Biología - Bioenergética - Fotosíntesis
Anuncio

BIOLOGÍA
FOTOSÍNTESIS
CARACTERES GENERALES
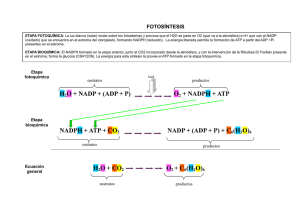
-La fotosíntesis es un conjunto de reacciones que conducen a la obtención de:
· energía química (ATP),
· poder reductor (NADP.H2) y
· precursores metabólicos, mediante procesos fotoquímicos.
-En la fotosíntesis diferenciamos dos fases:
a)Fase luminosa:
Los pigmentos de la membrana tilacoidal, junto con una cadena transportadora de electrones, captan
la energía solar que servirá para producir ATP y compuestos reducidos (NADP.H2).
b)Fase oscura o sintética:
La energía (ATP) y el poder reductor (NADP.H2) producidos en la fase lumínica se emplean para
reducir y asimilar el carbono que se encuentra en la naturaleza en un estado altamente oxidado (CO2).
-La fotosíntesis tiene lugar en los cloroplastos de las células eucariotas y en los mesosomas de las
procariotas. Se produce gracias a la presencia de la clorofila, que es capaz de absorber energía luminosa y
transformarla en energía química de enlace (ATP).
Como proceso anabólico es un proceso reductor, y requiere una fuente dadora de e-/H+ para llevar a cabo
esa reducción.
FASE LUMINOSA
Caracteres generales:
a)dependiendo de la naturaleza del dador de electrones se diferencian:
-fotosíntesis oxigénica, en la que la fuente de e- es el H2O (se libera O2) y que afecta a todos los
vegetales eucariotas (excepto hongos) y a cianobacterias.
-fotosíntesis anoxigénica, en la que la fuente de e- es un compuesto reducido (H2S) distinto del agua
(no se libera O2) y que afecta a determinados tipos de bacterias puesto que la mayoría son
heterótrofas.
b)en la membrana tilacoidea se encuentra la 'maquinaria molecular' que lleva a cabo las reacciones
fotoquímicas y que está integrada por tres tipos de componentes:
-los fotosistemas (PS I y PS II).
-la cadena transportadora de electrones.
-la ATP-sintetasa.
(A continuación veremos las características de estos elementos y, después, estudiaremos el mecanismo de
reacciones fotoquímicas).
1) FOTOSISTEMAS
-Cada fotosistema es un conjunto molecular muy complejo formado por un grupo de varios tipos de
pigmentos, que está unido a la membrana tilacoidal a través de proteínas.
-Los pigmentos fotosintéticos son de tres clases:
a) Clorofilas (a, b, c, d y bacterioclorofila, dan coloraciones verdes)
Se diferencian en los sustituyentes del anillo porfirínico (que posee Mg++ y una cadena de fitol).
Absorben y fijan eficazmente la energía luminosa, quedando "excitadas" (lo cual implica la liberación
de e- / H+ hacia un aceptor de los mismos y la necesidad de recuperarlos para restablecer su equilibrio
o estabilidad).
b) Carotenoides (anaranjados, rojos, amarillos)
También absorben energía lumínica, aunque menos eficazmente que las clorofilas.
la energía absorbida la transfieren a la clorofila.
c) Ficobilinas (rojas o azules)
También transfieren la energía luminosa que absorben a las clorofilas.
Estos pigmentos se hallan en las algas rojas (eucariotas) y en las algas verde-azules o cianobacterias
(procariotas).
-Los fotosistemas son de dos tipos:
a) PS II
Acepta e-/H+ del agua y por ello se asocia con la liberación de oxígeno.
Es propio de las células de organismos oxigénicos (cianobacterias, algas-eucariotas y arquegoniadas)
b) PS I
Es el responsable de la reducción del NADP, necesario para la fase oscura.
Se halla en todos los organismos fotosintéticos.
2) CADENA TRANSPORTADORA DE ELECTRONES
-Es similar a la de las mitocondrias.
-Los
transportadores
de
e-
(que
poseen
iones
metálicos
que
sufren
óxido-reducciones),
son
la
plastoquinona, el complejo citocromo b6-f y la plastocianina.
-El paso de e- por el complejo citocromo b6-f está acoplado a la entrada de H+ desde el estroma al tilacoide, a
través de la membrana tilacoidal.
De esta forma se genera un gradiente de H+ rico en energía electroquímica, que se utilizará en la síntesis de
ATP (en concreto, cuando los H+ vuelvan a salir al estroma por medio de la ATP-sintetasa).
3) ATP-SINTETASA
-Es una proteína canal-bomba de la membrana tilacoidal, que genera ATP con la energía liberada por la
salida de H+ desde el tilacoide al estroma.
-La salida de H+ por el canal origina la descarga de su gradiente electroquímico, y la energía producida por
esta descarga se utiliza para sintetizar ATP en el estroma, lo que se conoce como fotofosforilación o
fosforilación fotoquímica o fotosintética.
REACCIONES FOTOQUÍMICAS
-Básicamente, consisten en lo siguiente:
1)Al incidir la luz sobre los PS, la clorofila absorbe energía y se dice que queda excitada.
Como consecuencia, ambos PS sufren la pérdida de e- / H+, precisando recuperarlos para restablecer su
equilibrio.
2)Los e-/H+ perdidos por el PS I son capturados por un transportador electrónico (ferredoxina) que se
reduce.
La ferredoxina vuelve a oxidarse cuando cede los e- / H+ al NADP y éste se reduce transformándose en
NADP.H2 (poder reductor, que es necesario en la fase oscura).
3)El PS I ha de recuperar los e-/H+ perdidos para restablecer su equilibrio, y lo hace al incorporar los e- /
H+ procedentes del PSII que también los cede cuando sobre él incide la luz.
4)Los e-/H+ fluyen desde el PS II al PS I a través de un sistema redox (cadena de transporte) constituido
por la plastoquinona, el complejo citocromo b6-f y la plastocianina.
Cada vector o transportador se reduce cuando capta los e-/H+ y se oxida al cederlos.
5)En el flujo de e-/H+ anterior se libera la suficiente cantidad de energía como para que ésta quede
almacenada en forma de ATP.
A este fenómeno de atrapamiento energético y síntesis de ATP, se le llama fotofosforilación o
fosforilación fotosintética.
6)Los e-/H+
que necesita el PS II para recuperar su estabilidad proceden de la fotolisis del agua, es
decir, de la ruptura de estas moléculas por acción de los fotones lumínicos.
En esta reacción fotolítica se desprende oxígeno, que es liberado a la atmósfera.
2 H2O Æ O2 + 4 e- + 4 H+
-El tipo de fosforilación que hemos descrito es la fosforilación acíclica (transporte no cíclico de electrones),
porque la fuente de e-/H+ que fluyen y generan energía química de enlace (ATP) es una fuente externa (el
agua).
Actualmente sabemos que, simultáneamente y compatible con ese transporte no cíclico, se presenta otra
modalidad llamada fosforilación cíclica (transporte cíclico de electrones), porque los e-/H+ fluyen en un
circuito cerrado al ser desviados desde la ferredoxina al citocromo b6-f.
En este caso:
· no se forma NADP.H2
· no interviene el PS II (no hay fotolisis del agua)
· no se libera oxígeno (mecanismo anoxigénico)
· se produce igualmente ATP
-Es importante advertir que el agua sólo se precisa en la fotosíntesis en la medida que es una fuente de e/H+, los cuales son necesarios para la formación de NADP.H2 que, a su vez, es indispensable para la fijación
y reducción del CO2 en la fase oscura.
Es decir, el agua podría ser sustituida por cualquier otra sustancia dadora de e-/H+, como ocurre en la
práctica en la fotosíntesis bacteriana, en la cual los microorganismos utilizan H2S como fuente de e- / H+ (en
este caso no se libera oxígeno y, por tanto, la fotosíntesis es anoxigénica).
FASE OSCURA
-Excepto los hongos, las células vegetales poseen las enzimas necesarias para reducir y asimilar los
sustratos inorgánicos (minerales) oxidados, y transformarlos en las biomoléculas propias de la materia viva.
La fase oscura consiste, básicamente, en la transformación de los compuestos de C, N y S (CO2, NO-3 y SO=4)
en otros reducidos que se pueden incorporar a las rutas anabólicas de compuestos orgánicos.
-Caracteres generales de las reacciones 'oscuras':
a)tienen lugar en el protoplasma de procariotas o en el estroma de los cloroplastos de eucariotas.
b)están controladas enzimáticamente y, por tanto, dependientes de la temperatura.
c)se pueden producir sin necesidad de presencia de luz.
d)requieren el aporte de NADP.H2 y de ATP.
e)tienen por objeto la elaboración de precursores metabólicos.
f)configuran una secuencia cíclica (ciclo de Calvin-Benson).
g)este ciclo reductivo implica la incorporación de CO2 por los cloroplastos hasta formar compuestos
orgánicos (CH2O)n.
h)las plantas en las que tiene lugar el ciclo de Calvin se denominan 'plantas C3' porque el primer
compuesto de síntesis orgánica que se forma es el ác.3-fosfoglicérico (compuesto con 3 átomos de
carbono).
-En el ciclo de Calvin-Benson se distinguen tres etapas:
1) Fijación del CO2
-carboxilación de la ribulosa-1-5-di-P para originar dos moléculas de ácido 3-fosfoglicérico.
-esta reacción está catalizada por la enzima RuBisCo (ribulosa-1-5-di-P-carboxilasa-oxigenasa).
-la ribulosa1-5-di-P se forma por activación con ATP de la ribulosa-5-P.
2) Reducción del ác. 3-fosfoglicérico
-se realiza utilizando NADP.H2 y ATP.
-se produce a lo largo de dos momentos sucesivos:
a)el ác. 3-fosfoglicérico se transforma, con ATP, en 1-3-difosfoglicérico.
b)éste último ácido, a su vez, se reduce con NADP.H2 para formar gliceraldehído-3-P.
-el gliceraldehído-3-P, mediante una isomerasa se transforma en dioxicetona-3-P ('pool C3').
-una sexta parte del 'pool C3' es utilizada para la síntesis de osas, ác. grasos, AA, etc., y las cinco sextas
partes restantes se emplean en cerrar el ciclo de Calvin-Benson.
3) Regeneración de la ribulosa-5-P
-se realiza a partir de 5/6 partes del 'pool C3', por medio de una serie de reacciones que constituyen la
llamada 'vía de las pentosas'.
FOTORRESPIRACIÓN
-Es un proceso que se desencadena como consecuencia de la actividad oxigenasa que tiene la RuBisCo.
-Caracterísitcas de la RuBisCo:
a)está considerada como la proteína más abundante de la Tierra, ya que representa el 50% de las
proteínas que se encuentran en las hojas verdes.
b)la eficacia fotosintética depende en gran medida de su actividad.
c)es una enzima bifuncional, es decir, según las [CO2] y [O2] puede desarrollar una u otra actividad:
· Actividad carboxilasa: cataliza la combinación de la ribulosa-1-5-di-P con el CO2.
· Actividad oxigenasa: cataliza la combinación de la ribulosa-1-5-di-P con el O2.
Esta oxigenación de la ribulosa-1-5-di-P origina dos moléculas:
-una C3, fosfoglicérico (como en la carboxilación).
-otra C2, fosfoglicocólico.
(Las transformaciones de este ácido constituyen la vía metabólica de la fotorrespiración).
-Características de la fotorrespiración:
a)es una vía metabólica que se produce al mismo tiempo que la fotosíntesis, cuando el ambiente es cálido
y seco (los estomas de las hojas se cierran para evitar la pérdida de agua por transpiración).
b)por tanto, se produce en presencia de luz.
c)implica un consumo de O2 porque la RuBisCo actúa como oxigenasa (y oxida a la ribulossa-1-5-di-P
para transformarla en ác. 3-P-glicérico y ác. P-glicocólico).
d)el ác. P-glicocólico pasa a los peroxisomas, donde por cada dos moléculas del mismo se obtiene una de
P-glicérico y una de CO2.
e)es un mecanismo que disminuye la eficacia fotosintética de un 30 a un 50% y además consume ATP.
(No se conocen bien las funciones de la fotorrespiración, pero se acepta que es sólo una consecuencia de la
actividad oxigenasa de la enzima RuBisCo).
CICLO DE HATCH-SLACK
-En la fase oscura, al fijar el CO2, algunas plantas superiores han desarrollado una ruta metabólica auxiliar
que les permite crecer eficazmente en zonas tropicales (plantas C4) o en zonas desérticas (plantas CAM metabolismo ácido de las crasuláceas).
-En las plantas C4 la función oxigenasa de la RuBisCo es prácticamente nula y ello es debido a que poseen
vías metabólicas distintas.
Como consecuencia:
a)la vía auxiliar de Hatch-Slack actúa como una 'bomba' que impulsa el CO2 atmosférico hasta las células
(donde se produce el ciclo de Calvin).
b)al actuar como una bomba de CO2, aumenta la presión parcial del mismo y, por tanto, incrementa la
actividad carboxilasa de la RuBisCo (evitándose la fotorrespiración).
-En la vía Hatch-Slack se distinguen tres etapas:
1) Carboxilación
Consiste en la incorporación de CO2, al fosfoenolpirúvico (C3-PEP) para producir oxalacético (C4).
2) Descarboxilación
Consiste en la liberación de CO2 que, a su vez, se fija en el ciclo de Calvin-Benson (a la ribulosa-1-5-diP).
3) Regeneración del PEP
A partir del oxalacético, y a través de varios intermediarios C4, se forma pirúvico y, mediante ATP,
fosfoenolpirúvico.
-La fijación de CO2 por esta ruta C4 gasta más ATP que por la ruta C3, pero ...
· de cualquier modo, este gasto está sobradamente compensado por el ahorro que supone mantener la
fotorrespiración en niveles casi inexistentes, y ...
· como consecuencia, las plantas C4 pueden sobrevivir en concentraciones de CO2 muy bajas (zonas
tropicales y subtropicales)
-Las plantas CAM emplean el mismo proceso que las plantas C4 para fijar el CO2, pero lo hacen durante la
noche, cuando sus estomas están abiertos.
Por tanto, la difusión del CO2 atmosférico hacia el interior de la planta es nocturna. Y el primer compuesto
C4 lo producen al día siguiente, a la luz del día, cuando la RuBisCo puede actuar.
FIJACIÓN FOTOSINTÉTICA DEL N Y S
-En la fotosíntesis, además de glucosa, se elaboran otros compuestos, (aminoácidos y nucleótidos), que
contienen grupos amino (-NH) y tiol (-SH).
-Tanto en el agua y en el suelo, el N y el S se encuentran en forma de compuestos oxidados (NO-3 y SO=4),
que son absorbidos por las plantas y reducidos para ser incorporados a la materia orgánica.
En ambos casos, el proceso es lineal (no cíclico) y se sirve del NADP.H2 generado en la fase luminosa de la
fotosíntesis.
-La reducción fotosintética del N comprende dos etapas catalizadas por enzimas específicas:
1)La transformación de nitratos en nitritos y la de éstos en amoníaco.
2)El amoníaco se combina con el ác. alfa-cetoglutárico para formar glutámico.
(También el NH3 se combina con el ác. glutámico y forma glutamina, la cual puede transferir grupos
amino (mediante transaminasas) a otros compuestos para sintetizar aminoácidos y nucleótidos).
-La reducción fotosintética del S también es un proceso lineal en el que los sulfatos se reducen a sulfitos y
éstos a H2S. Requiere NADP.H2 y ATP.
El H2S se puede incorporar como grupo tiol (-HS) a la cisteína.
FACTORES CONDICIONANTES DE LA FOTOSÍNTESIS
1) La intensidad de la luz: la actividad fotosintética aumenta con la intensidad lumínica hasta alcanzar un
límite máximo característico de cada especie.
2) La temperatura: en general, el rendimiento fotosintético aumenta con la temperatura hasta alcanzar un
límite máximo por encima del cual puede producirse la desnaturalización de las enzimas.
3) La concentración del CO2: la actividad fotosintética aumenta conforme va creciendo la concentración de
CO2 hasta alcanzar un límite en el cual se estabiliza.
4) La concentración de O2: cuando ésta aumenta, el rendimiento fotosintético decae.
5) La humedad ambiental: cuando hay escasez de vapor de agua en la atmósfera, los estomas de las hojas
se cierran para evitar pérdidas de la misma por transpiración, y se dificulta el paso del CO2, con lo cual el
rendimiento fotosintético baja.
6) El fotoperíodo: el rendimiento está en relación directa a las horas de exposición a la luz que tenga la
planta.
http://www.loseskakeados.com