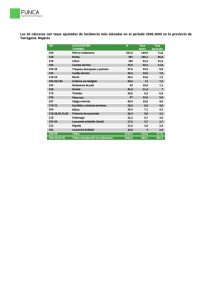
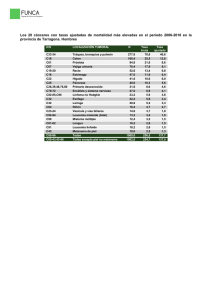
wendy espinosa hernández
Anuncio

UTILIZACIÓN DE ANTÍGENOS DEL VIRUS DE LA POLIOMIELITIS HUMANA PARA LA INMUNOTERAPIA DEL MELANOMA. WENDY ESPINOSA HERNÁNDEZ ASESOR INTERNO: M.B.E. Enrique Mendieta Márquez. ASESORA EXTERNA: Dra. Leticia Rocha Zavaleta. INSTITUTO DE INVESTIGACIONES BIOMÉDICAS. DEPARTAMENTO DE BIOLOGÍA MOLECULAR Y BIOTECNOLOGÍA. UNIVERSIDAD AUTONOMA METROPOLITANA ÍNDICE UTILIZACIÓN DE ANTÍGENOS DEL VIRUS DE LA POLIOMIELITIS HUMANA PARA LA INMUNOTERAPIA DEL MELANOMA. .................................................... 5 RESUMEN ....................................................................................................................... 5 UTILIZACIÓN DE ANTÍGENOS DEL VIRUS DE LA POLIOMIELITIS HUMANA PARA LA INMUNOTERAPIA DEL MELANOMA. .................................................... 6 ANTECEDENTES........................................................................................................... 6 INTRODUCCIÓN ........................................................................................................... 9 VIRUS DE LA POLIOMIELITIS HUMANA................................................................ 9 TRATAMIENTO DE LA POLIOMIELITIS ............................................................... 12 PREVENCIÓN DE LA POLIOMIELITIS................................................................... 13 CÁNCER DE PIEL........................................................................................................ 14 MELANOMA................................................................................................................. 15 FORMAS DE MELANOMA......................................................................................... 17 ESTADIAJE DEL MELANOMA ................................................................................. 18 FACTORES DE RIESGO ............................................................................................. 19 TRATAMIENTO ........................................................................................................... 20 ¿CÓMO PREVENIRLO? ............................................................................................. 20 PAPEL DEL SISTEMA INMUNE................................................................................ 21 RESPUESTA INMUNE HUMORAL ........................................................................... 23 EVASIÓN INMUNOLÓGICA...................................................................................... 24 TERAPIA GÉNICA....................................................................................................... 25 PROPIEDADES INMUNOGÉNICAS DE LAS PROTEÍNAS VP1 Y VP4. ............... 27 OBJETIVO GENERAL ................................................................................................ 30 OBJETIVOS PARTICULARES ................................................................................... 30 METODOLOGÍA .......................................................................................................... 31 SUBCLONACIÓN DE LAS PROTEÍNAS VP1 Y VP4 DEL PLÁSMIDO PCVP1 Y PCVP4 RESPECTIVAMENTE. ................................................................................... 31 CULTIVO DE BACTERIAS E. COLI ......................................................................... 31 PURIFICACIÓN DEL DNA PLASMÍDICO (PROCEDIMIENTO DE MINIPREPARACIÓN), MEDIANTE LISIS ALCALINA. ......................................... 31 RESTRICCIÓN ENZIMÁTICA................................................................................... 32 2 PURIFICACIÓN DE DNA POR EXTRACCIÓN FENOL-CLOROFORMO Y PRECIPITACIÓN POR ETANOL (MODIFICACIÓN DE MANIATIS ET. AL, (1989))............................................................................................................................. 32 REACCIONES DE LIGACIÓN.................................................................................... 33 TRANSFORMACIÓN CELULAR, UTILIZANDO CÉLULAS E. COLI DE LA CEPA BACTERIANA JM101....................................................................................... 33 SIEMBRA DE LAS CÉLULAS TRANSFORMADAS ................................................ 33 MASTER PLATE .......................................................................................................... 34 AMPLIFICACIÓN DE LA SECUENCIA CODIFICADORA DE LAS PROTEÍNAS VP1 Y VP4 DEL VIRUS DE LA POLIOMIELITIS.................................................... 34 STOCK EN GLICEROL DE CÉLULAS BACTERIANAS TRANSFORMADAS ... 34 DESCONGELAMIENTO DE LAS CÉLULAS B16FO............................................... 34 CONTEO DE CÉLULAS .............................................................................................. 35 VACUNACIÓN DE LOS ANIMALES ......................................................................... 35 INDUCCIÓN DE TUMORES ....................................................................................... 36 TRATAMIENTO DE TUMORES ................................................................................ 36 ANÁLISIS ESTADÍSTICO........................................................................................... 36 RESULTADOS .............................................................................................................. 37 OBTENCIÓN DE LOS FRAGMENTOS GÉNICOS VP1 Y VP4............................... 37 PREPARACIÓN DEL VECTOR PDISPLAY ............................................................. 40 SUBCLONACIÓN DE LOS GENES VP1 Y VP4 ........................................................ 43 SUBCLONACION DEL GENE VP1 ........................................................................... 43 SUBCLONACIÓN DEL GENE VP4 ........................................................................... 44 ANÁLISIS DE LAS COLONIAS RECOMBINANTES POR RESTRICCIÓN ENZIMÁTICA. .............................................................................................................. 45 ANÁLISIS PARA VP1 ................................................................................................ 45 ANÁLISIS DEL DNA DE LAS COLONIAS RECOMBINANTES ............................ 47 ANÁLISIS POR RESTRICCIÓN ENZIMÁTICA PARA VP4 ................................... 48 ANÁLISIS POR PCR PARA VP4................................................................................. 50 TRATAMIENTO DE LOS TUMORES ....................................................................... 51 EL DISEÑO EXPERIMENTAL DE VACUNACIÓN FUE EL SIGUIENTE:........... 52 DESARROLLO DE TUMORES DE MELANOMA.................................................... 53 TRATAMIENTO DE LOS TUMORES ....................................................................... 54 DISCUSIÓN ................................................................................................................... 65 3 CONCLUSIÓN .............................................................................................................. 69 PERSPECTIVAS ........................................................................................................... 70 BIBLIOGRAFÍA ........................................................................................................... 71 4 Utilización de Antígenos del Virus de la Poliomielitis Humana para la Inmunoterapia del Melanoma. RESUMEN De las diversas estrategias para combatir el cáncer, la inmunoterapia es una de las más prometedoras ya que busca estimular la respuesta inmune del propio paciente para combatir y eliminar a las células malignas que provocan alteraciones en el equilibrio interno. Esta terapia intenta aprovechar la especificidad, la capacidad de respuesta y la memoria de las células del sistema inmune y de sus moléculas efectoras. Además tiene la ventaja de que evita los efectos sistémicos indeseables de las terapias convencionales. Así, para despertar una respuesta inmune específica contra las células malignas es necesario que éstas expresen antígenos diferentes a los de las células sanas y así sean reconocidas como extrañas y posteriormente eliminadas. Para este trabajo se eligió al melanoma como modelo debido a que tiene la ventaja de que el tejido alterado se puede observar y medir sin dificultad, además de que este tipo de cáncer es un problema de salud pública a nivel mundial (cada año se producen en el mundo cien mil nuevos casos, su incidencia se sitúa entre el 1.5 y el 7 %, en varones es el sexto tipo de cáncer más común y en mujeres es el séptimo). En este experimento se indujo la expresión de antígenos del virus de la poliomielitis en células tumorales en ratones previamente inmunizados contra la poliomielitis y, considerando trabajos recientes en que se detectan células T y B específicas contra epítopes del virus, se determinó el efecto antitumoral. Para llevar a cabo lo anterior se provocó el desarrollo de melanoma en los ratones inmunizados y se administraron directamente en el tumor plásmidos que contienen las secuencias génicas de las proteínas estructurales VP1 y VP4 del virus de la polio. Los resultados indican que los ratones inmunizados, tratados con los plásmidos antes mencionados, presentaron una regresión del tumor y cicatrización total del tejido, lo que señala a los antígenos del virus expresados en el tumor como los blancos de las células inmunes y éstas como responsables del efecto antitumural. En conjunto, este trabajo abre la posibilidad de combatir ciertos tipos de cáncer aprovechando las cualidades del sistema inmunológico y de explorar cuáles son los tipos de células y moléculas que llevan a cabo las funciones efectoras para poder optimizar y aprovechar sus funciones. 5 Utilización de Antígenos del Virus de la Poliomielitis Humana para la Inmunoterapia del Melanoma. ANTECEDENTES Los Enterovirus constituyen un amplio grupo de virus causantes de importantes infecciones humanas, pertenecen a la familia Picornaviridae (pico=pequeño, rna=virus de RNA) e integran el género más virulento debido a que provocan una gran cantidad de manifestaciones clínicas y subclínicas donde los pacientes pueden sufrir desde una enfermedad febril leve hasta una parálisis muy severa y permanente que puede ser fatal; tal es el caso de la poliomielitis.2,3,10 Los Enterovirus presentan una cápside de simetría icosaédrica formada por 60 protómeros, sin envoltura lipídica; tienen un tamaño de 20-30 nm, poseen RNA de cadena única de 7209-18450 nucleótidos y polaridad positiva, por lo que se replican utilizando el propio RNA como mensajero. Su genoma se divide en 4 regiones que codifican proteínas estructurales y 2 regiones no codificadoras, reguladoras. La cápside está compuesta de 4 tipos diferentes de proteínas estructurales: VP1, VP2, VP3 y VP4 que se sintetizan como una poliproteína.3,9, 12,25,27 Salk desarrolló la primera vacuna contra la poliomielitis de administración intramuscular a partir de los tres virus inactivados con formol. Posteriormente en 1964 Sabin y Bougler desarrollaron una vacuna de virus activos de administración oral. Se la llamó Trivalente porque atacaba a los tres virus. La inmunidad producida por la vacunación es predominantemente, humoral, específica de serotipo y duradera [los anticuerpos persisten durante toda la vida.8,9,12,14,25 La generación de las vacunas altamente eficientes (Salk y Sabin) y su uso mundial permitió que la Organización Mundial de la Salud (OMS) declarará en 1988 la meta de eliminar todos los casos de poliomielitis en el mundo hacia el año 2000. Si bien esta meta no ha sido completamente realizada, en el continente americano ya no hay casos de esta enfermedad; gracias a que las compañas de inmunización serán llevadas a países en vías de desarrollo, esto pronto será posible.9,10,17 El melanoma es un tipo de cáncer de piel que aparece cuando los melanocitos se convierten en malignas. Representa el 2.5% de todos los cánceres. El melanoma se desarrolla de “novo” en el 75% de los casos y en el 25% se desarrolla sobre un nevus preexistente. Los individuos con piel clara, rubios o pelirrojos se ven más afectados. Los pacientes de raza negra tienen una incidencia 10 veces menor que los de raza blanca. Un 50% de melanomas se observan en personas menores de 50 años y el 35% en menores de 35 años.13 El 95% de melanomas se desarrolla en la piel, pero pueden desarrollarse en otras localizaciones como el ojo y mucosa oral o genital. Un 3% de pacientes 6 desarrollan melanomas ocultos (enfermedad metastásica sin evidencia de tumor). En total representa el 65% de todas las muertes por cáncer de piel, lo que indica su alto potencial de agresividad y capacidad metastásica. Existen varios factores de riesgo que aumentan la posibilidad de desarrollar un melanoma, entre los cuales están historia familiar de melanoma, tendencia a formar pecas, piel blanca, exposición excesiva al sol, agentes químicos, físicos, inmunológicos entre otros. Cada año se producen en el mundo 100.000 nuevos casos y su incidencia se sitúa entre el 1.5 y el 7%.13,15 A principios de los 90’s, gracias al conocimiento de todos los mecanismos moleculares que participan en el control normal del crecimiento, las diferenciación celular y la transformación maligna, se ha hecho factible al idea de tratar de intervenir a nivel molecular en el tratamiento de algunos tipos particulares de tumores malignos, entre ellos el melanoma, con novedosas alternativas por ejemplo la terapia génica.4 El concepto de terapia génica resulta de la observación de que ciertas enfermedades, entre ellas particularmente el cáncer, resultan de daños genéticos específicos estos se agrupan en tres grandes categorías: ¯ La activación de genes dominantes (proto-oncogenes oncogenes). ¯ La pérdida de genes supresores de tumores. ¯ La supresión local de la respuesta inmunes.1 Existe una enorme variedad de posibles aplicaciones de este tipo de terapia, pues ésta tiene el potencial de corregir defectos genéticos, combatir infecciones o destruir tumores.31 La elección de la estrategia de terapia génica anti-neoplásica depende del método que se desea emplear para destruir a las células tumorales. Según el método seleccionado, las células malignas pueden ser reconvertidas a un fenotipo no maligno, o ser eliminadas por el sistema inmune, o por inhibición de la neovascularización tumoral, o por efectos tóxicos directos.29 Una de ellas es la inmunoterapia. Esta estrategia involucra la activación de la respuesta inmune contra las células tumorales in vivo. En esta modalidad, las células que se han modificado ex vivo con genes que codifican paras citocinas, antígenos o moléculas alogénicas del complejo mayor de histocompatibilidad, potencian la inmunogenicidad del tumor. Otra opción es la de inmunizar activamente con antígenos asociados a tumor que han sido clonados.29 Existen muchos estudios pre-clínicos que han demostrado que las inmunizaciones basadas en DNA contra modelos de antígenos tumorales tales 7 como la albumina, β-galactosidasa, péptidos (IL-2)19, otros han utilizado diferentes los propios antígenos33 o superantígenos tumorales20, células manipuladas para secretar diferentes factores (GM-SCF)32, y diversidad de terapias que utilizan diversos al interferón34 o adyuvantes.6 Sin embargo a pesar de la existencia de diversos estrategias, aún no se ha encontrado un tratamiento eficaz contra el cáncer. En el Departamento de Biología Molecular y Biotecnología de la UNAM se realizó un proyecto basado en la inmunoterapia, en el cual se tratará de iniciar una respuesta inmune, por medio de la introducción de genes, que codifican proteínas (VP1 y VP4) del virus de poliomielitis humana clonados en el vector pDISPLAY Estos plásmidos serán inyectados en tumores para inducir la expresión de las proteínas de forma transmembranal en las células tumorales, mismas que el sistema inmune reconocerá como extrañas y eliminará; para así inducir regresión del melanoma. 8 INTRODUCCIÓN VIRUS DE LA POLIOMIELITIS HUMANA. Los Enterovirus constituyen un amplio grupo de virus causantes de importantes infecciones humanas, pertenecen a la familia Picornaviridae (pico=pequeño, rna=virus de RNA) e integran el género más virulento debido a que provocan una gran cantidad de manifestaciones clínicas y subclínicas donde los pacientes pueden sufrir desde una enfermedad febril leve hasta una parálisis muy severa y permanente que puede ser fatal.2,3,10 Los Enterovirus presentan una cápside de simetría icosaédrica formada por 60 protómeros [figura 1], sin envoltura lipídica por lo que son resistentes a éter u otros solventes; tienen un tamaño de 20-30 nm, poseen RNA de cadena única de 7209-18450 nucleótidos y polaridad positiva [figura 2], por lo que se replican utilizando el propio RNA como mensajero. Su genoma se divide en 4 regiones que codifican proteínas estructurales y 2 regiones no codificadoras, reguladoras. La cápside está compuesta de 4 tipos diferentes de proteínas complejas. Sus cuatro proteínas estructurales: VP1, VP2, VP3 y VP4 se sintetizan como una poliproteína en la que el extremo 5' está unido covalentemente a una pequeña proteína VPg de 20 a 24 aminoácidos (regulación genética). Poseen en el extremo 3’ residuos de ácidos adenílico (secuencia de Poli-A). La unión de las proteínas VP1 Y VP3 da lugar a la formación de un canal estructural que es utilizado por el virus para unirse al receptor celular.3,9,12,25,27,36,37 Figura 1. Imagen tridimensional de un poliovirus 9 El ciclo replicativo del virus es lítico y su receptor celular es una molécula perteneciente a la superfamilia de las inmunoglobulinas que se presenta en distintos órganos blanco, el sistema nervioso central, el hígado o el intestino. Son resistentes a todos los antivíricos y quimioterápicos conocidos, y a la activación por solventes de lípidos. Presentan una tendencia natural a la agregación espontánea que los del efecto de los agentes externos. Se inactivan rápidamente a más de 50°C, por la luz ultravioleta y en condiciones de desecación. Son estables a pH ácido.3 Figura 2. Esquema de un poliovirus donde se observa su RNA Los Enterovirus han sido responsables de epidemias como la poliomielitis (griego polios=gris), también llamada parálisis infantil, causada por los poliovirus; poliovirus es un nombre genérico que se aplica a tres tipos de virus: Brunhilde (tipo 1), Lansing (tipo 2), y León (tipo 3), el tipo 1 es el responsable de las epidemias, siendo la enfermedad más seria causada por éste género. Se llama infantil porque los enfermos que contraen la enfermedad son especialmente los niños entre cinco y diez años.8,9,14 Dentro de cada grupo, existe un número de serotipos definidos por los epitopos de la cápside que se deben a modificaciones estructurales de la superficie del virión. Los epitopos antigénicos, que definen a cada serotipo, estimulan la formación de anticuerpos específicos que se comportan como neutralizantes de la infección, ya que no permiten su unión al receptor celular. La evolución de los serotipos sigue un proceso de deriva antigénica, en el que durante el proceso de copia del genoma se producen errores que introducen mutaciones puntuales. Estas mutaciones son acumulativas y modifican progresivamente la estructura de la cápside [ figura 3]. La mayoría de las infecciones son abortivas, debido a la existencia de anticuerpos neutralizantes. Los anticuerpos IgG, IgM e IgA aparecen con bastante rapidez en el curso de la infección.3 10 Figura 3. Estructura de la cápside de un poliovirus en tercera dimensión. Los poliovirus infectan ciertos tipos de células nerviosas y durante su multiplicación pueden destruirlas afectando principalmente a las células de las astas anteriores de la médula espinal, pero en casos severos también se ve afectada la sustancia gris, así como los cuernos posteriores y dorsales de la médula espinal. Las lesiones se encuentran a nivel de hipótalamo y del tálamo. En al cerebro los órganos más afectados son la formación reticular, el núcleo vestibular, la vermis cerebelar y el núcleo cerebelar profundo. Pueden presentarse cambios en las neuronas, que van desde una ligera cromatoliosis hasta una neuronofagia y total destrucción.9 El registro más antiguo de data de 1300 A C en Egipto en el que se ve a un sacerdote con una deformidad en una pierna típica de poliomielitis. El primer caso de la era moderna apareció a finales del siglo XIX en países desarrollados y de allí sé extendió a países en vías de desarrollo. 10 La enfermedad fue descrita por primera vez por el alemán Jacob Heine en 1840 y se divide en tres clases: la primera, la poliomielitis abortiva, es una enfermedad febril inespecífica de 2-3 días de duración, sin signos en el Sistema Nervioso Central, apetito disminuido, vomito, estreñimiento y malestar general. Un segundo grupo de enfermos puede, además presentar una meningitis aséptica, fiebre, malestar general, dolor en los músculos del cuello, del tronco, los brazos y las piernas y rigidez del cuello y de la columna vertebral. La recuperación es rápida y completa, generalmente en unos días. La tercera clase o poliomielitis paralítica es la infección de consecuencias posibles más graves y suele ir precedida de un período de fiebre y "enfermedad menor" clásicamente, los síntomas desaparecen después de varios días. De los 5 a los 10 días, la fiebre reaparece seguida de signos de irritación meníngea debilidad generalizada, respiración debilitada, dificultad para tragar, y parálisis fláccida asimétrica como parálisis de la vejiga y de los músculos.9,10,12 11 Se transmite por contacto directo con las secreciones respiratorias, de garganta y las heces infectadas. El virus viaja con la tos y los estornudos. Se han presentado contagios a través del consumo de agua o de alimentos contaminados o por lavado de manos inadecuado. El hombre es el único reservorio conocido y la transmisión es por vía fecal-oral y respiratoria.8,12,35,36 Puede presentarse en forma endémica o en brotes epidémicos, siendo más frecuente en verano y otoño, en niveles socioeconómicos bajos y en lactante y niños (más frecuente en varones). De todas las personas (90-95%) que tienen la infección, el 2 % o menos puede desarrollar una enfermedad paralítica. La polio se presenta en todas partes del mundo. Sin embargo , en los últimos años no se ha reportado ningún caso en los Estados Unidos (cada año se presentan 8 casos de polio causada por la vacuna poliovirus oral). Se ha informado de algunos casos en hemisferio occidental.2,14 Anteriormente los principales afectados por esta enfermedad eran los bebés y los niños, pero en la actualidad son más frecuentes los caso en personas mayores de 15 años y es más común durante el verano y el otoño. En México no se han registrado casos de poliomielitis desde 1990.14 La poliomielitis empezó a controlarse en 1949 cuando el bacteriólogo John Franklin Enders logró hacer crecer los virus en el laboratorio dentro de tejidos. Basándose en esa técnica el epidemiólogo Jonás Salk desarrolló la primera vacuna contra la poliomielitis de administración intramuscular a partir de los tres virus inactivados con formol. Tras las pruebas clínicas pertinentes que demostraron que era segura, en 1954 se empezó la inoculación. Posteriormente en 1964 Sabin y Bougler desarrollaron una vacuna de virus activos de administración oral. Se la llamó Trivalente porque atacaba a los tres virus. La inmunidad producida por la vacunación es predominantemente, humoral, específica de serotipo y duradera [los anticuerpos persisten durante toda la vida].8,9,12,14,25 Tratamiento de la poliomielitis El tratamiento especifico para la poliomielitis será determinado por su médico basándose en lo siguiente: ¯ ¯ ¯ ¯ ¯ Estado general de salud e historia médica. Que tan avanzada esta la enfermedad. Tolerancia a determinados medicamentos, procedimientos o terapias. Expectativas para la trayectoria de la enfermedad. Opinión o preferencia.12 12 Prevención de la poliomielitis ¯ Higiene adecuada y lavarse bien las manos. ¯ La inmunización contra el poliovirus: en Estados Unidos, se recomienda administrar la vacuna contra la polio a las edades siguientes: 2 meses. 4 meses. Entre 6 y 18 meses. Entre 4 y 6 años.12 Como norma internacional el esquema básico de vacunación consiste en tres dosis, con intervalos de dos meses entre cada una, se inicia a los dos meses de vida, a los recién nacidos se les administra una dosis preliminar que no se considera dentro del esquema anterior, independientemente de los antecedentes de vacunación, en México se administran dosis adicionales en las Semanas Nacionales de Salud.8 La generación de las vacunas altamente eficientes (Salk y Sabin) y su uso mundial permitió que la Organización Mundial de la Salud (OMS) declarará en 1988 la meta de eliminar todos los casos de poliomielitis en el mundo hacia el año 2000. Si bien esta meta no ha sido completamente realizada, en el continente americano ya no hay casos de esta enfermedad; gracias a que las compañas de inmunización serán llevadas a países en vías de desarrollo, esto pronto será posible.9, 10,17 13 Cáncer De Piel El cáncer es un desorden que surge después de la acumulación de múltiples alteraciones en diferentes genes en las células somáticas a lo largo del tiempo en el cual estas células perdieron: la capacidad de regular la proliferación, las características del tejido de origen y su posición.13 El cáncer se clasifica de acuerdo al tejido de origen (figura 4). FIGURA 4 CLASIFICACIÓN DE LOS CÁNCERES DE ACUERDO A SU ORIGEN TIPO DE TEJIDO CÁNCER Cavidad oral. Órganos digestivos. colon y recto. Páncreas. Estómago. hígado y sistema biliar. sistema respiratorio. Pulmón. Seno. melanoma maligno. Tracto reproductivo. Glándula de la próstata. Ovario. Cervicouterino. Útero (endometrio). Órganos urinarios. Vejiga Leucemias. Linfomas. Mielomas. Eritroblastomas. Glioma. Retinoblastoma. Sarcomas. EPITELIO SISTEMA HEMATOPOYETICO E INMUNE SISTEMA NERVIOSO CENTRAL Y OJO TEJIDO CONECTIVO, MÚSCULO Y VASCULAR 14 Melanoma El melanoma es un tipo de cáncer de piel que aparece cuando los melanocitos se convierten en malignos ( forman un gránulo pigmentoso). Estas células elaboran un pigmento llamado melanina, responsable del color de la piel, el pelo y del iris de los ojos. La melanina, por su parte, funciona como un fotoprotector evitando que la radiación solar dañe las estructuras o los tejidos del cuerpo.15 La mayoría de los melanomas se localizan en la piel y menos frecuentemente en mucosas, pero dado el origen de los melanocitos –derivan de la cresta neural-, pueden observarse en otras localizaciones como retina y meninges donde también migran los melanocitos. Los melanocitos derivan de la cresta neural y durante el desarrollo embriológico se diferencian hacia melanoblastos y migran hacia la piel. Tras alcanzar la dermis migran a la epidermis o el folículo piloso y se diferencian hacia Nevomelanocitos. Las funciones de los melanocitos epidérmicos es la de producir melanina y transferirla a los queratinocitos que los rodean. Además de la función de producir melanina los melanocitos dan lugar a uno de los tumores más frecuentes del organismo: los nevus melanocíticos. 15 Según diversos estudios clínicos, histopatológicos, inmunológicos y genéticos, existen 5 niveles de progresión de los tumores melanocíticos: 1. Nevus melanocítico benigno 2. Nevus atípico 3. Melanoma maligno primario de crecimiento radial 4. Melanoma maligno primario en fase de crecimiento vertical 5. Melanoma metastásico El melanoma aparece como un lunar preexistente que cambia, o como un nuevo lunar en la piel previamente clara, del 20 al 40 % está asociado con nervi displástico (alteración en tamaño, forma y organización de los células) y puede originarse en cualquier edad pero más comúnmente ocurre después de la pubertad. Además puede presentar formas, volúmenes y colores variables y tiene la ventaja de que se puede observar y medir sin dificultad; en los hombres aparece generalmente en el tronco, desde la espalda hasta la cadera, en la cabeza o en el cuello; en cambio, en las mujeres se desarrolla principalmente en las piernas o en el tronco. 15 15 El melanoma tiene un comportamiento biológico que determina el pronóstico ya que presenta dos fases de crecimiento, una radial (intraepidérmica) y otra vertical (intradérmica). En la fase de crecimiento radial [figura. 5(A)], que puede durar meses o años, la tumoración está limitada a la epidermis sin presentar la capacidad de producir metástasis, en esta situación la extirpación quirúrgica es curativa. En la fase de crecimiento vertical [figura. 5(B)], la tumoración invade la dermis con la capacidad de producir metástasis linfáticas o sanguíneas. Este crecimiento del melanoma maligno hace necesario que al establecerse el diagnóstico, tenga que determinarse en que fase de crecimiento se encuentra, con la finalidad de establecer el pronóstico y el tratamiento. 15 A C B D Figura. 5 Fases de crecimiento del melanoma A) Los melanomas se inician en la epidermis con la proliferación de melanocitos en la unión dermo-epidérmica y B) de forma gradual presentan un crecimiento intraepidérmico para afectar a toda la epidermis. C) Cuando inician la fase de crecimiento vertical, rompen la unión dermo-epidérmica y D) pasan a invadir la dermis papilar, reticular y tejido graso subcutáneo. El melanoma representa el 2.5% de todos los cánceres. El melanoma se desarrolla de “novo” en el 75% de los casos y en el 25% se desarrolla sobre un nevus preexistente. Los individuos con piel clara, rubios o pelirrojos se ven más afectados. Los pacientes de raza negra tienen una incidencia 10 veces menor que los de raza blanca. Un 50% de melanomas se observan en personas menores de 50 años y el 35% en menores de 35 años. El 95% de melanomas se desarrolla en la piel, pero pueden desarrollarse en otras localizaciones como el ojo y mucosa oral o genital. Un 3% de pacientes desarrollan melanomas ocultos (enfermedad metastásica sin evidencia de tumor). 15 16 Formas De Melanoma. Clásicamente se describen 4 formas clínico-patológicas de melanoma cutáneo en relación con la localización anatómica y a su evolución.13, 15 MELANOMA DE EXTENSIÓN SUPERFICIAL Es la forma más frecuente de melanoma ya que representa el 70% de los casos de melanoma. entre la población blanca: constituye cerca del 50% de todos los melanomas. Aparece en cualquier zona de la piel, aunque en los hombres se localiza más en el tronco, y en las mujeres, más en las piernas. Se presenta entre los 40 y 50 años de edad, usualmente crece “oblicuamente” a lo largo de la superficie de la piel (epidermis) entonces penetra el tejido de la piel(dermis) y generalmente la lesión tiene una evolución de entre 1 y 5 años. La lesión clínica suele ser una placa pigmentada, palpable, de 1 a 2.5 cm de diámetro, de color marrón oscura, negra o parduzca, que puede presentar áreas claras que reflejan regresión tumoral. Esta forma de melanoma es curable cuando es removido antes de la invasión a la dermis. LENTIGO MALIGNO Afecta a las áreas de la piel que se han visto expuesta durante largo tiempo al sol(cara, cabeza, cuello, dorso de la mano y las piernas), se presenta entre los 60 y 70 años. comienza como una mancha moteada de pigmentación. ACRAL-LENTIGOSO El melanoma más común en la gente de piel negra. Aparece particularmente en palmas, uñas y dedos de la mano, uñas, dedos y plantas de los pies. Este tumor comprende el 2% del melanoma en caucasianos y 50% en razas de piel negra y es biológicamente muy agresivo. Se presenta a cualquier edad. NODULAR Se localiza en cualquier zona de la piel, aunque por lo general se sitúa en la cabeza y el tronco .Es más frecuente en hombres que en las mujeres. Se desarrolla entre los 50 y los 60 años de edad. Crece rápidamente el tumor sin apreciarse una mancha. Las tres formas clínicas: melanoma de extensión superficial, el léntigo maligno y el melanoma acral-lentigoso, tienen una fase de crecimiento intraepidérmica larga, en el cual el tumor está completamente limitado a la epidermis. El cuarto tipo de melanoma, el melanoma nodular, tiene una fase de crecimiento intraepidérmica muy corta con el paso al crecimiento nodular (dérmico) rápido. 17 Pero existen los siguientes subtipos de melanoma: AMELANOTICO Subtipo no común que no produce suficiente pigmento para tener el color de una lesión pigmentada. Puede ser piel coloreada o pequeñas manchas rojizas. Aparece usualmente como un nódulo rosa o rojo. NEUROTROPICO DESMOPLÁSTICO Subtipo no común que se observa como una lesión de cicatriz no pigmentada persistente. Puede ser o no pigmentada y aparece más comúnmente en piel dañada por el sol (cabeza y cuello en adultos mayores. MUCOSO Subtipo no común que ocurre en regiones mucosas de la boca, nariz y genitales. No puede ser reconocido por que ocurre en sitios que generalmente no son examinados. En la actualidad se considera el melanoma como una sola entidad que puede evolucionar hacia una forma clínico-patológica dependiendo de varios factores tales como la localización anatómica, y /o el daño solar asociado, todavía no bien establecidos. 15 Estadiaje Del Melanoma En el Estadiaje microscópico del tumor existen dos hallazgos que tienen una gran importancia pronóstica y que son el grosor tumoral y la presencia de ulceración. La ulceración es la ausencia histológica de epidermis por encima del tumor. El grosor tumoral se establece utilizando dos sistemas, un sistema basado en la medición milimétrica del grosor (índice de Breslow) y un sistema basado en el nivel cutáneo de invasión (niveles de Clark) [Figura 6]. 15 Figura 6 Estadiaje microscópico del melanoma El estadiaje microscópico establece el grosor del melanoma por medio de dos sistemas de medición. Un sistema milimétrico (Índice de Breslow) que establece el grosor tumoral en milímetros por medio de la utilización de una escala milimétrica adaptada al ocular de microscopio y un sistema basado en el nivel cutáneo de invasión. 18 La medición del espesor tumoral en milímetros, que se conoce como índice de Breslow, es una variable independiente de otros factores, que determina el pronóstico y la supervivencia de los enfermos con melanoma. Los niveles de invasión o niveles de Clark, describen la invasión de la tumoración en los diferentes niveles cutáneos. El nivel I describe los tumores puramente epidérmicos; el nivel II indica que la tumoración atraviesa la unión dermoepidérmica con invasión de la dermis papilar; el nivel III describe las tumoraciones que ocupan toda la dermis papilar expandiéndola; el nivel IV indica que la tumoración infiltra la dermis reticular y el nivel V indica la invasión del tejido graso subcutáneo. El pronóstico empeora al aumentar el nivel, pero tiene menos valor que el espesor medido por medio del índice de Breslow. 15 Factores De Riesgo Algunos atributos personales se correlacionan con un incremento en el riesgo para desarrollar cáncer de piel, entre ellos se encuentran: piel blanca con cabello rubio o rojo, ojos azules, grises o verdes, tendencia a pecas, tendencia a tostarse por el sol, lunares anormales (displásticos), historia personal o familiar de melanoma, un gran numero de lunares adquiridos y grandes o pequeños lunares congénitos.13, 15 Existen varios factores de riesgo que aumentan la posibilidad de desarrollar un melanoma, resumidos en la figura 7. La radiación solar –ultravioleta- es el factor etiopatogénico más importante en el desarrolló del melanoma maligno. 15 FIGURA 7 FACTORES QUE AUMENTAN EL RIESGO DE DESARROLLAR MELANOMA Número elevado de nevus. Presencia de nevus atípicos en pacientes Aumentan con antecedentes familiares de melanoma. mucho Lesiones pigmentadas con cambios clínicos. Historia previa de melanoma. Historia familiar de melanoma Moderadamente Nevus atípicos sin historia familiar de aumentado melanoma Fototipo de piel clara Quemadura solar fácil, sin pigmentación. Lengines múltiples. Inmunosupresión por trasplante Discretamente aumentado 19 Otros factores de riesgo identificados incluyen agentes químicos, físicos e inmunológicos. Los factores genéticos son muy importantes para el conocimiento de los factores de riesgo, incluyendo la tendencia familiar al desarrollo de melanoma, lunares prominentes, y lunares anormales. La exposición excesiva a la radiación ultravioleta de la luz del sol puede ser un factor que contribuye a algunos casos de melanoma. Los periodos cortos de exposición intensa están asociados con un incremento de dos veces el riesgo de melanoma. Cada año se producen en el mundo 100.000 nuevos casos y su incidencia se sitúa entre el 1.5 y el 7 %. El melanoma es el sexto cáncer más común en hombres y el séptimo cáncer más común en mujeres. En total representa el 65% de todas las muertes por cáncer de piel, lo que indica su alto potencial de agresividad y capacidad metastásica. 15 Tratamiento El diagnóstico precoz es esencial para la curación del melanoma. La extirpación del melanoma cuando éste esta limitado a la epidermis y no sobrepasa la membrana basal supone una supervivencia del 100%. Cuando el melanoma rompe la membrana basal e inicia su fase de crecimiento vertical el pronóstico empeora y debe realizarse un tratamiento del tumor primitivo, de la afectación ganglionar y de las metástasis sí existen. 13 En general el tratamiento del melanoma depende del tipo y del estadio del tumor, así como de otros factores como el estado de salud y la edad del paciente. Existen cuatro tipos de tratamiento que pueden utilizarse de forma combinada: cirugía, quimioterapia, inmunoterapia y radioterapia. 13 ¿Cómo prevenirlo? El sol constituye el principal factor de riesgo para desarrollar un melanoma, por lo que es recomendable tomar una serie de medidas básicas a la hora de exponerse a los rayos solares: o Evitar el efecto acumulativo del sol en la piel y las quemaduras, especialmente durante la niñez y la adolescencia, ya que la piel es mucho más sensible. o Los rayos solares son más dañinos entre las 10 de la mañana y las 2 de la tarde. o Durante los baños de sol hay que utilizar cremas, lociones o geles protectores tanto para los rayos UVA como para los UVB y que tengan como mínimo, un factor 15 de protección. Los productos más adecuados son los que tienen filtros físicos o químicos, resistentes al agua y dermatológicamente probados. o Se debe restringir el contacto con productos potencialmente fotosensibles, como los jabones, los desodorantes y los perfumes, que pueden crear una especial sensibilidad a la luz del sol. o Las gafas de sol y los sombreros son unos accesorios muy útiles ya que, además de la cara protegen el cuello y las orejas. 13 20 PAPEL DEL SISTEMA INMUNE La respuesta inmunitaria es el resultado de la interacción de células presentadoras de antígeno (APC) que capturan a los materiales antigénicos y los fagocitan para luego procesarlos. Los péptidos antigénicos o epitopos resultantes de este procesamiento se acoplan a moléculas del complejo de histocompatibilidad (MHC-II)y así son transportados hasta la membrana de las APC donde pueden ser reconocidos por las células T respondedoras (aquellas que portan el receptor complementario al epitope). Mientras procesan al antígeno, las APC producen y liberan citocinas como las interleucinas (IL) IL-1, IL-6 e IL-12, además de otras. La IL-1, junto con moléculas coestimulatorias de las APC inducen la síntesis de IL-2 por las células Th1 y de su receptor IL-TR, por las células Th1 y Th2. La IL-2 es un factor de proliferación de las células y también de las células B. Las células estimuladas simultáneamente por antígeno (MHC-IIepitope), por citocinas de las APC y por moléculas coestimulatorias de las APC (como la molécula B7), proliferan y producen otras citocinas que sostienen y expanden la respuesta inmunitaria. Algunas de las citocinas producidas por las células Th1, como las quimiocinas, atraen a otras células linfoides al sitio de reacción inmunitaria; otras, como la IL-2, estimulan la proliferación de las células reclutadas; otras más, como el interferón gamma (IFNγ), se activan macrófagos , y aun otras, como el factor de necrosis tumoral beta (TNFβ), son citolíticas. La actividad del IFNγ sobre los macrófagos induce en estas células la producción d citocinas y de metabolitos adicionales como el factor de necrosis tumoral alfa (TNFα) y el óxido nítrico, además de la sobreexpresión de moléculas MHC-II. Los macrófagos activados por las citocinas de tipo Th1, incrementan su metabolismo y su capacidad para matar microorganismos de todo tipo, pero sobre todo a aquellos de vida intracelular como los virus, algunas bacterias y algunos hongos y protozoarios. 28 En estudios sobre melanoma, se ha encontrado que la inducción de la inmunidad anti-tumoral sistémica optima involucra la sensibilización de células CD4+ y CD8+ especificas de antígenos asociados al tumor. Además de que las células CD4+ cooperadoras proveen señales reguladoras requeridas para la sensibilización de CD8+-HMC I, como para activar eosinófilos y macrófagos.18 Las células T CD4+ son requeridas para la generación y mantenimiento de las células T CD8+ citotóxicas y son esenciales para la generación tanto de la respuesta inmune anti-tumoral humoral y celular.16 LINFOCITOS T CITOTÓXICOS Dentro de las células T que proliferan en respuesta a la estimulación antigénica, hay algunas que adquieren propiedades citotóxicas. Las células T citotóxicas incluyen tanto a los linfocitos Tc/CD8+ como a los linfocitos Tc/CD4+. Cuando están activadas, estas células son capaces de matar a las células portadoras de antígenos extraños como por ejemplo algunas células tumorales . La muerte de las células blanco ocurre cuando menos a través de dos mecanismos generales: por necrosis o por apoptosis. 28 21 Necrosis En la necrosis, la célula citotóxica “inyecta” en la célula blanco el contenido de sus gránulos que incluye perforinas, granzimas y enzimas hidrolíticas, además de los radicales libres del oxígeno y el nitrógeno neoformados. Estos y otros factores externos lesionan indiscriminadamente a las células y ocasionan la ruptura de sus membranas, organelos y moléculas estructurales. Durante el proceso de muerte por necrosis las células dejan escapar su contenido citoplásmico, lo que estimula la acumulación de neutrófilos y la amplificación del daño tisular. Algunos de los factores causantes de muerte por necrosis son también inductores de apoptosis; dentro de estos factores están las granzimas, el peróxido de hidrógeno y el óxido nítrico, y el resultado parece depender de la dosis; las dosis más altas provocan la necrosis, mientras que las más bajas inducen apoptosis. 28 Apoptosis La apoptosis es un proceso de muerte más sutil que se induce por señales químicas derivadas del contacto con otras células o con substancias producidas por ellas (también pueden inducirse por señales originadas en el núcleo). Mientras que la mayoría de las células expresan en su membrana la molécula Fas (CD95), las células citotóxicas expresan el ligando de Fas (CD95L). La interacción FasFasL, causa la muerte de las células portadoras de Fas por activación del sistema de las caspasas. Las caspasas son enzimas dependientes de cisteína que escinden a otras caspasas en secuencias del extremo C-terminal que contienen residuos de aspártico y que de esta manera resultan activadas. Al recibir el estímulo apoptótico, las células inician un proceso de señalización que converge en las mitocondrias para promover la liberación de citocromo-c y de otros factores inductores de apoptosis (AIF, apoptosis inducer factor) al citoplasma; el citocromoc liberado interactúa con el factor activador de apoptosis (Apaf-1) y lo oligomeriza. El factor Apaf-1 oligomerizado activa a pro-caspasas iniciadoras de apoptosis (procaspasas-8 y -9), las cuales continúan el proceso de activación de cascada de otras caspasas. Alguna de las caspasas activadas (como la caspasa-3) alteran la estructura de la membrana de las mitocondrias, aumentan su permeabilidad y propician la liberación adicional de citocromo-c y AIF, lo que amplifica las señales de apoptosis. Las caspasas del final de la cascada, funcionan como nucleasas o activas a otras nucleasas, las cuales efectúan la hidrólisis del ADN. En esta etapa la célula comienza a mostrar los cambios morfológicos típicos de la apoptosis. 28 En las células apoptóticas la cromatina se condensa antes de escindirse en fragmentos que difieren entre sí en aproximadamente 200 pares de bases (pb), la membrana celular se contrae sin fragmentarse formando vesículas dentro de las cuales se empaca el contenido citoplásmico; como no hay ruptura de membranas, no hay escape del contenido celular y tampoco hay reacción inflamatoria. El sistema de caspasas promotoras de apoptosis está regulado por varias proteínas anti-apoptóticas; una de las más prominentes, la molécula Bcl-2, se encuentra asociada al superficie externa de las mitocondrias y evita la liberación de citocromo-c para estabilizar la membrana mitocondrial. 28 22 En estudios realizados en la línea celular U937 promonocitica humana, la cual es moderadamente susceptible a la infección por poliovirus. La infección de células U937 con poliovirus induce cambios morfológicos típicos de apoptosis. Y dispara la producción de óxido nítrico así como la activación de la pro-caspasa 3 y la polimerasa poli (ADP-ribosa) dirigiendo a la muerte celular.23 Respuesta inmune humoral Un rasgo fundamental de la inmunidad humoral es que la producción de todas las clases de anticuerpos se inicia por la interacción del antígeno con un numero pequeño de linfocitos B maduros que expresan inmunoglobulinas especificas para cada antígeno. Simultáneamente a la activación de las células Th1 mediadoras de la inmunidad celular, ocurre la activación de las células Th2. Como las primeras, las células Th2 responden a la estimulación antigénica y a las señales derivadas de las APC, proliferando y produciendo nuevas citocinas, dentro de las que se encuentran las interleucinas IL-4,IL-5,IL-6,IL-10 e IL-13. Algunas de estas citocinas y moléculas coestimulatorias presentes en los linfocitos B (como B7-1 o CD80 y B7-2 o CD86) participan en la activación de las células Th1 y en la transformación de las células B en células plasmáticas (CP), productoras de anticuerpos específicos. Ciertas citocinas como la IL-4, la IL-6 y el IFNγ modulan además la clase de anticuerpo producido por las CP. Los anticuerpos ejercen su función protectora cuando al unirse a un microorganismo lo inmovilizan y facilitan se eliminación por fagocitosis o su lisis por complemento. 28 Los anticuerpos no solo operan en el interior del cuerpo , también hay anticuerpos en la superficie de los epitelios mucosos de las tractos respiratorio y digestivo, en la conjuntiva, y en el tracto genitourinario. Aquí, los anticuerpos se producen en el tejido linfoide que se encuentra internamente asociado a las mucosas y se transportan a la superficie de las mismas acarreados por una proteína de transporte de origen epitelial. Aunque la secreción mucosa puede contener anticuerpos de todos los isotipos, los anticuerpos más abundantes son los de la clase IgA (seguidos por los de la clase IgM). 28 Los anticuerpos desempeñan además un papel primordial a. como receptores para antígeno de los linfocitos B, b. como reguladores de la respuesta inmunitaria y c. como depuradores de los autoantígenos que circulan en la linfa y en la sangre en cantidades diminutas al funcionar como opsoninas. La ausencia de anticuerpos en estos casos propiciaría a. la inmunodeficiencia humoral, b. la desregulación de la respuesta inmunitaria y el riesgo de hipersensibilidad, y c. la autoinmunidad. 28 En diversos estudios se ha encontrado que la vacunación con la vacuna de poliovirus inactivos (IPV) es capaz de inducir una fuerte respuesta de IgA de memoria en el suero de personas, que han sido expuestas de manera natural a el poliovirus de tipo silvestre. Esta vacunación, vía parenteral, fue capaz de favorecer la respuesta de IgA sistémica y mucosa en personas vacunadas con la vacuna de poliovirus oral (OPV). Donde la vacunación IPV es suficiente para 23 inducir una respuesta mucosa de IgA contra poliovirus y en los individuos sensibilizados con OPV mucosa, sin embargo, un refuerzo de la vacunación con IPV dirige a una fuerte respuesta de IgA en mucosas.7 Sin embargo existen muchos cánceres, entre ellos el melanoma, que pueden evadir o atenuar la respuesta inmunológica de diversas formas. Evasión Inmunológica La ineficiencia del sistema inmune para combatir el tumor puede estar asociada a múltiples fallas de los componentes del sistema inmune o de las células tumorales, las cuales se van acumulando durante la evolución de la enfermedad. Las células tumorales en etapas avanzadas utilizan cuando menos tres caminos diferentes para evadir la respuesta inmune: a) La modulación de la expresión de moléculas MHC, b) La modulación de otras moléculas de unión en la superficie celular, y c) La modulación de la secreción de citocinas celulares. Estos cambios inducen una depleción localizada de células inmunocompetentes y una disminución de su capacidad de respuesta a las linfocinas reguladoras, lo cual pudiera contribuir a la progresión tumoral. Si bien es cierto que las células tumorales presentan epitopos inmunogénicos, existen una serie de elementos que en general atenúan la respuesta inmunológica del huésped. En primer lugar, el carcinógeno responsable de iniciar el proceso maligno puede también atenuar la respuesta inmune. Segundo, la gran mayoría de los cánceres se originan en tejidos que no presentan con eficiencia algunos antígenos tumorales, de tal manera que en general hay insuficiente producción de factores coestimuladores. Esto puede llevar a que las células T se vuelvan tolerantes hacia los antígenos tumorales, lo cual atenúa severamente la respuesta inmune anti-tumoral. Tercero, aunque exista la presencia de determinantes antigénicos tumorales, éstos pueden ser subdominantes, crípticos, o bien poco inmunogénicos. 1,4,28,29 Actualmente, la mayoría de las formas clínicas del cáncer no tienen cura y más aún, en la gran parte de los casos representan un gran problema de salud pública, tanto en los países desarrollados, así como en aquellos en vías de desarrollo. La propuesta curar el cáncer o al menos tratarlo de manera individual, evitando los efectos sistémicos indeseables de las terapias convencionales. mediante terapia génica se fundamenta en sus orígenes moleculares. 28 Estudios en diversas líneas de melanoma, la existencia de mutación en lagunos genes supresores de tumores como p53, p21, INK4a/p16, INK4a/p14AR, INK4a/p15. Además la familia del oncogen ras ha sido implicada en la patogénesis de melanoma a causa de la alta frecuencia de genes ras activados en melanoma, predomiantemente N-ras. Por último se ha visto la expresión masiva de c-myc, el cual, es capaz de incrementar la tumoriginicidad y el potencial metastásico de algunas líneas de melanoma. Todos ellos participan en la evasión inmunológica del melanoma.11,30 24 Terapia Génica. El concepto de terapia génica resulta de la observación de que ciertas enfermedades, entre ellas particularmente el cáncer, resultan de daños genéticos. Los daños genéticos involucrados se pueden agrupar en tres grandes categorías: ¯ La activación de genes dominantes (proto-oncogenes oncogenes). ¯ La pérdida de genes supresores de tumores. ¯ La supresión local de la respuesta inmune. Concretamente el término terapia génica unifica los principios de la farmacología con los de la genética, pues implica el empleo de ácidos nucleicos como el agente farmacológico para el tratamiento de estados patológicos.1 La introducción del gene terapéutico puede realizarse directamente en las células tumorales, llamada terapia génica in vivo, o en las células extraídas del paciente que son reinoculadas después de la manipulación genética, llamada terapia génica ex vivo. Normalmente se necesita un vector , que es el elemento que permite introducir en la célula el material genético que pretendemos.11 El vector ideal que por el momento características: no existe debería reunir una serie de ¯ Permitir la incorporación y expresión regulada durante el tiempo conveniente de uno o más genes necesarios para la aplicación clínica requerida. ¯ Ser específico en su transferencia génica para la célula blanco. ¯ Ser estable y fácil de obtener. ¯ Ser irreconocible por el Sistema Inmune y no inducir respuesta inflamatoria. 11 Existe una enorme variedad de posibles aplicaciones de este tipo de terapia, pues ésta tiene el potencial de corregir defectos genéticos, combatir infecciones o destruir tumores. 31 La elección de la estrategia de terapia génica anti-neoplásica depende del método que se desea emplear para destruir a las células tumorales. Según el método seleccionado, las células malignas pueden ser reconvertidas a un fenotipo no maligno, o ser eliminadas por el sistema inmune, o por inhibición de la neovascularización tumoral, o por efectos tóxicos directos.24 a) Citotóxicidad condicionada. Esta modalidad también se conoce como terapia génica suicida y consiste en la administración de genes tóxicos para eliminar células tumorales. Estos genes codifican para enzimas que convierten una por-droga inocua en un agente tóxico. 25 b) Corrección fenotípica o compensación de mutaciones. Consiste en la adición de genes y en el uso de moléculas antisentido, para anular el fenotipo maligno. Esto se logra mediante la sobreexpresión de genes supresores de tumores ola inactivación de oncogenes involucrados en la progresión tumoral. c) Inhibición de la neoangiogénesis tumoral. Se están estudiando moléculas y mecanismos que bloquearían la perfusión sanguínea de la neoplasia y provocarían una inhibición persistente del crecmiento tumoral. Así como tratamientos para alteraer la neovasculatura tumoral tendrían un efecto localizado y probablemente no afectarían a los tejidos normales. d) Quimioprotección. Consiste en la administración de genes de resistencia a drogas para proteger la médula ósea de la mielosupresión inducida por quimioterapia. En esta estrategia se introducen genes que incrementan la resistencia a medicamentos. e) Terapia potenciadora. Combinada con otras modalidades complementarias, la terapia génica puede jugar un papel preponderante en el tratamento antitumoral. Tal es el caso de la quimiosensibilización y la radiosensibilización. En la primera, se combinan agentes quimioterapéuticos convencionales y la quimioterapia molecular. En la segunda, se combinan esquemas de terapia suicida más radioterapia, puede lograr un tratamiento anti-tumoral más eficaz. f) Inmunoterapia. Se basa en el control del crecimiento tumoral mediante la activación del sistema inmunológico. La activación de una respuesta antitumoral específica será consecuencia de las expresión por las células tumorales de nuevos antígenos, denominados tumor-específicos o tumorasociados. La inmunoterapia, busca la estimulación de la respuesta inmune del propio paciente produciendo un estímulo lo suficientemente potente que active la respuesta del sistema inmunológico en forma de anticuerpos séricos, proteínas circulantes que se unen al antígeno inactivándolo o destruyendo la célula tumoral que posee dicho antígeno, o respuesta celular basada en los linfocitos y células de la inflamación que producen una respuesta de células inmunitarias contra las células tumorales. 29,30,31 Para ello se han ensayado o están en curso de estudio, diferentes modalidades desde la utilización de células tumorales enteras inyectadas al paciente, generalmente con sustancias adyuvantes (capaces de potenciar inespecíficamente la respuesta inmunológica) 6 y procedentes del propio paciente o de otros, extractos de células tumorales lisadas o los antígenos purificados si se conocen o incluso péptidos, incluyendo superantigenos.20 26 Además existen muchos estudios pre-clínicos que han demostrado que las inmunizaciones basadas en DNA contra modelos de antígenos tumorales tales como la albumina, β-galactosidasa, péptidos (IL-2),19 células manipuladas para secretar diferentes factores (GM-SCF)32, y diversidad de terapias que utilizan diversos al interferón.34 La inmunoterapia se utiliza como tratamiento complementario en fases de remisión o cuando aparece un tumor difícilmente eliminable por otros procedimientos. Puede hacerse bajo diferentes enfoques terapéuticos: ¯ La inmunoterapia adoptiva, mediante la administración de anticuerpos monoclonales. ¯ La inmunoterapia pasiva, donde se intenta la estimulación de los componentes del sistema inmunitario responsables de la respuesta antitumoral y donde quedaría englobada la administración de las vacunas (inmunización activa específica) y de los interferones, interleucina-2 y BCG (inmunización activa inespecífica).4 Para potenciar la capacidad inmunológica de los antígenos, la introducción de los genes responsables de su codificación se realizará en el interior de vectores víricos o bacterianos. La utilización de virus, alterados genéticamente, como vectores, se caracteriza por la ausencia de tropismo tisular, la rápida inactivación por los anticuerpos cuando se administran sistemáticamente y un tamaño reducido que se asocian a una escasa carga antigénica que puede limitar la eficacia clínica del tratamiento. 4 El principal problema de esta metodología es conseguir que todas las células tumorales se encuentren infectadas por estos vectores. Si alguna célula tumoral escapa de la infección presentará una mayor capacidad de supervivencia y podrá general líneas celulares resistentes. Por ello los tratamientos deberán estar basados en esquemas terapéuticos que requerirán al administración repetida de los vectores, de forma similar a los actuales tratamientos radioterápicos y quimioterápicos fraccionados. 4 Propiedades Inmunogénicas De Las Proteínas Vp1 Y Vp4. La vacunación contra los poliovirus es capaz de generar inmunidad humoral. Numerosos estudios han mencionado un papel para las células T en la inmunidad de los poliovirus a través de su función colaboradora en la producción de anticuerpos. 18 La mayoría de la información acerca de los determinantes antigénicos involucrados en la neutralización de los poliovirus ha derivado de estudios conducidos por anticuerpos monoclonales generados por ratones inmunizados vía parenteral. 5 27 Los anticuerpos específicos son capaces de neutralizar los virus y son considerados para ser el principal mecanismo de protección contra la infección por poliovirus. Además, muchos estudios han sido llevados a cabo para definir las estructuras antigénicas involucradas en la neutralización de los poliovirus y han resultado en la identificación de varios epitopos de neutralización. Los cuatro sitios antigénicos principales para la neutralización han sido identificados en las 3 proteínas principales de la cápside de los tres serotipos de poliovirus. El sitio 1 esta compuesto de residuos de VP1, mientras las estructuras de los otros sitios antigénicos parecen ser más complejas, involucrando residuos de VP1, VP2, y/ o VP3. Se sabe que estos inducen una respuesta proliferativa específica de células T CD4+.5,18 Análisis específicos contra las proteínas de poliovirus (los tres serotipos) purificadas, demuestran que las clonas de las células T reconocen determinantes en las proteínas superficiales de la cápside: VP1, VP2 y VP3 y en la proteína interna de la cápside VP4 24 La caracterización inmunológica de los linfocitos T específicos de poliovirus indica que la población efectora involucrada en la respuesta proliferativa especifica de poliovirus observada fue de las células T colaboradoras CD4+. La respuesta proliferativa también fue inducida dentro de estas poblaciones de linfocitos T bajo estimulación con la proteína VP1 o con péptidos sintéticos de la misma. Al usar péptidos sintéticos, varios epítopes de linfocitos T colaboradores fueron identificados. 21 Generalmente, la respuesta proliferativa fue observada en 3 regiones de la proteína VP1. El análisis usando péptidos sintéticos de las secuencias descubiertas indican que la región abarca los residuos 201 a 241 puede contener varios epitopes para los linfocitos T y puede contar para la fuerte respuesta proliferativa observada. 21 La respuesta proliferativa de las células T hacia los poliovirus en varios linajes de ratones ha sido analizada por uso de virus muertos purificados o péptidos sintéticos de la proteína de la cápside VP1. Seguida de la inmunización de ratones con el poliovirus tipo 1 inactivado, una respuesta proliferativa de sus células CD4+ en nódulos linfáticos fue obtenida después de la estimulación con el virus purificado. En ratones inmunizados con poliovirus tipo 1,2 y 3, una fuerte reactividad cruzada de la respuesta de células T fue observada antes in vitro con la estimulación con virus hetérologos. Un epitope dominante para las células T fue identificado en la región 103-115 aminoácidos del polipéptido VP1 de la cápside muy cerca del epitope de neutralización C3. La inmunización de ratones BALB/c (H-2d) con el péptido p86-115 sin acoplar, el cual representa los aminoácidos 86 a 115 de VP1 y contiene tanto el epitopo de las células T como el epitopo de neutralización C3, induce respuesta de células T y células B especificas de poliovirus. Además, estos ratones desarrollaron anticuerpos que neutralizan a los poliovirus. El sitio antigénico 1 del poliovirus tipo 1 inactivado contiene un 28 importante epitopo para células B por lo que estos ratones desarrollan anticuerpos que neutralizan a los poliovirus. 22 El análisis específico contra las proteínas purificadas de los poliovirus demuestra que las clonas de las células T reconocen determinantes en las proteínas de la superficie de la cápside VP1, VP2 y VP3 y de la proteína interna de la cápside VP4. Los epitopes en VP4 fueron conservados entre los serotipos y esto puede contar para reactiviadad–cruzada observada con la población de las células T policlonal. En contraste, las clonas de las células T que reconocen epitopes en VP1 o VP3 fueron grandemente especificas de serotipos, con simples o múltiples sustituciones de aminoácidos fueron encontradas para ser criticas en el reconocimiento por las células T. 24 Por lo anterior, podemos decir que al utilizar las proteínas VP1 y VP4, como antígenos en las células tumorales podría inducir una fuerte respuesta inmunológica, gracias a la presencia de determinantes antigénicos en estas proteínas, que reconocen las células T. En el Departamento de Biología Molecular y Biotecnología de la UNAM se realizará un proyecto basado en la inmunoterapia, en el cual se tratará de iniciar una respuesta inmune, por medio de la introducción de genes, que codifican proteínas (VP1 y VP4) del virus de poliomielitis humana clonados en el vector pDISPLAY. Estos plásmidos serán inyectados en tumores para inducir la expresión de las proteínas de forma transmenbranal en las células tumorales, mismas que el sistema inmune reconocerá como extrañas y eliminara; para así inducir regresión del melanoma. 29 OBJETIVO GENERAL Determinar el efecto que tienen los antígenos del virus de la poliomielitis humana sobre las células cancerosas. OBJETIVOS PARTICULARES Ø Aislar las secuencias génicas codificadoras para las proteínas VP1 y VP4 del virus de la poliomielitis humana por restricción enzimática. Ø Clonar las secuencias aisladas en los sitios Bgl ll y Sal l del vector pDISPLAY. Ø Transformar bacterias E.coli (cepa JM101) con cada uno de los plásmidos construidos . Ø Extraer los plásmidos por la técnica de lisis alcalina. Ø Analizar los vectores recombinantes por restricción enzimática y PCR. Ø Establecer el stock de los vectores recombinantes . Ø Vacunar a los animales en su etapa infantil con una vacuna génica contra la polio. Ø Inducir tumores con las células B16FO. Ø Tratar los tumores con los plásmidos pDISVP1, pDISVP4, pVAXVP1 y pVAXVP4. Ø Medir el efecto del tratamiento sobre las masas tumorales. 30 METODOLOGÍA SUBCLONACIÓN DE LAS PROTEÍNAS VP1 Y VP4 DEL PLÁSMIDO pCVP1 Y pCVP4 RESPECTIVAMENTE. Los genes que codifican para las proteínas VP1 y VP4 del virus de la poliomielitis humana fueron aislados por restricción enzimática con Bamh I y Xho I a partir de los plásmidos pCVP1 y pCVP4 respectivamente. Los fragmentos génicos obtenidos fueron subclonados en los sitios Bgl II y Sal I del vector pDISPLAY, el cual contiene una secuencia que codifica para Cultivo De Bacterias E. Coli Inicialmente se inoculó una alícuota de bacterias E. coli. (cepa JM101), que contenia el plásmido pCVP1, en un tubo con 5ml de medio LB con 20µl del antibiótico ampicilina y el crecimiento celular se logró a 250rpm, 37°C, durante toda la noche. Purificación Del DNA Plasmídico (Procedimiento De Minipreparación), Mediante Lisis Alcalina. Después de haber obtenido el cultivo bacteriano, se centrífugo a 5000g durante 8 minutos, el DNA plasmídico se aisló resuspendiendo los botones bacterianos, en solución glucosada [ 50Mm TRIS-HCL (pH 8.0), 10mM EDTA (pH 8.0)], a la cual se adicionó una porción pequeña de lisozima sólida, para favorecer la lisis celular. A este volumen se le adiciono 200µl de esta solución se mezcló por agitación invertida 5 veces y se incuban en hielo durante 5 minutos. Se le adicionó 400µl de solución 0.2M NaOH/15(w/v)SDS, e inmediatamente se mezcló su contenido invirtiendo los tubos rápidamente. Para precipitar las proteínas y el DNA genómico, se adicionaron 300µl de solución de potasio 5M ( pH 5.1) a cada tubo, Para dispersar el acetato de potasio se removieron los tubos fuerte y rápidamente, hasta que se observo un lisado bacterial viscoso y después de este lisado, los tubos fueron incubados en hielo durante 10 minutos, para después centrifugar a 12000g durante 5 minutos. El sobrenadante que contenía el plásmido se transfirió tubos nuevos y estériles y se les adiciono 500µl de isopropanol y se incubaron en hielo durante 10minutos. Los tubos se centrifugaron a 12000g durante 5 minutos, el sobrenadante se desecho y los botones se resuspendieron en una mezcla de 200µl de Buffer TE + 100µl de acetato de amonio 7.5 M por tubo. Para precipitar el RNA, los tubos se incubaron en hielo durante 20 minutos y se centrifugaron a 12000g durante 5 minutos y el sobrenadante se transfirió a tubos nuevos. Posteriormente se adicionaron 600µl de etanol al 95 %, se mezclaron por agitación y los tubos se incubaron a -70°C durante 1 hora. Después de la incubación , los tubos se centrifugaron a 1200g durante 15 minutos y los botones resultantes se resuspendieron en 30µl de H2O guardándose a -20°C hasta su uso posterior. Se realizó el mismo procedimiento para cada uno de los plásmidos y para el vector. 31 Restricción Enzimática. Para clonar el fragmento VP1, fue necesario por una parte, digerir el fragmento de pCVP1 en los sitios BamH I y Xho I, y por otro lado, fue necesario linearizar el vector pDISPLAY por restricción enzimática en los sitios Bgl II y Sal I, para poder clonar a VP1. La restricción enzimática se llevó a cabo, utilizando 2 µl de cada enzima (10U/µl),10 % de Buffer de reacción, para un volumen final de 20µl de reacción. Electroelusión Del DNA Con el fin de purificar bandas de DNA de tamaños específicos, se realizó una electroforesis en gel de agarosa al 1%, utilizando100µl del volumen de reacción de la restricción enzimatica. Puesto que se trataba de 100µl de volumen, fue necesario fusionar varios pozos para colocar la muestra. Después de 45 minutos de corrimiento electroforético, la banda de interés fue extraída del gel con un bisturí y se colocó, en una membrana de diálisis, a la que se adicionó 1ml de buffer de electroelusión (TAE 1%). El DNA se aisló de la agarosa, por medio de un nuevo corrimiento electroforético a 100 Volts, voltaje constante durante 30 minutos (método de electroelusión). Después de éste tiempo, la membrana fue observada en el transiluminador (emisión UV), en donde se pudo verificar que el DNA se encontraba en el buffer de electroelusión y no en la agarosa. El DNA se purificó usando la técnica de extracción con fenol-cloroformo y precipitación por etanol. PURIFICACIÓN DE DNA POR EXTRACCIÓN FENOL-CLOROFORMO Y PRECIPITACIÓN POR ETANOL (Modificación de Maniatis et. Al, (1989)). En un volumen igual de fenol-cloroformo (1:1), se adiciona la muestra de DNA (aislado por electroelusión) en un tubo de polipropileno. El contenido del tubo se agitó hasta que se formó una emulsión y los tubos se centrifugaron a 12 000g durante 5 minutos. El ácido nucléico estaba contenido en la fase acuosa que se formó en la parte superior, mientras la fase orgánica permaneció en la parte inferior. La fase acuosa se transfirió a un tubo nuevo y limpio con un volumen igual de cloroformo + alcohol isoamílico y las fases se mezclaron agitando los tubos y posteriormente centrifugando a 12000g durante 10 segundos. El DNA que estaba contenido en la fase acuosa (fase superior) nuevamente se transfirió a un tubo nuevo y limpio. El DNA se precipitó con 750µl de etanol. El contenido se mezcló rápidamente por inversión de los tubos de 4 a5 veces y posteriormente se incubaron a -70°C durante 1 hora. El DNA se centrífugo a 12000g durante 15 minutos. Se decantó el sobrenadante y los botones se resuspendieron en agua ultra pura. 32 REACCIONES DE LIGACIÓN La concentración de DNA fue medida por espectrofotometría, a 260nm aplicando la siguiente formula. 1 A 260 Unit de DNA DOBLE = 50µg (OD 260NM)(Factor de Dilución)(50)= µg/ml Factor de Dilución 4µl DNA EN 1000µL DE H20= 1000/4= 250 Se hizo una reacción 1:3, es decir, usando 100ng de vector digerido contra 300 ng de inserto, para un volumen final de 20µl de reacción. Se utilizaron 2 unidades de la enzima T4 DNA ligasa (1U/µl, Gibco BRL) y 20% del volumen de reacción del buffer T4 DNA ligasa. Las reacciones de ligación se incubaron 1-2 semanas a 16°C y después de la incubación se hizo transformación bacteriana, adicionando las células directamente a 10µl de la reacción de ligación. TRANSFORMACIÓN CELULAR, UTILIZANDO CÉLULAS E. coli DE LA CEPA BACTERIANA JM101 Las células competentes se almacenaron en hielo y solo 100µl de éstas fueron incubadas con 10µl de la reacción de ligación (pDISPLAY e inserto), en condiciones de esterilidad. Las muestras se incubaron en hielo durante 30 minutos y posteriormente la transformación de las células bacterianas fue llevada a cabo por tratamiento de choque térmico a 42°C durante 2 minutos. Inmediatamente después se adicionaron 500µl de medio LB (Extracto de levadura, Peptona de caseína y Cloruro de Sodio estéril, sin antibiótico ) y los tubos fueron incubados 1 hora a 37°C con agitación constante. SIEMBRA DE LAS CÉLULAS TRANSFORMADAS Después de la incubación, en una caja Petri con 20ml de medio LB y 1.5% de agar bacteriológico, mas 20µl de ampicilina, se colocó 300µl del volumen total de la mezcla, para luego dispersarlo por toda la superficie de la caja. La caja de Petri se incubó a 37°C durante toda la noche en posición invertida. Una vez obtenidas las colonias celulares, se colectaron 10 clonas y se hizo una reacción de PCR, utilizando directamente las bacterias para corroborar la positividad de las colonias recombinantes con la fracción génica de VP1 o VP4. 33 MASTER PLATE En condiciones de esterilidad se adiciona una gota de cultivo de cada colonia a una caja petri con LB-ampicilina que anteriormente fue divida en 10 partes . Sen incuba a 37°C toda la noche, se sella con parafilm y se guarda a 4°C para su uso posterior. Se eligieron las clonas recombinantes más representativas para purificar el DNA plasmídico mediante la técnica de miniprep, con la cual se obtuvo un DNA para hacerles RESTRICCIÓN ENZIMÁTICA y PCR AMPLIFICACIÓN DE LA SECUENCIA CODIFICADORA DE LAS PROTEÍNAS VP1 Y VP4 DEL VIRUS DE LA POLIOMIELITIS. El Marco de Lectura Abierto de interés (proteínas VP1 y VP4) fueron amplificadas por PCR utilizando 2 unidades de Taq DNA polimerasa (PerKin Elmer), 2µl de buffer PCR 10X.25mMde cloruro de magnesio (MgCl2), 10µM de cada uno de los oligonucleótidos, 2mM de dNTPs y 1µl de DNA genómico, previamente aislado. La reacción de PCR fue realizada en una reacción de 20µl totales por tubo, en 30 ciclos de amplificación (95°C, 30 segundos;40°C, 60segundos; 72°C, 90 segundos). El oligonucleótido sentido: VP1 F( 5’ C GAA TTC ATG GGG TTA GGT CAG 3’ ) y el antisentido: VP1 R ( 5’ C CTC GAG GTG TCC GAA TCC ATA 3’ ) y el oligonucleótido sentido: VP4 F( 5’ C GAA TCC ATG GGT GCT CAG GTT 3’ ) y el antisentido: VP4 R ( 5’ C CTC GAG GTT TAG CAT TGG GGC 3’ ), utilizados para realizar la reacción de PCR fueron diseñados con base en la secuencia prototipo del genoma de el virus de poliomielitis, reportada en el banco de datos GenBank. STOCK EN GLICEROL DE CÉLULAS BACTERIANAS TRANSFORMADAS Una vez confirmada la positividad de las colonias recombinantes mediante restricción enzimatica, reacción de PCR, se hizo un stock en glicerol de las clonas más representativas. Esto evito transformar bacterias cada vez que se requería el plásmido recombinante. Para vello de utilizaron tubos de crioconservación en donde se colocaron 850µl de células transformadas con cada uno de los plásmidos, Una vez adicionadas las células se adicionaron 150µl de glicerol estéril. Los tubos se almacenaron a -70°C hasta el momento de su uso. DESCONGELAMIENTO DE LAS CÉLULAS B16FO Se descongeló 1ml de células de mamífero B16FO contenidas en un microvial a 70°C en Η2Ο a 37°C, agitando hasta disolver. Se transfirió 1 ml de las células a dos tubos eppendorf estériles (500µl cada uno) + 1ml de medio DMEM, se centrifugaron a 25000rpm durante 4 minutos. Se desechó el sobrenadante y 34 resuspendió el botón en 1 ml de medio DMEM nuevo. Se transferir todo el contenido de ambos tubos a una caja Petri, se incubó a 37°C con 5% de CO2 toda la noche. CAMBIO DE MEDIO Para cambiar el medio, se desechó 1ml del medio que contenía la caja Petri, se agregó medio nuevo y se incubó a 37°C con 5% de CO2 hasta que las células ocuparon toda la superficie de la caja para hacer resiembra. RESIEMBRA Se retiró todo el medio (2ml) y se lavó dos veces con PBS, se incubó con 2ml de Tripsina (1ml de Tripsina en 13 ml PBS) a 37°C durante 3 minutos. Se transfirió 1ml de las células a un tubo eppendorf nuevo y el otro ml a otro tubo para neutralizar la Tripsina se agregó 300µl de medio DMEM nuevo, se centrifugaron a 2500rpm durante 4 minutos, se desechó sobrenadante y el botón de cada tubo se resuspendió en 1ml de medio DMEM nuevo. Se sembró todo el contenido de los dos tubos en una caja Petri estéril, se incubo a 37°C con 5% de CO2 toda la noche. CONTEO DE CÉLULAS Se retira todo el medio (2ml) de la caja Petri y se lavó dos veces con PBS, se incubo con 2ml de Tripsina (1ml de Tripsina en 13 ml PBS) a 37°C durante 3 minutos. Se transfirió 1ml de las células a un tubo eppendorf nuevo y el otro ml a otro tubo para neutralizar la Tripsina se agregó 200µl de medio DMEM nuevo, se centrifugaron a 2500rpm durante 4 minutos, se desechó sobrenadante y el botón de cada tubo se resuspendió en 1ml de medio DMEM nuevo. El contenido se dividió en 4 tubos: 3 tubos se congelaron a –70 °C y el último se resembró y se contó. En dos cajas Petri con 5 ml de medio DMEM se agregó 500µl de células a cada una y se incubaron a 37°C con 5% de CO2 toda la noche. VACUNACIÓN DE LOS ANIMALES Para este trabajo se utilizaron ratones hembra de la cepa C57Bl/6 de 6 semanas de vida. Un grupo de animales fue vacunado con una vacuna génica contra el virus de la poliomielitis humana previamente construida en nuestro laboratorio, por la vía oral. Otro grupo recibió solución salina estéril (PBS) por la misma vía, utilizando una sonda intragástrica. Los animales recibieron 2 dosis de 100µg, de la vacuna génica o 100µl de la solución salina estéril a intervalos de 2 semanas. 35 INDUCCIÓN DE TUMORES Para la inducción de tumores en ratones se usaron células B16FO, derivadas de un melanoma de ratones C57Bl/6. 50,000 x 105 células fueron inyectadas intradérmicamente en el costado derecho de cada animal. La inducción de los tumores se llevó a cabo 12 meses después de que los animales fueron vacunados contra la polio. El desarrollo de los tumores fue monitoreado cada dos días. TRATAMIENTO DE TUMORES Cuando los tumores alcanzaron un área de mm2 se realizó su tratamiento. Los tumores fueron inyectados directamente con 100µg de pDISVP1+ 100µg de pDISVP4, utilizando para este fin una jeringa de tuberculina y procurando depositar los plásmidos en el centro de la tumoración. Grupos control de animales recibieron 100µg de los plásmidos pVAXVP1 y pVAXVP4, los cuales expresan a VP1 y VP4 pero no la presentan en la membrana celular. Otro grupo de animales recibió PBS en sus tumores. Finalmente un último grupo fue dejado sin tratar. ANÁLISIS ESTADÍSTICO Los datos obtenidos serán analizados por la prueba t de student. Se considerará un nivel de significancia básico para p= 0.005 con un intervalo de confianza de 95%. 36 RESULTADOS OBTENCIÓN DE LOS FRAGMENTOS GÉNICOS VP1 Y VP4. Los genes de las proteínas VP1 y VP4 del virus de la poliomielitis humana están presentes en la región de policlonación de los vectores pCVP1 Y pCVP4 respectivamente (figura 8). VP Figura 8. Las fracciones génicas de VP1 y VP4 están clonadas en la región de policlonación del vector, en los sitios de EcoRI y Xho I. La figura 8, esquematiza el vector utilizado para la obtención de cada secuencia desde la región de policlonación, es decir, que hay un vector para extraer cada secuencia génica. 37 pCVP1/BamH I, Xho I DNA Ladder 100pb Los fragmentos génicos que codifican para las proteínas VP1 y VP4 fueron aislados por restricción enzimática con BamH I y Xho I de los vectores pCVP1 Y pCVP4 respectivamente. La digestión de pCVP1 produjo la liberación de un fragmento de 917pb (figura 9), el cual corresponde con el peso esperado para el gene VP1. Se hizo una electroforesis en gel de agarosa para visualizar el DNA previamente aislado (miniprep). 5.4Kb (VECTOR) 1500pb 1000pb 917pb (VP1) 600pb Figura 9. DNA de VP1 aislado de la bacteria E. coli., digerido con BamH I Y Xho I. G el de agarosa 1.0% En la figura 9, se observa el fragmento BamH I Y Xho I después de haber sido liberado del vector de expresión. Hay una banda de aproximadamente 5.4Kb que corresponde al tamaño del vector y otra banda de 917pb que corresponde al tamaño del inserto VP1. 38 DNA Ladder 100pb pCVP4 NOcortado pCVP4/BamH, xho I Por otra parte, la digestión de pCVP4 liberó un fragmento de 833pb (figura 10), el cual corresponde con el peso esperado para el gene VP4. Se hizo una electroforesis en gel de agarosa para visualizar el DNA previamente aislado (miniprep) 5.4Kb (VECTOR) 1500pb 600pb 200pb 205pb (VP4) Figura 10. DNA de VP4 aislado de la bacteria E. coli., digerido con BamH I Y Xho I. Gel de agarosa 1.0% En la figura 10, se observa el fragmento BamH I Y Xho I después de haber sido liberado del vector de expresión. Hay una banda de aproximadamente 5.4Kb que corresponde al tamaño del vector y otra banda de 833pb que corresponde al tamaño del inserto VP4. 39 PREPARACIÓN DEL VECTOR pDISPLAY VISIÓN GENERAL DEL VECTOR pDISPLAY El vector pDISPLAY se muestra en la figura 11, donde se observa la región de policlonación. La cual fue una guía importante para establecer los sitios de restricción para linealizar el vector y los sitios en los cuales se ligaría cada una de las secuencias génicas de VP1 y VP4 en dicho vector [ver figura 13 y 14 respectivamente]. Figura 11. Esquema general del vector pDISPLAY, así como la representación de la región de policlonación del mismo. El vector utilizado contiene: ¯ El promotor Citomegalovirus humano (pCMV) permite la traducción en las células de mamífero. ¯ El promotor T7 permite la transcripción. ¯ La secuencia de la cadena-κ de Ig murina permite que la proteína clonada sea secretada. ¯ El epitope A de hematoglutinina (HA) induce la detección de nuestra proteína por los anticuerpos monoclonales. ¯ Una región de policlonación permite la inserción de nuestro gene y facilita su clonacióin (Sfl I- Acc I). 40 ¯ El epitope myc permite la detección de pDISVP con los anticuerpo Antimyc. ¯ El domino transmembranal del recertor del factor de crecimiento derivado de plaquetas (PDGFR-TM) ¯ La señal de poliadenilación de la hormona de crecimiento bovina (BGH) ¯ El origen pUC induce replicación y crecimiento en E. coli. ¯ La señal de poliadenilación (TkpA) permite la terminación de la transcripción y la maduración del mRNA del gnen de resistencia Kanamiciana. ¯ El origen y promotor SV40 permite la expresión del gene de resistencia Kanamicina/Neomicina en las células de mamífero además induce la replicación en las células que contengan el antígeno + de SV40. ¯ El gene de resistencia a Ampicilina (β-lactamasa) permite la selección de las colonias recombinantes en E. coli. ¯ El origen f1 permite el rescate del DNA de una sola cadena. 41 pDISPLAY/ Bgl II y Sal I DNA Ladder 100pb El vector pDISPLAY fue digerido usando las endonucleasas de restricción Bgl II y Sal I, el producto de la digestión puede verse al observar la banda de ∼5.4Kb (figura 12). El vector fue previamente aislado de la bacteria E. coli. 5.3Kb 1500pb 600pb Figura 12. Restricción enzimática del vector pDISPLAY con las enzimas Bgl II y Sal I. Gel de agarosa 1.0% En la figura 12, se observa el vector de expresión linearizado después de haber sido digerido con las enzimas Bgl II y Sal I. Hay una banda de aproximadamente 5.3Kb que corresponde al tamaño del vector. 42 SUBCLONACIÓN DE LOS GENES VP1 Y VP4 SUBCLONACION DEL GENE VP1 La secuencia génica de la proteína VP1 del virus de la poliomielitis humana fue ligada en los sitios Bgl II y Sal I de la región de policlonación del vector pDISPLAY, para generar el plásmido pDISVP1 (figura 13). VP1 Figura 13. Clonación del fragmento génica VP1 en el vector de expresión pDISPLAY 43 SUBCLONACIÓN DEL GENE VP4 La secuencia génica de la proteína VP4 del virus de la poliomielitis humana fue ligada en los sitios Bgl II y Sal I de la región de policlonación del vector pDISPLAY, para generar el plásmido pDISVP4 (figura 14). VP4 Figura 14. Clonación del fragmento génica VP4 en vector de expresión pDISPLAY 44 ANÁLISIS DE LAS COLONIAS RECOMBINANTES POR RESTRICCIÓN ENZIMÁTICA. pDISVP1/Xba I (clona 4) pDISVP1 NO cortado (clona 2) pDISVP1/Xba I (clona 2) pDISVP1 NO cortado (clona 3) pDISVP1/Xba I (clona 3) pDISVP1 NO cortado (clona 4) DNA Ladder X pDISVP1 NO cortado (clona 1) pDISVP1/Xba I (clona 1) ANÁLISIS PARA VP1 Para determinar la presencia de la secuencia génica VP1 en el plásmido de las colonias recombinantes,se analizaron por restricción con la enzima de restricción Xba I. Esta enzima corta sólo al inserto (VP1), en el sitio 2546 y 2861pb por lo que se espera 1 banda representativa de la digestión con el siguiente tamaño; una banda de 315pb (figura 15). Si el fragmento de 315pb es liberado, significa que las colonias son recombinantes, es decir, el gene de la proteína VP1 esta clonado. En cambio si el fragmento de 315pb no es liberado, por lo tanto las colonias no son recombinantes. En la figura 12 se muestra la electroforesis de la restricción con la enzima Xba I, donde se observa el fragmento esperado de 315pb solo en la colonias 1-3, lo cual indica que estas colonia son recombinantes. 1018pb 517pb 315pb 298pb Figura 15. DNA de las colonias (1-4) E.coli, cepa JM101, digerido con la enzima Xba I. Gel de agarosa 1.0% En la figura 15, se observa una banda de 315pb en las colonias 1-3, lo cual indica que son recombinantes. 45 pDISVP1/Xba I (colonia 6) pDISVP1 NO cortado (colonia 7) pDISVP1/Xba I (colonia 7) pDISVP1 NO cortado (colonia 8) pDISVP1/Xba I (colonia 8) pDISVP1 NO cortado (colonia 9) pDISVP1/Xba I (colonia 9) pDISVP1 NO cortado (colonia 10) pDISVP1/Xba I (colonia 10) DNA Ladder X pDISVP1 NO cortado (colonia 5) pDISVP1/Xba I (colonia 5) pDISVP1 NO cortado (colonia 6) En la figura 16, se observa una banda de 315pb en las colonias 4-5 y 7-10, después de haber sido liberada del DNA de la bacterias E.coli (cepa JM101),el cual fue digerido con Xba I, lo cual indica que estas colonias son transformadas, es decir, presentan el vector recombínate pCVP1. 1018pb 517pb 315pb 220pb Figura 16. DNA de las colonias 5-10, digerido con la enzima Xba I. Gel de agarosa 1.0% 46 ANÁLISIS DEL DNA DE LAS COLONIAS RECOMBINANTES PCR VP1 (clona6) PCR VP1 (clona7) CONTROL (-) DNA Ladder X CONTROL (+) Para corroborar la presencia de la secuencias génica VP1 en el plásmido, de las colonias (6 y 7), se realizó una reacción de PCR. El producto de la reacción de PCR fue analizado mediante un corrimiento electroforético en un gel de agarosa al 1.0% y visualizado en un equipo de documentación de geles(figura 17). 4070pb 1018pb 915pb (VP1) 517pb Figura 17. Secuencia génica VP1, amplificada por PCR. Gel de agarosa 1.0% En la figura 17, se observa una banda de 915pb, que corresponde a la amplificación del gene que codifica a la proteína VP1 del virus de la poliomielitis humana, comparada con una banda de 915 (control positivo [+]) que corresponde al vector pXpA que contiene el genoma completo del virus de la polio. Al amplificar el fragmento génico VP1, pudimos estar completamente seguros que el gene había sido clonado. 47 ANÁLISIS POR RESTRICCIÓN ENZIMÁTICA PARA VP4 pDISVP4/Sma I (colonia 6 ) pDISVP4 NO cortado (colonia 7 ) pDISVP4/Sma I (colonia 7) pDISVP4 NO cortado (colonia 6 ) pDISVP4/Sma I (colonia 4) pDISVP4 NO cortado (colonia 5 ) pDISVP4/Sma I (colonia 5 ) pDISVP4 NO cortado (colonia 3) pDISVP4/Sma I (colonia 3) pDISVP4 NO cortado (colonia 4) pDISVP4/Sma I (colonia 2) pDISVP4 NO cortado (colonia 2) DNA Ladder XVII pDISVP4 NO cortado (colonia 1) pDISVP4/Sma I (colonia 1) Para determinar la presencia de la secuencia génica VP4 en el plásmido de las colonias recombinantes, se analizaron por restricción con la enzima Sma I. Esta enzima corta sólo al vector (pDISPLAY), en la región de policlonación, en el sitio de restricción 815pb, por lo que sí hay corte el vector se linearizará observándose una banda de aproximadamente 5.3Kb lo cual significa que las colonias NO son recombinantes, es decir, el gene de la proteína VP4 no esta clonado. En cambio si no hay corte, entonces las colonias son recombinantes, debido a que se perdió el sitio de restricción que reconoce esta enzima y por lo tanto no se observará ninguna banda. En la figura 18 y 19 se muestra la electroforesis de la restricción con la enzima Sma I, donde se observa el DNA de las 10 colonias transformadas. 4000pb 2000p 500pb Figura 18. DNA de las colonias (1-7) E.coli, cepa JM101, digerido con la enzima Sma I. Gel de agarosa 1.0% En la figura 18, no se observa ninguna banda de 5.3Kb en las colonias 1-7, después de haber digerido el DNA de las bacterias E.coli (cepa JM101) con la enzima Sma I, lo cual indica que estas colonias son transformadas, es decir, presentan el vector recombínate pCVP4. 48 pDISVP4/Sma I (colonia 8) pDISVP4 NO cortado (colonia 9 ) pDISVP4/Sma I (colonia 9) pDISVP4 NO cortado (colonia 10 ) pDISVP4/Sma I (colonia 10) pDISVP4 NO cortado (colonia 8 ) DNA Ladder X En la figura 19, no se observa ninguna banda en las colonias 9 y 10 pero si se observa una banda de 5.3Kp en la colonia 8, liberada del DNA de la bacterias E.coli (cepa JM101) después de haber sido digerido con Sma I, lo cual indica que esta colonia no esta transformada pero las colonias 9 y 10 si son colonias transformadas, es decir, presentan el vector recombínate pCVP4. 5.3Kp ( vector) 4070pb 517pb 220pb Figura 19. DNA de las colonias (1-7) E.coli, cepa JM101, digerido con la enzima Sma I. Gel de agarosa 1.0% En la figura 19, no se observa ninguna banda en las colonias 9-10, lo cual indica que son recombinantes. 49 ANÁLISIS POR PCR PARA VP4 PCR VP4 ( clona 10) PCR VP4 ( clona 5) PCR VP4 ( clona 6) PCR VP4 ( clona 7) PCR VP4 ( clona 8) PCR VP4 ( clona 9) DNA Ladder X DNA Ladder 100pb Control (+) PCR VP4 ( clona 4) PCR VP4 ( clona 3) PCR VP4 ( clona 2) PCR VP4 ( clona 1) DNA Ladder X Control (-) Para corroborar la presencia de la secuencias génica VP4 en el plásmido, en las 10 colonias rtransformadas, se realizó una reacción de PCR. El producto de la reacción de PCR fue analizado mediante un corrimiento electroforético en un gel de agarosa al 1.0% y visualizado en un equipo de documentación de geles (figura 20). 4070pb 517pb 220pb 206pb (VP4) Figura 20. Secuencia génica VP4, amplificada por PCR. Gel de agarosa 1.0% En la figura 20, se observa una banda de 206pb, que corresponde a la amplificación del gene que codifica a la proteína VP4 del virus de la poliomielitis human, comparada con una banda de 206pb (control positivo [+]) que corresponde al vector pXpA que contiene el genoma completo del virus de la polio. Al amplificar el fragmento génico VP4, pudimos estar completamente seguros que el gene había sido clonado. 50 TRATAMIENTO DE LOS TUMORES PCR pVAXVP1 PCR pVAXVP4 Control (+) VP1 PCR pDISVP1 DNA Ladder X Control (+) VP4 PCR pDISVP4 El DNA plasmídico utilizado para el tratamiento de los tumores fue analizado por PCR para corroborar la presencia de los genes clonados antes de ser inyectados en los tumores (figura 21). 1018pb 517pb 220pb 915pb 206pb Figura 21. Secuencias génicas de VP1 y VP4 amplificadas por PCR. Gel de agarosa 1.0% En la figura 21, se observa una banda de 915pb, que corresponde a la amplificación del gene que codifica a la proteína VP1 del virus de la poliomielitis humana, así como una banda de 206pb, que corresponde a la amplificación del gene que codifica a la proteína VP4 del virus de la polio, comparadas con una banda de 915pb (control positivo [+] para VP1) y otra banda de 206pb (control positivo [+] para VP4 ) que corresponden al vector pXpA que contiene el genoma completo del virus de la polio. Al amplificar los fragmentos VP1 y VP4, pudimos estar completamente seguros que los genes estaban presentes y que los plásmidos podían ser usados para el tratamiento de los tumores. 51 EL DISEÑO EXPERIMENTAL DE VACUNACIÓN FUE EL SIGUIENTE: Ratones C57Bl/6 N= 20 Controles SIN Vacunar N= 10 Vacunados contra la polio N= 10 Inducción de Tumores N= 10 Inducción de Tumores N= 10 SIN Tratamiento N= 2 Tratamiento de Tumores con pDISVP1 y pDISVP4 N= 4 SIN Tratamiento N= 2 Tratamiento de Tumores con pDISVP1 y pDIS VP4 N= 4 Tratamiento de Tumores con pVAXVP1 y pVAXVP4 N= 4 Tratamiento de Tumores con pVAXVP1 y pVAXVP4 N= 4 52 DESARROLLO DE TUMORES DE MELANOMA Para inducir el crecimiento de tumores en los ratones experimentales, se inyectaron 5 x 104 células de melanoma B16FO. El desarrollo de los tumores fue monitoreado cada 2 días. A los 10 días los tumores tenían un promedio de 1.0 mm2 de área, el máximo observado fue de 6.5 mm2 a los 34 días. El crecimiento promedio de los tumores se muestra en la figura 22. ÁREA DEL TUMOR (mm2) CRECIMIENTO DEL TUMOR 7 6 5 4 3 2 1 0 6 12 18 TIEMPO (días) 24 30 36 Figura 22. Desarrollo de tumores de melanoma en ratones. Los tumores fueron inducidos mediante la inyección de 5x104 células B16FO. En la figura 22, se observa el promedio del área de los ratones, donde el crecimiento del tumor es proporcional al tiempo. 53 TRATAMIENTO DE LOS TUMORES Cuando los tumores alcanzaron 1.5mm2 fueron sometidos al tratamiento con los plásmidos pDISVP1 Y pDISVP4. Como controles, un grupo de tumores fueron tratados con los plásmidos pVAXVP1 y pVAXVP4, los cuales expresan las proteínas VP1 y VP4 pero no inducen su anclaje a la membrana celular. Adicionalmente, un grupo de tumores fue tratado con el vector pDISPLAY vacío. Estos tratamientos se dieron a ratones vacunados contra la polio y a ratones control no vacunados. (figura 23) ÁREA DEL TUMOR (mm2) RATONES NO VACUNADOS 8 7 6 5 4 3 2 1 0 SIN tratamiento pVAXVP1 y pVAXVP4 pDISVP1 y pDISVP4 6 12 18 24 30 36 pDISPLAY vacío TIEMPO (días) A) ÁREA DEL TUMOR (mm2) RATONES VACUNADOS 8 SIN tratamiento pVAXVP1 y pVAXVP4 pDISVP1 y pDISVP4 pDISPLAY vacío 6 4 2 0 TIEMPO (días) B) Figura 23. Desarrollo de tumores A)en ratones no vacunados con 4 diferentes tratamientos. B) ratones vacunados con 4 diferentes tratamientos. Los tumores fueron inducidos 4 mediante la inyección de 5 x10 células B16FO. 54 En el caso de los ratones control no vacunados, el grupo sin tratamiento muestran un crecimiento proporcional al tiempo, como se observa en la figura 24, donde el área del tumor alcanza 6.5 mm2. El análisis estadístico mostró que la diferencia entre el promedio de las áreas de los tumores de este grupo con respecto al promedio del grupo vacunado no es significativa (p= 0.589), es decir, que el promedio de ambos grupos es similar. El grupo tratado con pVAXVP1 y pVAXVP4 muestra un crecimiento proporcional al tiempo semejante al grupo sin tratamiento, como se observa en la figura 25. El análisis estadístico mostró que la diferencia entre el promedio de las áreas del grupo vacunado sin tratamiento y este grupo no es significativa (p= 0.62), es decir, que el promedio de las áreas de ambos grupos son estadísticamente iguales. El grupo tratado con pDISVP1 y pDISVP4 muestra una ulceración como se observa en la figura 26, la cual pudo ser causada por lo agresivo del tumor o por la agresividad entre los animales, lo que no permitió la medición adecuada. Por lo anterior será necesario separar a los animales. El análisis estadístico mostró que la diferencia entre el promedio de las áreas de este grupo con respecto al promedio de las áreas del grupo vacunado sin tratamiento si es significativa (p<0.0001), lo que indica que las áreas de estos tumores son diferentes entre sí, por lo que se piensa que el tratamiento es efectivo. Para el caso de los ratones vacunados, el grupo sin tratamiento muestra un crecimiento proporcional al tiempo semejante al del grupo no vacunado sin tratamiento, como se observa en la figura 27. El grupo tratado con pVAXVP1 y pVAXVP4 presentan un crecimiento proporcional al tiempo semejante al del grupo no vacunado sin tratamiento, como se observa en la figura 28. El análisis estadístico mostró que la diferencia entre el promedio de las áreas de este grupo con respecto al promedio del grupo no vacunado sin tratamiento si es significativa (p<0.0001), es decir, que el promedio de las áreas de ambos grupos son diferentes, lo que indica que el tratamiento funcionó. Los ratones tratados con pDISVP1 y pDISVP4 presentan una regresión del tumor después de 3 días de haber sido tratados, como se observa en la figura 29. El análisis estadístico mostró que la diferencia entre el promedio de las áreas del grupo vacunado sin tratamiento y este grupo es significativa (p= 0.001), lo que indica que el promedio de las áreas de ambos grupos son diferentes entre sí, sugiriendo que el tratamiento es efectivo. En la figura 30, se compara el tamaño de los tumores de un ratón vacunado sin tratamiento y un ratón vacunado tratado con pDISVP1 y pDISVP4 y en la figura 31, se observan los tumores extraídos de dichos ratones. Por último en la figura 32, se muestra el tumor de un ratón vacunado tratado con pDISPLAY vacío. 55 Figura 24. Se observa el tumor del ratón no vacunado sin tratamiento, el cual muestra un crecimiento proporcional al tiempo semejante al tumor del ratón vacunado sin tratamiento. Figura 24. Se muestra el melanoma de un ratón no vacunado sin tratar, el cual presenta un tamaño de tumor proporcional al tiempo semejante al tumor del ratón vacunado sin tratamiento. observado al final de los tratamientos. 56 Figura 25. Se observa el tumor del ratón no vacunado tratado con pVAXVP1 y pVAXVP4, el cual muestra un crecimiento semejante al de los ratones vacunados sin tratamiento. Figura 25. Se observa el tumor de un ratón no vacunado tratado con pVAXVP1 y pVAXVP4, el crecimiento del tumor es proporcional al tiempo, semejante al crecimiento de los ratones vacunados sin tratar. visto al final de los tratamientos. 57 Figura 26. Se observa el tumor del raton no vacunado tratado con pDISVP1 y pDISVP4, el cual muestra un tumor de crecimiento semejante al de los ratones vacunado sin tratamiento. también se observa una ulcera debido a lo agresivo del tumor. Figura 26. se muestra el melanoma de un ratón no vacunado tratado con pDISVP1 y pDISVP4, el cual presenta un crecimiento tumoral semejante al de los ratones vacunados sin tratamiento, se observa una ulcera debido a lo agresivo del tumor, visto al final del tratamiento. 58 Figura 27. se observa el tumor del ratón vacunado sin tratamiento, el cual muestra un crecimiento proporcional al tiempo. Figura 27. Se muestra el melanoma de un ratón vacunado sin tratar, el cual presenta un tamaño proporcional al tiempo, observado después de 4 semanas después de la inoculación de las células B16FO. 59 Figura 28. Se observa el tumor del ratón vacunado y tratado con pVAXVP1 y con pVAXVP4, el cual muestra una coloración distinta por la sobre-expresión de melanina, cuyo crecimiento es diferente al de los ratones vacunados sin tratamiento. Figura 28. Se muestra el melanoma de un ratón vacunado y tratado con pVAXVP1 y con pVAXVP4, el cual muestra una coloración distinta por la sobre-expresión de melanina, y cuyo tamaño es diferente al de los ratones vacunados sin tratar, observado al final del tratamiento. 60 Figura 29. Se observa el tumor del ratón vacunado tratado con pDISVP1 y pDISVP4, el cual muestra regresión y cicatrización total del tejido. el crecimiento es diferente al de los ratones vacunados sin tratamiento. visto al final del tratamiento. Figura 29. Se muestra el melanoma de un ratón vacunado tratado con pDISVP1 y pDISVP4, el cual presenta regresión y cicatrización total del tejido. el tamaño diferente al de los ratones control sin tratamiento, observado al final del tratamiento. 61 Figura 30. Se muestra al ratón vacunado sin tratar en la parte superior de la imagen, donde se aprecia el tumor sin ningún cambio en el tamaño, en la parte inferior de la imagen se observa el tumor del ratón vacunado tratado con pDISVP1 y pDISVP4, el cual presenta una gran diferencia de tamaño como una cauterización del tejido. Figura 30. en la parte superior de la imagen se muestra el melanoma de un ratón vacunado sin tratar y en la parte inferior de la imagen se muestra el melanoma de un ratón vacunado tratado con pDISVP1 y pDISVP4. se observa una gran diferencia en tamaño, como la cauterización del tejido en el melanoma ,esto se observo a los 4 días después del tratamiento. 62 Figura 31. Se observa un acercamiento de los dos tumores extraídos de los ratones de la imagen 30, al final del tratamiento. A la izquierda se muestra el tumor del ratón vacunado sin tratamiento y a la derecha se muestra el tumor del ratón vacunado tratado con pDISVP1 y pDISVP4, el cual se presenta una regresión del tumor así como cicatrización del tejido, además de apreciar gran diferencia de tamaño. Figura 31. A la izquierda se muestra el melanoma de un ratón vacunado sin tratamiento y a la derecha se muestra el melanoma de un ratón vacunado tratado con pDISVP1 y pDISVP4. Se observa una gran diferencia en tamaño, como la cicatrización del tejido en el melanoma tratado. 63 Figura 32. Se observa el tumor del ratón vacunado tratado con pDISPLAY vacío, el cual muestra un crecimiento proporcional al tiempo semejante al tumor del ratón vacunado sin tratamiento. Figura 32. Se muestra el melanoma de un ratón vacunado tratado con pDISPLAY vacío, el cual presenta un tamaño proporcional al tiempo, observado al final de los tratamientos. 64 DISCUSIÓN Existe un gran interés en obtener un tratamiento contra el cáncer basado en la estimulación de Sistema Inmune. Pero uno de los principales inconvenientes es la escasa inmunogenicidad de las células tumorales. Actualmente se han realizado más de un centenar de ensayos utilizando los propios antígeno del tumor, células tumorales dirigidas a producir diversas interleucinas y diversas terapias. Sin embargo, muy pocos estudios consiguen resultados satisfactorios ya que falta superar mucos obstáculos metodológicos y esquemas terapéuticos más efectivos, con potencial a corto plazo o por lo menos disminuir la tasa de recurrencia y metástasis tumoral. Al evaluar la capacidad inmunoestimulante de los vectores de expresión recombinante utilizados como posible tratamiento contra el melanoma, se puede observar que la expresión de las proteínas VP1 y VP4 del virus de la poliomielitis human en las células tumorales fue satisfactoria para inducir una respuesta antitumoral. Donde los ratones vacunados tratados sólo con los plásmidos construidos (pDISVP1 y pDISVP4) presentaron tanto una regresión del tumor como cicatrización total del tejido, por lo que se piensa que el grupo tratado con los plásmidos construidos presentan una diferencia significativa del área con respecto al grupo vacunado sin tratamiento. Con respecto a los grupo no vacunado sin tratamiento, vacunado sin tratamiento o tratado con pVAXVP1 y pVAXVP4 o tratado con el vector vacío no presentaron efecto alguno. Sin embargo, los resultados son contradictorios respecto a los grupos no vacunados y vacunados tratados con pVAXVP1 y pVAXVP4 ya que en el primero este tratamiento no afecta el desarrollo del tumor pero en el segundo si lo afecta disminuyendo su crecimiento, por lo que se piensa que el grupo tratado con estos plásmidos no presentan diferencia significativa del promedio de las áreas de los tumor con respecto a los tratados con los plásmidos construidos, por lo cual podría servir también como un posible tratamiento contra el cáncer. Por otro lado, el grupo no vacunado tratado con pDISVP1 y pDISVP4 presentaron una diferencia significativa del área con respecto al grupo control sin tratamiento por causa de la presencia de una ulcera tal vez debido a lo agresivo del tumor o a la agresividad entre animales, por lo cual el área del tumor no fue medida adecuadamente por lo que será necesario separar a los animales en futuros estudios. El grupo vacunado tratado con el vector pDISPLAY vacío no presento efecto alguno, esto indica que el efecto visto en el grupo tratados con pDISVP1 Y pDISVP4 se debió a la presencia de las proteínas clonadas en dicho vector y no algún componente del mismo. 65 La posible explicación de este efecto podría ser que es originado por la participación de linfocitos, tanto T como B, ejerciendo un papel fundamental en la respuesta inmune. Las células tumorales son susceptibles de ser destruidas por casi todos los mecanismos efectores de la respuesta inmunológica. ¯ Complemento ( en presencia de anticuerpo). ¯ Células asesinas naturales, macrófagos y leucocitos PMN, a través del proceso de la ADCC (citotoxicidad celular dependiente de anticuerpo). ¯ Linfocitos T citotóxicos CD4+ y CD8+. ¯ Células asesinas naturales (por citotoxicidad directa). ¯ Citocinas. IFNγ, TNFα y TNFβ (linfotoxina). INDUCCIÓN DE LA RESPUESTA INMUNITARIA: Los macrófagos, las células dendríticas y las células de Langerhans (APC) podrían estar capturando y procesando los antígenos (VP1 y VP4) y presentándolos como oligopéptidos o epitopos a los linfocitos T. Los linfocitos B también podrían estar capturando estos antígenos, procesándolos y presentándolos a las células T. Estos epitopos serían presentados a las células T, asociados a moléculas MHC de clase I. Los cuales serían reconocidos por los linfocitos T a través de un complejo macromolecular formado por las moléculas TCR-CDA o TCR-CD8 que se encuentran en íntima asociación con el complejo CD3. Durante el procesamiento de los antígenos, el macrófago sintetizará y liberará las citocinas IL-1, IL-6 e IL-12, entre otras. Para la activación de las células T se requiere de varias señales simultáneas que derivan del reconocimiento de los epitopos VP1 y VP4 asociados a moléculas MHC de clase I, de la interacción entre las moléculas coestimulatorias de las APC (B7-1 y CD40) y sus receptores en las células T (CD28 y CD154, respectivamente) y del efecto de las citocinas liberadas por las APC. Una de las citocinas, IL-1, podría estimular a los linfocitos Th1 y Th2 y los inducirá a producir y expresar receptores para IL-2 (IL-2R). La misma IL-1 o alguna otra (¿IL-6?) estimulará a las células Th1 y las inducirá a producir IL-2, un factor de proliferación de las células T, B y otras células. Muchas otras citocinas participan en el proceso de inducción de la respuesta inmunitaria, activando y desactivando células, esto es, regulando su función. Las células activadas presentarán cambios metabólicos que las inducirán a proliferar y a producir y liberar las citocinas que están programadas a producir. 66 Estas citocinas participarán en las fases subsecuentes de las respuestas inmunitarias, incluyendo la promoción de la inflamación, la activación de macrófagos y la coestimulación de los linfocitos B para la producción de anticuerpos (anti-VP1 y anti-VP4), además de las muchas otras actividades que conforman la respuesta inmunitaria. Por otra parte, las células Th1 ayudarán a la destrucción de células tumorales y a la activación de macrófagos, además de ayudarán a los linfocitos B en su tarea de producir anticuerpos, al producir citocinas para estimular a las células B induciéndolas a proliferar y diferenciarse en células plasmáticas para producir los anticuerpos (anti-VP1 y anti-VP4). La coexistencia de los antígenos y su anticuerpos propiciará la formación de complejos inmunes, tanto fijos en los sitios de lesión como solubles en la circulación. Estos complejos son usualmente fijadores de complemento, de tal manera que su activación puede ocurrir localmente o en circulación. Lo que conlleva a la activación del complemento que llevará a lisis celular, esta activación se inicia cuando dos o más fragmento Fc de los anticuerpos en los complejos inmunes reaccionan con el componente C1 del complemento; la interacción ocurre a través del subcomponente C1q pero para que ocurra se requiere al menos una molécula de IgM o dos moléculas de IgG situadas en estrecha proximidad. El daño celular será causado por el complejo de ataque a la membrana o complejo MAC. Este propiciará alteraciones en la estructura de la membrana de la célula tumoral que facilitarán la penetración parcial de C8 y la polimerización e inserción C9. El complejo de ataque a la membrana es el responsable de la destrucción lítica de las células al favorecer la desorganización de los lípidos de la membrana, al producir en ella poros o agujeros a través de los cuales ocurre la salida y entrada de agua, iones y macromoléculas además de incremento en la permeabilidad vascular, asfixia y quimiotaxis, opsonización, endocitosis que promueven la agregación y remoción por fagocitosis, adherencia inmune y por último lisis celular. Por ultimo la asesinas naturales (NK) son capaces de matar algunas células tumorales. El mecanismo de daño incluye la interacción de anticuerpos de la clase IgG con los antígenos VP1 y VP4 presentes en las células tumorales. Las regiones Fc de los anticuerpos son reconocidos por receptores (FcgammaRII y FcgammaRIII) presentes en algunas células citotóxicas como los macrófagos, los neutrófilos y las células NK. Mientras que los macrófagos y los neutrófilos destruyen a las “células blanco” fundamentalmente por fagocitosis, las células NK lo hacen por contacto directo. Se ha visto que las células inyectan componentes de sus gránulos (granzimas, perforinas y otras proteínas catiónicas) y materiales tóxicos del tipo de los radicales libres del oxígeno a las células blanco, que finalmente son destruidas por mecanismos tanto necróticos como apoptóticos. 67 En otras ocasiones, cuando la célula blanco es de mayor talla que la célula fagocítica, ocurrirá un fenómeno descrito como “ataque citotóxico”, en el cual los fagocitos hacen contacto con las células y, mientras hacen el vano intento de endocitarlas, se estimulan metabólicamente y liberan radicales libres del oxígeno, además de sus componente lisosomales con todo su arsenal citocida. Al recibir el estimulo los linfocitos Th1 además podrían funcionar como células citotóxicas (Thc) y son capaces de reconocer y matar a las células portadoras de los antígenos VP1 y VP4. Para esto los linfocitos Thc liberan citocinas tóxicas como el factor de necrosis tumoral beta (TNFβ) o linfotoxina, perforinas, y enzimas hidrolíticas (hidrolasas). Los linfocitos Th1 activados (Th1*) también producen otras citocinas, entre ellas, interferón gamma (IFNγ). Esta citosina induce, en los macrófagos, la producción de factor de necrosis tumoral alfa (TNFα), el cual, actuando autocrinamente, estimula la síntesis de la óxido nítrico cintaza inducible (iNOS). La iNOS incrementa el metabolismo de la arginina, promoviendo la producción y liberación de óxido nítrico (.NO). El óxido nítrico induce a la muerte de la célula. Otro vía por la cual se induciría la muerte de las células tumorales que expresan en antígeno VP1y VP4 es inducida por Fas: La célula T citotóxica (LcT) interacciona con la célula tumoral a través de el reconocimiento de la molécula Fas. La interacción FasL-Fas induce señales que se transducen al citoplasma de la célula tumoral y que finalmente inciden en las mitocondrias, lesionándolas. Las mitocondrias lesionadas dejan escapar citocromo c y éste interacciona con el factor promotor de apoptosis Apaf-1 y lo aligomeriza [(Apaf-1)n]. El Apaf-1 oligomerizado activa a las caspasa 8 y 9 (Pro c-8/c-9). Las caspas 8 y 9 activadas (c-8*/c-9*) activan a la por-caspasa 3 (Pro c-3). La caspasa 3 (c-3*) interacciona con la membrana mitocondrial, promoviendo la liberación adicional de citocromo c y amplificando las señales de apoptosis. La caspasa 3 activa también otras caspasas . Algunas de estas caspasas funcionan como nucleasas en tanto que otras activan a otras nucleasas. Las caspasas con actividad de nucleasas y las nucleasas fragmentan al ADN iniciando los cambios morfológicos que caracterizan a las células apoptóticas. Cabe señalar que no se puede afirmar que participaron tanto los linfocitos T y B ni precisar el efecto causado (apoptosis o/ y necrosis).También debe tomarse en cuenta la vía de administración, el numero de dosis, así como una serie de posibilidades para determinar si los plásmidos de expresión recombinante utilizados para estimular la Respuesta Inmune, funcionan como una posible terapia génica contra el cáncer y para demostrar lo anterior sería necesario llevar acabo algunos experimentos como: la determinación de la expresión de los genes VP1 y VP4 para estar seguros de su presencia, la evaluación de anticuerpos por ELISA, linfocitos T, así como la determinación de la inducción de apoptosis, todo esto para asegurar que la regresión del tumor se debió a la participación tanto de linfocitos T y que la presencia de4 anticuerpos activa alas células asesinas naturales para inducir apoptosis. 68 CONCLUSIÓN La expresión de las proteínas VP1 y VP4 del virus de la poliomielitis humana en las células tumorales indujo una respuesta anti-tumoral. Donde los ratones vacunados tratados sólo con los plásmidos construidos (pDISVP1 y pDISVP4) presentaron tanto regresión del tumor como cicatrización total del tejido. En el grupo no vacunado tratado con pVAXVP1y pVAXVP4 presentó un crecimiento muy lento, por lo cual podría servir también como un posible tratamiento contra el cáncer. En el presente estudio, la administración por vía intradérmica de los plásmidos recombinantes utilizados como tratamiento, demostró su potencial como agente anti-cancerígeno al observar regresión en el melanoma de ratones inmunizados con la vacuna génica. 69 PERSPECTIVAS ¯ Determinar la expresión in vitro e in vivo de los genes de las proteínas VP1 y VP4 clonadas, mediante su transfección en líneas celulares y la detección de los productos génicos inmunohistoquímica. ¯ Evaluar la presencia de anticuerpos. ¯ Evaluar la presencia de linfocitos T citotóxicos. ¯ Determinar la inducción de apoptosis en las células tumorales. 70 BIBLIOGRAFÍA 1. Cabrera G y García-Carranca A. La terapia del cáncer. Oxfor University Press Inc. New York, N. Y. 111-119 pp. 2. Delves P.J. Celular Immunology. 1994. Bios Scientific Publishers. Oxford United Kingdom. 3. Evans A. S. 1991. Viral Infection of Humans Epidemiology and Control. Tercera Edición. Plenum Medical Book Company. New York, N. Y. 191263 pp. 4. Ferriols F, Ferriols R y Colomina J. 1997 Inmunoterapia Tumoral. Vacunas y Terapia Génica. Farm Hosp.21:166-170. 5. Fiore L, Ridolfi B, Genovese D, Buttinelli, G, Lucioli S, Lahm A and Ruggeri F. M . 1997. Poliovirus Sabin Type 1 Neutralization Epitopes Recognized by Immunoglobulin A Monoclonal Antibodies. J Virol 71:6905-6912. 6. Hersey P. 2003. Adjuvant therapy for high-risk primary and resected metastatic melanoma. J Med Inter.33:33-43. 7. Herremans T, Reimerink J,, Buisman A, Kimman T and Koopmans M.1999. Induction of Mucosal Immunity by Inactivated Poliovirus Vaccine is Dependent on Previous Mucosal Contact with Live Virus. J Immunol 162:5011-5018. 8. http://www.bbmundo.com/articulos/polio-p.htm 9. http://encyclopedia.uv.es/wiki.phtml?title=Poliomielitis&action=print 10. http://www.facmed.unam.mx/deptos/microbiologia/librovir/enterovirus.html 11. http://www.fefoc.org/educacion/rincon/terapia_genica.jsp 12. http://www.smmhc.com/healthcontent/sadult/infectious/polio.htm 13. http://www.ondasalud.com/edicion/noticia/0,2458,5261,00.html 14. http://www.umm.edu/esp_ency/article/001402.htm 15. http://www.uv.es/∼vicalegr/CLindex/CLmelanoma/CLmmpbl1.htm 16. Hu, Hing-Ming., Winter, H., Urba, W. J. and Fox, B.A. 2000 Divergent Roles for CD4+ T cells in the Priming and Effector/Memory Phases of Adoptive Immunotherapy. J Immunol.165:4246-4253. 71 17. Hull H. F and Aylward R. B.2001 Progress towards global polio eradication. Vaccine 19:4378-4384. 18. Hung K. G, Hayashi R, Lafond-Walker A, Lowenstein C, Pardoll D and Levitsky H. 1998. The Central Role of CD4+ T Cells in The Antitumor Immune Response. J. Exp. Med. 188:2357-2368. 19. Kammula U, White D. E, Rosenberg S. A. 1988. Trends in the Safety of High Dose Bolus Interlukin-2 Administration in Patients with Metastatic Cancer. Cancer. 83:797-805. 20. Kominsky S. L, Torres B. A. Hobeika A. C, Lake F.A and Johnson, H. M. 2001. Superantigen Enhanced Protection Against a Weak TumorSpecific Melanoma Antigen Implications for Prophylactic Vaccination Against Cancer. Int. J. Cancer. 94:834-841. 21. Kutubuddin M, Simons J and Chow M. 1992. Identification of T-Helper Epitopes in lhe VP1 Capsid Protein of Poliovirus. J Virol. 66:3042-3047. 22. Leclerc C, Deriaud E, Mimic, V and Werf S. Van Der. 1991. Identification of a T-Cell Epitope Adjacent to Neutralization Antigenic Site 1 of Poliovirus Type 1. J Virol. 65:711-718. 23. López-Guerrero J A, Alonso M, Martín-Belmonte F, and Carrasco L. 2000 Poliovirus Induces Apoptosis in the Human U937 Promonocytic Cell Line . Virol. 272:250-256. 24. Mahon B. P, Katrak K. and Mills H.G. 1992. Antigenic Sequences of Poliovirus Recognized by T Cells: Serotype-Specific Epitopes on VP1 and VP3 and Cross-Reactive Epitopes on Vp4 Defined by Using CD4+ T-Cell Clones. J Virol 66:7012-7020. 25. Minor P.D. 1996. Poliovirus Biology. Structure.4:775-778. 26. Nielsen M. B and Marincola F. M. 2000. Melanoma vaccines: the paradox of T cell activation without clinical response. Cancer Chemother Pharmacol 46:S62-S66. 27. Ohka S and Nomoto A. 2001. Recent insights into poliovirus pathogenesis. TRENDS in Microbiology.9:501-506. 72 28. Rojas-Espinosa O. 2001. Inmunología (de memoria) 2a . Edición. Edirorial Médica Panamericana. México,. D. F. 101-121, 319-335, 343pp. 29. Rojas-Martínez A, Martínez-Dávila E, Hernández-García A, Aguilar-Córdova A y Barrera-Saldaña H A. 2002 Terapia génica del cáncer. Rev Invest Clin;54:57-67. 30. Schneeberger A, Goos M, Stingl and Wagner SN.. 2000. Management of malignant melanoma: new developments in immune and gene therapy. Clin and Exp Dermat 25.509-519. 31. Smith C and Cerundolo V. 2001. Immunotherapy of melanoma. Immunol.104:1-7. 32. Soiffer R., et al.1998. Vaccination with irradiated autologous melanoma cells engineered to secrete human granulocyte-macrophage colonystimulating factor generates potent antitumor immunity in patients with metastatic melanoma. Proc. Natl. Acad. Sci. USA.95:13141-13146. 33. Touloukian, C. E., Leitner, W. W., Robbins, P. F., et al. 2002. Expression of a “Self”-Antigen by Human Tumor Cells Enhances Tumor Antigenspecific CD4+ T-Cell Function. Cancer Res. 62:5144-5147. 34. Urugel S, Rebmann V, Ferrone S, Tilgen W, Grosse-Wilde Hand Reinhold U. 2001. Soluble Human Leukocyte Antigen-G Serum Level Is Elevated in Melanoma Patients and is Further Increased by Interferon-α Immunotherapy. Cancer. 92:369-376. 35. White, D. O., Fenner, F. J. 1994. Medical Virology. Cuarta Edición. Inc, Academic Pres. San Diego, California. 382- 439 pp. 36. Wilson, J. 1991. Principios de Medicina Interna. 12a Edición. Nueva Editorial Interamericana Mc Graw-Hill. México, D., F. 558-565, 786-794, 836-837 pp. 37. Wimmer E and Nomoto A. 1993. Molecular Biology and Cell-free Synthesis of Poliovirus. Biologicals. 21:349-356. 73