XX. Integración metabólica en los mamíferos. Introducción. Aparato
Anuncio

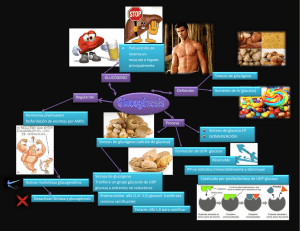
XX. Integración metabólica en los mamíferos. Introducción. Todos los organismos vivos del planeta, dependen absolutamente de la energía y elementos estructurales básicos que se encuentran en el medio ambiente. Debido a los variados requerimientos nutricionales que presenta, el cuerpo de los mamíferos utiliza una elaborada maquinaria para obtener de la ingesta los elementos necesarios para sobrevivir. Es por ello que se lo define como un sistema complejo abierto. La asimilación de los nutrientes por el organismo se conoce como nutrición. Este proceso se lleva a cabo gracias a la participación de una serie de órganos que conforman el aparato digestivo, con la colaboración de otros sistemas, fundamentalmente el aparato circulatorio, que transporta los alimentos entre las diferentes partes del cuerpo una vez que fueron absorbidos, y el aparato respiratorio que facilita el sustrato (oxígeno) y elimina el producto final (dióxido de carbono) del proceso de utilización de los alimentos en las distintas células. Aparato digestivo. En los vertebrados el tubo digestivo (conocido mucho menos elegantemente pero con igual precisión como tripa) está formado por un tubo largo y tortuoso que se extiende desde la boca hasta el ano. Se denomina aparato digestivo al conjunto de órganos encargados de absorber los nutrientes presentes en la alimentación. En términos generales es el encargado de asimilar moléculas y energía del entorno y ponerlas a disposición del organismo. Así, el aparato digestivo incluye también a las glándulas salivales, el páncreas, el hígado y la vesícula biliar, órganos accesorios que proporcionan las enzimas y otras sustancias esenciales para la digestión. La superficie interna del tubo digestivo se continúa con la superficie externa del cuerpo, y así, técnicamente, la cavidad del tubo está fuera del cuerpo. Por ello, los contenidos del tubo, pueden ser sometidos a la acción de enzimas y bacterias y a condiciones de pH que, aunque resultan óptimas para la desintegración del alimento, destruirían rápidamente a las células y tejidos vivos del cuerpo propiamente dicho. Las moléculas nutrientes realmente entran al cuerpo sólo cuando pasan a través del revestimiento epitelial del tubo digestivo y son captadas por la circulación. Así, el proceso de digestión implica dos componentes: la descomposición de las moléculas de alimento y su absorción en el cuerpo. El tubo digestivo comienza en la cavidad oral e incluye la boca, la faringe, el esófago, el estómago, el intestino delgado, el intestino grueso y el ano. Cada una de estas áreas está especializada para una fase particular del proceso general de la digestión, pero la estructura de cada una es similar. El epitelio del tubo digestivo contiene muchas células caliciformes secretoras de moco y, en algunas partes del intestino, glándulas que secretan enzimas digestivas. A lo largo de la mayor parte del tubo digestivo hay dos capas de músculo liso: una capa interna, en la que la orientación es circular, y una capa externa, en la que las células están dispuestas longitudinalmente. Las contracciones coordinadas de estos músculos producen constricciones anulares que mezclan el alimento, al igual que los movimientos ondulatorios, conocidos como peristálticos, que mueven el alimento a lo largo del tubo digestivo. En varios puntos la capa muscular circular se engrosa formando bandas anchas, llamadas esfínteres. Estos esfínteres, relajándose o contrayéndose, actúan como válvulas que controlan el paso del alimento de un área del tubo digestivo a otra. Tubo digestivo o tracto gastrointestinal. Como ya se mencionó, el tubo digestivo está compuesto por órganos huecos que se comunican entre sí dando lugar de esta forma a un tubo cuyos extremos son la boca y el ano. La cavidad buco-nasal y la faringe son estructuras que participan también en el aparato respiratorio. En la boca se realiza principalmente el proceso de ruptura mecánica de los alimentos, también se lleva a cabo una primera digestión enzimática gracias a enzimas presentes en la saliva. Estas enzimas son producidas por las glándulas salivales. La secreción de éstas glándulas es un jugo acuoso con cierta actividad degradativa. Los componentes mayoritarios en la saliva son: agua, bicarbonatos y fosfatos que ayudan a mantener el pH bucal en un valor ligeramente alcalino (entre 7 y 8), Inmunoglubulinas (IgA) con actividad antibacteriana y antifúngica, lisozima, una enzima que degrada la pared bacteriana, amilasa salival, otra enzima que comienza la degradación de polisacáridos. y mucus para lubricar los alimentos que ingresaran al tracto digestivo. La cavidad oral incluye labios, lengua, dientes, paladar y carrillos. La lengua es una estructura con alto contenido de tejido muscular de tipo voluntario en cuya superficie están presentes estructuras denominadas papilas. Las papilas pueden ser táctiles o gustativas. Las papilas filiformes son estructuras pequeñas que presentan terminaciones nerviosas sensibles al tacto, es por ello que son las responsables de la gran sensibilidad táctil de la lengua. Dentro de las papilas gustativas pueden describirse dos tipos: las papilas fungiformes y las papilas caliciformes. Las papilas fungiformes tienen forma de hongo y se encuentran dispersas en toda la lengua. En cambio las papilas caliciformes, de mayor tamaño con forma de copa (o cáliz, de ahí su nombre), se encuentran exclusivamente en la región profunda de la lengua formando la V lingual. Las papilas gustativas son terminales nerviosas modificadas (como las células de la retina, o las células olfativas) que nos permiten diferenciar al menos cuatro sabores básicos, dulce, amargo, ácido, salado. La faringe solo comunica la nariz y la boca con el esófago y la tráquea siendo la glotis la estructura que determina que el pasaje de los alimentos discurra hacia el esófago y no hacia las vías respiratorias. El esófago conecta la faringe con el estómago, no teniendo ninguna actividad digestiva. El estómago es un órgano con forma de saco que continúa al esófago. En él se mezcla el alimento con el jugo gástrico. Todo el alimento ingerido es retenido en el estómago y luego pasa de forma regulada al intestino delgado a través de la válvula pilórica que comunica el estómago con el intestino delgado. El jugo gástrico es un líquido súmamente ácido que es producido por las células epiteliales del estómago que expulsan activamente iones H+ , los que son acompañados por Cl—, hacia la luz de la cavidad con gasto de energía. De hecho, este gasto de energía realizado en la formación del ácido estomacal es uno de los procesos importantes que contribuyen al cálculo total de gasto energético diario en un individuo adulto. El pH del jugo gástrico tiene un valor prácticamente igual a 2, lo que lo convierte en único en comparación con los otros líquidos celulares que o son neutros o ligeramente alcalinos. Las células de la pared intestinal se encuentran recubiertas por una capa de moco muy gruesa para no verse afectadas por la acidéz del jugo que ellas mismas generan. El intestino delgado es el órgano que presenta la mayor longitud de todo el tubo digestivo, (alrededor de 2,5 mts.). En él se realiza la ruptura enzimática de las macromoléculas y la absorción de los nutrientes liberados de la ingesta. Presenta prolongaciones en su superficie que son denominadas vellosidades intestinales las cuales aumentan enormemente la superficie de absorción del intestino –esta superficie de absorción en un adulto ronda los 300 mts 2 similar a la superficie que presenta una cancha de tenis–. El intestino delgado puede dividirse en tres zonas: el duodeno que es la región que está en contacto con el estómago a través de la válvula pilórica, el yeyuno y el ileon. Estas regiones se diferencian unas de otras por la presencia o ausencia de ciertas estructuras celulares. Gracias a la estructura muscular y nerviosa que presenta, este órgano tiene una conspicua actividad contráctil zonal lo que le permite desplazar el bolo alimenticio de forma anteroposterior, movimiento que se conoce como peristálsis. Las enzimas digestivas producidas por el páncreas y las sustancias de solubilización de los alimentos grasos producidas por la vesícula biliar, son volcadas primariamente en el duodeno. El páncreas también vuelca en el duodeno un líquido muy alcalino, tanto que es capaz de neutralizar la acidéz del jugo gástrico con el que está en contacto a través del píloro y deja en el duodeno un pH alcalino, que tiene un valor aproximado de 8. Las principales enzimas del jugo pancreático sirven para digerir los cuatro tipos de macromoléculas y liberar, a partir de ellas sus monómeros constituyentes o, en algunos casos, oligómeros pequeños. Por ejemplo, la tripsina, la quimiotripsina, y las carboxipeptidasas A y B, son proteasas que degradan las proteínas y liberan aminoácidos libres (a veces dipéptidos); la amilasa pancreática termina de degradar los polisacáridos que no fueron todavía procesados, y libera azúcares simples; la lipasa y la fosfolipasa degradan los acil-glicéridos liberando sus ácidos grasos constituyentes, y la desoxirribonucleasa y la ribonucleasa degradan al DNA y al RNA liberando los nucleótidos. Así, los monómeros pueden ser absorbidos en el resto del intestino delgado sin problemas. El intestino grueso continua al ileon (la última porción del intestino delgado) y se comunica con éste a través de la válvula iliocecal. Mide alrededor de 1,5 mts. de longitud. Tiene poca o nula capacidad de digestión, aunque si presenta actividad de absorción, principalmente de líquidos. Se divide en ciego, colon, recto para terminar en el canal anal. El hígado es la principal glándula anexa del organismo. La secreción exógena del hígado, se acumula, antes de llegar al duodeno, en la vesícula biliar. Esta sustancia, denominada bilis, es un líquido amarillento y alcalino que contiene bicarbonato de sodio, sales biliares, pigmentos biliares, colesterol, lecitinas, mucinas y bilirrubina. En la secreción biliar se mezclan principios activos, como son las sales biliares que disuelven los componentes grasos de la dieta, y productos de desechos, como son los pigmentos biliares y la bilirrubina. La función principal de la bilis es permitir la absorción de las grasas, lo cual se logra por la actividad emulsificante de las sales biliares. El hígado es el órgano que sensa, almacena y distribuye los componentes alimentarios para la dieta de todas las células del cuerpo. Además, como ya se mencionó, participa activamente en la digestión de alimentos a través de su secreción. Alimentac ión y nutrición. Los alimentos ingeridos en la dieta contienen nutrientes que son los elementos aprovechados por el organismo. Los nutrientes proveen energía y componentes básicos estructurales. En la comida están presente seis clases de nutrientes esenciales que son vitales para mantener en condiciones óptimas al cuerpo. Ellos son: carbohidratos, proteínas, lípidos, vitaminas, minerales y agua. Estos nutrientes no están presentes en todos los alimentos por lo que mantener una dieta variada evita que se produzcan deficiencias alimenticias. Los componentes de los alimentos son estructuralmente similares a los del propio organismo, por lo cual la actividad degradativa –que tiene como fin atacar al alimento– podría potencialmente atacar a distintas estructuras propias. Esto no ocurre normalmente gracias a que la digestión se da en la luz del tubo digestivo (o sea en el exterior del cuerpo), que presenta barreras para evitar la autodigestión. Los mamíferos requieren, ya que no pueden sintetizar, un grupo grande de aminoácidos, vitaminas, y ciertas grasas poliinsaturadas. Los aminoácidos y las proteínas (y los aminoácidos escenciales) normalmente se encuentran en una dieta variada que contenga frutas, vegetales y carnes. Los ácidos grasos esenciales pueden obtenerse comiendo plantas o insectos (o comiendo otros animales que se han alimentado de plantas o de insectos, lo que a veces es mas sabroso). Los excesos en el consumo de alimentos pueden llevar a la obesidad y se correlacionan con un inrecremento significativo en la enfermedad coronaria, la diabetes y otros trastornos. El exceso de sal ha sido relacionado con la hipertensión. Otro riesgo es la grasa animal, como la que se encuentra presente en la carne vacuna y de cerdo. Se sabe, desde hace cierto tiempo, que las dietas ricas en grasa animal interfieren en la regulación del colesterol sanguíneo, implicado en la aterosclerosis y en los ataques cardíacos. Más recientemente se ha enfocado la atención en el papel de las fibras de la dieta (carbohidratos no digeribles tipo la celulosa para nosotros) en la disminución del cáncer de colon. Otro riesgo nutricional, es el deseo de experimentar con nuestros propios cuerpos y adoptar dietas extremas y seguir las modas nutricionales: el ayuno, las proteínas líquidas, la “macrobiótica”, las dosis proteínicas elevadas o bajas, la alta cantidad de fibras, las megavitaminas, sin considerar nuestros requerimientos como organismos biológicos y los dictados de nuestra larga historia evolutiva omnívora. Otros auxiliares de la digestión. Comparadas con las células de otros tejidos, el material vegetal es duro y difícil de digerir. Por lógica, se esperaría que los hervívoros terrestres tuviesen conjuntos de enzimas capaces de desintegrar la celulosa y otros polisacáridos estructurales que se encuentran en las plantas. Sin embargo no es así. Los animales terrestres han elaborado otra solución: relaciones simbióticas con bacterias y protistas que desempeñan estas actividades en su beneficio y reciben en retribución un abastecimiento alimenticio gratuito y protección. En el interior cálido, húmedo, sin aire, del tracto digestivo de un mamífero, los simbiontes degradan los productos vegetales de manera anaerobia. La glucosa la utilizan ellos mismos; la parte que le queda al huésped del alimento está formada principalmente por los ácidos grasos, y en parte por algunos aminoácidos que éste absorbe a través de su intestino. Además, los microorganismos sintetizan muchas vitaminas, especialmente la vitamina K y las del grupo B, que son utilizadas por el animal. El proceso de fermentación, la oxidación anaerobia de moléculas orgánicas, ocurre en diferentes áreas del tubo intestinal de diferentes especies. En la familia del caballo ocurre en el colon, que es grande, y en el ciego, un divertículo lateral que se encuentra en la unión de los intestinos delgado y grueso. El ciego persiste en los seres humanos en forma vestigial como el apéndice, aparentemente sin una función importante pero causante de problemas agudos. Por otro lado, los conejos y otros lagomorfos tienen un ciego agrandado, en el que ocurre la fermentación bacteriana y también una adaptación curiosa, peculiar de este grupo. Durante la noche, en sus cuevas, los conejos producen desde el ciego un tipo especial de heces, diferentes de las que producen en las horas diurnas, que consisten casi completamente en bacterias. Los conejos comen estas heces, digiriendo y absorbiendo así nutrientes adicionales, obteniendo vitaminas producidas por las bacterias y recirculando sus simbiontes beneficiosos. La especialización más compleja, anatómicamente hablando, se encuentra entre los rumiantes, tales como los vacunos, los antílopes, las ovejas y los venados. Probablemente no sea una coincidencia que los rumiantes se encuentren entre los herbívoros terrestres más exitosos, cuya elevación a la prominencia evolutiva tuvo lugar cuando las praderas graminosas estaban experimentando una rápida expansión. Los rumiantes desgarran precipitadamente las hojas de gramíneas y otras partes vegetales y las almacenan, regurgitándolas en forma de bolo alimenticio y completando el proceso de masticación sin prisa. La fermentación real ocurre en el rumen, un saco esofágico grande. El contenido del rumen pasa luego al estómago, donde comienza la absorción. Las bacterias y los ciliados del rumen no solamente degradan la celulosa y otros polisacáridos, sino que además sintetizan proteínas, utilizando amoníaco y urea como fuentes de nitrógeno. Algunos de estos microorganismos también son digeridos por el hospedador, proveyéndole así de proteínas. Los omnívoros humanos no tienen simbiontes de este tipo y, por tanto, son incapaces de utilizar la celulosa, que es eliminada como sustancia indigesta. Sin embargo, al igual que otros mamíferos, dependemos de las bacterias de nuestro tubo digestivo para sintetizar vitaminas y, como en el conejo, las bacterias simbióticas son un componente principal de nuestras heces, aunque por suerte no tendamos a alimentarnos de ellas. Carbohidratos. Con la excepción de la lactosa, presente en la leche, de la melasa producida por insectos o del glucógeno del hígado y músculos de los animales, los carbohidratos se asimilan por ingestión de productos elaborados por las plantas. Las frutas en general son ricas en glucosa, fructosa y sacarosa, mientras que los tubérculos, los granos y las legumbres aportan mayoritariamente almidón. La celulosa es un polisacárido rico en glucosa que debido a su estructura molecular (las uniones entre las glucosas son a través de enlaces beta en lugar de enlaces alfa como en el caso de el almidón y el glucógeno) es indigerible para el ser humano. A estos constituyentes indigeribles de los alimentos se los conoce como fibras. Los alimentos que presentan altas cantidades de fibra no tienen aparentemente propiedades nutricionales. Sin embargo, una dieta carente de fibras puede ocacionar graves trastornos intestinales ya que su ingesta es escencial para facilitar el tránsito fecal y la peristalsis. Proteínas. Normalmente, son los componentes mayoritarios en la ingesta. Algunos aminoácidos pueden degradarse con el fin de producir energía. De los 20 aminoácidos diferentes que conforman los distintos tipos de proteínas, 12 son producidos por el organismo, el resto debe ser adquirido en la dieta. Este hecho determina las series de aminoácidos esenciales y no esenciales, o sea aquellos que deben ser asimilados por la dieta necesariamente y aquellos cuya ingesta no es absolutamente necesaria. Las fuentes animales de proteínas incluyen carnes rojas y blancas, lácteos, huevos, etc. Las fuentes vegetales que contienen altas cantidades de proteínas incluyen maníes, cereales, frutas secas, etc. Aunque las proteínas son parte vital de la dieta, los requerimientos diarios de proteínas son relativamente bajos –en el adulto se sitúan alrededor de 0,8 gr por kilo de peso–. Lípidos. Los lípidos son las moléculas con mayor carga energética de la dieta. Se encuentran principalmente en las mantecas, los aceites y las margarinas. También están presentes en la crema y el chocolate, y por supuesto en la grasa animal. Los lípidos tienen varias funciones. Además de ser moléculas con una gran carga energética, algunos lípidos facilitan la absorción de vitaminas hidrofóbicas, como las vitaminas A, D, E y K. Los lípidos mantienen la salud de la piel y permiten un saludable crecimiento. El colesterol –precursor de la vitamina D- y los fosfolípidos son constituyentes de las membranas celulares. Vitaminas. Las vitaminas son pequeñas moléculas orgánicas que se requieren en poca cantidad para el funcionamiento del metabolismo. Muchas vitaminas son convertidas en coenzimas después de entrar en el organismo. Como las vitaminas no pueden ser sintetizadas por el organismo se hace necesario el consumo de éstas moléculas en la dieta. Sin embargo, la vitamina K, es sintetizada por las bacterias presentes en el intestino grueso y absorbida por el organismo para su uso. Pero esta síntesis se lleva a cabo sólo en presencia de ciertos precursores (pro-vitaminas) que se convierten así en elementos tan importantes en la ingesta como las vitaminas. Minerales. Los principales minerales necesarios en la dieta son el sodio, el calcio y el potasio que participan en la regulación homeostática del organismo. Otros minerales como el iodo, el cobre, el zinc, el hierro y el cobalto se clasifican dentro de los grupos de necesidad minoritaria. Este concepto no implica que no sean importantes en la dieta sino que, al existir como trazas en varios alimentos, son ingeridos naturalmente. Agua. Ningún nutriente es más importante que el agua. Más del 60% del volumen corporal es de agua. El agua participa en todos los procesos que se realizan dentro del organismo y es un elemento fundamental para la existencia de la vida. Energía La energía presente en los alimentos es medida en calorías. Una caloría es la cantidad de energía necesaria para elevar la temperatura de 1 gramo de agua en 1°C. La energía tiene dos costados uno está directamente relacionado con la nutrición, y es el consumo energético (CE). El otro está solo indirectamente relacionado y es lo que se denomina el gasto energético (GE), que corresponde a la suma de actividades del organismo. En un individuo sano el gasto debe ser aproximadamente igual al consumo. GE ≈ CE Cada uno de los constituyentes de los alimentos que mencionamos, excepto los minerales el agua y (porque se encuentran en cantidades mínimas) las vitaminas, aportan al contenido energético de un alimento. El contenido energético de un alimento puede calcularse de acuerdo a la composición relativa de las distintos grupos de moléculas, carbohidratos, proteínas, lípidos y, en menor medida, ácidos nucleicos. El potencial energético de los carbohidratos, de las proteínas y de los ácidos nucleicos es similar, situándose alrededor de 4 kcal por gramo de materia metabolizada. En cambio, los lípidos son las moléculas con mayor carga energética de la dieta ya que por gramo de lípido el organismo aprovecha 9 kcal. Un componente que también hay que considerar (ya que es usualmente ingerido acompañando las dietas aunque no sea ”esencial“), es el alcohol, que aporta aproximadamente 7 kcal por gramo, bastante más que las proteínas e hidratos de carbono. Como dijimos, para mantener la salud el gasto energético debe aproximarse al consumo. Si el gasto es siempre mayor que el consumo, se pueden producir enfermedades crónicas como la anorexia. En el caso contrario, se puede producir obesidad. Obviamente, la igualdad exacta no se cumple casi nunca y los individuos adelgazan o engordan durante determinados períodos sin que esto implique una patología. Los componentes principales del gasto energético de un cuerpo son, en orden de importancia: 1 Metabolismo Basal, corresponde a todas las actividades que el cuerpo realiza aún cuando está en reposo absoluto, se incluyen aquí, al mantenimiento de la actividad de los músculos involuntarios (como corazón o diafragma), a la regulación de la temperatura corporal y al mantenimiento del balance ionico en cada una de las células del cuerpo. Algunos autores incluyen aquí la energía gastada en mantener la temperatura corporal o termoregulación. Otros la consideran aparte ya que a diferencia del resto de los componentes del metabolismo basal, la termoregulación depende de la temperatura externa. 2 Acción dinámica, que corresponde a la energía gastada por el cuerpo para digerir un alimento. Es el gasto de energía asociado con la producción del jugo gástrico y con la síntesis de las enzimas pancreáticas. 3 Trabajo, corresponde al gasto energético producto del movimiento de los músculos voluntarios. 4 Crecimiento, esta energía se considera sólo en los niños y en las mujeres embarazadas o en lactancia. Con respecto a las dietas, un punto a tener en cuenta es que cuando estas se basan en el aumento del gasto energético para desbalancear la ecuación mostrada más arriba y producir así una disminución del peso, el punto 3, es el único ítem que podemos manejar y, para colmo, no es el que más aporta. Por ello, cuando nos embarcamos en una dieta de este tipo se requiere un esfuerzo continuado durante mucho tiempo para poder lograr el efecto buscado. Asimilación de los componentes de la dieta. Los componentes presentes en la dieta son absorbidos, como ya se mencionó, en el intestino delgado. La mayoría de los nutrientes son recogidos por el sistema sanguíneo. Las proteínas degradadas son absorbidas como tripéptidos, dipéptidos o aminoácidos libres. En la célula intestinal los di y tripéptidos son degradados a aminoácidos y así pasan a la circulación. Los azúcares son absorbidos por las células como hexosas y los derivados de polinucleótidos como bases y pentosas libres. Los monosacáridos y las bases pasan como los aminoácidos a la circulación sanguínea. Los capilares sanguíneos del intestino, por su parte se unen entre sí y en lugar de conectarse directamente con la circulación general, lo hacen con el hígado que es el órgano que almacena y distribuye los nutrientes. Así esta vena ingresa al hígado en donde se recapilariza para ponerse en contacto con cada una de las células hepáticas. Esta estructura anatómica en donde una red capilar forma un vaso sanguíneo que vuelve a ramificarse en otro órgano se denomina sistema porta. Es por ello que la estructura que une al intestino delgado con el hígado se conoce como sistema porta hepático, conformado por la red capilar mesentérica del intestino, la vena porta hepática y la red capilar hepática. Como se deduce de lo dicho anteriormente el órgano que recibe casi todos los elementos absorbidos en el intestino es el hígado. Este hecho explica la función detoxificadora de éste órgano, porque así como recibe todos los nutrientes también recibe las toxinas que puedan haberse ingerido o aquellas que naturalmente presentan los alimentos. Además, el hígado tiene una marcada actividad metabólica de obtención y almacenamiento de energía y de nutrientes. Por otra parte las grasas de la dieta no son transferidas al sistema porta hepático. En cambio, los ácidos grasos libres, o los monoacilgliceroles, que son el producto de la degradación de las grasas en la luz intestinal, son absorbidos por las células intestinales, y dentro de ellas son re-esterificados para formas triacilgliceroles. Estos, en conjunto con el colesterol y otros lípidos de la dieta, son incorporados en la pared interna de la célula intestinal a partículas que contienen proteínas y se denominan Quilomicrones. Los quilomicrones por su gran tamaño no pueden ser incorporados por los capilares sanguíneos y por ello, son recuperados por el sistema linfático. Los capilares del sistema linfático, recogen los quilomicrones como también diferentes partículas del líquido instersticial. Estos capilares terminan asociándose para convertirse en las vías de circulación linfática centrales, que, pasando por los ganglios terminan volcando su contenido en la vena cava superior, justo antes de la entrada de la sangre venosa en el corazón. Desde aquí, las grasas son distribuidas por la circulación arterial a todo el cuerpo, son absorbidas por prácticamente todos los tipos celulares que las usan como fuente principal de energía. Si existe un exceso de grasa dietaria, terminan siendo almacenadas en las células adiposas del organismo, aumentando el volumen de este tejido (que lamentablemente no tiene límite de crecimiento). Homeostasis de la glucos a sanguínea. La principal función de la digestión es, naturalmente, suministrarle a cada célula del cuerpo las moléculas orgánicas que puedan servir como fuente de energía y materias primas. Aunque los vertebrados raramente comen durante las 24 horas del día, la concentración de la glucosa en la sangre —que es la fuente de la energía casi exclusiva de las neuronas, de los glóbulos rojos y de las células de la retina, y además es una molécula estructural fundamental— permanece extraordinariamente constante. Nuevamente el hígado desempeña un papel central en este proceso crítico. Como dijimos, la glucosa y otros monosacáridos entran en la sangre por absorción desde el tubo intestinal y pasan directamente al hígado por intermedio de la vena porta hepática. El hígado convierte parte de estos monosacáridos en glucógeno de reserva. En esta etapa, los azúcares son absorbidos además por otros órganos, como el músculo donde también se convierten en glucógeno, o por los adipocitos, donde se utilizan para sintetizar los restos de glicerol que van a formar parte de las grasas acumuladas como triacilgliceroles. El exceso de azúcares en la sangre es convertido, fundamentalmente por el hígado, en ácidos grasos, los que terminan siendo almacenados en las células grasas, y como dijimos antes, acumulándose indefinidamente. De modo análogo, el hígado degrada los aminoácidos en exceso (que no se almacenan) y los convierte en glucosa. El nitrógeno de los aminoácidos es excretado en forma de urea y la glucosa es almacenada como glucógeno o, también cuando está en exceso, convertida en ácidos grasos. Para identificar todos los procesos posibles que ocurren para mantener la homeostasis de la glucosa en la sangre, deberíamos considerar que adaptaciones metabólicas ocurren en el organismo después de una comida hasta llegar a un ayuno prolongado. En este caso, tomando en cuenta los perfiles metabólicos diferentes de un grupo de órganos clave, podemos identificar al menos 5 fases diferentes de la homeostasis de la glucosa. La primera de ellas es la fase que ocurre inmediatamente después de una comida, perdura entre 2 y 4 hs. En esta fase se produce la absorción y el almacenamiento de los componentes de la dieta y por ello es denominada fase absortiva. La fase siguiente, o postabsortiva esta caracterizada por la utilización de los compuestos almacenados en la primer fase para mantener la homeostasis. Esta fase dura como máximo hasta 24 hs. después de la comida. Estas son las fases que normalmente atraviesa cualquier organismo. De no mediar una comida antes que termine la fase postabsortiva, el organismo entra en las fases de ayuno. Estas fases son tres, ayuno primario, secundario y terciario y se diferencian en los metabolitos que el cuerpo utiliza para mantener la homeostasis. Fase absortiva. Esta fase está caracterizada por un aumento en la concentración de los metabolitos alimenticios en la sangre. Por lo mismo se estimula su captación por los tejidos. Vamos a considerar lo que ocurre principalmente con la glucosa, los ácidos grasos y los aminoácidos. La concentración de glucosa y la de ácidos grasos, está regulada por diversas hormonas y está influida por el sistema nervioso autónomo. Entre las hormonas que intervienen en este proceso están la insulina, el glucagón y la somatostatina, todas ellas producidas por el páncreas. La insulina actúa sobre varios órganos, y estimula la absorción de glucosa por las células, disminuyendo la glucosa sanguínea. También estimula la captación de ácidos grasos y la síntesis de triacilgliceroles en los adipocitos. Por el contrario, el glucagón actúa sobre el hígado exclusivamente. Allí promueve la degradación del glucógeno y la desfosforilación de la glucosa-6-fosfato, incrementando así la concentración de glucosa sanguínea. La insulina indica abundancia de alimento mientras que el glucagón indica hambre. Por su parte, la somatostatina, que fue la primera proteína de los vertebrados sintetizada usando tecnología de DNA recombinante, tiene una variedad de efectos inhibitorios que colectivamente ayudan a regular la tasa a la que la glucosa y otros nutrientes son absorbidos desde el tubo digestivo. En esta fase se produce un aumento en la liberación de la insulina y una disminución en la concentración del glucagón en la sangre, las dos principales hormonas reguladoras de la concentración de glucosa sanguínea y de la captación o liberación de ácidos grasos. Las células de distintos órganos modifican su actividad de diferente forma para aumentar su capacidad de recuperar glucosa de la sangre en este período. Casi todas estas modificaciones son impulsadas por la insulina que actúa en forma diferente sobre distintas células. Sobre los hepatocitos la insulina activa a la hexokinasa IV (antes denominada glucokinasa) e inhibe a la glucosa-6-fosfatasa. La hexokinasa IV es la isoforma de las hexokinasas que posee una mayor velocidad máxima y por lo mismo tiene una muy alta capacidad de fosforilar glucosa. Al mismo tiempo posee un KM también elevado, por lo que sólo actúa cuando la concentración de glucosa dentro de la célula supera determinado valor. En el caso de esta enzima el valor límite mencionado, en el que comienza a ser importante su actividad, corresponde a aproximadamente 8 mM que está por encima del valor promedio de la concentración de la glucosa sanguínea (5 mM). Las células del hígado poseen transportadores pasivos de glucosa que funcionan en los dos sentidos dependiendo del gradiente de glucosa generado. La actividad de la hexokinasa IV, aparece sólo cuando la glucosa en sangre se encuentra en exceso y por su acción disminuye la concentración de glucosa interna, tornándola inferior a la concentración de glucosa sanguínea. De esta manera se produce un desplazamiento del equilibrio alrededor del transportador de glucosa de la célula hepática que lo hace funcionar captando glucosa de la sangre. Esta glucosa es convertida primariamente en glucógeno. Como el glucógeno tiene un límite en su acumulación, el resto de la glucosa no convertida en glucógeno se utiliza para sintetizar acetil-CoA. El acetil-CoA puede ser utilizado para generar la energía necesaria para el funcionamiento de la célula hepática y el exceso se convierte en ácidos grasos, que son liberados a circulación como lipoproteínas. Otros tipos de células como las células musculares o los adipocitos tienen un mecanismo diferente para aumentar la capacidad de absorción de glucosa en la fase absortiva. Estas células prácticamente no captan glucosa en otras fases homeostáticas diferentes a la fase absortiva. Esto se debe a que no poseen transportadores de membrana para la glucosa, expuestos en la superficie de las células. Por el contrario, los transportadores de glucosa de estas células se encuentran ubicados en vesículas en el interior del citoplasma por lo que no cumplen ninguna función. La insulina al impactar sobre estas células, impulsa la fusión de estas vesículas con la membrana plasmática, lo que provoca la externalización de los receptores para glucosa. De esta manera, estos órganos son capaces de captar activamente glucosa solamente cuando la insulina está presente en la sangre, lo que ocurre, normalmente, inmediatamente después de una comida en la fase absortiva. La glucosa que ingresa al músculo, principalmente, se transforma en glucógeno y en parte aporta la energía necesaria para el funcionamiento de la célula muscular. Considerado como un conjunto, la mayor parte del glucógeno almacenado en un organismo, el 85% aproximadamente, se encuentra en los músculos; el resto mayoritariamente es el glucógeno hepático. Sin embargo, el glucógeno muscular sólo puede ser utilizado dentro de la célula donde está acumulado. Esto lo diferencia claramente del glucógeno hepático que puede ser exportado a la sangre. Las células musculares no pueden exportar la glucosa acumulada en la forma de glucógeno ya que no poseen la enzima glucosa-6-fosfatasa, que si es activa en el hígado. El otro órgano que posee esta enzima es el riñón. La glucosa que no es captada ni por el hígado ni por los otros órganos, llega al riñón donde es filtrada (al mismo tiempo que todas las moléculas pequeñas también lo son) e, inicialmente, es transportada hacia los túbulos renales, donde forma parte del líquido que después va a transformarse en orina. Las células que recubren los túbulos renales poseen transportadores activos, que, asociados con el gasto de ATP, reabsorben la glucosa presente en los túbulos renales. Este mecanismo de reabsorción diferencia a la glucosa (y a un conjunto pequeño de otras moléculas) de otras moléculas pequeñas que no pueden ser recuperadas y por lo mismo se pierden en la orina. La glucosa reabsorbida vuelve a la sangre circulante gracias a la acción de la enzima glucosa-6-fosfatasa renal. Los adipocitos, tienen el mismo tipo de transportadores de glucosa (que se exponen en respuesta a la insulina) que las células musculares. Sin embargo el destino de la glucosa absorbida durante la fase absortiva en este órgano, es diferente, ya que su principal utilidad es en la síntesis del esqueleto del glicerol que va a formar parte de los triacilglicéridos. En los adipocitos durante esta fase también se absorben activamente ácidos grasos aportados por los quilomicrones del plasma. Los triglicéridos transportados por cualquiera de las lipoproteínas plasmáticas1, llegan a todos los tejidos. Antes de ser absorbidos, son degradados a glicerol y ácidos grasos por la acción de una lipoproteín-lipasa externa, que está unida a la capa externa de la membrana plasmática por medio de un puente de oligosacáridos. Estas lipasas son activas siempre, independientemente del nivel hormonal y de la fase alimentaria que se analice. Además como dijimos estas lipasas se encuentran en la superficie externa de la mayoría de los tipos celulares ya que permiten la incorporación del alimento energético principal de la mayoría de las células que son los ácidos grasos. Una vez que los ácidos grasos son incorporados a la célula adiposa, tienen que ser reesterificados con el glicerol, para formar los triacil-gliceroles que conforman las mencionadas gotas de grasa intracelulares. Este proceso de esterificación comienza con la reacción entre un acil graso-CoA y el glicerol-3-fosfato, para producir un monoacil-fosfatidil-glicerol. Este compuesto reacciona dos veces más con derivados de CoA de otros ácidos grasos (la última reacción con liberación del fosfato del glicerol-fosfato), para formar los triacil-gliceroles. Una característica particular de los adipocitos es que el glicerol que se obtiene de la degradación de los triacil-gliceroles de los alimentos en el exterior celular, no puede ser utilizado para la reesterificación de los ácidos grasos en el interior de la célula. Esto es debido a que los adipocitos carecen de la enzima glicerol-kinasa, necesaria para convertir el glicerol libre en glicerol-3-fosfato que es el primer sustrato del proceso de esterificación. Aquí es donde entra a jugar la captación de glucosa externa (impulsada por la insulina, por exposición de los receptores) en los adipocitos. El glicerol-3-fosfato solo puede ser obtenido en los adipocitos a partir de la glucosa captada, y por lo mismo, la síntesis de triacilgliceroles en estas células depende exclusivamente del aporte externo de glucosa. 1 El quilomicrón es uno de los distintos tipos de lipoproteínas plasmáticas. Los quilomicrones son más grandes que otras lipoproteínas y se diferencian de ellas en que son sintetizadas en las células intestinales y transporta lípidos dietarios. Las otras lipoproteínas (HDL, LDL, IDL y VLDL) son sintetizadas en el hígado y transportan lípidos generados en el propio organismo. Para ello, la glucosa, se convierte en dihidroxiacetona-fosfato y la reducción de esta produce el glicerol-3-fosfato. El glicerol liberado por la degradación de los triacilgliceroles externos (o internos en las siguientes fases), es transportado por la sangre al hígado donde sí existe la enzima glicerol-kinasa. Por lo mismo en el hígado, el glicerol puede ser convertido en glucosa, por medio de las mismas reacciones que ocurren en el adipocito pero en sentido inverso. Los adipocitos poseen, además, otra lipasa, que es una enzima interna y que se utiliza para degradar las gotas de grasa interna de estas células. Estas gotas de grasa que están formadas por triacilgliceroles, son la reserva de energía más importante del cuerpo y su degradación se utiliza en las etapas posteriores a la fase absortiva para generar los ácidos grasos que puedan llevar energía metabólica al resto del cuerpo. La enzima que degrada estos triacilglicéridos a glicerol y ácidos grasos, en el interior del adipocito, es una lipasa cuya actividad se encuentra inhibida cuando hay insulina en la sangre. De esta manera la insulina regula el metabolismo de las grasas en las células adiposas. En la fase absortiva, al permitir el ingreso de glucosa, permite la síntesis del glicerol-3-fosfato que es esencial para la síntesis de los triacilglicéridos. Además en esta fase inhibe a las lipasas internas evitando la degradación de las grasas, con lo que el flujo metabólico en la fase absortiva tiende a la acumulación de grasas. Por el contrario en las fases siguientes, a causa de la ausencia de insulina, no hay suficientes receptores de glucosa en las células adiposas y además se elimina la inhibición de las lipasas internas con lo que el flujo metabólico en las demás fases tiende a la eliminación de grasas. Además, la oxidación de los ácidos grasos por otros tejidos es regulada en los sitios de liberación de los mismos en el adipocito. La liberación de los ácidos grasos de las células de grasa por medio de la degradación de los triacilglicéridos es estimulada sobre todo por la epinefrina. Como resultado, se incrementa la concentración de ácidos grasos fijados a la albúmina en la sangre ocasionando que una mayor cantidad de ácidos grasos penetre en los tejidos donde son oxidados. La epinefrina provoca el aumento en la concentración de AMPcíclico en la célula adiposa, lo que luego causa el efecto de aumentar la liberación de ácidos grasos en circulación. Uno de los efectos de la cafeína en el organismo es justamente inhibir la acción de la enzima fosfodiesterasa, que actúa degradando el AMPcíclico y terminando con la acción de la epinefrina. Las proteínas absorbidas en la dieta, son degradadas a aminoácidos en las células de la pared intestinal antes de ser volcadas a la vena porta que los transporta hacia el hígado. A diferencia de las otras moléculas que consideramos hasta ahora (los glúcidos y los lípidos) los aminoácidos no son almacenados en ninguna célula. Algunos aminoácidos no esenciales son rápidamente metabolizados, en el intestino mismo (cuya principal fuente de energía es la glutamina, en todas las fases alimenticias) o en el hígado (cuya principal fuente de energía en la fase absortiva son también los aminoácidos). La degradación de la glutamina en el intestino, provoca la liberación de amonio a la vena porta que llega al hígado, así como el metabolismo de otros aminoácidos en el hígado, produce también la liberación de amonio en la célula hepática. El amonio es en definitiva, metabolizado en el hígado por el ciclo de la urea para poder ser eliminado sin causar toxicidad al resto del cuerpo. Aproximadamente la mitad de los aminoácidos dietarios corresponden a aminoácidos esenciales que no son metabolizados ni en el intestino ni en el hígado y pasan a la circulación general para ser utilizados en distintos tejidos para la síntesis de las proteínas corporales. 16 14 12 10 8 Gl uconeogénesis hepática Gl uconeogénesis renal Cuer pos cetón icos 5,0 glucosa dietaria glucogenólisis gluconeogénesis 4 ,0 3 ,0 6 4 2 ,0 2 1,0 0 0 10 20 30 40 Absortiva Tiempo de ayuno (en horas) 0 ,0 0 10 20 30 40 50 Tiempo de ayuno (en días) Post-Absortiva Ayuno Prim ario Secundario Terciario Ayuno Fase postabsortiva. Esta fase dura mientras existe una cantidad suficiente de glucógeno hepático para mantener la concentración de glucosa en la sangre. El hígado tiene una capacidad limitada de acumular glucógeno. Normalmente puede almacenar suficiente glucógeno como para satisfacer las necesidades de glucosa del cuerpo durante unas 24 horas sin otra fuente de alimentación. Cuando el glucógeno se consume, el hígado comienza a degradar proteínas y usa sus aminoácidos para convertirlos en glucosa. Aquí se considera que comienza el ayuno. Esta fase que comienza después que los alimentos dietarios fueron eliminados de la circulación y abarca hasta que la provisión de glucógeno hepático permite controlar la concentración de la glucosa sanguínea, se denomina fase post-absortiva. Por lo que dijimos antes esta fase acaba aproximadamente a las 24 hs después de haberse realizado la alimentación. Como dijimos, la absorción o la liberación de glucosa por el hígado y en que proporción ocurre alguno de estos procesos están determinadas primariamente por su concentración en la sangre e influenciada por la acción de hormonas como la insulina, el glucagón y la somatostatina, todas ellas producidas por el páncreas. La insulina estimula la absorción de glucosa por las células, disminuyendo la glucosa sanguínea. Por el contrario, el glucagón promueve la degradación del glucógeno, incrementando así la glucosa sanguínea. En esta fase comienza la gluconeogénesis hepática que (a diferencia de lo que va a ocurrir en el ayuno) utiliza como fuente de esqueletos carbonados al lactato producido en otros tejidos por fermentación de la glucosa. De esta manera se establece un ciclo metabólico entre órganos que tiene al hígado como centro. Este ciclo conocido como ciclo de Cori en honor a los investigadores que lo describieron, provoca un menor gasto de glucosa pero no implica la generación de nueva glucosa en el cuerpo ya que simplemente esta se recicla. Los órganos periféricos utilizan glucosa y por fermentación producen lactato, al que vuelcan a circulación. El lactato llega al hígado donde, utilizando energía proveniente de la degradación de los ácidos grasos, es convertido nuevamente en glucosa. Finalmente el hígado libera la glucosa sintetizada nuevamente a la circulación general. Un ciclo similar al de Cori se establece utilizando alanina en lugar de lactato. El ciclo de la alanina requiere de reacciones de transaminación en los órganos que la producen y en el hígado para la conversión de la misma en piruvato, el cual después es convertido en glucosa en el hígado. Ayuno. En la primer fase del ayuno la glucosa es aportada fundamentalmente por la gluconeogénesis hepática a partir de aminoácidos. Estos aminoácidos provienen mayoritariamente de proteínas musculares que son las que el organismo puede descartar con menor dificultad. Es importante recordar que el cuerpo no posee un sistema de almacenamiento de proteínas por lo que cualquier actividad metabólica en este período que tienda a mantener la concentración de la glucosa en la sangre implica la degradación de componentes útiles para el cuerpo y por lo mismo la pérdida de funciones. Por lo mismo en un ayuno prolongado (más de un día), donde sólo se consume agua, lo que más sustancialmente disminuye es la masa muscular y, con ello, la actividad corporal, que corresponden a las actividades menos esenciales. El ayuno prolongado puede durar hasta 1 mes y medio aproximadamente. Al comenzar el ayuno el hígado comienza a sintetizar cuerpos cetónicos. Estos son enviados a la sangre y desde allí llegan a todos los tejidos. Los cuerpos cetónicos pueden ser procesados en todos los órganos aeróbicos de la misma manera que los ácidos grasos, ya que de hecho son ácidos grasos de cadena muy corta. Justamente los cuerpos cetónicos pueden ser utilizados como combustible por las células del sistema nervioso central, a pesar que las mismas son incapaces de ser alimentadas por ácidos grasos. Estas células son completamente aeróbicas, sin embargo su combustible principal es la glucosa ya que en condiciones normales, los ácidos grasos no pueden llegar hasta la superficie de estas células debido a que no pueden atravesar la barrera hemato-encefálica. Esta es una barrera celular que protege al cerebro y al sistema nervioso central, y que posibilita el paso únicamente de moléculas pequeñas como la glucosa y los cuerpos cetónicos. Los ácidos grasos debido a su insolubilidad en solventes acuosos son transportados por proteínas en la sangre y los complejos lipoproteicos son demasiado grandes como para atravesar la barrera mencionada. De esta manera las células del cerebro comienzan a ser alimentadas por otros combustible y la tasa de utilización de glucosa disminuye, disminuyendo la necesidad de su síntesis en el hígado. Los tejidos que no pueden dejar de consumir glucosa son los tejidos estrictamente anaeróbicos en el organismo, como por ejemplo los glóbulos rojos, que carecen de núcleos y mitocondrias, o las células de la retina del ojo que poseen una irrigación (y por lo mismo una oxigenación) defectuosa para no entorpecer su función. De esta manera la utilización de la glucosa en el ayuno se va progresivamente limitando a los tejidos que dependen exclusivamente de ella. Los cuerpos cetónicos se constituyen en el principal combustible de las diferentes células del organismo en las etapas de ayuno secundario. En este momento también empieza a funcionar más activamente la gluconeogénesis renal. Cuando este último proceso supera en importancia a la gluconeogénesis hepática, comienza la fase final del ayuno, o ayuno terciario. La aparición de la gluconeogénesis renal está íntimamente relacionada con el aumento de la concentración de cuerpos cetónicos en la sangre. Los cuerpos cetónicos presentas dos problemas principales para el organismo que intenta utilizarlos como moléculas combustibles en lugar de la glucosa. En primer lugar, los cuerpos cetónicos son filtrados de la sangre en los riñones, pero a diferencia de la glucosa, el cuerpo no posee un mecanismo para reabsorberlos. Así, los cuerpos cetónicos se pierden activamente en la orina. Por lo mismo, el cuerpo debe realizar un esfuerzo mayor para restaurar la energía metabólica que está perdiendo sin que haya sido utilizada. En segundo lugar, los cuerpos cetónicos son ácidos, y su aumento en concentración provoca el consumo de la capacidad buffer de la sangre y genera una disminución en el pH sanguíneo. Para evitar la acidificación de la sangre y la consiguiente acidosis metabólica que se genera, la realización de la gluconeogénesis en el riñón tiende a generar bicarbonato que puede ser volcado en la sangre y compensar así el cambio de pH. La gluconeogénesis renal se realiza a partir de glutamina. La reacción global de la conversión de glutamina en glucosa implica la generación de 4 CO2 y de 4 NH4+ por cada glucosa sintetizada. Al disolverse en sangre el CO2 se convierte en CO3H-. Si todos los productos fueran liberados en sangre, existiría el problema de la alta toxicidad del NH4+ y de la incapacidad de controlar el pH ya que se liberaría O 2 C H2 N C OH H O 2 NH 4+ 2 CH2 O C H2N OH O 2 NH4 C H CH2 C C NH2 O glutamina OH C O 2 CH2 CH2 C + CH2 CH2 2 NADH OH O C OH α-cetoglutarato glutamato 2 NADH 2 CO3H- 2 ATP 2 QH2 O H C H 2 NADH 2 CO3 C OH O HO C H H C OH H OH C 2 2 NADH 2 ATP C OH O C O P CH2 O 2 OH OH O CH2 2 ATP O fosfo-enolpiruvato C C OH CH2 OH glucosa H- C OH oxalacetato bicarbonato de amonio al medio. Sin embargo el riñón se encuentra estratégicamente ubicado para obtener el máximo beneficio de esta reacción. En las células renales donde se realiza la gluconeogénesis existen sistemas que permiten controlar la excreción del amonio hacia el exterior del cuerpo (la orina) y del bicarbonato hacia el interior (la sangre). Como el riñón es el otro órgano, además del hígado, que posee la enzima glucosa-6-fosfatasa, también es capaz de enviar la glucosa libre a la sangre aportando al mantenimiento de su homeostasis. El proceso de la gluconeogénesis renal nunca aporta demasiada glucosa al organismo. Sin embargo se torna más importante que la gluconeogénesis hepática al comenzar la fase terciaria del ayuno. De cualquier manera, en esta etapa comienzan a producirse pérdidas de función más trascendentes que complican cada vez más el cuadro del individuo en ayuno, y que pueden desembocar en comas irreversibles.