedat JW, Blackburn EH. Block in znaphase chromosome separation caused by telomerase template mutation. Science 1997;275:1478-81. 19. Chiu CP, Harley CB. Replicative senescence and cell inmortality: the role of telomeres and telomerase. Proc Soc Exp Biol Med 1997;214:99-106. 20. Counter CM. The roles of telomeres and telomerase in cell life span. Mutat Res 1996;366:45-63. 21. Wright WE, Piatyszeck MA, Rainey WE, Byrd W, Shay JW. Telomerase activity in human germline and embryonic tissues and cells. Dev Genet 1996;18:173-9. 128 22. Hsiao R, Sharma HW, Ramakrishnan S, Keith E, Narayanan R. Telomerase activity in normal human endothelial cells. Anticancer Res 1997;17:827-32. 23. Saito T, Scheneider A, Martel N, Mizumoto H, Bulgay-Moeschel M, Kudo R, et al. Proliferation-associated regulation of telomerase activity in human endometrium and its potencial implication in early cancer diagnosis. Biochem Biophys Res Commun 1997;321:610-4. 24. Weng N, Levine BL, June CH, Hodes RJ. Regulation of telomerase RNA template expression in human T lumphocyute development and activation. J immunol 1997;158:3215-20. 25. Igarashi H, Sakagushi N. Telomerase activity is induced in human peripheral B lumphocytes by the stimulation to antigen receptor. Blood 1997;89:1299-307. 26. King NW, Piatyszeck MA, Prowse KR, Harley CB, West MD, Ho PLC et al. Specific assoiation of human telomerase activity with inmortal cells and cancer. Science 1994;266:2911-5. 27. Nawaz S, Hashizumi TL, Markkhan NE, Shroyer AL, Shroyer KR. Telomerase expression in human breast cancer with and without lumph node metastases. Am J Clin Pathol 1997;107:542-7. 28. Lin Y, Uemura H, Fujinami K, Hosaka M, Harada M, Kubota Y. Telomerase activity in primary prostate cancer. J Urol 1997;157:1161-5. 29. Sallinen P, Miettinen H, Sallinen SL, Haapasalo H, Helin H, Kononen J. Increased expression of telomerase RNA component is associated with increased cell proliferation in human astrocytomas. Am J Pathol 1997;150:1159-64. 30. Shay JW, Gazdar AF. Telomerase in the early detection of cancer. J Clin Pathol 1997;50:106-9. Recibido: 29 de septiembre de 1998. Aprobado: 19 de julio de 1999. " Dr. Rolando A. Hernández Fernández. Instituto de Ciencias Básicas y Pereclínicas " Victoria de Girón", Calle 146 y Ave. 31, Playa, Ciudad de La Habana, Cuba. 129 Rev Cubana Invest Biomed 1999;18(2):130-7 Centro de Investigaciones de Energía Solar. Santiago de Cuba ADYUVANTES INMUNOLÓGICOS Lic. Humberto Joaquín Morris Quevedo, Lic. Clara Martínez Manrique, Lic. Roberto T. Abdala Díaz e Ing. Denia Campos Orama RESUMEN La búsqueda de nuevas sustancias con actividad adyuvante/inmunopotenciadora constituye una de las tendencias más importantes en la investigación inmunológica actual. El presente trabajo considera los criterios fundamentales que determinan la selección de un adyuvante con fines investigativos, en el campo de la salud veterinaria y en la formulación de vacunas humanas, con el objetivo de incrementar la efectividad de la respuesta inmune. Se analizan brevemente las principales dificultades identificadas en la literatura, relacionadas con la comparación de la efectividad de diferentes adyuvantes, y se destacan las amplias posibilidades que ofrecen los productos naturales, especialmente polisacáridos de diversas fuentes, en la tamización de sustancias con propiedades inmunoestimulantes. Descriptores DeCS: ADYUVANTES INMUNOLOGICOS; VETERINARIA EN SALUD PUBLICA. Las sustancias inmunomoduladoras constituyen una familia muy heterogénea si se toman en consideración su origen, naturaleza química y actividad biológica específca.1 Dentro de ellas ocupan un lugar muy importante los agentes inmunopotenciadores, a los cuales se les atribuyen 2 funciones fundamentales: la estimulación de la resistencia no específica del huésped contra las enfermedades infecciosas y el cáncer; y, por otra parte, la potenciación de la inmunogenicidad de las vacunas comerciales y de la respuesta de los animales de laboratorio durante la inmunización experimental con vistas a la producción de antisueros. 2 Los avances experimentados en la última década en la 130 síntesis de péptidos, la secuenciación de nucleótidos y la ingeniería genética han posibilitado el desarrollo de las denominadas "vacunas de nueva generación";3 sin embargo, la utilización exitosa de estas nuevas tecnologías requiere de un esfuerzo investigativo sostenido para hallar sustancias con actividad inmunopotenciadora (adyuvantes), eficaces en la estimulación de la respuesta inmune y desprovistas de propiedades biológicas adversas.4 Los adyuvantes son sustancias o preparados químicos que, incorporados al antígeno o inyectados simultáneamente con él, hacen más efectiva la respuesta inmune. Con su empleo se logra una economía de antígeno y de tiempo, así como un mayor nivel de anticuerpos específicos.5 El mecanismo de acción de estas sustancias ha sido objeto de numerosos estudios y, al parecer, existen diversos factores que explican su modo de acción. El antígeno libre normalmente difunde con mucha rapidez desde los tejidos locales que rodean el sitio de inoculación, y una de sus funciones importantes es crear un reservorio o depósito del antígeno de larga vida. Las investigaciones realizadas han demostrado que virtualmente todos los adyuvantes activan o estimulan los macrófagos;6 éstos cuando son activados estimulan la respuesta inmune por un incremento de la cantidad de antígeno expresado en la membrana celular y de la eficiencia de su presentación a los linfocitos. El macrófago también libera factores solubles estimulantes, que amplifican la proliferación de los linfocitos.7 Por otro lado, algunos adyuvantes poseen la capacidad de actuar específicamente sobre los linfocitos; pero, en general, éstas funcionan mejor si facilitan la liberación simultánea del antígeno y de sustancias inmunomoduladoras al tejido linfoide.8 En el nivel internacional, la lista de productos naturales y derivados de la síntesis química, con propiedades adyuvantes/inmunopotenciadoras, es cada vez mayor; sin embargo, sólo un reducido número se utiliza en la formulación de vacunas veterinarias y humanas, existiendo una tendencia relacionada con la evaluación de nuevas sustancias con esta finalidad. En función del campo de aplicación y de la relación eficiencia/seguridad se pueden reconocer diferentes categorías de adyuvantes (tabla),que serán examinadas a continuación con más detenimiento. SELECCIÓN DE UN ADYUVANTE CON FINES INVESTIGATIVOS Con fines investigativos, las exigencias más importantes se relacionan con una elevada eficacia, un amplio espectro de aplicación, una fácil manipulación y, por supuesto, la disponibilidad comercial. El adyuvante completo de Freud (FCA) ha sido utilizado durante más de 50 años TABLA. Relación entre las aplicaciones de los adyuvantes y la relación eficiencia/seguridad Adyuvantes FCA Emulsiones aceite/agua Emulsiones agua/aceite Materiales inertes Compuestos de aluminio Liposomas Saponinas DDA NBP La eficiencia es más importante que la seguridad Animales Animales para de laboratorio la alimentación La seguridad es más importante que la eficiencia Animales Humanos de compañía + + - - + (-) - + + - - + + + + + + (+) + + (+) (+) (+) (+) + (+) (+) (+) (+) (-) + (-) (-) (-) (-) Leyenda +: aplicado de rutina en productos comerciales; (+): no aplicados, pero se considera aplicable en función de su seguridad; –: no aplicado y considerado no aplicable por problemas de seguridad; (-): no aplicado y considerado no aplicable, excepto para fines muy específicos. 131 en la producción de antisueros en animales9 y es empleado con frecuencia cuando se dispone de cantidades limitadas de antígenos o cuando presentan una baja inmunogenicidad. La gravedad de su toxicidad fue reconocida inmediatamente, pero los esfuerzos por encontrar opciones igualmente efectivas y menos tóxicas no han resultado del todo exitosos. Con los avances alcanzados en numerosas áreas de las ciencias biológicas y el creciente interés por el bienestar de los animales de experimentación, existe una considerable presión para restringir el uso del FCA, y aunque no se cuenta en la actualidad con alternativas definitorias para este adyuvante, en diferentes estudios comparativos se informan resultados estimulantes en cuanto a eficiencia, menor número de efectos adversos y mayor facilidad en la manipulación.8 En la investigación pueden utilizarse, además, los compuestos de aluminio, la formulación "Syntex-1" (SAF-1) que contiene el treonil análogo del dipéptido murámico, polímeros no iónicos en bloque (NBP), saponinas, bromuro de dimetil dioctadecil amonio (DDA), lipopéptidos, etc. SELECCIÓN DE UN ADYUVANTE EN EL CAMPO DE LA SALUD VETERINARIA En este campo, la eficacia es un elemento de gran importancia y se toleran ciertos niveles de efectos colaterales. Los adyuvantes más ampliamente utilizados en las vacunas veterinarias son las emulsiones de aceite mineral (del tipo aceite en agua o agua en aceite) y los adsorbentes (hidróxido y fosfato de aluminio). En algunos casos, se emplean liposomas, saponinas, vitamina E, complejos inmunoestimulantes 132 (ISCOMs), así como diferentes emulsiones de aceites de origen vegetal o animal. Las emulsiones de aceite mineral, especialmente las del tipo agua en aceite, si bien inducen una fuerte respuesta inmune, pueden provocar riesgos y efectos no deseados, a causa posiblemente de su limitada biodegradabilidad y biocompatibilidad.10 Por tal razón, se han realizado numerosos intentos para desarrollar adyuvantes eficaces y a la vez seguros. Ejemplo de ello es la formulación compuesta por sulfolipopolisacáridos sintéticos, con características hidrofóbicas, incorporados a una emulsión de escualeno en agua, desarrollada por la compañía belga Solvay SA. Este adyuvante ha sido validado exitosamente en animales de laboratorio con antígenos proteicos y virales, y resulta, al menos, tan efectivo como los adyuvantes basados en aceite mineral, empleados hoy día en diversas vacunas veterinarias.11 ADYUVANTES PARA VACUNAS HUMANAS El criterio más importante, sin lugar a dudas, para la selección de un adyuvante destinado a vacunas humanas es la bioseguridad y, en la práctica, los compuestos de aluminio son los únicos adyuvantes licenciados para uso humano.3 El hidróxido de aluminio tiene un punto isoeléctrico de 11,1 y, por tanto, está cargado positivamente en las condiciones biológicas, por lo que puede adsorber proteínas cargadas negativamente y adsorber débilmente las proteínas básicas.El grado de adsorción depende de la naturaleza y concentración del antígeno, de la presencia de sales e iones bufferantes y del pH de la mezcla resultante.4 Las fuerzas de atracción electrostáticas y las interacciones hidrofóbicas son las responsables de la adsorción de los antígenos a los adyuvantes que contienen aluminio, pero probablemente las fuerzas de Van der Waals y los enlaces de hidrógenos también contribuyan a la adsorción en estos sistemas;12 sin embargo, se ha señalado que el alumbre puede inducir anticuerpos de la clase IgE, así como reacciones alérgicas.13 Según las regulaciones de la FDA, son permisibles cantidades no mayores de 0,85 mg de aluminio por dosis de vacuna.14 Las nuevas vacunas recombinantes y sintéticas poseen una baja inmunogenicidad en comparación con las vacunas tradicionales, consistentes en los microorganismos intactos, atenuados o inactivados por el calor; de ahí el interés por la búsqueda de diferentes inmunoadyuvantes para incrementar la efectividad de estas nuevas vacunas para uso humano.3 En este sentido, Tamura y otros,15 notifican en estudios realizados en ratones, la utilidad de 2 derivados del dipéptido murámico como adyuvantes para la vacuna recombinante de la hepatitis B, primera de esta categoría en ser licenciada para uso en humanos y disponible comercialmente en forma adsorbida al alumbre. La respuesta de anticuerpos obtenida con los derivados del MDP resultó similar a la inducida por la vacuna adsorbida en alumbre y se discute, por los autores, el posible papel de estos compuestos como su sustituto. ASPECTOS PRÁCTICOS En los estudios relacionados con la evaluación de la efectividad de diferentes adyuvantes se impone dejar reglamentadas líneas generales, que permitan solucionar las principales dificultades existentes en este campo en la actualidad, entre las que pueden citarse: 1. No existencia de criterios estandarizados para determinar la eficacia de los adyuvantes; 2. la mayoría de los estudios adolecen de formulaciones estandarizadas como referencias apropiadas; 3. los niveles de inmunoestimulación alcanzados con los distintos adyuvantes no se indican de forma explícita en los trabajos publicados; 4. en general, las condiciones para lograr la eficiencia óptima (tipo de antígeno, ruta de inmunización, especies animales, etc.) no han sido establecidas. Estos aspectos han sido reconocidos por Stewart-Tull,4-16 quien propuso una estrategia para la estandarización de la eficacia de los adyuvantes, la cual contempla la utilización de la ovoalbúmina y el virus de la influenza como antígenos estándares, el empleo de adyuvantes de referencia y parámetros definidos por la inmunidad (determinación de los títulos de anticuerpos por ELISA), entre otros criterios. PROPIEDADES INMUNOPOTENCIADORAS DE POLISACÁRIDOS NATURALES Las propiedades inmunomoduladoras han sido demostradas en numerosos polisacáridos naturales y algunos derivados obtenidos por hidrólisis o modificación química. La actividad estimulante de la hematopoyesis y de la respuesta inmune humoral y celular es uno de los rasgos más significativos de los glucanos aislados de la pared celular de levaduras. 17 Ha sido mostrado por diferentes autores que la administración simultánea del glucano y un antígeno estimula la formación de anticuerpos específicos contra el antígeno en cuestión; prueba de ello es la estimulación por los glucanos de la respuesta humoral y mediada por células contra Francisella tularensis y Pseudomonas 133 pseudomallei.18 Además, se ha descrito la estimulación de la síntesis de anticuerpos contra Entamoeba histolytica en curieles después de la administración del glucano19 y se ha reconocido el efecto adyuvante en Leishmania major, Mycobacterium leprae, Candida albicans, Herpes simplex y el virus de la hepatitis murina.20 Por otra parte, la administración oral de un extracto de glucanos (Glucanil) de Candida albicans ATCC 20995, suministrado a ratones 10 días antes de la infección experimental con Candida albicans y Staphylococcus aureus, incrementó significativamente la resistencia a la infección sistémica con estos microorganismos y los estudios toxicológicos revelaron que el producto era altamente tolerado.21 En algunos experimentos, la actividad del glucano resultó superior a la de adyuvantes usados comúnmente. El efecto del glucano depende del modo de administración, y aunque este último puede ser realizado de forma repetida, el efecto estimulatorio suele ocurrir después de una inyección única.22 Los hongos constituyen otra fuente valiosa de polisacáridos con actividad inmunomoduladora. En este sentido, posiblemente el ejemplo más estudiado sea el lentinan, un B (1-3) D-glucano con ramificaciones B(1-6) D-glucopiranósidas, aislado del hongo comestible Lentinus edodes, el cual es capaz de restablecer o aumentar la capacidad de respuesta del huésped a las citoquinas u otros factores bioactivos intrínsecos mediante la estimulación de la maduración, diferenciación o proliferación, o ambas, de las células involucradas en los mecanismos de defensa del huésped.23 El lentinan también incrementa la resistencia del huésped a varios tipos de infecciones bacterianas, virales y parasíticas y su contribución a la restauración de la disfunción inmunológica ha sido demostrada en pacientes con neoplasias.24 134 En la literatura médica existen referencias, además, de la actividad inmunomoduladora del esquizofilan, aislado del hongo Schizophyllum commune y de 2 polisacáridos (AT-HW y AT-AL) de los cuerpos fructíferos de Armillariella tabescens, los que ejercen un efecto potenciador del sistema reticuloendotelial y de la actividad mitogénica y su administración está acompañada de un incremento en el número de células del exudado peritoneal y de la activación de los macrófagos.25 La activación del sistema del complemento (vías clásica y alternativa) por B(1-3) D-glucanos, presentando diferente ultraestructura y grado de ramificación aislados de la pared celular de hongos, ha sido demostrada con el empleo de suero y plasma humanos.26 Por otra parte, en estudios realizados tanto in vitro como in vivo, la fracción polisacarídica ha estimulado las funciones fagocíticas de los macrófagos murinos, las cuales dependen en gran medida del contenido de grupos sulfato en su estructura.27 También se ha comprobado que las propiedades inmunoestimulantes de numerosos preparados que se usan tradicionalmente en la Medicina Oriental, son atribuibles a diferentes fracciones polisacarídicas presentes en su composición.28 Desde el punto de vista de la inmunización de especies de interés económico, se han alcanzado resultados muy prometedores con el empleo de antígenos enlazados a polisacáridos, como el J-carragenano en aves y el sulfato de dextrano en el ganado,29 los cuales confirieron protección a los animales en un encuentro posterior con los citados antígenos. En ese contexto, durante los últimos años se han dado importantes pasos en la utilización de polisacáridos como adyuvantes inmunológicos. A modo de ejemplo pode- mos mencionar la formulación AdjuvaxTM (Alpha-Beta Technology, Worcester, MA, USA), cuyo componente con actividad inmunomoduladora es un polisacárido de la clase de los glucanos, donde el antígeno se incorpora a la matriz polisacarídica con una breve retención; y el polímero acetilado de la manosa, registrado comercialmente como Acemannan (Carrington Labs. Inc., TX, USA), que se emplea en la vacuna contra el virus de la enfermedad de Marek en las aves.30 Dentro del universo microbiano, las algas microscópicas comprenden un grupo muy diverso de microorganismos fotosintetizadores de más de 30 000 especies, que se caracterizan por presentar una elevada velocidad de crecimiento y una gran eficiencia en el uso de la energía luminosa. Los efectos promotores de la salud humana atribuidos a las algas se conocen desde tiempos inmemoriales, a causa de las antiguas experiencias en su uso por los aztecas y agrupaciones del África Central.31 Estas acciones positivas sobre el estado fisiológico general del organismo se han relacionado con la activación de los mecanismos de protección inmunitaria inherentes a él por sustancias naturales de elevado potencial terapéutico, entre las cuales los polisacáridos adquieren un valor especial.32 Tomando en consideración estos elementos, existe actualmente un gran interés por el estudio de la posible actividad inmunopotenciadora de las fracciones polisacarídicas de las microalgas (tanto intracelulares como exocelulares), pues la evaluación del efecto farmacológico de estas sustancias depende de manera primaria del establecimiento de condiciones definidas de cultivo. Al respecto, las propiedades inmunorreguladoras de ciertos extractos de Chlorella han sido atribuidas a su importante composición polisacárida [Centro de Investigaciones de Energía So- lar. Informe de la revisión de patentes sobre extractos de microalgas. Santiago de Cuba, 1994] y, por otra parte, se ha observado que la fracción polisacarídica intracelular de Spirulina platensis estimula la proliferación de los linfocitos T y las células NK, por lo cual podría resultar sumamente válido para la investigación sobre tumores.33 Por otro lado, en experimentos in vitro con leucocitos mononucleados de sangre periférica humana se ha demostrado un efecto estimulante de la síntesis de anticuerpos (inmunoglobulinas) de 3 fracciones polisacarídicas del alga roja unicelular Porphyridium cruentum, sugerente de una actividad moduladora sobre los linfocitos B.33 El mecanismo de dicha estimulación es aún desconocido. Por todo lo anterior concluimos que por razones de naturaleza práctica y económica, tanto en la investigación inmunológica fundamental como en la aplicación de los conocimientos derivados de ella en la producción de antisueros y vacunas, reviste una importancia primordial la utilización de los adyuvantes/inmunopotenciadores. De particular interés resulta la valoración de nuevos inmunoadyuvantes para incrementar la efectividad de las vacunas recombinantes o sintéticas, las que poseen una baja inmunogenicidad en comparación con las vacunas formuladas tradicionalmente. En este sentido, se impone establecer líneas generales que permitan solucionar las principales dificultades existentes hoy día, relacionadas con la comparación de la efectividad de diferentes adyuvantes. El estudio de los productos naturales, incluidos polisacáridos de diversas fuentes, ofrece amplias perspectivas para la búsqueda de nuevas sustancias con actividad inmunopotenciadora y desprovistas de propiedades biológicas adversas. 135 SUMMARY The search for new substances with inmunological adjuvant activity constitutes one of the most important trends in today’s inmunological research work. The present paper analyzes the fundamental criteria that determine the selection of an adjuvant for research purposes in the veterinary health field and in human vaccine formulation, with a view to increasing effectiveness of inmune response. The main difficulties mentioned in literature and related to the comparison of the effectiveness of various adjuvants are briefly analyzed, and also the wide possibilities of natural products especially polysaccharides of various sources in the selection of substances with inmunostimulating qualities. Subject headings: ADJUVANTS, INMUNOLOGIC; PUBLIC HEALTH VETERINARY. REFERENCIAS BIBLIOGRÁFICAS 1. Takx-Kohlen BC. Immunomodulators. Future prospects. Pharm Weekbl Sci 1992;14:24-52. 2. Azuma I. Synthetic immunoadjuvants: application to nonspecific host stimulation and potentiation of vaccine immunogenicity. Vaccine 1992;10:1000-6. 3. Grupta RK, Relyveld EH, Lindblad BE, Bizzine B, Ben-Efraim S, Grupta CK. Adjuvants-a balance between toxicity and adjuvancicity. Vaccine 1993;11:293-306. 4. Stewart-Tull DES. Recommendations for the assessment of adjuvants (immunopotentiators). En: Gregoriadis G, Allison AC, Poste G, eds. Immunological adjuvants and vaccines. New York: Plenum, 1989:213-26. 5. Roitt IM. essential immunologyu. 7 ed. Oxford: Blackwell Scientific, 1991:326. 6. Guerrero J, Gattas CR. Immunolomodulating substances: an overview. Rev Microbiol 1982;13(2):110-5. 7. Wagnerová J, Ferencík M. Secretory and regulatory products of macrophagues in the immune and inflamatory reactions. Biol Bratislav 1993;48(6):709-17. 8. Bennet B, Check IJ, Olsen MR, Hunter RL. A comparison of comercially available adjuvants for use in research. J Immunol Methods 1992;153:31-40. 9. Freund J. the mode of action of immunological adjuvants. Adv Tuberc Res 1956;7:130-48. 10. Straw BE, Maclachlan NJ, Cobertt WT, Carter PB, Schey HM. comparison of tissue reactions produced by Haemophilus pleuropneumoniae vaccines made with six different adjuvants in swine. Can J Comp Med 1985;49:149-51. 11. Hilgers LAT, Platenburg PLI, Luitjens A, Groenveld B, Dazelle T, Ferrari-Laloux M, et al. A novel nonmineral cil-based adjuvant. I. Efficacy of a synthetic sulfoli-popolysaccharide in a squalene-in water emulsion in laboratory animals. Vaccine 1994;12:653-60. 12. Al-Shakshir RH, Regnier FE, White JL, Hem SL. Contribution of electrostatic and hydrofobic interactions to the adorption of proteins by aluminium-containing adjuvants. Vaccine 199513:41-4. 13. Veien NK, Hattel T, Justesen O, Norholm A. Aluminium allergy. Contact Dermatitis 1986;15:295-7. 14. May JC, Progar JJ, Chin R. The aluminium content of biological products containing aluminium adjuvants: determination by atomic absorption spectrometry. J Biol Stand 1984;12:175-83. 15. Tamura M, Yoo GC, Yoshimatsu K, Yoshida R, Oka T, Ohkuma K, et al. Effects of muramyi dipeptide derivatives as adjuvants on the induction of antibody response to recombinant hppatitis B surface antigen. Vaccine 1995;13:77-82. 16. Stewart-Tull DES. The assessment and use of adjuvants. En: Gregoriadis G, allison AC, Poste G, eds. Vaccine, recent trends and progress. New York: Planum Press, 1991:85-92. 17. DiLuzio NR. Update on the immunomodulating activities of glucans. Springer Semin Immunopathol 1985;8:387-400. 18. Reynolds JA, Kastello MD, Herrigton DG, Crabbs CL, Peters CJ, Jemski JV, et al. Glucan-induced enhancement of host resistance to selected infecious diseases. Infect Immun 1980;30:51-7. 19. Sharma A, Haq AU, Siddiqui MV, Ahmad S. Immunization of Guinea pigs against Entamoeba histolytica using glucan as an adjuvant. Int J Immunopharm 1984;6:483-91. 20. Di-Luzio NR. Immunopharmacology of glucan: A broad spectrum enhancer of host defense mnechanisms. Trends Pharmacol Sci 1983;4:344-7. 21. Nicolette A, Nicoletti G, Ferraro G, Palmiei G, Mattaboni P, Germogli R. Preliminary evaluation of immunoadjuvant activity of an orally administeered glucan extracted from Candida albicans. Arzneimittelforscgung 1992;42:1246-50. 136 22. Wagnerová J, Lískova A, gervenakova L, Trnovec T, Ferencik M. The immunoadjuvant effect of soluble glucan derivatives in mice. Folia Microbiol 1991;36:198-204. 23. Chihara G. Recent progress in immunopharmacology and therapeutic effects of polysaccharide. Develop Biol Stand 1992;77:191-7. 24. Susuki M, Takatsuki F, Maeda YY, Hamuro J, Chihara G. Lentinan-rationale for development and therapeutic potential. Clin Immunother 1994;2:121-33. 25. Kiho T, Shiose Y, Nagai K, Ukai S. Polysaccharides in fungi. XXX. 1) Antitumor and immunomodulating activities of two polysaccharides from the fruiting bodies of Armilariella tabescens. Chem Pharm Bull 1992;40:2110-4. 26. Susuki T, Ohno N, Saito K, Yadomae T. Activation of the complement system by (1-3) beta D-glucans having different degrees of branching and different ultrasturctures. J Pharmacobiodyn 1992;15:277-85. 27. Yoshizawa Y, Ametani A, Tsunehiro J, Nomura K, Itoh M, Fukui F, et al. Macrophage stimulation activity of the polysaccharide fraction from a marine alga Polphyra yezoensis: structure-function relationsphips and improved solubility. Biosci Biotechnol Biochem 1995;59(10):1933-7. 28. Yamada H. Chemical and pharmacologicalk studies on efficacy Japanese and Chinese herbal medicines. Kitasato Arch of Exp Med 1992;65:159-80. 29. Kungú MW, Goodger BV, Bushell GR, Wright IG, Waltisbuhl DJ. Vaccination of cattle with dextran sulphatebinding Babesia bigemina. Int J Parasitol 1992;22:621-5. 30. Muirihead S. Solvay introduces new immune stimulant product. Feedstuffs 1992;64:2. 31. Furst PT. Spirulina. Hum Nature 1978;60:60-5. 32. Pulz O, Koehler E. Microalgae as a source of pharmacologically valuable polusaccharides. Proceeding of the 6th European Congress on biotechnology. firenze June 13-17. 1993,40-1. 33. Pulz O, Koehler E. Pharmakologische Wirkung von Polysacchariden aus Microalgen. Deutsche Gesellschaft Qualitätsforschung (Pflänzliche Nahrungsmittel) Quidlinburg März 21-22. 1994,253-64. Recibido: 9 de febrero de 1999. Aprobado: 19 de julio de 1999. Lic. Humberto Joaquín Morris Quevedo. Centro de Investigaciones de Energía Solar, Santiago de Cuba. 137 Rev Cubana Invest Biomed 1999;18(2):138-45 Hospital General "Ciro Redondo". Artemisa BOCIO DIFUSO EUTIROIDEO: 100 AÑOS DE TRATAMIENTO Dr. Mayque Guzmán Cayado RESUMEN El Bocio Difuso Eutiroideo (BDE) constituye hasta hoy día uno de los temas polémicos en cuanto a su patogenia, y por tanto, a su tratamiento. La terapéutica a base de hormonas tiroideas (HT), está sujeta a una serie de indicaciones que han variado con el decursar de los años y para muchos es puesta en duda por la alta tasa de recidiva tras su suspensión, así como por la aparición de reacciones adversas. La cirugía consta de indicaciones cada vez más precisas y es cuestionable su operatividad. Con el presente trabajo nos proponemos poner de manifiesto los principales factores involucrados en su patogenia y los criterios seguidos para su tratamiento así como los resultados de algunos autores con ambos tratamientos: médico y quirúrgico. Descriptores DeCS: BOCIO/etiología; BOCIO/terapia; HORMONAS TIROIDEAS/uso terapéutico. DEFINICIÓN, ETIOLOGÍA Y PATOGENIA El bocio difuso eutiroideo (BDE), bocio simple, bocio no tóxico o bocio esporádico, se define como todo aquel aumento de volumen de la glándula tiroides, no asociado a hiper o hipotiroidismo, y que además no resulta de un proceso infeccioso o neoplásico de dicha glándula; sus causas son las siguientes:1 1. Déficit de yodo. 2. Alimentos o medicamentos bociógenos: yoduros (tratamiento de la EPOC), carbonato de litio (antipsicótico), verduras (tioglucósidos de coles y bocioína de raíces y semillas) y mandioca (glucósidos cianógenos). 3. Menarquia, embarazo y anticonceptivos orales (AO). 138 4. Tiroiditis de Hashimoto. 5. Tiroiditis subaguda. 6. Síntesis hormonal inadecuada por defecto enzimático hereditario: Disminución del transporte de yodo, déficit de peroxidasa (TPO), disminución del acoplamiento de tirosinas, disminución o ausencia de desyodasa de yodotirosinas de forma que el yodo no se conserve dentro de la gándula y aumento de la producción de yodoproteínas metabólicamente inactivas en el tiroides. 7. Deficiencia hereditaria en los receptores de tiroxina en la membrana de la célula. 8. Neoplasias benignas y malignas (esta última causa aparece excluida conceptualmente). La teoría más tradicional, en cuanto a su patogenia, plantea que el bocio representa una respuesta compensadora ante una gama de factores que, de una forma u otra, afectan su eficacia en la elaboración de hormonas tiroideas (HT), lo que provocaría disminución de sus niveles plasmáticos y activación del mecanismo de retroalimentación hipofisario con hipersecreción de tirotropina (TSH), o quizás un aumento de la sensibilidad de los tirocitos a esta última, lo cual incrementa la actividad hormonosintetizadora y, además, produce aumento de volumen de la gándula; resultando en un paciente eutiroideo pero bocioso. En muchos de estos sujetos resulta imposible demostrar la presencia de un factor bociogénico extrínseco, por lo que desde los estudios iniciados por McGir y Hutchinson y sobre todo por Stanbury en 1955, se ha especulado que causas intrínsecas, probablemente congénitas sean las responsables del bocio.2-8 Se piensa en una causa autoinmune del BDE en aquellos pacientes en quienes concomitan otros fenómenos de tipo inmunológico y en los que el bocio recidiva después de la cirugía. Los estudios que apoyan esta teoría son pocos y se requiere de su ampliación.9,10 En la década del 80, Drexhage y otros y McMullan describieron la presencia de un tipo de inmunoglobulinas (IgG) estimuladoras del crecimiento tiroideo (TGI) en pacientes con enfermedad de Graves y de Hashimoto, con bocios grandes, y en algunos pacientes con BDE.11 Más recientemente Davies y otros, reportan haberlas encontrado en el 64 % de los pacientes con bocio esporádico. 12 Estas inmunoglobulinas parecen no actuar por el mecanismo de la adenilciclasa ni unirse específicamente al receptor de TSH.13 La posibilidad de cultivar células tiroideas en medios libres de suero ha permitido demostrar que no sólo la TSH influye en el crecimiento tiroideo, sino que existen otros factores de crecimiento como: el de tipo insulínico (IGF-1), el epidérmico (EGF) y el de fibroblastos (FGF), por mencionar algunos, capaces de lograr el mismo efecto.14-18 PREVALENCIA Y CUADRO CLÍNICO Su prevalencia es variable, según tipo de población. Se estima que el 4 % de la población general19 y alrededor del 5 % de la población de Estados Unidos,20 sufre esta afección. Al igual que el resto de las tiroidopatías es más frecuente en el sexo femenino.21-25 Es también más frecuente durante la adolescencia y durante el embarazo,26-29 en el primero de los casos se debe a un aumento de los estrógenos en esta etapa, lo que conlleva a un efecto en el nivel hipofisario aumentando la concentración de los receptores para la TRH y la respuesta de las células tirotropas a dicho péptido; en el segundo de los casos hay un aumento en el aclaramiento de yodo y gran parte del yoduro y las yodotironinas pasan al feto a través de la placenta lo que crea en la madre un defecto de yodo y yodotironinas (hipotiroidismo relativo) que es compensado al activar el eje hipófisis-tiroides, aumenta la TSH y el tiroides se torna bocioso. 30 En algunos pacientes regresa después de estos eventos y en otros persiste. El cuadro clínico, en general, resulta del aumento de volumen de la glándula y los síntomas pueden ser de tipo mecánico: disfagia, por compresión del esófago; disnea, por compresión de la tráquea; dilatación venosa que se exacerba al alzar los 139 brazos (signo de Pemberton), por compromiso del retorno venoso; mareos y síncope, por compromiso arterial y de tipo estético. La compresión del nervio laríngeo recurrente sugiere carcinoma y el aumento de volumen y el dolor localizados, hemorragia. Desde el punto de vista bioquímico las determinaciones hormonales se encuentran en el rango normal. TRATAMIENTO MÉDICO La historia de las enfermedades tiroideas se remonta a las primeras épocas conocidas de nuestra historia. El papiro de Ebers (1 500 años a.n.e.) describe el bocio y señala, desde ese entonces, 2 posibles tratamientos: la resección quirúrgica y la ingestión de sales de un sitio particular del Bajo Egipto (presumiblemente ricas en yodo).31 Nuestros antepasados, de forma general, empleaban algas, esponjas y otros productos marinos que hoy conocemos son ricos en yodo, sin embargo en la segunda mitad del siglo XIX dicho tratamiento fue rechazado por la frecuente aparición del fenómeno de "Jod-basedow", de manera que las HT devinieron los medicamentos de mayor uso. En 1891, Murray comenzó a tratar a los pacientes mixedematosos con extracto de tiroides y posteriormente un médico de Edimburgo, el doctor Moyan Sunderland, fue el primero en sugerir el empleo de este medicamento en el bocio simple. Si realmente el paciente tiene una hipersecreción de TSH, el tratamiento con HT estaría justificado con el objetivo de suprimir esta hormona, sin embargo, sus concentraciones están dentro de límites normales en la mayoría de los pacientes,9,32,33 lo que hace cuestionable la utilidad terapéutica de este proceder. 140 La administración de las HT para ser eficaz debe ajustarse a las condiciones siguientes:34 ! Iniciarse en la fase temprana cuando no se ha producido la transformación nodular. ! La cantidad de HT debe ser una dosis subtóxica capaz de suprimir TSH, aproximadamente entre 180 mg de tiroides desecado (TD) o 200 mg de levotiroxina (L-T4). ! El tratamiento debe ser prolongado. La mejoría es proporcional al período de administración y el plazo para justificar la eficacia del tratamiento oscila entre 6 meses y 1 año. Las HT son recetadas por los médicos con frecuencia, unas veces con sólidos criterios de uso, otras no.35 La L-T4 sintética es la preparación tiroidea de uso más común en el tratamiento del hipotiroidismo y para la supresión de los niveles elevados de TSH en pacientes con nódulos tiroideos y/o aumento de volumen de la glándula.36 El 1,2 % de la población en Suecia recibe L-T4 sintética37 y alrededor de 18 000 000 de recetas de HT se llenan en Estados Unidos.38 Todo esto nos da una medida de la magnitud del uso de las HT en la práctica médica. En los pacientes provenientes de áreas endémicas de déficit de yodo, la yodoterapia ha demostrado ser el tratamiento de elección del BDE.39,40 Vitti y otros, en 1991 propusieron: – Pacientes < 40 años: dosis supresivas. – Pacientes > 40 años: dosis más juiciosa. – TSH basal para descartar autonomía. Después de los 60 años no se recomienda el uso de las HT por lo dudoso de su utilidad y por el peligro de las sobredosis. A pesar de que el paciente esté eutiroideo existe lo que se ha dado en llamar "hipertiroidismo químico" en pacientes tratados con L-T4 en los cuales las T4 está elevada por el agregado exógeno y clínicamente asintomáticos por estar la conversión de T4 a T3 disminuida; la T3 es el mayor indicador de posibilidad de hipertiroidismo en pacientes tratados con L-T4.36 En nuestro país se ha seguido históricamente la siguiente pauta [Navarro D, Álvarez E. Bocio eutiroideo. Conducta clínico-terapéutica. En: La glándula tiroides. La Habana:Editorial Ciencias Médicas, 1992:48-59]: – Bocio entre 40-60 g: dosis sustitutiva. – Bocio mayor que 60 g o nodular: dosis supresiva. Los reportes cada día más frecuentes, sobre la aparición de reacciones adversas a causa del uso crónico de las HT y a su sobredosificación como: taquicardia, arritmias, hipertensión arterial, embolismo cerebral, trastornos psiquiátricos e incluso muertes en ancianos 41-45 así como la desmineralización ósea en mujeres posmenopáusicas, 46 han llevado a la reconsideración de las bondades de dicha terapéutica en el BDE. El valor de la TSH basal o la respuesta de esta hormona a la estimulación con TRH, parecen ser de un valor indiscutible al descartar la presencia de autonomía tiroidea con supresión del eje hipotálamohipófisis-tiroides. La abstención terapéutica constituye una indicación cuando el bocio ya no es tributario de tratamiento médico, pero aún no existe una indicación clara de cirugía.30 TRATAMIENTO QUIRÚRGICO La cirugía de tiroides no comenzó a ser efectiva hasta finales del siglo XIX ya que, entre otras razones, la riqueza vascular del tejido hacía que ante cualquier maniobra quirúrgica se produjeran grandes hemorragias.31 Hoy día se plantea que el tratamiento quirúrgico del bocio es fisiológicamente inconsistente ya que reduce la habilidad del tejido para satisfacer las necesidades hormonales del individuo, sin embargo es indicada en los casos siguientes: 1. Síntomas obstructivos severos (disfagia y disfonía) a pesar del tratamiento médico con HT). 2. Cuestiones cosméticas. 3. Sospecha de que en su estadio multinodular albergue un carcinoma. 4. Nódulos únicos. 5. No tolerancia a las dosis requeridas para el tratamiento médico. Con respecto al uso preventivo de las HT en la recidiva posquirúrgica del bocio, un estudio prospectivo en el año 1994, de 9 años previos de seguimiento, concluyó en que el uso de L-T4 (100 mg/d) no logró ningún efecto sobre la incidencia de recidiva posquirúrgica.47 La tasa de recurrencia es baja, sin embargo, la tasa de morbilidad posquirúrgica es alta (disfonía frecuente); y en segunda, la enfermedad no parece curarse con la tiroidectomía. En la tabla se recogen los resultados de algunos autores, con el uso de las HT, en el bocio simple. Por todo lo anterior se concluye que el tratamiento del BDE, aparejado a los recientes descubrimientos en cuanto a su patogenia, ha resultado en nuevos enfoques terapéuticos. La cirugía ha perdido lugar y cada día se hace más cuestionable el empleo de las HT durante toda la vida. Si bien es cierto que en un porcentaje de los casos el bocio se reduce, en otros la aparición de peligrosas reacciones adversas 141 TABLA. Resultados de algunos autores durante estos 100 años de tratamiento en el BDE Ref. Autor(Año) n Medicamento Dosis diaria Duración Respuesta satisfactoria Observaciones * Bruns 1896 326 TD – – 75 48 Greer 1953 50 TD 2!3 g – 76 No influyeron la edad, sexo ni No TTO L-T3 Placebo L-T4 – 12 sem 15 57 el tiempo de evolución Posible inclusión de 200 mg 6-36 sem 10 57 hipotiroideos Mejor respuesta en bocios pequeños de etiología autoinmune L-T4 – – 75 Mejor respuesta en jovénes con menor T. Evol. TodosT. de Hashimoto Todos los pacientes tenían nódulos Disminución significativa en los primeros 3 meses. En los tratados con L-T4 hubo recidiva al suspender el TTO 49 Badillo 1963 40 21 16 21 75 mg – 50 Gutteridge 1978 51 Doniach 1979 – 52 Ashcrsft 1981 446 – – – 37 53 Perrild 1982 30 L-T4 100 mg 12m NS 33 L-T4 150 mg NS L-T4 L-T3 L-T4 Yodo L!T3 200 mg 60 mg 100 mg 500 mg 60 mg 12 m 55 17 30 Olbricht 1984 20 20 Faldt!Rasmussen 1984 19 p<0,05 p<0,01 70 4 p<0,01 56 Berghout 1989 Cirugía – – 82,3 Placebo L-T4 L-T4+CBZ L-T4 Yodo L-T4 L-T4 + Yodo – 9m 2,5 mg/Peg 2,5 mg/kg+40 mg 90-150 mg 150 mg 100 mg 50 mg + 100 mg 24 m 6m aumentó 58 35 p<0,001 18,5 a 8,8 ml 14,1 a 8,3 ml 17,2 a 8,3 ml 54 57 Berghout 1990 113 26 26 26 13 30 30 30 12 m Mejor respuesta en jóvenes con bocios recientes Respuesta evidente con L-T4 a los 3 meses No recidiva al suspender TTO En el 17,7 % recidivó el bocio Recidiva tras suspender el tratamiento 58 59 Hegedus 1991 Hinenkel 1992 60 Papini 51 L-T4 – – NS T.Hashimoto hipotiroideas. Posible inclusión de pacientes bocio endémico. Recidiva al suspender TTO en el grupo con L-T4 Nódulos tiroideos Placebo L-T4 – – Bistrop 1994 50 40 100 mg 9 años No cambios 85,5 años Casos operados previamente Pavón de Paz 1996 60 39 No TTo L-T4 12 m 78,2 NS 49 61 * – 100 mg Nódulos tiroideos Referido por Greer 195348 L-T4= Levotiroxina sódica L-T3= Liotironina TD= Tiroides desecado conspira contra su indicación. A nuestro juicio esta modalidad terapéutica debe explotarse cuando las determinaciones de TSH lo justifiquen y en períodos críticos, díganse la pubertad y el embarazo. Todo parece in- 142 dicar que en los pacientes jóvenes, con bocios pequeños y de corto tiempo de evolución, y donde se demuestre su etiología autoinmune, se obtienen los mejores resultados. SUMMARY The diffuse euthyroid goiter (DEG) is up now one of the most controversial topics as to the pathogenesis and treatment of this disease. Thyroid hormones therapy depends on a series of indications that have changed with the years and many specialists doubt the effectiveness of this treatment because of the high rate of relapses after being suspended, as well as the occurence of adverse reactions. The surgical method has very precise indications and its operativeness is questionable. In the present paper, we intend to set forth the main factors involved in the pathogenesis of and the treatment criteria followed in this disease, and also the results that some authors have achieved with the use of both therapies-medical and surgical. Subject headings: GOITER/etiology; GOITER/therapy; THYROID HORMONES/therapeutic use. REFERENCIAS BIBLIOGRÁFICAS 1. Greenspan FS, Rapoport B. Tiroides. En: Greenspan FS, Forsham PH, eds. Endocrinología básica y clínica. México DF: El Manual Moderno, 1988:142-200. 2. Corral J, Martín C, Pérez R, Sánchez I, Mories MT, San Millan JL, et al. Thyroglobulin gen point mutation associated with non-endimic simple goitre. Lancet 1993;341:462-4. 3. Beckers C. Non-toxic goitre. En: De Vischer M, ed. Comprehensive endocrinology. The thyroid gland. New York: Raven Press, 1980:231-55. 4. Miralles JM, Pérez Centeno C, Mories M, Burgos R, González-Sarmiento R. Detección de una mutación reconocida por la endonucleasa de restricción BgIII en el gen de la tiroperoxidasa (TPO) en un paciente con bocio esporádico (abstract). Endocrinología 1996;43:79. 5. Ohyama Y, Hosoya Tk, Kameya T, Susuki N, Nakamura S, Kazahari K, et al. Congenital euthyro8id goitre with impaired thyroglobulin transport. Clin Endocrinol 1994;41:129-35. 6. Ieiri T. thyroglobulin (Tg) gene and familial Tg synthesis defect [abstract]. Nippon Rinsho 1994;52:869-74. 7. de Carvalho DP, Rego KG, Rosenthal D. Thyroid peroxidase in dyshormonogenetic goiters with organification and ghyroglobulin defects. Thyroid 1994;4:421-6. 8. Medeiros Neto G, Targovnik HM, Vassart G. Defective thyroglobulin sinthesis and secretion causing goiter and hypothyroidism. Endocrine Rev 1993;14:165-83. 9. Larsen PR, Ingbar SH. The thyroid gland. En: Wilson JD, Foster DW (eds). Textbook of endocrinology 8 ed. Philadelphia: WB Saunders, 1992:463-5. 10. VanderGaag RD, Drexhage HA, Wiersinga WM. Further studies on thyroid growth-stimulating immunoglobulins in euthyroid nonendemic goiters. J Clin Endocrinol Metabol 1985;60:972-9. 11. Mc Mullan NM, Smyuth PPA. In vitro generation of NADPH as an index of thyroid stimulatin immunolobulins (TSI) in goitrous disease. Clin Endocrinol 1984;20:269-80. 12. Davies R, Lawry J, Bhatia V, Weetman A P. Growth stimulating antibodies in endemic gotre: a reappraisal. Clinical Endocrinol 1995;43:189-95. 13. Zalkarija M, Mc Kenzie JM. Do thyroid growth-promoting inmunoglobulings exist?. J Clin Endocrinol Metabol 1990;70:308-10. 14. Eggo MC, Bachrach LK, Burrow GN. Interaction of TSH, insulin, and insulin like growth factors in regulating thyroid growth and function [bstract]. Growth Factors 1990;2:99-109. 15. Wu J, Dent P, Jeline T, Wolfmann A, Weber MJ, Sturgill TW. Inhibition of EGF-activated MAP kinase signalling pathway by adenosine 3’5' [abstract]. Monophosphate Sc 1993;262:1065-8. 16. De Vito WJ, Chanoine JP, Alex S, Fang SL, Stone S, Huber CA, et al. Effect of in vivo administration of recombinant acidic fibroblast growth factor in thyroid function in the rat: induction of colloid goiter. Endocrinology 1992;131:729-35. 17. Rybakowa M, Soltysik-Wilk E, Tylek D, Krzanowska M, Kowalczyk M. Incidence of goiter in children of the Tamobrzed district and environmental factors [abstract]. Endocrynol Pol 1992;43:12-7. 18. Phillips ID, Becks GP, Logan A, Wang JF, Smith C, Hill DJ. Altered expression of insulin-like growth factorI (IGF-I) and IGF binding proteins during rat thyroid hyperplasia and involution [abstract]. Growth Factors 1994;10:207-22. 19. Tunbridge WMG. The epidemiology of thyroid diseases. En: Ingbar SH, Braverman LE (eds). The thyroid. 5 ed. Philadelphia: Lippincott, 1986:625-33. 143 20. Larsen PR. The thyroid. En: Wyngaarden JB, Smith LLH, Bennett JC, eds. Cecil texbook of medicine. 19 ed. Philadelphia: WB Saunders, 1992:1248-71. 21. Gras C, Hellec C, Spiegel A, Chungue E, Boutin JP, Cardines R, et al. Goiter in Tahiti [abstract]Med Trop Mars 1992;52:57-61. 22. Katugampola SL, Balasuriya S, Fernando MA. Prevalence of goiter in a rural community in Sri Lanka [abstract]. Asia Pac J Public Health 1992;6:182-7. 23. Wolde-Gebril Z, Demeke T, West CE, VanderHaar F. Goitre in Ethiopia. Br J Nutr 1993;69:257-68. 24. Hsiao YL, Chang TC. Prevalence of goiter in Taiwanese adults: a preliminary study [abstract]. J Formos Med Assoc 1995;94:1979. 25. Wong GW. Leung SS. Kwok MM, Openheimer SJ. Goitre in Southern Chinese children in Hong Kong [abstract]. Ann Trop Paediatr 1995;15:27-31. 26. Syrenicz M, Syrenicz A, Gozdzik J, Pynka S, Czekalski S. Frequencyof goiter in children and adolescents in the Szczcecin region [abstract]. Endocrinol Pol 1992;43:38-46. 27. Foley TP. Goiter in adolescents. Endocrinol Metabol Clin North Am 1993;22:593-606. 28. Glinoer D, Lemone M. Goiter and pregnancy: a new insight into an old problem. Thyroid 1992;2:65-70. 29. Castelli V. Hypothyroidism and simple goiter in pregnancy [abstract]. Recenti Prog Med 1994;85:190-5. 30. Burrow GN, Fisher DA, Larsen PR. Maternal and fetal thyroid function. Engl J Med 1994;331:1072-8. 31. Amaro Méndez S. Bocio. En: Breve historia de la endocrinología. La Habana: Editorial Científico Técnica, 1975:67-73. 32. Schaaf L, Trojan J, Helton TE, Usadel KH, Magner JA. Serumn thyrotropin (TSH) heterogeneity in euthyroid subjects and patients with subclinical hypothyroidism: the core fucose content of TSH-releasing hormonereleased TSH is altered, but not the net charge of TSH. J Endocrinol 1995;144:561-7. 33. Toft AD, Irvine WJ, Hunter WM. A comparison of plasma thyrotropin levels in patients with diffuse and nodula non-toxic goiter. J Clin Endocrinol Metabol 1976:42:973-6. 34. Foz M. Tiroides: introducción anatomofisiológica. Bocio Simple. En: Balibrea Cantero JL, Tratado de Cirugía. 1 ed. Barcelona: Ediciones toray, 1989:1364-73. 35. Roti E, Minelli R, Gardini E, Braverman LE. The use and misuse of thyroid hormone. Endocrin Rev 1993;14:401-23. 36. Rendell M, Salmon D. Chemical hyperthyroidism: the significance of elevated serum thyroxine levels in LThyroxine treated indivuduals. Clin Endocrinol 1985;22:693-700. 37. Petersen K, Bengtsson C, Lapidus L. Morbidity, mortality and quality of life for patients treated with L thyroxine. Arch Intern Med 1990;150:2077-81. 38. Kaufman SC, Gross GP. Kennedy DL. Thyroid hormone use: trends in the USA from 1960 through 1988. Thyroid 1991;1:285-91. 39. Hitze G, Emrich D, Kobberling J. Therapy of endemic goitre: controlled wstudy on the effect of iodline and thyroxine. Horm Metabol Res 1985;17:362-5. 40. Hintze G, Emrich D, Kobberling J. Treatment of endemic goiter due toiodine deficiency with iodine, levothyroxine or both results of multicenter trial. Eur J Clin Invest 1989;19:527-34. 41. Cohen JH, Ingbar SH, Bravenman LE. Thyrotoxicosis due to ingestion of excess thyroid hormone. Endocrin Rev 1989;10:113-24. 42. Burch HB. Evaluation and management ofthe solid thyroid nodule. Endocrinol Metabol Clin North Am 1995;24:663-710. 43. Finucane P, Anderson C. Thyroid disease in older pawtients. Diagnosis and treatment. Drugs Aging 1995;6:26877. 44. Sawing CT. Thyroid dysfunction in older persons. Adv Inter Med 1991;37:223-48. 45. Fazio S. Biondi B. Carella C, Amato G, Cittadini A, Lupoli G, et al Diastolic dysfunctionn in patients on thyroid-stimulating hormonc suppressive therapy with levothyroxine: beneficiaaleffect of B-Blockade. J Clin Endocrinol Metabol 1995;80:2222-6. 46. Greenspan SL, Greenspan FS, Resnick NM, Block JE, Friedlander AL, Genant HK. Skeletal integrity in premenopausal and postmenopausal women receiving long-term L-thyroxine therapy. Am J M 1991;91:5-14. 47. Bistrop C, Nielsen JD, Gregersen G, Franch P. Prevetive effect of levothyroxine in patients operated for nontoxic goitre: a randomized trial of one hundred patients with nine years follow up. Clin Endocrinol 1994;40:323-7. 48. Greer MA, Astwood EB. Treatment of simple goiter with thyroid. J Clin Endocrinol Metabol 1953;3:1312-31. 144 49. Badillo J, Shimaoka K, Lessmann EM, Marchetta FC, Sokal JE. Treatment of nontoxic goiter with soidum liothyronine. a double-blind study. JAMA 1963;184:151-8. 50. Gutteridge DH, Orell SR. Non-toxic goitre: Diagnostic role of aspiration cytology, antibodies and serum thyrotropin [abstract]. Clin Endocrinol 1978;9:505-14. 51. Doniach D, Bottazo GF, Russell RCG. Goitrous autoinmune thyroiditis (Hashimoto’s disease). Clin Endocrinol Metabol 1979;8:63-80. 52. Asheraft MW, vanHerle AJ. Managemement of thyroid nodules. II. Scanning techniques, thyroid suppres8ive therapy and fine needle aspiration Head Neck Surg 1981;3:297-322. 53. Perrild H, Hansen JM, Hegedus L, Rytter L, Holm B, Gundtofte E, Hohansen IK. Triiodothyronine and thyroxine treatment of diffuse non-toxic goitre evaluated by ultrasoni scanning. Acta Endocrinol 1982;100:382-7. 54. Olbricht T, Schmitka T, Benker G, Mellinghoff U, Reinwein D. Ultrasonic determination of thyroid volume in non-toxic goitre: treatment with thyroxine versus iodide [abstract] Acta Endocrinol 1984;105:81. 55. Feldt-Rasmussen U, Hegedus L, Hansen JM, Perrild H, Relationship between thyroid volume and serum thyroglobulin during long-term supression with triiodothyronine in patients with diffuse non-toxic goitre. Acta Endocrinol 1984;105:184-9. 56. Berghout A, Wiersinga WM, Drexhage HA, van Trotsenbur P, Smits NJ, vanderGaag RD The long term outcome of thyroidectomy for sporadic non-toxic goitre. Clin Endocrinol 1989;31:193-9. 57. Berghout A, Wiersinga WM, Drexhage HA, Smits NJ, Toubert JL. Comparison of placebo with L. Thyroxine alone or with carbimazole for treatment of sporadic non-toxc goitre. Lancet 1990;336:193-7. 58. Hegedus L, Hansen JM, Feldt-Rasmussen U, Hansen BM, Hoier-Madsen M. Influence of thyroxine treatment on thyroid peroxidase antibodies in Hashimoto thyroiditis. Clin Endocrinol 1991;35:235-8. 59. Einnenkel D, Bauch KH, Benker G. Treatment of juvenile goitre withlevothyroxine, iodide or a combination of both: the valeu of ultrasound grey-scale analysis. Acta Endocrinol 1992; 127: 301-6. 60. Papini E, Bacci V, Panuzi C. A prospective randomized trial of levothyroxine suppresive therapy for solitary thyroid nodules. Clin Endocrinol 1993;38:507-13. 61. Pavón de Paz I, Monereo S Vega B, Vascones S, Elviro R. Tratamiento con levotiroxinna del nódulo tiroideo solitario. Endocrinología 1996;43(1):13-5. Recibido: 8 de mayo de 1998. Aprobado: 19 de julio de 1999. Dr. Mayque Guzmán Cayado. Hospital General "Ciro Redondo", Artemisa, La Habana, Cuba. 145 Rev Cubana Invest Biomed 1999;18(2):146-50 Hospital Universitario General "Calixto García " CARDIOPROTECCIÓN: UN TRIUNFO DE LA BIOMEDICINA DEL SIGLO XX Dra. Iliana Cabrera Rojo RESUMEN La cardioprotección significa prevenir el daño vascular coronario y de los miocitos cardíacos, lo cual se obtiene por mecanismos internos del organismo y a través de diversos fármacos. Esto involucra la vasodilatación, inhibición de la generación de radicales libres de oxígeno, aumento de los niveles de ATP tisular y reducción del daño microvascular. Se revisan los mecanismos de la cardioprotección endógena, metabólica y farmacológica y se destaca la función de los betabloqueadores, anticálcicos, inhibidores de la enzima de conversión de angiotensina y por último la trimetazidina como antiisquémico celular. Descriptores DeCS: ISQUEMIA MIOCARDICA/prevención & control; INSULINA/farmacología; GLUCOSA/farmacología; BLOQUEADORES DE LOS CANALES DE CALCIO/farmacología; BETAANTAGONISTAS ADRENERGICOS/farmacología; INHIBIDORES DE LA ENZIMA CONVERTIDORA DE ANGIOTENSINA/ farmacología; TRIMETACIDINA/ farmacología. METABOLISMO ENERGÉTICO DURANTE LA ISQUEMIA CARDÍACA En condiciones aerobia las 2 vías metabólicas principales del miocito cardíaco son la beta oxidación de los ácidos grasos libres, que aporta 2/3 de ATP, y la oxidación de la glucosa que representa 1/3 del ATP producido en la célula. A causa de que las concentraciones sanguíneas de estos sustratos están relativamente altas, el suplemento de oxígeno es el limitante en la síntesis de ATP. Si el aporte de oxígeno es interrumpido y el balance entre la producción y el 146 consumo de ATP se altera, como sucede en la isquemia, el metabolismo se modifica gradualmente con paso de la respiración aerobia a la glucólisis anaerobia. La concentración de ATP declina y una cascada de fenómenos ocurre: aumento de ADP, AMP, adenosina y pirofosfato, producción de lactato y acidosis; la bomba Na+/K+ se afecta y aumenta la concentración intracelular de Na+, y por consiguiente del calcio, a causa del intercambio Na+/Ca++. El exceso de calcio intracelular interfiere en la capacidad de las mitocondrias de generar ATP. Además, durante la isquemia, disminuye la betaoxidación de los ácidos grasos libres por la inhibición de la enzima trasferasa de acil carnitina, que es necesaria en el transporte de la acil-CoA del citosol a la mitocondria. Sin embargo, hay diferentes mecanismos que protegen al corazón y pueden contrarrestar estos efecto deletéreos. Kubler y Haass1 definieron la cardioprotección como la "preservación del corazón por reducción o prevención del daño miocárdico"; por lo que todos los mecanismos adaptativos y compensatorios que directa o indirectamente contribuyen a la preservación del miocardio han de ser clasificados como "cardioprotectores". A continuación se resumen los mecanismos de cardioprotección: endógeno, metabólico y farmacológico. – Cardiopatía endógena Precondicionamiento isquémico. • "Segunda ventana" de precondicionamiento isquémico. • Hipoxia. • Estimulación vagal. – Cardioprotección metabólica • Infusión de insulina y glucosa – Cardioprotección farmacológica • Anticálcicos. • Betabloqueadores. • Inhibidores de la enzima conversora de angiotensina. • Trimetazidina. CARDIOPROTECCIÓN ENDÓGENA 1. El precondicionamiento isquémico es un fenómeno que se manifiesta después de períodos cortos, de oclusión arterial coronaria seguido de reperfusión, previo a un período prolongado de isquemia, lo que puede proteger al corazón y disminuir el tamaño de la necrosis. Los beneficios se encuentran dentro de un período de 2 h. El mecanismo principal es explicado por la estimulación del receptor A1 de adenosina y activación de la proteína G y la fosfolipasa C, con liberación de sustancias protectoras endógenas, inducción de la sintetasa de óxido nítrico (NO) que resulta en la producción de dicha sustancia y translocación de la proteína kinasa C desde el citosol al sarcolema, lo que favorece la apertura de los canales de K+ dependientes de ATP, se acorta el potencial de acción y hay reducción de la entrada de Ca++.2 Esto conduce a una depresión de la contractilidad, reducción del consumo energético, preservación del ATP y mantenimiento de la función cardíaca. 2. La denominada "segunda ventana" de precondicionamiento isquémico que brinda protección en las primeras 24 h. Lo interesante de este mecanismo es que hay cambios en la expresión genética de sustancias protectoras tales como los heat shock proteins (HSP) o los antioxidantes endógenos, ejemplo: Mn-SOD.3 3. De forma análoga a los 2 mecanismos anteriores la hipoxia por si sola induce cardioprotección. En la clínica, los ejemplos de los mecanismos anteriores resultan el miocardio aturdido y el hibernado, condiciones que se caracterizan por disfunción ventricular izquierda regional causada por la isquemia, pero con reintegración completa de la función contráctil posterior a la recuperación del flujo sanguíneo coronario, ya que los miocitos permanecen viables.4 4. El mediador de la estimulación vagal, la acetilcolina, a través del receptor muscarínico M2 libera NO de las células endoteliales, lo que estimula la gunailciclasa, aumenta el SMPc, hay vasodilatación, apertura de los canales 147 de K+ y depresión de la contractilidad miocárdica y se reduce la demanda de energía. CARDIOPROTECCIÓN METABÓLICA En la isquemia disminuye el contenido de glucógeno y, por tanto, hay reducción del sustrato glicolítico. La infusión de insulina y glucosa, durante la isquemia moderada, aumenta el contenido de ATP celular destinado a la ATP asa de la bomba Na+/K+, por lo que desciende la carga osmolar y mejoran las consecuencias del fenómeno isquemia-reperfusión.5 CARDIOPROTECCIÓN FARMACOLÓGICA La mayoría de las drogas con efectividad en la cardioprotección: anticálcicos, betabloqueadores e inhibidores de la enzima de conversión de angiotensina (IECA), actúan por 2 mecanismos, uno indirecto por efecto hemodinámico, y otro directo por acción tisular específica, en el cual se incluye la trimetazidina. Los anticálcicos actúan de forma específica sobre los canales de calcio voltaje dependiente del músculo liso vascular, que disminuye la corriente de entrada del ion, causando descenso de la resistencia vascular periférica a expensas de las arteriolas de resistencia, y que conlleva a la caída de la presión arterial (PA). Los beta-bloqueadores inhiben el tono simpático al bloquear los receptores beta 2 adrenérgicos presinápticos, inhiben la liberación de renina y aumentan la síntesis vascular de prostaglandina I2 y el NO con propiedades vasodilatadoras y antiagregantes plaquetarias. Además, al reducir la frecuencia cardíaca, la presión de pulso y la velocidad del flujo sanguíneo, 148 conlleva a un efecto protector frente a las fuerzas físicas de presión, cizallamiento y estiramiento.6 Los IECA evitan la formación de angiotensina II, potente vasoconstrictor y activador de la síntesis de aldosterona, lo que impide por una vía la contracción del músculo liso vascular y por otra la retención hidrosalina. Además, con el perindopril, captopril, enalapril y ramipril se ha demostrado un aumento en la producción de bradikinina con la consecuente liberación de NO y prostaciclina.6 Todo esto contribuye al descenso de la PA. Entre los mecanismos directos evaluados en la protección de los miocitos cardíacos y en la regresión de la HVI, se señalan que los betabloqueadores y los IECA interfieren con la acción trófica de la noradrenalina y la angiotensina II respectivamente. También están demostradas fuertes propiedades antioxidantes del propanolol y el carvedilol.6 Por otra parte, en relación con la protección de las arterias coronarias, se conoce que el estrés aumentado de la pared (ejemplo: hipertensión arterial), favorece el transporte de lipoproteínas hacia el interior celular. Todos los fármacos antihipertensivos reducen dicho estrés pero sólo algunos tienen un efecto enlentecedor de la placa de ateroma. Los betabloqueadores y diuréticos tienden a incrementar los lípidos, pero los IECA y anticálcicos no afectan el perfil lipídico. Un hecho a favor de los anticálcicos, es que existe una relación positiva entre el aumento del calcio intracelular y la deposición de lípidos en la lesión aterosclerótica; y un aspecto de interés en relación con los IECA es que el sistema renina angiotensina desempeña un importante papel en el engrosamiento de la capa media de las coronarias y en la fibrosis perivascular, por lo tanto, admi- nistrados a largo plazo en pacientes con cardiopatía isquémica, ejercerán un efecto beneficioso en el remodelado coronario.6-8 Antes de concluir con estos fármacos antihipertensivos hay que mencionar su acción en el síndrome de insulinorresistencia, defecto donde fracasa la respuesta tisular normal a la insulina, y ocurre un hiperinsulinismo. La insulina es aterogénica: altera el transporte de colesterol en la membrana endotelial e incrementa la síntesis endógena de lípidos. Estimula la proliferación del músculo liso arteriolar, aumenta la síntesis de colágenos y factores de crecimiento. Todo lo anterior conduce a hipertensión arterial, diabetes mellitus, obesidad y aceleración de la aterosclerosis. Los diuréticos tiazidas y betabloqueadores están asociados con el desarrollo de la insulinorresistencia y la diabetes, mientras que los calcios antagonistas son neutrales y los IECA aumentan la sensibilidad a la insulina.9 TRIMETAZIDINA Se reporta un efecto protector experimentalmente y en modelos clínicos de isquemia cardíaca: cirugía cardíaca, 10 angioplastia transluminal perscutánea11 e insuficiencia cardíaca.12 En el mecanismo de acción se plantea un efecto directo en los sistemas anzimáticos mitocondriales cuando éstos se deterioran por la isquemia.13 Es un modo de acción diferente al de las drogas antianginosas convencionales, las cuales corrigen el desbalance entre el aporte y la demanda de oxígeno del miocardio a través de un mecanismo de acción hemodinámico. En estudios experimentales, el efecto protector del fármaco fue demostrado en mitocondrias aisladas de corazones hipertrofiados de ratas, sensibles al daño inducido por la isquemia y el estrés oxidativo, donde restauró la fosforilación de ADP, sugestivo de una acción directa sobre la ATPasa mitocondrial.14 Del resultado de investigaciones biomédicas se conoce que las 2 bombas del sarcolema más importantes son las que promueven el transporte de Na+ y Ca++ hacia el exterior celular y que la energía necesaria para mantener su actividad deriva principalmente de la glucólisis, ya que aunque se encuentre al máximo dicha vía metabólica, no se pueden mantener los requerimientos de energía para la contracción cardíaca; la que dependerá de la beta oxidación de los ácidos grasos. Este conocimiento explica la prevención de la acidosis miocárdica con trimetazidina en células isquémicas que se acompaña de reducción de la entrada de sodio a través de la membrana celular, relacionado con la activación de la bomba Na+/H+ y consecuentemente disminuye la acumulación celular de calcio inducida por la bomba de Na+/Ca++. El resultado directo de estas acciones es que los gradientes iónicos a través de la membrana (H+, Ca2+, Na+) se mantienen, con lo cual se garantiza la función contráctil y eléctrica. Concluimos que en la última mitad del sigo XX se han desarrollado múltiples investigaciones biomédicas especialmente en lo concerniente a los mecanismos de protección cardiovascular, lo que condiciona modificaciones en la historia natural de las enfermedades del corazón y los vasos sanguíneos. Además de los mecanismos de cardioprotección endógena y metabólica, los fármacos antihipertensivos (anticálcicos, betabloqueadores, IECA) y los de acción antiisquémica celular directa como la trimetazidina, representan en la 149 actualidad un triunfo de la biomedicina contemporánea, ya que logran reducir, detener y/o revertir los efectos dañinos que tienen diversos factores y moléculas en la fisiopatología de las enfermedades del sistema cardiovascular. SUMMARY Cardioprotection means prevention of vascular coronary damage and of cardiac cyocites, which is achieved by internal mechanism of the body and through various pharmaceuticals. This involves vasodilation, inhibition of oxygen free radical generation, incresase of tissue ATP and reduction of microvascular damage. The mechanisms of endogenous, metabolic and phatmacological cardioprotection are reviewed, and also the function of betablokers, calcium channel blockers, angiotensin-converting enzime inhibitors, and the use of trimetazidine as anti-cell ischemia are underlined. Subject headings: MIOCARDIAL ISCHEMIA/preventive control; INSULIN/pharmacology; GLUCOSE/ pharmacology; CALCIUM CHANNEL BLOCKERS/pharmacology; ADRENERGIC BETA-ANTAGONISTS/ pharmacology; ANGIOTENSIN-CONVERTING ENZYME INHIBITORS/pharmacology; TRIMETAZIDINE/ pharmacology. REFERENCIAS BIBLIOGRÁFICAS 1. Kubler W, Haass M. Cardioprotection: definition, classification and fundamental principles. Heart 1996;75:330-3. 2. Niroomand F. Strasser R. Impaired functions of inhibitory G- proteins. Cir Res 1995;76:861-70. 3. Zarco P, Zarco HM. Biochemical aspects of cardioprotection. Medicographia 1996:18(2):18-21. 4. Heyndrickx GR. Subcellular basis of myocardial stunning and hibernation. Medicographia 1996;18(2):10-2. 5. Apstein CS, Gravino FN, Haudenschild CC. Determinants of a protective effect of glucose and insulin on the ischemic myocardium. Effects on contractile function, diastolic compliance, metabolism and ultrastructure during ischemia and reperfusion. Cir Res 1983;52:515-26. 6. Tamargo J. Análisis de la protección vascular de los farmacos antihipertensivos. En: Dies J ed. Enfermedad vascular e hipertensión arterial. Madrid: Harcourt Brace,1997:331-48. 7. Shim I, Schroeder AP, Aalkjaer C, Holm M, Murn B, Mulvany M, et al. Normalization of structural cardiovascular chamges during antihypertensive treatment with a regimen based on the ACE-inhibitor perindopril. Blood press 1995;4:241-8. 8. Kiowshi w. Efectos coronarios de los inhibidor de la enzima conversiva de la angiotensina (ECA). Inhibición ECA 1993;1(5):66-7. 9. Beiras CA, Muñiz J, Juana R. Contraversias en el tratamiento de la hipertensión arterial: otros fármacos como primer eslabón (calcioantagonistas, IECA y alfabloqueantes). Rev Esp Cardiol 1995;48(Supl 4):72-80. 10. Fabiani JN, Ponzio O, Emerit I. Cardioprotective effect of trimetazidine during coronary artery graft aurgery. J Cardiovasc Surg 1992;33:486-91. 11. Kober B, Buck T, Sievert H, Vallbracht C. Myocardial protection during percutaneous transluminal coronary amgioplasty: effect of trimetazidine. Eur Heart J 1992;13:1109-15. 12. Brottier L, Barat J1, Combe C, Boussens B, Bonnet J,Bricaud H. Therapeutic value of a cardioprotective agent in patiens with severe ischaemic cardiomyopathy. Eur Heart J 1990;11:207-12. 13. Tellier P. Trimetazidine: from cytoprotective effect to anti-ischemic efficacy. Medicographia 1996;18(2):46-50. 14. Mody FW, Sherbel H, Cole K. Mechanisms of action of a novel metabolically active antianginal agent (trimetazidine) delineated by PET. J Am Coll Cardiol 1996;27(Suppl A):132A. Recibido: 3 de junio de 1998. Aprobado: 19 de julio de 1999. Dra Iliana Cabrera Rojo. Hospital Universitario " General Calixto García", Calle J y Ave. Universidad, El Vedado. Ciudad de La Habana, Cuba. 150