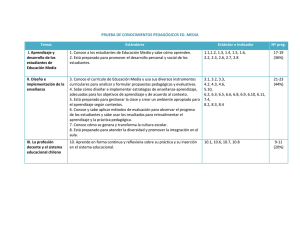
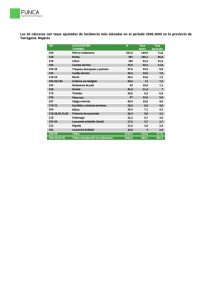
Ver/Abrir
Anuncio

República de Cuba Universidad de La Habana Facultad de Biología Mecanismos inmunológicos asociados a la vacunación terapéutica de cáncer con NGcGM3/VSSP Tesis en opción al título de Doctor en Ciencias Biológicas Por Autora: Lic. Mayrel Labrada Mon, M. C. Tutor: Lic. Luis Enrique Fernández Molina, Dr. C. Centro de Inmunología Molecular La Habana 2014 No pretendo más que lo que el hombre de ciencia, que aun cuando realiza sus experimentos con la máxima precisión, minuciosidad y previsión, jamás proclama haber alcanzado conclusiones definitivas, sino que los contempla con la mente alerta y espíritu crítico. M. K. Gandhi A Camila y Carlos La gratitud, como ciertas flores, no se da en la altura y mejor reverdece en la tierra buena de los humildes. José Martí En este documento cité las fuentes de los resultados científicos que aparecen publicados en la literatura especializada, pero ¿y las ideas y sugerencias recogidas en presentaciones, encuentros y conversaciones con compañeros y amigos? Estoy en deuda con mi tutor Luis Enrique Fernández; con las oponentes de la predefensa, Isis Torrens y Marta Ayala, del CIGB; con los amigos y colegas del CIM: Adriana Carr, Zaima Mazorra, Joel de León, Beatriz García, Circe Mesa, Belinda Sánchez, Kalet León, Ana María Vázquez, Tays Hernández, Liliana Oliver, Yanín Bebelagua, Arlhee Díaz, Giselle Hevia e Isabel Pablos; con Federica Benvenuti y Francesca Prete, del ICGEB en Triste, Italia. Agradezco también las críticas y los consejos de las profesoras de la Facultad de Biología de la Universidad de La Habana: Fabiola Pazos, Maria Eliana Lanio, María del Carmen Luzardo y María Cristina Pico. Agradecimiento aparte hago a Marylin Clavell y Judith Raymond, por su ayuda inmensa con los ratones. Gratitud también a mis compañeras de doctorado: Audry Fernández y Tania Carmenate, con quienes disfruté esta competencia de entrega de expedientes, exámenes, artículos enviados, aceptados y aparecidos, entrevistas con los oponentes y defensas, porque ninguna podía quedarse rezagada. Las tres llegamos juntas a la meta. Agradezco además la paciencia de mi familia, pues fueron muchos los momentos de ausencia para llegar aquí. Síntesis Los tejidos normales de humanos no expresan niveles apreciables de gangliósidos Nglicolilados, sin embargo, su presencia se describe en varios tipos de tumores de pacientes. Esto, unido al papel de los gangliósidos en la evasión de la inmunovigilancia y en la progresión tumoral, los convierte en blanco atractivo para la inmunoterapia del cáncer. En el Centro de Inmunología Molecular se desarrolla la vacuna NGcGM3/VSSP, que incluye al gangliósido NGcGM3 en las vesículas de membrana externa de la bacteria Neisseria meningitidis. En este trabajo se estudia el mecanismo de acción de la vacuna NGcGM3/VSSP en ratones, utilizando el modelo de metástasis espontánea a pulmón del carcinoma de Lewis. La comprensión del mismo permitirá el empleo de terapias óptimas y el uso de estrategias de combinación con inmunomoduladores. Se demostró la presencia del gangliósido NGcGM3 en las diferentes etapas por las que transita el modelo tumoral. La administración terapéutica de la vacuna NGcGM3/VSSP redujo las metástasis en los pulmones de los animales tratados, independientemente del carácter emulsivo de la formulación empleada y de la ruta de administración. Los resultados obtenidos son la primera evidencia de reclutamiento de células efectoras de la inmunidad adaptativa al sitio metastásico, por una vacuna basada en gangliósidos. Abreviaturas AAT: antígenos asociados a tumores ADCC: citotoxicidad cellular dependiente de anticuerpo, del inglés: antibody-dependent cellular cytotoxicity) Ac: anticuerpo AcM: anticuerpo monoclonal AET: antígeno específico de tumores Ag: antígeno APC: células presentadoras de antígenos (del inglés: Antigen Presenting Cells) AS: ácido siálico ANOVA: análisis de varianza (del inglés: Analysis of Variance) ATCC: Colección Americana de Tipos Celulares (del inglés: American Type Culture Collection) BCG: Bacilo Calmette-Guérin bm-DC: células dendríticas derivadas de precursores de monocitos de médula ósea (del inglés: bone marrow dendritic cells) BSA: albúmina de suero bovino (del inglés: bovine serum albumin) CD: moléculas de diferenciación expresadas en células del sistema inmunitario (del inglés: cluster differentiation) cDC: células dendríticas convencionales CDC: citotoxicidad dependiente de complemento (del inglés: cytotoxicity dependent complement) CEA: antígeno carcino-embrionario (del inglés: carcinoembrionic antigen) CIM: Centro de Inmunología Molecular CMAH: enzima mono fosforil citidina-N-Acetil ácido siálico hidroxilasa (del inglés: cytidine monophospho-N-acetylneuraminic acid hydroxylase) Cmah: gen que codifica para la enzima CMAH CpG: ácido desoxirribonucleico bacteriano no metilado con secuencias CpG CTL: linfocitos T citotóxicos (del inglés: cytotoxic T lymphocytes) DC: células dendríticas (del inglés: dendritic cells) ELISA: ensayo inmunoenzimático colorimétrico (del inglés: enzime immunosorbent assay) FACS: citometría de flujo (del inglés: flourescence activated cell sorter) FITC: isotiocianato de fluoresceína (del inglés: fluorescein iso-thiocyanate) FLT3-L: ligando de tirosina cinasa Fms-3 (del inglés: Fms-related tyrosine kinase 3 ligand) Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP GM-CSF: factor estimulador de colonias de granulocitos y macrófagos (del inglés: granulocytemacrophage colony stimulating factor) HPTLC: Cromatografía de alta resolución en capa delgada (del inglés: high performance thin layer cromatography) HSP proteínas de estrés térmico (del inglés: heat shock proteins) IFN: interferón IT: inmunoterapia ITAE: inmunoterapia activa específica KLH: hemocianina de la linfa del cangrejo Bayoneta (del inglés: keyhole limpet hemocyanin) LPS: lipopolisacárido MCI: muerte celular inmunogénica MDSC: células supresoras de origen mieloide (del inglés: myeloid-derived suppressor cells) NAcGM3: gangliósido N-acetil GM3 Neu5Ac: ácido N-acetil neuramínico (del inglés: N-acetyl neuraminic acid) Neu5Gc: ácido N-glicolil neuramínico (del inglés: N-glycolyl neuraminic acid) NGcGM3: gangliósido N-glicolil GM3 NK: células asesinas naturales (del inglés: Natural Killer) NKT: células asesinas naturales que expresan el receptor de células T (del inglés: natural killer T cells) Nm: Neisseria meningitidis pDC: células dendríticas plasmacitoides PE: ficoeritrina (del inglés: phycoerithrin) PECy5: ficoeritrina-cianina (del inglés: phycoerythrin and cyanine) poli I:C: ácido polyinosinoico-polycitidílico QS-21: fracción saponina purificada de la corteza del árbol Quillaja saponaria SFT: suero fetal de ternera SSTF: solución salina tamponada con fosfatos TCR: receptor de células T (del inglés: T cell receptor) Th: linfocitos T auxiliadores (del inglés: T helper) TLR: receptores tipo Toll (del inglés: Toll-like receptors) VLDL: lipoproteínas de muy baja densidad (del inglés: very low density lipoprotein) VME: vesículas de membrana externa VSSP: proteoliposomas de muy pequeño tamaño (del inglés: very small size proteoliposomes) Índice 1. INTRODUCCIÓN ...................................................................................................... 1 2. REVISIÓN BIBLIOGRÁFICA ....................................................................................... 9 2.1 Inmunoterapia de cáncer ........................................................................................... 9 2.1.1 Vacunas de cáncer ...............................................................................................10 2.1.2 Adyuvantes en las vacunas de cáncer ..................................................................11 2.1.3 Los VSSP..............................................................................................................15 2.1.4 Mecanismos de acción de las vacunas de cáncer. ................................................16 2.2 Gangliósidos ............................................................................................................ 20 2.2.1 Localización, características estructurales, biosíntesis y función ...........................20 2.2.2 Los gangliósidos como antígenos asociados a tumores ........................................23 2.2.3 La función N-glicolilada en tumores humanos .......................................................27 2.2.4 Inmunosupresión inducida por los gangliósidos ....................................................29 2.2.5 Papel del gangliósido NGcGM3 en la progresión tumoral .....................................32 2.3 Vacunas de cáncer basadas en gangliósidos .......................................................... 33 2.4 Mecanismos de acción de las vacunas de cáncer basadas en gangliósidos............ 36 2.5 El gangliósido GM3 como blanco de la IT de cáncer ............................................... 38 2.5.1 El preparado vacunal NAcGM3/VSSP ...................................................................39 2.5.2 Racotumumab .......................................................................................................40 2.5.3 El preparado vacunal NGcGM3/VSSP ..................................................................41 3. MATERIALES Y MÉTODOS ..................................................................................... 43 3.1 Animales .................................................................................................................. 43 3.2 Líneas celulares y medios de cultivo ....................................................................... 43 3.3 AcM utilizados para la eliminación de poblaciones linfocitarias ................................ 43 3.4 AcM utilizados en la citometría de flujo .................................................................... 44 3.4.1 Para la detección del gangliósido NGcGM3 en la membrana plasmática de las células tumorales ................................................................................................................44 3.4.2 Para la determinación de moléculas de superficie en las DC ................................44 3.4.3 Para la detección de linfocitos infiltrantes en pulmón ............................................44 3.5 Gangliósidos NGcGM3 y NAcGM3 .......................................................................... 45 i Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 3.6 Los preparados vacunales NGcGM3/VSSP y NAcGM3/VSSP ................................. 45 3.7 Desarrollo de tumores primarios del carcinoma de pulmón de Lewis 3LL-D122 ....... 46 3.8 Modelo de metástasis espontánea del carcinoma de pulmón de Lewis 3LL-D122 ... 46 3.9 Extracción de gangliósidos de las muestras tumorales............................................. 47 3.10 Identificación química del gangliósido NGcGM3 ....................................................... 47 3.11 Identificación inmuno-química del gangliósido NGcGM3 en HPTLC......................... 48 3.12 Identificación inmuno-histoquímica del gangliósido NGcGM3 ................................... 48 3.13 Detección del gangliósido NGcGM3 por espectrometría de masa ............................ 49 3.14 Procedimientos de inmunización en los experimentos con animales ........................ 49 3.14.1 Experimentos de inmunogenicidad en pollos ........................................................ 49 3.14.2 Experimentos de inmunogenicidad en ratones...................................................... 50 3.14.3 Experimentos de evaluación del efecto del preparado vacunal en el modelo murino de metástasis espontánea a pulmón del carcinoma de Lewis 3LL-D122 ................. 50 3.14.4 Experimentos de medición in vivo de maduración de DC...................................... 51 3.14.5 Experimentos de medición de citocinas en suero ................................................. 51 3.15 3.15.1 Determinación de anticuerpos séricos ...................................................................... 51 Preparación de las muestras para la titulación de los Ac anti-gangliósido en los pollos ........................................................................................................................ 52 3.15.2 Preparación de las muestras para la evaluación de los Ac anti-gangliósido en los ratones ..................................................................................................................... 52 3.16 Reconocimiento de la línea tumoral P3X63 por los sueros de los animales vacunados .......................................................................................................................... 53 3.17 Obtención de las células dendríticas murinas a partir de la médula ósea ................. 53 3.17.1 Obtención de las DC convencionales.................................................................... 53 3.17.2 Obtención de las DC plasmacitoides .................................................................... 54 3.18 Maduración de células dendríticas con NGcGM3/VSSP ........................................... 54 3.19 Determinación de la expresión de moléculas de superficie por FACS ...................... 55 3.20 Determinación de la frecuencia de células inmunes en pulmones ............................ 55 3.21 Medición de citocinas producidas por las DC ........................................................... 56 3.22 Experimentos de eliminación de poblaciones celulares in vivo ................................. 56 3.23 Análisis estadístico ................................................................................................... 56 ii Índice 4. RESULTADOS....................................................................................................... 59 4.1 La expresión del gangliósido NGcGM3 en el modelo de carcinoma pulmonar de Lewis 3LL-D122 se incrementa en las metástasis respecto a los tumores primarios .......... 59 4.2 El preparado vacunal NGcGM3/VSSP es inmunogénico en modelos animales ....... 62 4.3 El preparado vacunal NGcGM3/VSSP disminuye las metástasis pulmonares en el modelo murino estudiado ............................................................................................... 67 4.4 La presencia del gangliósido NGcGM3 en la vacuna es necesaria para la actividad antitumoral del preparado NGcGM3/VSSP ........................................................................ 70 4.5 La actividad antimetastásica del preparado vacunal NGcGM3/VSSP es independiente de la vía de administración.......................................................................... 71 4.6 La vacunación con NGcGM3/VSSP induce la maduración de células dendríticas ... 72 4.7 Las células NK y NKT están involucradas en la actividad anti-metastásica del preparado vacunal NGcGM3/VSSP ................................................................................... 76 4.8 Las células CD8 y CD4 participan en la actividad anti-metastásica del preparado vacunal NGcGM3/VSSP .................................................................................................... 79 4.9 La vacunación con NGcGM3/VSSP promueve la infiltración de células efectoras de la inmunidad adquirida al sitio metastásico.................................................................... 81 4.10 El infiltrado de leucocitos correlaciona con la inhibición de la metastización provocada por la vacunación con NGcGM3/VSSP ............................................................. 83 5. DISCUSIÓN .......................................................................................................... 85 6. CONCLUSIONES ................................................................................................... 97 7. RECOMENDACIONES ............................................................................................ 99 8. BIBLIOGRAFÍA .................................................................................................... 101 9. ANEXOS ............................................................................................................ 127 iii 1. ÍNTRODUCCÍON El cáncer constituye un conjunto de enfermedades que afecta a millones de personas en el mundo y genera un alto número de muertes cada año. Por la incidencia y la mortalidad crecientes, así como por el impacto psicológico sobre la población y el hecho de que, por lo general, los pacientes no mueren por el tumor primario, sino por el desarrollo de metástasis, se considera un serio problema de salud para la humanidad. En Cuba, durante las últimas décadas, las neoplasias malignas constituyeron una de las principales causas de mortalidad, situándose en el 2012 en la primera causa de muerte, con un total de 22 532 fallecidos para todas las edades (MINSAP, 2013). La terapia del cáncer se basa en el uso simultáneo de diversas alternativas, entre las que se destacan la cirugía, la quimioterapia, la radioterapia, la hormonoterapia y la bioterapia; que incluye la inmunoterapia (IT). Numerosas moléculas son consideradas como blanco para la IT de cáncer por sus niveles de expresión y la posibilidad de ser reconocidas por los componentes del sistema inmunológico. En este trabajo se hace referencia a los gangliósidos, glicoesfingolípidos presentes en la cara externa de las membranas plasmáticas de las células de los vertebrados (Sonnino et al., 2007). La elección de los gangliósidos como antígenos asociados a tumores (AAT) y su utilización como blancos de la IT activa antitumoral, está determinada por su densidad de expresión en la célula tumoral (Nores et al., 1987; Zhang et al., 1997). Esto constituye la base de la IT activa específica (ITAE) contra los gangliósidos, que tiene el propósito de generar una respuesta efectiva de tipo humoral y celular contra el tumor. En particular, la variante N-glicolilada del gangliósido GM3 (NGcGM3) está ausente en los tejidos de individuos sanos, mientras que está presente en tumores de mama, melanoma, colon, recto, hepatocelular, estómago y páncreas, glioblastoma, neuroblastoma, sarcomas, 1 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP cáncer de pulmón de células no pequeñas, linfomas y mielomas, así como en metástasis (Carr et al., 2000; Carr et al., 2003; Diaz et al., 2003; Oliva et al., 2006; Blanco et al., 2011a; Blanco et al., 2011b; Scursoni et al., 2011; Blanco et al., 2012; Blanco et al., 2013). Esta expresión diferencial permite clasificarlo como un antígeno específico de tumores (AET). Estudios recientes reconocen además el papel del gangliósido NGcGM3 en la progresión tumoral (Gabri et al., 2009; Segatori et al., 2012a; Segatori et al., 2012b; Casadesus et al., 2013). Adicionalmente, de León y col. demostraron que el gangliósido NGcGM3 tiene funciones inmunosupresoras sobre células dendríticas y células T CD4+ (de Leon et al., 2006; de Leon et al., 2008). El desarrollo de terapias antitumorales en el Centro de Inmunología Molecular (CIM, Cuba), con el fin de incrementar la supervivencia y la calidad de vida de los pacientes con cáncer, es una de las proyecciones fundamentales que tiene la institución. En el CIM se desarrollan dos preparados vacunales para el tratamiento del cáncer que usan al gangliósido GM3 en su composición (Estevez et al., 1999). Uno utiliza la variante N-glicolilada y el otro, la variante N-acetilada (NAcGM3), que es un AAT. Ambos preparados (NGcGM3/VSSP y NAcGM3/VSSP) se obtienen a partir de la incorporación hidrofóbica del gangliósido purificado de eritrocitos de caballo (NGcGM3) o de perro (NAcGM3), a las vesículas de membrana externa (VME) de la bacteria Neisseria meningitidis (Nm), que tiene características inmunoestimuladoras (Durand et al., 2009). El estudio del mecanismo de acción del preparado vacunal NGcGM3/VSSP en un modelo animal que exprese el gangliósido NGcGM3 y que haga énfasis en los mediadores inmunológicos celulares asociados a la vacunación es relevante para el avance de la ITAE contra este antígeno (Ag) tumoral. El modelo murino de metástasis espontánea a pulmón con el carcinoma de Lewis 3LL-D122 (Gorelik et al., 1980; Eisenbach et al., 1983) ha sido 2 Introducción ampliamente empleado para estudiar terapias antitumorales, pues simula un escenario similar al desarrollo de los tumores en pacientes, donde en muchos casos luego de la remoción del tumor primario, aparecen metástasis en órganos distantes. La comprensión del mecanismo de acción permitirá el empleo de terapias óptimas, así como el uso de distintas estrategias de combinación con inmunomoduladores. El campo de las vacunas de cáncer es de gran actualidad y los resultados alcanzados en el desarrollo de estas terapias en el CIM lo colocan en la primera línea de las investigaciones mundiales relacionadas con este campo. Con el preparado vacunal NGcGM3/VSSP emulsionado en Montanide ISA 51 (NGcGM3/VSSP/ Montanide) se realizaron varios ensayos clínicos en Cuba, en pacientes de melanoma (Osorio et al., 2008; Osorio et al., 2012; Perez et al., 2013) y cáncer de mama (Carr et al., 2003; Mulens et al., 2010). Los pacientes tratados con NGcGM3/VSSP/ Montanide desarrollan títulos de anticuerpos (Ac) IgM e IgG anti-NGcGM3 y el suero inmune reconoce a las líneas tumorales murinas L1210 y P3X63, que expresan el gangliósido NGcGM3 (Carr et al., 2003; Mulens et al., 2010). La ITAE con otras terapias basadas en gangliósidos, también genera altos títulos de Ac IgM e IgG (Ritter et al., 1991; Helling et al., 1994; Livingston, 1995a). Sin embargo, en todos los casos sólo se logró la generación de respuesta humoral (Chapman et al., 2000a; Chapman et al., 2000b; Ragupathi et al., 2003b). En un ensayo clínico Fase Ib/IIa con el preparado vacunal NGcGM3/VSSP/ Montanide en pacientes con melanoma cutáneo avanzado, se encontró vitiligo en tres de 12 pacientes vacunados (Osorio et al., 2008), aunque el gangliósido NGcGM3 no está presente en el melanocito. Por otra parte, evidencias experimentales avalan la participación de los linfocitos T CD8+ citotóxicos (CTL) en la fisiopatología del vitiligo (Steitz et al., 2005), desorden de la pigmentación con pérdida local de los melanocitos en la piel. Los CTL no se 3 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP asocian con la respuesta immune anti-gangliósidos, debido a la naturaleza de Ag Tindependiente de estos glicoesfingolípidos. Por tanto, la aparición de vitiligo en los pacientes tratados con NGcGM3/VSSP/ Montanide sugiere la generación de una respuesta CTL contra otros Ag presentes en el melanocito. Sobre la base de los antecedentes descritos se formuló la siguiente hipótesis de trabajo: La vacunación con NGcGM3/VSSP tiene efecto anti-metastásico en un modelo antígeno positivo, involucrando a la rama celular efectora de la respuesta inmune. A partir de esta hipótesis se plantearon los siguientes objetivos: 1. Demostrar que el modelo murino de metástasis espontánea con la línea de carcinoma pulmonar de Lewis 3LL-D122 es adecuado para este estudio. 2. Evaluar la inmunogenicidad y el efecto anti-metastásico del preparado vacunal NGcGM3/VSSP en ratones portadores del tumor Lewis 3LL-D122. 3. Caracterizar los mecanismos inmunológicos celulares asociados a la actividad antimetastásica del preparado vacunal NGcGM3/VSSP. Para dar cumplimiento a los objetivos se desarrollaron las siguientes tareas experimentales: Para el Objetivo 1: 1.1-Aislamiento del gangliósido NGcGM3, mediante métodos bioquímicos de extracción, de la línea celular de carcinoma pulmonar de Lewis 3LL-D122, de los tumores primarios generados a partir de la inoculación subcutánea de la misma en ratones C57BL/6 y de las metástasis espontáneas generadas en ratones portadores de estos tumores. 4 Introducción 1.2-Determinación de la presencia del gangliósido NGcGM3 en la línea celular 3LL-D122, en sus tumores primarios y en las metástasis espontáneas generadas en ratones C57BL/6 portadores de estos tumores mediante técnicas inmunoquímicas y de espectrometría de masa. Para el Objetivo 2: 2.1-Determinación de la inmunogenicidad en animales del preparado vacunal NGcGM3/VSSP, formado con los gangliósidos NGcGM3 natural y sintético, emulsionado o no en Montanide. Caracterización de la respuesta de anticuerpos inducidos, en cuanto a: título, isotipo, especificidad y reconocimiento in vitro de líneas tumorales NGcGM3 positivas. 2.2-Evaluación del efecto de la administración terapéutica del preparado vacunal NGcGM3/VSSP, con o sin Montanide, en el modelo murino de metastización espontánea a pulmón con la línea tumoral 3LL-D122. 2.3-Comparación del efecto de la administración terapéutica de los inmunógenos NGcGM3/VSSP, NAcGM3/VSSP y el complejo de proteínas de las VME de Nm sobre la metastización espontánea del carcinoma pulmonar de Lewis 3LL-D122 en ratones C57BL/6. 2.4-Determinación del impacto de la vía de administración del preparado vacunal NGcGM3/VSSP sobre la metastización espontánea a pulmón del carcinoma pulmonar de Lewis 3LL-D122 en ratones C57BL/6. Para el Objetivo 3: 3.1-Evaluación de la maduración de células dendríticas murinas y su patrón de secreción de citocinas, después de la estimulación in vitro e in vivo con el preparado vacunal NGcGM3/VSSP. 5 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 3.2-Evaluación de la participación de poblaciones efectoras de la inmunidad innata (células NK y NKT) en el efecto de la vacuna sobre la metastización espontánea del carcinoma pulmonar de Lewis 3LL-D122 en ratones, mediante experimentos de eliminación de poblaciones con anticuerpos específicos y de citometría de flujo. 3.3-Evaluación de la participación de poblaciones efectoras de la inmunidad adaptativa (células T CD8+ y T CD4+) en el efecto de la vacuna sobre la metastización espontánea del carcinoma pulmonar de Lewis 3LL-D122 en ratones C57BL/6, mediante experimentos de eliminación de poblaciones con anticuerpos específicos y de citometría de flujo. 3.4-Determinación de la correlación entre la inhibición de la metastización espontánea del carcinoma pulmonar de Lewis 3LL-D122 en ratones C57BL/6, provocada por el tratamiento con el preparado vacunal NGcGM3/VSSP, y el infiltrado leucocitario en pulmones. La principal novedad científica de este trabajo de Tesis consiste en la demostración, por primera vez, de que una vacuna de cáncer basada en un gangliósido es capaz de inhibir la diseminación metastásica mediante un mecanismo efector que involucra células T CD4+ y T CD8+. Constituye además un aporte al conocimiento la evidencia sin precedente de que el efecto antitumoral de una vacuna de cáncer basada en gangliósidos se correlaciona con el incremento de la infiltración de efectores de la inmunidad adquirida, principalmente células T CD4+, en el órgano diana. La descripción de la expresión del gangliósido NGcGM3 en el modelo murino de metástasis espontánea con la línea de carcinoma pulmonar de Lewis 3LL-D122, es también una novedad de esta Tesis, así como la demostración del enriquecimiento de este gangliósido en las células tumorales en la medida que progresa el modelo. La disminución de la metastización en el modelo experimental, lograda con la administración de una formulación del preparado vacunal NGcGM3/VSSP en forma 6 Introducción terapéutica y sin la necesidad de formar una emulsión con un adyuvante oleoso, como el Montanide ISA 51, sustentó decisivamente un cambio en la concepción del producto y la forma de administrarlo a pacientes, que ya ha sido implementado en la experimentación clínica y ha permitido constatar una disminución relativa de la toxicidad local, acompañada de un aparente incremento en la eficacia de este preparado vacunal. En esto consiste el principal aporte práctico de este trabajo. Adicionalmente, el hecho de haber obtenido resultados similares, al comparar los perfiles de inmunogenicidad alcanzados con los preparados vacunales formados por los gangliósidos NGcGM3 de origen natural o sintético, apoya un cambio favorable en la concepción del producto desde el punto de vista regulatorio. Los resultados de este trabajo están recogidos en dos publicaciones aparecidas en Expert Opinion in Biological Therapies (Labrada et al., 2010), y en Human Vaccines and Inmunotherapeutics (Labrada et al., 2014). Además, los mismos han sido presentados en eventos nacionales e internacionales. 7 2. REVÍSÍON BÍBLÍOGRA FÍCA 2.1 Inmunoterapia de cáncer El Instituto Nacional del Cáncer, perteneciente al Instituto Nacional de Salud de los Estados Unidos, define al cáncer como un conjunto de enfermedades en las cuales el organismo produce un exceso de células malignas, también llamadas cancerígenas o cancerosas, las cuales presentan un crecimiento incontrolado más allá de los límites normales. Estas células son capaces de invadir los tejidos adyacentes normales y propagarse a distancia, fundamentalmente por vía linfática o sanguínea, lo que se conoce como metástasis. La terapia del cáncer se basa en el uso simultáneo de diversas alternativas, entre las cuales se destacan: la cirugía, la quimioterapia, la radioterapia, la hormonoterapia y la bioterapia; incluyéndose dentro de esta última la IT. Las diversas terapias hasta hoy no han producido más que logros parciales, a pesar de los muchos años de investigación dedicados a ella. La IT, por sus peculiares características, puede significar una terapia con resultados superiores a los tradicionales. Los inicios de la IT de cáncer se remontan a los finales del siglo XIX, pero no fue hasta finales del siglo XX que se retomó con fuerza el tema de la IT activa específica de cáncer. Esto se debió al descubrimiento y la acumulación de una gran cantidad de evidencias sobre el reconocimiento de los tumores por el sistema inmunitario. Se han descubierto y caracterizado muchos Ag que se sobre-expresan en las células malignas comparado con el nivel de expresión en células normales. Además, se ha verificado la hipótesis de la vigilancia inmunológica del cáncer, en la cual las células presentadoras de antígenos (APC, del inglés: antigen presenting cells) reconocen células transformadas y mutágenos, y activan a otros componentes del sistema inmunitario para eliminarlos (Sahin et al., 1995; Kaplan et al., 1998). 9 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 2.1.1 Vacunas de cáncer La IT activa específica ha resurgido vigorosamente desde finales de la década de 1990. Más de 50 vacunas son evaluadas actualmente en ensayos clínicos. Esto ha estado motivado por las múltiples evidencias científicas que demuestran la existencia de la inmunovigilancia tumoral (Burnet, 1970) y la posibilidad de definir y caracterizar AAT (Ko et al., 2003). Se considera que existen dos generaciones de vacunas de cáncer, que en la actualidad co-existen en diferentes ensayos clínicos. Una primera generación se basa en preparaciones celulares, y la otra en entidades moleculares. Las vacunas de la primera generación, han utilizado células tumorales autólogas irradiadas o extractos de las mismas como fuente de Ag tumorales, en combinación con adyuvantes (Mitchell et al., 1988; Morton et al., 2002). Posteriormente, debido a la poca inmunogenicidad de las células tumorales y a la poca definición molecular de los Ag tumorales, las investigaciones desviaron su atención a la segunda generación de vacunas, basada en entidades moleculares para la terapia activa contra el cáncer (Lage et al., 2005). Ejemplo de estas vacunas son: las compuestas por péptidos derivados de varios Ag tumorales (los más usados: MART-1, gp100, tirosinasa) (Slingluff et al., 2003; Akiyama et al., 2005), las compuestas por proteínas sintéticas o purificadas del tumor, donde se incluyen proteínas de estrés térmico (HSP, del inglés: heat shock protein); oncoproteínas mutadas como K-ras y p53; glicoproteínas de la familia de las mucinas tales como MUC-1; entre otras (Naftzger et al., 1996). En los estudios de IT activa contra cáncer tienen un papel esencial los blancos tumorales, los cuales se encuentran clasificados de dos formas fundamentales: los AET y los AAT. Los AAT son aquellas moléculas las cuales se encuentran expresadas en el tejido normal que le dio origen al tumor pero están presentes en mayores cantidades en los tumores. Los AET se restringen a Ag del tumor que no se encuentra en los tejidos normales. 10 Revisión bibliográfica El microambiente tumoral es otro factor importante a valorar al diseñar una IT activa específica, ya que éste se ve afectado por la liberación de AAT y AET de la superficie de la célula tumoral. Se ha comprobado que los linfocitos recuperados de tumores proliferan menos que los de sangre periférica al mezclarlos con mitógenos o interleucina 2 (IL-2) (Miescher et al., 1986). Por otra parte, se conoce que existe gran heterogeneidad entre las células de diferentes tumores y las de un mismo tumor por lo cual algunas variantes celulares pueden evadir la acción de las terapias dirigidas contra ellas (Nowell, 1986). Se han observado además, diferencias entre las características antigénicas del tumor primario y sus metástasis (Chan et al., 1986). 2.1.2 Adyuvantes en las vacunas de cáncer El desarrollo de vacunas basadas en AAT, que constituyen a fin de cuentas moléculas propias para las cuales, en su mayoría, operan los mecanismos de deleción tímica central, encuentra su marco teórico en la teoría del daño, postulada por Polly Matzinger (Matzinger, 1994). Según esta, el real problema del sistema inmunitario no es distinguir entre lo propio y lo no propio, sino entre lo dañino y lo inocuo. De esta forma, la vacunación contra lo propio pudiera ser exitosa si ocurre en un contexto de “señales de peligro”, y en esto desempeñan un papel fundamental los adyuvantes. Por tanto, las estrategias de vacunación utilizadas contra el cáncer dependen de la calidad del Ag seleccionado (Berzofsky et al., 2004) y del adyuvante utilizado para la activación de la respuesta inmune, para obtener una eficiente activación de las APC y una consiguiente activación de los linfocitos CD4+ y CD8+ para ejercer sus funciones. Muchos productos han sido empleados como estimuladores inespecíficos del sistema inmunitario, pero tienen efectos colaterales adversos, por lo que su utilización en humanos pasa por una compleja evaluación y selección. 11 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP Entre los adyuvantes de origen bacteriano que han demostrado una excelente capacidad inmunoestimuladora se encuentran el Bacilo Calmette-Guérin (BCG) y el ácido desoxirribonucleico bacteriano no metilado con secuencias CpG (CpG). El BCG ha sido utilizado como un agente antitumoral en el tratamiento del cáncer superficial de vejiga y del melanoma (Duda et al., 1995; Alexandroff et al., 1999), y el CpG ha mostrado la capacidad de estimular potentes respuestas específicas contra varios tipos de tumores en ensayos preclínicos (Brunner et al., 2000). Los adyuvantes bacterianos, tanto las bacterias completas (BCG, Corynebacterium parvum, Salmonella minessota) o componente de bacterias, que incluyen el esqueleto de paredes celulares de bacteria, el dimicolato de trealosa, el monofosforil lípido A y el adyuvante completo de Freund, han sido ampliamente empleados por su gran capacidad de ayudar a desencadenar una respuesta inmune específica y potente contra el tumor tanto celular como de Ac. Sin embargo, al utilizarlos, algunos de ellos han provocado ulceraciones en los sitios de inyección, por lo que se ha debido eliminar el componente bacteriano en las inmunizaciones (Aucouturier et al., 2002). La capacidad de todos estos derivados de activar las células dendríticas (DC, del inglés: dendritic cells) se ha demostrado en diversos escenarios. Por ejemplo, tanto el BCG como el CpG activan a las DC humanas y murinas, y promueven el aumento en la expresión de las moléculas de MHC clase I y II, CD80 y CD86 y la secreción de IL-12 (Jakob et al., 1998; Martin-Orozco et al., 1999; Tsuji et al., 2000). Está descrito que las células necróticas, pero no las apoptóticas, liberan las HSP (Basu et al., 2000). Numerosos grupos han demostrado que diferentes HSP pueden activar a las DC y a los macrófagos, calificando así a las HSP como la primera gran familia de señales de peligro endógenas (Colaco, 1998). Los complejos HSP-péptidos derivados de tumores se 12 Revisión bibliográfica han administrado como vacunas de cáncer personalizadas y su acción ha sido evaluada en múltiples indicaciones, entre las que se incluyen el carcinoma renal, el melanoma metastásico, el cáncer gástrico y algunos tipos de leucemias (Hoos et al., 2003). Los estudios dedicados a investigar los mecanismos efectores efectivos contra el cáncer indican que los AAT son reconocidos por los CTL. Esto sugiere que la actividad de estos linfocitos es un mecanismo efector contra las células tumorales. Además, se considera el único fenómeno capaz de tener algún efecto sobre los tumores sólidos (Heath et al., 1999). Entre los adyuvantes que han demostrado facilitar el fenómeno de presentación cruzada se encuentran las secuencias de CpG, las HSP, la OmpA y el Detox (Aebischer et al., 2000). Los Ag peptídicos “chaperoneados” por las HSP purificadas activan respuestas CTL cuando se inyectaron en ratones (Arnold et al., 1995; Schild et al., 1999). También se ha demostrado que el CpG conjugado a proteínas solubles media la presentación cruzada de las mismas, mediante el cambio del mecanismo de captura de las proteínas de pinocitocis a endocitosis mediada por un receptor del conjugado (Pawelec, 2004). Otro compuesto que se ha estudiado como adyuvante para vacunas de cáncer es el αGalCer, un glicolípido sintético que potencia la respuesta de los linfocitos auxiliadores tipo 1 (Th1) estimulando a las células T asesinas naturales (NKT, del inglés: Natural Killer T cells) humanas, que pueden transactivarse con gran velocidad e inducir la producción de interferón gamma (IFNγ) por las células asesinas naturales (NK, del inglés: natural killer cells) (Carnaud et al., 1999). Los adyuvantes vacunales actualmente más utilizados en ensayos clínicos son las sales de Aluminio (Cottrez et al., 2000), el Montanide (Birkett et al., 2002) y la fracción saponina purificada de la corteza del árbol Quillaja saponaria (QS21-) (Moore et al., 1999), que estimulan la respuesta de Ac. 13 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP La bacteria Nm es una de las dos especies patógenas del género Neisseria. El meningococo, como comúnmente se le conoce, es un diplococo Gram-negativo que presenta dos membranas celulares separadas por una lámina de peptidoglicano. El 50% del exterior de la membrana externa lo constituyen moléculas anfifílicas de lipopolisacárido (LPS), endotoxina principal de esta bacteria. Otros componentes importantes de la membrana externa son las proteínas transmembranosas, entre las que se encuentran las porinas, las proteínas unidoras de transferrina, la Opa, la Opc, entre otras (Robinson et al., 1993). Los componentes aislados de la membrana externa de Nm se han estudiado como Ag en el diseño de vacunas contra la meningitis, debido a su alta inmunogenicidad. Sin embargo, hasta el momento, las únicas vacunas que han mostrado cierta efectividad consisten en preparados más complejos (Bjune et al., 1991; Sierra et al., 1991). Las VME derivadas de los meningococos se han utilizado tanto como adyuvantes o transportadores de Ag, como en la generación de vacunas antimeningocóccicas. Las vacunas antimeningocóccicas en uso consisten en VME a las que se les eliminó parte del LPS (Bjune et al., 1991; Sierra et al., 1991). Al evaluar estas preparaciones en humanos, se demostró su inmunogenicidad y su capacidad de generar títulos de Ac bactericidas. Las VME han mostrado gran capacidad adyuvante al ser conjugadas a Ag importantes para otras enfermedades. Por ejemplo, al conjugar el polisacárido tipo B de Haemophilus influenzae a las VME se indujeron Ac protectores en ratones jóvenes (Ambrosino et al., 1992). De forma general, las propiedades adyuvantes de estas vesículas y sus variantes parecen estar asociadas a la presencia del LPS. Por ejemplo, estudios de la capacidad de activar a las DC de las vesículas derivadas de bacterias salvajes y mutantes que no expresan el LPS, demostraron que estas últimas eran muy poco efectivas (Steeghs et al., 1999). 14 Revisión bibliográfica Además, sólo las bacterias salvajes estimulaban la secreción de IL-12, IL-6 y TNFα (Dixon et al., 2001; Uronen-Hansson et al., 2004). En esta dirección, también hay varios trabajos, pero con derivados vesiculares de estas bacterias y en ratones deficientes de la señalización a través de la molécula TLR4, los cuales indican que en ausencia del estímulo del LPS la activación de las DC es marginal. Los autores de estos estudios, reclaman que los componentes distintos al LPS tienen alguna contribución a este fenómeno. Sin embargo, los datos mostrados no son convincentes. Además, en ambos casos se reafirma un papel protagónico e indispensable del LPS para la polarización hacia Th1 de las células Th (AlBader et al., 2004; Rodriguez et al., 2005). 2.1.3 Los VSSP Otro adyuvante derivado de las VME lo constituyen los proteoliposomas de muy pequeño tamaño (VSSP, del inglés: very small size proteoliposomes). Estos VSSP son obtenidos a partir de la combinación de las VME de Nm y el gangliósido NAcGM3 (Estevez et al., 1999). La composición molecular de los VSSP ha sido poco caracterizada. Sin embargo, los VSSP se han evaluado como adyuvantes promotores de la respuesta de Ac específicos por el propio NAcGM3 (NAcGM3/VSSP) (Estevez et al., 1999; Carr et al., 2001) y contra otros Ag vacunales (Torrens et al., 2005; Ramirez et al., 2008; Morera et al., 2010; Aguilar et al., 2012; Fuentes et al., 2012). Se ha demostrado que los VSSP contienen ligandos de los TLR4 y TLR2, los cuales juegan un papel muy importante en las propiedades inmunomoduladoras de estos compuestos. Al evaluar in vitro el efecto de los VSSP sobre las DC, se demostró que éstos constituyen un estímulo tan potente como el LPS en la inducción de la maduración de estas células. Los VSSP aumentan la expresión de las moléculas coestimularorias CD40, CD80, CD86 y la expresión de la molécula MHC-II (Mesa et al., 2004), potencian la producción de las citocinas IL-1β, IL-12, IL-6 e IL-18, mientras que disminuyen la secreción de IL-10 (Mesa et 15 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP al., 2004; Venier et al., 2007). Experimentos con células T transgénicas Ag-específicas demostraron que las DC tratadas con los VSSP inducen un fenotipo Th1 en células T vírgenes CD4+ y facilitan la presentación cruzada de Ag protéicos (Mesa et al., 2004; Mesa et al., 2006b). Por otra parte, los VSSP potencian la respuestas CTL en ratones sanos (Mesa et al., 2006a), generan similares respuestas CTL en ratones portadores de tumor y sanos (Fernandez et al., 2011), y expanden las células T CD8+ con fenotipo de memoria, a la vez que protegen la respuesta CTL en ratones leucopénicos (Oliver et al., 2012). Particular atención se ha puesto en la interacción de los VSSP con las células supresoras de origen mieloide (MDSC, del inglés: myeloid-derived suppressor cells) identificadas en ratones por la co-expresión de los marcadores CD11b y Gr1 (Fernández et al., 2014a). El tratamiento de ratones sanos con los VSSP aumenta la frecuencia de células esplénicas CD11b+Gr1+, sin embargo las células expandidas son poco supresoras (Fernandez et al., 2011). Adicionalmente se conoce que los VSSP reducen la capacidad supresora de las MDCS inducidas por los tumores, afectan su migración al microambiente tumoral y promueven su diferenciación a DC maduras (Fernandez et al., 2011; Fernández et al., 2014b). 2.1.4 Mecanismos de acción de las vacunas de cáncer. El descubrimiento e identificación de los AET y los AAT hizo posible atacar los tumores de manera específica mediante Ac o células citotóxicas (Rosenberg, 1999; Rosenberg et al., 2004). La unión de los Ac al Ag tumoral puede resultar en la activación de la cascada del complemento. Este fenómeno trae como consecuencia la inflamación (incluye opsonización, 16 Revisión bibliográfica activación de leucocitos y macrófagos, y el incremento en la permeabilidad vascular) y la formación del complejo de ataque a la membrana que conduce a la citotoxicidad dependiente de complemento (CDC, del inglés: cytotoxicity dependent complement). Estos eventos pueden promover la eliminación de células tumorales circulantes y las micrometástasis. Los Ac IgM son los mayores activadores del complemento debido a su naturaleza pentamérica. En el caso de los Ac IgG, la propiedad de mediar CDC la tienen las IgG1 e IgG3 en humanos y las IgG2a e IgG3 en ratones. Los receptores Fc de estas subclases de IgG son además los blancos primarios para las células efectoras que median la citotoxicidad celular dependiente de anticuerpos (ADCC, del inglés: antibody-dependent cellular cytotoxicity). Los receptores FCδR1 (CD64), FCδRII (CD32) y FCδRIII (CD16), expresados en un amplio rango de células que incluyen células NK, células T y células del linaje mieloide, se unen a los Ac que reconocen Ag sobre la superficie del tumor, con la consiguiente activación del mecanismo citotóxico intrínseco de estas células efectoras. (Livingston et al., 1997b). Debido al gran potencial que tiene la respuesta celular para erradicar células tumorales, una gran cantidad de estudios clínicos y preclínicos están enfocados a la inducción de respuesta celular contra el tumor, donde se incluye la generación de NK y CTL específicos. La generación de CTL es un proceso complejo que requiere de al menos dos señales. La primera es el reconocimiento, mediado por la interacción del receptor de células T CD8+ (TCR, del inglés: T cell receptor) y los péptidos presentados en el MHC I de las APC. La segunda señal es la coestimulación, mediada por determinadas moléculas en la APC y en las células T CD8+ o por citocinas derivadas de células Th que se activan al reconocer péptidos presentados en el MHC II de las APC y moléculas coestimulatorias. En sujetos con tumor, los Ag tumorales pueden ser presentados por el tumor, pero en la mayoría de los casos, la interacción del tumor con el sistema inmunitario produce tolerancia en lugar de 17 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP activación debido a, entre otros factores, que las señales coestimulatorias requeridas para la activación de la células T no son las adecuadas (Dutoit et al., 2002). El objetivo principal de las estrategias vacunales es convertir a la célula T anérgica en célula efectora contra el tumor. Sin embargo, en pocos estudios se ha correlacionado la inducción de estas células y la respuesta clínica. En este sentido, en un ensayo realizado con una vacuna peptídica en pacientes de melanoma se correlacionó el incremento de la respuesta CTL en sangre periférica contra los Ag MAGE-1 y MAGE-3 con una respuesta clínica favorable en un paciente tratado (Jager et al., 2001). En otro estudio con vacunas de péptidos derivados de Melan A/MART-1, tirosinasa o gp100, se observó una asociación entre la respuesta clínica y la respuesta CTL específica (Jager et al., 1999). Algunos pacientes con respuesta clínica favorable mostraron además desarrollo de vitiligo. En otro trabajo publicado en pacientes de cáncer de colon con metástasis hepáticas, inmunizados con la vacuna Oncophage, se demostró la activación de células T CD8+ específicas por los Ag tumorales. El incremento en la frecuencia de estas células después de la vacunación correlacionó con un aumento en la supervivencia de estos pacientes (Mazzaferro et al., 2003). El fenómeno de autoinmunidad provocado por las terapias se ha asociado con un aumento de la supervivencia de los pacientes. Este fenómeno se ha visto más frecuentemente en pacientes de melanoma, lo que sugiere que una respuesta inmune potente puede prolongar la supervivencia pero también producir autoinmunidad (Gogas et al., 2006). Por otra parte se ha señalado que la diversificación del reconocimiento antigénico acontecida con algunas estrategias vacunales es beneficioso para la respuesta antitumoral. Este fenómeno muy bien descrito en los modelos de enfermedades autoinmunes, se inicia con la respuesta contra un simple epítopo antigénico. Este epítopo es presentado en el 18 Revisión bibliográfica MHC II de las APC, lo cual activa células Th y células T efectoras que causan la inflamación crónica y la muerte de la célula blanco. El debris celular, que contiene otros Ag, es capturado por las APC y presentado de forma cruzada por el MHC I, con la consiguiente activación de nuevas células T CD8+ específicas por otros epítopos en el mismo Ag (diversificación intramolecular) o en otras moléculas (diversificación intermolecular). Uno de los trabajos que abordan este fenómeno fue publicado por El-Shami y cols. que vacunaron ratones retados con EG7-OVA (línea de timoma EL-4 transfectada con la ovalbúmina) con el péptido OVA257-264 (de la ovalbúmina) y mostraron una respuesta CTL protectiva frente al reto con EL-4 que no expresa OVA (el-Shami et al., 1999). Otro de los mecanismos prominentes para activar al sistema inmunitario contra los tumores es la muerte celular inmunogénica (MCI). Dependiendo del estímulo inicial, la muerte de las células de cáncer puede ser inmunogénica o no inmunogénica. La MCI involucra cambios en la superficie celular (exposición de calreticulina) y la liberación de mediadores solubles (ATP y la proteína nuclear HMGB1), que ocurre en una secuencia temporal definida, y operan sobre una serie de receptores expresados por DC (CD91, P2R7 y TLR4), y facilita el reclutamiento de DC al sitio tumoral (estimulado por ATP), la captura de Ag tumorales (estimulado por calreticulina), y la presentación óptima de Ag a las células T (estimulado por HMGB1) (Kroemer et al., 2013). Este proceso resulta en una potente respuesta inmune, mediada por IFNδ, y dependiente de IL-1β e IL-17; donde participan células T δγ y CTL. Esto significa que el tumor o las células moribundas del paciente operan como una vacuna que estimula una respuesta tumor específica, que puede controlar y en ocaciones erradicar las células de cáncer residuales. Varios tratamientos antitumorales provocan MCI, entre los que se encuentran quimioterapéuticos como las antraciclinas (Casares et al., 2005), la ciclofosfamida (Schiavoni et al., 2011), la irradiación (Perez et al., 2009), el ácido 19 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP polyinosinoico-polycitidílico (poli I:C) (Kubler et al., 2011), así como los AcM anti-EGFR 7A7 (Garrido et al., 2011) y el cetuximab (Lee et al., 2011). 2.2 2.2.1 Gangliósidos Localización, características estructurales, biosíntesis y función Los gangliósidos son constituyentes característicos de la membrana plasmática de las células de los mamíferos y, prácticamente, de todos los vertebrados (Daniotti et al., 2011). En 1942, Ernest Klent aplicó el término gangliósidos a unos lípidos nuevos que aisló de las células ganglionales del cerebro. Están particularmente en el cerebro, donde son el 10-12% de los lípidos totales y el 20-25% de los lípidos de la capa externa de las membranas neuronales. Fuera de ahí aparecen en muy bajos niveles en el resto de los tejidos animales, constituyendo el 1-2% de los lípidos totales, y se concentran en la cara externa de la membrana plasmática, en los llamados parches lipídicos (Sonnino et al., 2007). Los que aparecen en la leche son derivados de las células secretorias de la glándula mamaria y tienen importancia nutricional para los recién nacidos, pero ahí han sido poco caracterizados, así como sucede en general con los provenientes de las comidas. No aparecen fuera del reino animal. Son compuestos de naturaleza anfipática (Voet D, 1995), al igual que otros lípidos formadores de membranas. Por sus características estructurales pertenecen a la familia de los glicoesfingolípidos (GEL) y están constituidas en su porción lipofílica por la ceramida (una esfingosina unida a un ácido graso), mientras que la región hidrofílica expuesta al espacio intercelular posee un oligosacárido lineal de hasta cuatro residuos neutros, al que se encuentran unidos uno o más residuos de ácido siálico (AS). Se distribuyen en la superficie celular de manera tal que las cadenas de azúcares sialiladas quedan expuestas hacia el exterior celular (Sonnino et al., 2007) y presentan patrones de expresión 20 Revisión bibliográfica específicos, en dependencia del tipo de célula y la etapa de diferenciación en que se encuentren (Reglero et al., 1982; de Mello Coelho et al., 2004). La biosíntesis de los GEL comienza a partir de la ceramida (Cer). Lo más común es que se adicione una molécula de glucosa a la Cer por la proteína transmembrana glucosil-ceramida sintasa Tipo I, formando la glucosil-ceramida (GlcCer). Se adiciona entonces una galactosa (Gal) a la GlcCer por la β-1,4-galactosil transfrasa formando la lactosil-ceramida (LacCer), en el lumen del aparato de Golgi (Durrant et al., 2012). La LacCer es el aceptor de varias transferasas que generan las principales series de GEL: globo-, isoglobo-, lacto-, neolacto-, y ganglio-GEL. La LacCer es convertida, por la adición secuencial de diferentes unidades de monosacáridos, a miembros neutrales o acídicos de la serie ganglio, proceso mediado por proteínas integrales de membrana glicosil-transferasas Tipo II. Los gangliósidos son miembros acídicos de la serie ganglio, siendo así la LAcCer la base de todos los gangliósidos, los cuales son elongados posteriormente por la adición de azúcares y AS. Este proceso involucra actividades consecutivas de enzimas sialil-transferasas y glicosiltransferasas. Estas enzimas están unidas a la membrana del aparato de Golgi, en una secuencia que corresponde con el orden de adición de varios de los componentes carbohidratos. Se han descrito varias sialil-transferasas que usan AS-citidin-monofosfato (CMP) para transferir el residuo de AS a la cadena oligosacarídica. Alternativamente, la Gal puede ser adicionada a la ceramida, formando galactosil-ceramida (GalCer), que da lugar a la síntesis de estructuras menos frecuentes, como el gangliósido GM4 y los sulfátidos (Daniotti et al., 2011). La complejidad estructural de estos compuestos ha estimulado la elaboración de varias nomenclaturas. Son ellas la de Svennerholm (Svennerholm, 1963), la establecida por la IUPAC-IUB (1978), y la de Wiegandt (Wiegandt et al., 1969). La establecida por Svennerholm es la más utilizada en la designación de los gangliósidos y entre los más 21 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP frecuentes están: GM3, GD3, GM2, GD2, GM1, GD1a, GD1b, GT1a. Las letras M, D, T y Q se refieren a la cantidad de residuos de AS: M-mono, D-di, T-tri, y Q-tetra-sialogangliósidos; y los números 1, 2, 3, 4 se refieren al orden de migración en una cromatografía de placa delgada (TLC): GM3 > GM2 > GM1. En general, la estuctura de la Cer tiende a ser relativamente simple. La esfingosina es usualmente la principal base esfingoide, acompañada por un ácido graso, los cuales pueden ser de longitudes variables (entre 16-24 carbonos) con pocas o ninguna poliinsaturaciones. La naturaleza de la Cer es importante para su función biológica, pero es el motivo carbohidrático quien más ha determinado en su función (Hakomori, 1996). Los AS constituyen una familia muy diversa de aproximadamente 50 tipos de carbohidratos ácidos alfa-ceto aldónicos con un esqueleto carboxilo de 9 carbonos, presentes en glicoproteínas y glicoesfingolípidos (Angata et al., 2001). La gran diversidad de los AS está dada por modificaciones del grupo amino del C5 (con acetil o glicolil), o de los grupos hidroxilo, en los C4, C7, C8 y C9, por acetato, lactato, sulfato o ésteres de fosfato o ésteres de metilo (Angata et al., 2001; Wipfler et al., 2011). Los dos AS más frecuentes en mamíferos son las variantes N-acetiladas (NAc) y N-glicoliladas (NGc) del ácido neuramínico (ácidos NAc- y NGc-neuramínicos, Neu5Ac y Neu5Gc) (Varki et al., 2001). Las funciones biológicas en las que se encuentran implicados los gangliósidos han sido ampliamente abordadas en la literatura. En la superficie celular interactúan con ligandos solubles u organismos enteros, independientemente de su pequeño tamaño en comparación con glicoproteínas también expresadas en la membrana (Fishman, 1986; Siebert et al., 1996). Estas moléculas son también receptores específicos de exotoxinas bacterianas (Holmgren et al., 1973; van Heyningen, 1980), virus (Schnaar, 1991) y bacterias (Karlsson, 1989; Teneberg et al., 1990); así como pueden constituir receptores de 22 Revisión bibliográfica interferones (Andrade et al., 2006), actuar como mediadores de las interacciones célulacélula, donde se destaca su participación en la transmisión de impulsos nerviosos (Thomas et al., 1990). La interacción de gangliósidos con receptores de factores de crecimiento con actividad enzimática asociada (Chung et al., 2009; Guan et al., 2011; Huang et al., 2013; Wang et al., 2013; Kim et al., 2014) vincula a los gangliósidos con la maquinaria transduccional. El gangliósido NAcGM3 está involucrado en la actividad fosfatasa del receptor del factor de crecimiento epidérmico (EGF), al inhibir el crecimiento en líneas celulares que expresan este último con Ac anti-EGF (Rebbaa et al., 1996; Alves et al., 2002; Liu et al., 2008; Chung et al., 2009; Huang et al., 2013) Se ha descrito su papel en la regulación de los eventos relacionados con la proliferación celular a través de la modulación de proteínas de la membrana que funcionan como receptores, transductores y transportadores (Hakomori, 1985). Su estructura sufre diversas alteraciones durante el desarrollo celular, la diferenciación, la ontogenia, las transformaciones oncogénicas y el envejecimiento, lo cual evidencia una acción reguladora por parte de estas moléculas sobre las funciones asociadas al crecimiento (Hakomori, 1985; Saito, 1993) y la regulación del ciclo celular (Spiegel et al., 1996) Los gangliósidos, han sido también descritos como inmunosupresores de la respuesta inmune (Shurin et al., 2001). 2.2.2 Los gangliósidos como antígenos asociados a tumores Los gangliósidos están ampliamente distribuidos en gran cantidad de tumores de diferentes orígenes. La conformación y configuración de los gangliósidos cambia durante el proceso de malignización, originando patrones de glicosilación alterados, fenómeno conocido como glicosilación aberrante y que forma parte de los cambios moleculares asociados a la transformación oncogénica que han sido descritos durante las últimas décadas (Hakomori, 23 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 1985; Reis et al., 2010). Esta es una de las principales razones que ha permitido su empleo como blancos de la terapia contra el cáncer (Hakomori, 1985). Estos cambios que sufren durante el desarrollo tumoral han permitido identificarlos como AAT. Se conoce que aunque la región hidrofóbica de los gangliósidos es poco inmunogénica, cualquier cambio en la misma puede variar la orientación o el ángulo de la cadena sacarídica, exponiéndola para la neosíntesis o la degradación por las enzimas glicosiltransferasas y desglicosilantes, lo cual puede crear nuevos determinantes antigénicos en la superficie de la células tumorales (Wiegandt, 1980). Las primeras evidencias de la asociación entre los gangliósidos y el cáncer provienen de estudios de los niveles de expresión de lípidos en los melanomas humanos realizados por Portoukalian y cols. (Portoukalian et al., 1979), en los que se demostró la particular abundancia del gangliósido GD3. Durante los años posteriores, el GD3 fue identificado como uno de los Ag reconocidos por anticuerpos monoclonales (AcM) generados en ratones inmunizados con células de melanoma maligno humano (Pukel et al., 1982). Por esa misma época, los intentos realizados por varios grupos con células tumorales autólogas o heterólogas, en la inmunización de pacientes de melanoma (Livingston et al., 1982), y la identificación serológica de los Ag reconocidos, contribuyeron al interés en los gangliósidos como marcadores tumorales y blancos para la terapia antitumoral. Las células utilizadas en las inmunizaciones habían sido seleccionadas por su elevada expresión de glicoproteínas, proteínas y gangliósidos. Sin embargo, los únicos Ag reconocidos por los Ac producidos por varios pacientes fueron los gangliósidos GM2 y GD2. La inmunogenicidad particular de estas moléculas en pacientes con melanoma ha sido demostrada una y otra vez desde entonces (Cahan et al., 1982; Tai et al., 1983; Livingston et al., 1989a; Livingston et al., 1989b; Portoukalian et al., 1991). 24 Revisión bibliográfica La glicosilación aberrante de manera general se manifiesta en la síntesis incompleta de algunos compuestos (Itai et al., 1991; Kannagi, 2004; Kawamura et al., 2008; Wei et al., 2010; St Hill et al., 2011), en la neosíntesis de estructuras aberrantes que resulta en la acumulación de ciertos carbohidratos en altas densidades en la superficie (Kannagi et al., 2008). Este fenómeno ocurre como resultado de alteraciones metabólicas que incluyen la pérdida de enzimas, la inducción de nuevas enzimas o el incremento anormal en la actividad de enzimas ya existentes que puedan estar involucradas, tanto en las vías de síntesis de los glicocompuestos, como en su degradación. Este proceso parece activar diferentes enzimas, de lo cual resulta en una expresión alterada del patrón de gangliósidos presente en los tumores (Ogawa et al., 1997; Koike et al., 2004). La fucosilación anormal es otra de las modificaciones comunes de las glicoproteínas presentes en las células tumorales, producto de la desregulación de enzimas involucradas en la adición de residuos de fucosa a los glicanos (Miyoshi et al., 2008). Las sobrexpresión de los glicoesfingolípidos, en particular de los gangliósidos, es uno de los cambios cuantitativos en la composición de las células neoplásicas, e incluye alteraciones en su densidad y distribución en la superficie celular (Dubois et al., 1986; Tsuchida et al., 1989; Kono et al., 1990; Hamilton et al., 1993). Se ha descrito que el GD2 se expresa en la superficie de los tumores de origen neuroectodérmico, incluyendo el neuroblastoma y el melanoma; así como en glioma y en el cáncer de pulmón de células no pequeñas (Mujoo et al., 1987). Su expresión es abundante en el 100% de los tumores de neuroblastoma, independientemente de la edad (Wu et al., 1986). Sin embargo, su expresión en los tejidos normales es baja y se limita a las neuronas y los melanocitos normales (Tsuchida et al., 1984; Tsuchida et al., 1989). Adicionalmente, el gangliósido GD2 se ha visto asociado al crecimiento celular, la diferenciación y la apoptosis (Yoshida et al., 2002). El gangliósido GD3 ha sido descrito como un marcador de melanoma debido a su sobrexpresión 25 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP (Tsuchida et al., 1989) en este tumor y su baja o restringida expresión en los melanocitos. Se ha descrito además, sobrexpresión del gangliósido GM2 en varios tipos de tumores, entre los que se incluyen el melanoma maligno y el neuroblastoma (Portoukalian et al., 1979). Los gangliósidos NAcGM3 y GM1 son de amplia distribución corporal (Tai et al., 1988). Los melanocitos normales contienen, predominantemente, al gangliósido NAcGM3, sin embargo, en el melanoma el gangliósido NAcGM3 alcanza el 80 % del total de gangliósidos de las células tumorales. En tanto, la expresión del gangliósido GD3 se incrementa después de la transformación neoplásica. Cuando el melanoma progresa a variantes altamente metastásicas hay una variación pronunciada en la relación de gangliósidos NAcGM3:GD3 (Ravindranath et al., 1991). La proporción de gangliósidos NAcGM3:GD3 se invierte durante la desdiferenciación de melanocitos (15:1) a células de melanoma (1:16) (Ravindranath et al., 1989). La presencia en estos tumores de cantidades menores de los gangliósidos GM2, GD2 y 9-O-acetil GD3 puede entonces explicarse por la alteración producida en la proporción de gangliósidos NAcGM3:GD3. A pesar de que muchos de los gangliósidos mencionados se expresan en tejidos normales, muchos de éstos han sido evaluados como blancos para la IT pasiva y activa de pacientes con melanoma. La evaluación de la presencia de gangliósidos tales como GM2, GD2, NAcGM3 y GD3 en tumores humanos, así como su distribución en tejidos normales, ha sido posible en gran medida por el uso de AcM dirigidos contra estos gangliósidos (Durrant et al., 2012). Otros estudios han demostrado que algunos gangliósidos pueden ser liberados al microambiente tumoral. Por ejemplo, se ha descrito que los gangliósidos GM2, NAcGM3 y GD1a son liberados por líneas de meduloblastoma (Chang et al., 1997). Los gangliósidos así secretados pueden ser internalizados por células T activadas, desencadenando su 26 Revisión bibliográfica apoptosis y propiciando que los tumores puedan evadir la acción del sistema inmunitario (Sa et al., 2009). 2.2.3 La función N-glicolilada en tumores humanos Dentro de los cambios en la glicosilación asociados a las células tumorales se ha descrito la presencia de derivados sialilados en gran variedad de tumores humanos. En los vertebrados, los glicanos usualmente terminan con AS, que funcionan como marcadores de lo propio y que pueden ser reconocidos por varios receptores (nombrados Siglecs), que median la comunicación inter e intracelular (Paulson et al., 2012). Los glicoconjugados portadores de la variante NGc del AS se encuentran distribuidos ampliamente en la mayoría de los mamíferos, pero es difícil su detección en tejidos normales de humanos y pollos (Feeney et al., 1960; Rosner, 1975; Muchmore et al., 1987; Brinkman-Van der Linden et al., 2000). Sin embargo, un conjunto de investigaciones demuestran su carácter de Ag específicos de células tumorales en estas dos especies (Kikuta, 1985; Hirabayashi et al., 1986; Higashi et al., 1987; Shida et al., 2010). Los humanos no pueden sintetizar de novo el AS Neu5Gc por hidroxilación oxidativa del AS Neu5Ac, debido a una pérdida irreversible del exón del gen Cmah que codifica para la enzima mono fosforil citidina-N-Acetil AS hidroxilasa (CMAH) que participa en esta transformación (Chou et al., 2002). De forma similar, ratones modificados genéticamente para que no expresan el gen de la hidroxilasa (Cmah-/-) son incapaces de sintetizar el AS Neu5Gc (Hedlund et al., 2007; Naito et al., 2007), lo cual sugiere que la fuente de AS Neu5Gc presente en los tejidos de humanos es exógena (Tangvoranuntakul et al., 2003). Los tejidos humanos sanos han sido extensivamente evaluados para determinar la presencia de AS Neu5Gc. La tinción inmuno-histoquímica (IHQ) con Ac específicos contra el AS Neu5Gc generados en pollos estableció que los niveles de AS Neu5Gc son bajos en 27 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP los tejidos normales, y que aparece fundamentalmente en epitelio y endotelio de vasos sanguíneos o en células secretoras del epitelio de pulmón, piel, colon, próstata, útero, riñón, bazo, testículos, páncreas, hígado y estómago fetal (Tangvoranuntakul et al., 2003; Diaz et al., 2009). El AcM murino 14F7 (IgG1), generado por inmunización con el gangliósido Neu5Gc-GM3 (NGcGM3) conjugado hidrofóbicamente a lipoproteínas de muy baja densidad (VLDL, del inglés: very low density lipoprotein) es otra herramienta que permite el estudio de la presencia de AS Neu5Gc en los tejidos humanos (Carr et al., 2000). Este AcM ha mostrado ser altamente específico por el gangliósido NGcGM3, sin reactividad cruzada con el Neu5Gc-GM2 (NGcGM2) ni con su contraparte Neu5Ac-GM3 (NAcGM3) ni con glicolípidos sulfatados (Carr et al., 2000; Rojas et al., 2013). En un estudio realizado en ratones por Roque-Navarro y cols. (2008) se observó que el AcM 14F7 induce la muerte celular de la línea tumoral murina de leucemia L1210 y no de las células normales (linfocitos B y T CD4) que expresan el gangliósido NGcGM3, por un mecanismo independiente de complemento que provoca la formación de lesiones en la membrana de la célula tumoral, sugiriendo un fenómeno tipo oncosis (Roque-Navarro et al., 2008) La tinción de tejidos humanos con el AcM 14F7 ha permitido identificar la presencia del gangliósido NGcGM3 en muy bajos niveles en muestras de tejidos sanos de riñón y ovario, pero no en testículo, próstata ni vejiga (Blanco et al., 2011a). Todos estos trabajos revisados señalan que en los tejidos normales, solamente se expresa la variante NAc de los gangliósidos, pero no la variante NGc (Ozawa et al., 1992b). En contraste a la limitada evidencia de la presencia del AS Neu5Gc en los tejidos normales humanos, su presencia en los tumores ha sido ampliamente descrita (Malykh et al., 2001). El AS Neu5Gc fue detectado, al usar inmunofluorescencia, en tumores de mama, colorectal, 28 Revisión bibliográfica nasofaríngeo, uterino, leucemia, y linfoma maligno, así como en ovario, páncreas y teratoma (Higashi et al., 1988). Utilizando la inmunotinción sobre TLC, el AS Neu5Gc ha sido detectado en colon, melanoma, retinoblastoma, tumores del saco de yolk y en el carcinoma de mama (Amon et al., 2014). Algunos investigadores han obtenido resultados que confirman la existencia de variantes Nglicoliladas de los gangliósidos GM2, NAcGM3 y GD3 en las células tumorales procedentes de melanomas, retinoblastomas y de tumores colorectales, de pulmón y provenientes de la vesícula seminal (Higashi et al., 1987; Hirabayashi et al., 1987; Kawashima et al., 1993; Hayashi et al., 2013). El AcM 14F7 ha permitido detectar al gangliósido NGcGM3 en muestras de tumores de mama y melanoma (Carr et al., 2000; Oliva et al., 2006); en tumores epiteliales malignos del sistema digestivo como los adenocarcinomas de colon, estómago y páncreas, así como en tumores gastroesofágicos, carcinomas epidermoides del recto, y en el carcinoma hepatocelular (Blanco et al., 2011a); en tumores pediátricos como el neuroblastoma y de la familia de sarcomas de Ewing (Scursoni et al., 2011); en muestras de cáncer de pulmón de células no pequeñas (Blanco et al., 2012); en muestras de melanoma mucosal oral (Zhong et al., 2012); así como en linfomas, linfonodos metastásicos y otros sitios metastásicos (Blanco et al., 2013). 2.2.4 Inmunosupresión inducida por los gangliósidos La interacción entre las células del tumor y las células del sistema inmunitario es un fenómeno crítico para el crecimiento tumoral. La hipótesis de que los gangliósidos juegan un papel fundamental en la supresión de la respuesta inmune antitumoral está soportada por varios estudios que demuestran su liberación al microambiente tumoral por parte de las células tumorales (Black, 1980). Los gangliósidos liberados de manera espontánea y en 29 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP cantidades considerables en forma de monómeros, micelas y vesículas de membrana (Kong et al., 1998; Dolo et al., 2000) poseen cadenas cortas de ácido graso en la porción Cer (Li et al., 1991; Li et al., 1997). Esto favorece no sólo su liberación al microambiente tumoral, sino además, su inserción en la membrana de las células del hospedero, e induce los fenómenos de supresión inmune (Chang et al., 1997). Estudios realizados en pacientes de melanoma han demostrado el incremento en el suero de la concentración de los gangliósidos mayoritarios NAcGM3 y GD3, hasta valores entre 20 y 50 μg/mL (Portoukalian et al., 1993). Además, la concentración de estos gangliósidos se incrementa por encima de estos valores en el microambiente tumoral y en los ganglios linfáticos de pacientes con este tipo de tumor (Hamilton et al., 1993). Junto con estas observaciones muchos estudios han mostrado que los gangliósidos liberados por el tumor inhiben múltiples pasos de la respuesta inmune celular in vitro. En este sentido se ha visto la inhibición de las células T auxiliadoras (Th, del inglés: T helper) (Offner et al., 1987; Chu et al., 1995), de la citotoxicidad de las células (Grayson et al., 1992) y de la activación de los linfocitos T y B, ya sean estimulados por mitógeno o de manera Ag específico (Dyatlovitskaya et al., 1987; Ladisch et al., 1992). Se ha observado además que estos gangliósidos bloquean la producción del factor de necrosis tumoral alfa (TNFα, del inglés: tumor necrosis factor), así como la presentación de Ag por los monocitos humanos (Ladisch et al., 1984; Ziegler-Heitbrock et al., 1992). Ellos inhiben además la generación de las DC funcionalmente activas (Shurin et al., 2001). Los mecanismos por los cuales los gangliósidos llevan a cabo la supresión del sistema inmunitario no están totalmente esclarecidos aún, pero existen numerosas evidencias que confirman su papel inmunosupresor. Uno de estos estudios es el descrito por de León y cols. (2006), que demostraron la capacidad del gangliósido NGcGM3 de modular 30 Revisión bibliográfica negativamente la expresión de la molécula CD4 en linfocitos T de ratón y humanos, afectación que se se correlaciona con su inserción en la membrana plasmática de estas células. Además, estos autores encontraron que el gangliósido NGcGM3 modifica la funcionalidad de los linfocitos Th CD4+CD25-lo que no está mediado por un incremento en la actividad supresora de los linfocitos T reguladores naturales (de Leon et al., 2006). Se ha demostrado también que la exposición de esplenocitos de ratón a algunos gangliósidos, conduce a una reducción de la transcripción de los genes que codifican para las citocinas asociadas a las células Th1: IL-2 e IFNα. Sin embargo, la transcripción de los genes que codifican para las citocinas asociadas a las células Th tipo 2 (Th2), como la IL-4, así como para la citocina inmunosupresora IL-10, no se afecta por el tratamiento de las células del bazo con los gangliósidos (Irani et al., 1996). Estos resultados sugieren que los gangliósidos liberados por los tumores modifican el balance de la respuesta inmune antitumoral de un patrón Th1 a un patrón Th2, lo que provoca una reducción en la respuesta inmune celular que puede resultar crítica para la eliminación del tumor (Birkle et al., 2003). Las DC también son afectadas por los gangliósidos. Se ha informado que los gangliósidos NAcGM3 y GD3 purificados de melanoma humano inhiben la diferenciación fenotípica y funcional de las DC derivadas de monocitos, e inducen su apoptosis (Peguet-Navarro et al., 2003; Bennaceur et al., 2006). Se ha descrito además, que el gangliósido NGcGM3 afecta la diferenciación de las DC, así como su maduración ante una señal inflamatoria (de Leon et al. 2008). El gangliósido GD1a es capaz de inhibir la función de DC, las que tienen un papel crucial en la generación de la respuesta inmune. Las DC humanas incubadas con el gangliósido GD1a muestran una expresión constitutiva reducida del CD40 y del CD80 pero no del CD86, así como una marcada inhibición de la secreción de las citocinas IL6 e IL12 (Caldwell et al., 2003). 31 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP Los linfocitos B también resultan afectados por los gangliósidos, los cuales inhiben su proliferación y la producción de Ac (Berenson et al., 1991; Frugoni et al., 1994). De hecho el gangliósido GM2 inhibe la producción de Ac por células B linfoblastoides humanas y por células B activadas in vivo, en alguna medida como consecuencia del bloqueo de la producción autocrina de IL10 (Kimata et al., 1996). 2.2.5 Papel del gangliósido NGcGM3 en la progresión tumoral El papel de los gangliósidos en la progresión tumoral no está ligado sólo a su demostrado efecto supresor sobre la actividad del sistema inmunitario. Los gangliósidos asociados a los tumores tienen un papel importante en la ocurrencia de los eventos relacionados con el establecimiento de los tumores y las metástasis (Cheresh, 1985; Birkle et al., 2003). La relación del gangliósido NGcGM3 y la progresión de los tumores ha sido poco abordada. Los primeros estudios en esta dirección fueron realizaados por de León y cols. (2006), que estudiaron la influencia del gangliósido NGcGM3 en el modelo celular de mieloma de ratón P3X63. Estos autores demostraron una disminución del crecimiento de los tumores desarrollados en los animales por el tratamiento de las células tumorales P3X63 con un inhibidor de la glucosil-ceramida sintasa (D-PDMP, del inglés: D-threo-1-phenyl-2decanoylamino-3-morpholino-1-propanol), efecto que se revirtió eliminando la población de células T CD4+ (de Leon et al., 2006; de Leon et al., 2008). Posteriormente, Gabri y cols. (2009) demostraron que las células tumorales de ratón de melanoma B16 y de carcinoma mamario F3II, incubadas con mucinas ricas en AS Nglicolilados, incorporan este AS y expresan, de manera transciente, el gangliósido NGcGM3 en la membrana. Ambas líneas tumorales implantadas in vivo en ratones mostraron un aumento del número de metástasis experimentales en el pulmón con respecto a los 32 Revisión bibliográfica controles carentes del gangliósido NGcGM3; así como una disminución de la carga tumoral necesaria para lograr el implante del tumor SC (Gabri et al., 2009). Estudios ulteriores, con el carcinoma de ratón de pulmón de Lewis 3LL-D122, demostraron que la preincubación de estas las células tumorales con AS N-glicolilado, da lugar a células 3LL-D122 que expresan el gangliósido NGcGM3 y las mismas forman más nódulos metastásicos pulmonares, que sus controles no tratados (Segatori et al., 2012b). En cambio, este mismo grupo de autores demostraron que las células de melanoma murino B16 transfectadas con el gen Cmah expresan el gangliósido NGcGM3, pero disminuyen su proliferación y adhesión in vitro, y muestran una menor habilidad de desarrollar tumores en ratones, comparadas con la línea celular parental (Segatori et al., 2012a). Adicionalmente, Casadesús y cols. (2013) mostraron que los cambios en la biología celular debido a la disminución de la síntesis de AS N-glicolilado disminuyen la tumorigenicidad tanto in vitro como in vivo. Estos autores demostraron que la línea parental de leucemia linfocítica murina L1210, la cual expresa altos niveles del gangliósido NGcGM3, tiene una mayor ventaja de crecimiento en ratones comparada con la línea L1210 modificada, que carece permanentemente de la expresión de la enzima CMAH y muestra una disminución notable del gangliósido NGcGM3 con un aumento del gangliósido NAcGM3. La línea L1210 modificada muestra una disminución de la frecuencia de tumores desarrollados por su implante en ratones (Casadesus et al., 2013). 2.3 Vacunas de cáncer basadas en gangliósidos Los cambios que ocurren en la expresión de los gangliósidos durante las transformaciones oncogénicas los convierten en candidatos atractivos para la IT del cáncer. Muchos aspectos complican el uso de los gangliósidos como vacunas de cáncer. En primer lugar, su aislamiento de las células tumorales es muy difícil, debido a su heterogeneidad en 33 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP la superficie celular. Una solución a este problema es el empleo de la síntesis química para la obtención de los mismos (Buskas et al., 2009). En segundo lugar, los gangliósidos son Ag T-independientes, dando lugar a respuestas de Ac de baja afinidad y de corta duración, en la que predomina el isotipo IgM a pesar de repetidas inmunizaciones (Livingston, 1995a). Adicionalmente, como los gangliósidos son Ag propios, expresados en bajos niveles en las células normales, son tolerados por el sistema inmunitario, con inmunogenicidad baja. Esta tolerancia se refuerza además porque se liberan hacia el microambiente del tumor y hacia la circulación sistémica, inactivando las células del sistema inmunitario. Además, la presencia en las células tumorales de otros autoantígenos rodeando a los gangliósidos contribuye a la poca inmunogenicidad de estos últimos (Livingston et al., 1997b). La posibilidad de romper esta tolerancia en individuos normales y en pacientes con tumores, ha sido demostrada por la presencia en el suero de Ac naturales específicos contra estos Ag propios y de las células B que los producen, lo que indica que los linfocitos B antigangliósidos no han sido eliminados por deleción clonal y se encuentran funcionalmente silentes (Marcus, 1984; Alfonso et al., 1996). Varias estrategias vacunales se han llevado a cabo con el objetivo de eliminar la tolerancia y aumentar la inmunogenicidad de los gangliósidos frente a los tumores (Tabla 1). Uno de los candidatos vacunales evaluados es el GD2-KLH. La vacunación de pacientes con este preparado dio lugar a una pequeña respuesta de Ac, mientras que la variante GD2-lactona-KLH logró respuesta de Ac en el 83% de los pacientes tratados. Los pacientes inmunizados con altas dosis de GD2-lactona-KLH tuvieron una alta respuesta de Ac con capacidad de inducir CDC en células tumorales GD2 positivas (Ragupathi et al., 2003a; Ragupathi et al., 2003b). 34 Revisión bibliográfica Pacientes de cáncer de pulmón de células pequeñas, inmunizados con la variante sintética y fucosilada de GM1-KLH en presencia de QS-21, mostraron respuestas de Ac IgM, pero un solo paciente demostró una respuesta tipo IgG, de bajo título (Krug et al., 2004). Otro resultado fallido se obtuvo con el candidato GD3-KLH, que no levantó respuesta de Ac en los pacientes tratados (Livingston, 1995a). Sin embargo, pacientes de melanoma inyectados con GD3-lactona-KLH en presencia de QS-21, dieron lugar a respuestas de IgM. Con el objetivo de levantar la respuesta de Ac anti-GD3, se realizó otro intento, usando el AcM anti-idiotipo BEC2, que mimetiza al gangliósido, como inmunógeno (Chapman et al., 2004). A pesar de que los primeros resultados en pacientes de cáncer de pulmón fueron prometedores, el ensayo clínico fase III no demostró ninguna ventaja de supervivencia para los pacientes vacunados (Giaccone et al., 2005). Tabla 1. Gangliósidos identificados como antígenos tumorales que han sido blancos de la ITAE. Gangliósido Tipo de Tumor GD2 Melanoma Fucosil-GM1 SCLC GD3 Melanoma Pulmón GM2 Melanoma Tratamiento Respuesta inmune Ac IgM GD2-lactona-KLH AcM anti-idiotipo 1A7 Fucosil-GM1Ac IgM KLH/QS-21 GD3-lactonaAc IgM KLH/QS-21 AcM anti-idiotipo BEC2 GM2-KLH/QS-21 Ac IgM IgG Referencia (Ragupathi et al., 2003a; Ragupathi et al., 2003b) (Dickler et al., 1999); (Krug et al., 2004) (Livingston, 1995a) (Chapman et al., 2004; Giaccone et al., 2005) e (Slovin et al., 2005; Eggermont et al., 2013) El preparado más prometedor usando la KLH fue con el gangliósido GM2. Las primeras preparaciones vacunales con el gangliósido GM2 utilizaron BCG o ciclofosfamida, y lograron respuestas de Ac IgM, predominantemente. A pesar de la ausencia de respuesta de Ac de afinidad madurada, o cambio de clase de los Ac generados, se observó un significativo aumento en el intervalo libre de enfermedad y en la sobrevida total en el 17% 35 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP de los pacientes tratados con GM2/BCG (Livingston et al., 1994a). Con el objetivo de aumentar la inmunogenicidad del GM2, el mismo se conjugó con la KLH. La administración de GM2-KLH resultó en el incremento de los títulos de Ac IgM e IgG en los pacientes (Kitamura et al., 1995). Otros adyuvantes se emplearon también para incrementar la inmunogenicidad del GM2, tal es el caso del Detox y de QS-21 (Helling et al., 1995). La vacuna GM2-KLH mostró resultados prometedores en los primeros ensayos clínicos realizados (Slovin et al., 2005), sin embargo, en el ensayo clínico fase III, a pesar de demostrar una correlación entre la respuesta de Ac y la sobrevida, no se obtuvo beneficio clínico con la vacunación cuando se comparó con los grupos placebo y control no tratado (Eggermont et al., 2013). 2.4 Mecanismos de acción de las vacunas de cáncer basadas en gangliósidos El papel de los Ac en el efecto antitumoral inducido por las vacunas basadas en gangliósidos ha sido considerado predominante en este tipo de terapia (Livingston et al., 1997a). Evidencias procedentes de los modelos animales y los estudios clínicos han mostrado que la frecuencia de inducción de Ac anti-gangliósidos correlaciona con la supervivencia (Zhang et al., 1998; Takahashi et al., 1999). Está descrito en la literatura que los niveles de Ac IgM específicos por los gangliósidos GD2, GM2, GD3 y NAcGM3 correlaciona con la supervivencia de pacientes de melanoma al ser tratados con una vacuna de células completas (Takahashi et al., 1999). Por otra parte, los trabajos de Livingston y cols., demostraron que pacientes de melanoma en estadío III, que desarrollaban Ac naturales o inducidos por vacunación contra el gangliósido GM2 mostraban un incremento en el intervalo libre de enfermedad y en la supervivencia (Livingston et al., 1994b). 36 Revisión bibliográfica Los Ac específicos por los gangliósidos son capaces de mediar la CDC de células positivas a la expresión del Ag. En este sentido, Hsueh y cols. describieron una correlación significativa entre el intervalo libre de enfermedad y la propiedad del suero de lisar células tumorales mediante CDC, en pacientes de melanoma estadio III vacunados con Canvaxin como terapia adyuvante (Hsueh et al., 1998). En otro estudio sobre el papel de los Ac anti-gangliósidos de isotipo IgM, Kawashima y cols. demostraron que en ratones transgénicos para Ac IgM contra el gangliósido GD2 se inhibía el crecimiento de la línea de linfoma T EL-4, la cual expresa este gangliósido. Además, en el bazo de estos ratones se encontró un incremento del número de células NK comparado con animales no transgénicos. La eliminación de esta población redujo o eliminó el efecto antitumoral, lo que sugiere que las células NK desempeñan un importante papel en la erradicación del tumor en este tipo de ratones (Kawashima et al., 2003). Se ha publicado la detección de células T específicas por los gangliósidos presentados en moléculas de presentación no clásica como el CD1. Estas células se han encontrado en donantes sanos y pacientes de esclerosis múltiple (Shamshiev et al., 1999). Chapman y cols. demostraron que ratones inmunizados con células de melanoma humano las cuales expresaban GD3 desarrollaron una respuesta de células NKT restringidas a CD1d y específicas por este gangliósido. En estos estudios se demostró además que el GD3 era presentado de forma cruzada por las APC murinas (Wu et al., 2003). Los resultados obtenidos en los modelos preclínicos sugieren que aunque no se ha demostrado la inducción de células T ni NKT específicas por los gangliósidos con la inmunización con vacunas compuestas por este tipo de moléculas, no se puede descartar la posibilidad de que la rama celular de la respuesta inmune esté involucrada en los mecanismos antitumorales inducidos por estas terapias. 37 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 2.5 El gangliósido GM3 como blanco de la IT de cáncer El gangliósido GM3, en sus variantes N-acetilada y N-glicolilada, se ha utilizado como blanco de la IT de cáncer (Fernandez et al., 2010; Fuentes et al., 2010; Hernandez et al., 2011). La efectividad de vacunas que contienen el gangliósido NAcGM3 en su composición ha sido estudiada en el modelo del melanoma murino B16 puesto que es el gangliósido mayoritario (Vedralova et al., 1995). Los esfuerzos para aumentar la inmunogenicidad de esta molécula no han sido del todo exitosos debido a su abundancia en los tejidos normales. Los resultados obtenidos por Livingston y cols., demostraron que la inmunización de ratones con el gangliósido NAcGM3 unido a un mutante de Salmonella minnesota no indujo respuesta de Ac contra el gangliósido (Livingston et al., 1989a). Por otra parte, Ravindranath y cols., inmunizaron ratones con vacunas celulares de melanoma B16 que contenían monofosforil lípido A, logrando respuesta de Ac solo de isotipo IgM contra el gangliósido en todos los ratones inmunizados y rechazo del tumor en menos del 40 por ciento de ellos (Ravindranath et al., 1994). Otra variante fue la vacuna Canvaxin, compuesta por tres líneas alogénicas de melanoma humano adyuvada con BCG, que se administró a pacientes de melanoma avanzado. Los resultados del ensayo clínico en un pequeño número de pacientes demostraron una mayor supervivencia en aquellos sujetos con altos títulos de Ac contra el gangliósido NAcGM3 (Chung et al., 2003). Sin embargo, estos hallazgos no se corroboraron en un ensayo clínico fase III, el cual no mostró ningun beneficio clínico para los pacientes vacunados respecto a los pacientes que recibieron placebo (Kelland, 2006). 38 Revisión bibliográfica 2.5.1 El preparado vacunal NAcGM3/VSSP Otra terapia contra el gangliósido NAcGM3 es el preparado vacunal NAcGM3/VSSP. Los estudios realizados con el NAcGM3/VSSP demostraron que la inmunización de ratones, monos y humanos con este preparado en adyuvante oleoso estimuló la producción de Ac de tipo IgM e IgG específicos tanto por el gangliósido NAcGM3 como por las proteínas de Nm (Estevez et al., 1999; Guthmann et al., 2004). Por otra parte, la inmunización de ratones C57BL/6 con NAcGM3/VSSP adyuvado con Montanide ISA 51, indujo la protección de estos animales ante el reto con la línea tumoral de melanoma B16 (Carr et al., 2001). Por otra parte, en un ensayo clínico fase I en pacientes de melanoma avanzado, la vacuna NAcGM3/VSSP emulsionada en Montanide ISA 51 indujo altos títulos de anticuerpos de isotipo IgM en el 44 % de los pacientes, sin evidencias de respuesta celular contra el gangliósido. Desde el punto de vista clínico en un paciente se indujo primero estabilización de la enfermedad y luego una reducción del tumor en lesiones de tejido blando acompañada por aparición de vitiligo (Guthmann et al., 2004). Sin embargo, el fracaso de la inmunoterapia activa en pacientes con estadíos avanzados de la enfermedad, motivó a buscar un inmunógeno efectivo contra el melanoma en el escenario de enfermedad residual mínima e incluso en el preventivo. Mazorra y cols (2008) demostraron en estudios en ratones C57 con el modelo de melanoma murino B16 que el preparado NAcGM3/VSSP, emulsionado en Montanide ISA 51, protege a los ratones de la recurrencia del melanoma B16, lo cual sugiere una mayor posibilidad de éxito terapéutico en el escenario de enfermedad residual mínima. Estos autores señalaron que los Ac de isotipo IgG específicos al gangliósido NAcGM3 participan en el mecanismo inmunológico antitumoral inducido por el inmunógeno en el escenario preventivo. En este mismo trabajo, se demostró que el preparado vacunal NAcGM3/VSSP, emulsionado en Montanide ISA 51, estimuló la expansión de células T CD8+ secretoras de IFNγ específicas al melanoma B16 en los 39 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP animales portadores de tumor, que están involucradas en la capacidad antitumoral del preparado (Mazorra et al., 2008). 2.5.2 Racotumumab Una de las estrategias vacunales que tiene como blanco a la variante N-glicolilada del gangliósido GM3 consiste en el uso del AcM anti-idiotípico Racotumumab (inicialmente conocido como 1E10), también referido como Ab2, que funciona como subrogado del gangliósido NGcGM3 (Hernandez et al., 2005) e induce una respuesta específica de Ac IgM e IgG anti-NGcGM3. Vaxira, que es el nombre que se le dió a la formulación de Racotumumab en hidróxido de aluminio (Alúmina) fue registrada en Cuba y Argentina en el año 2013 para el tratamiento de pacientes de cáncer de pulmón de células no pequeñas. Vaxira es segura y bien tolerada, con mínima toxicidad (Alfonso et al., 2014). La eficacia de Racotumumab en Alúmina se evaluó en varios ensayos clínicos fase I en pacientes de melanoma maligno avanzado, cáncer de pulmón de células pequeñas, en cáncer de mama estadíos III/IV y metastásico, y en cáncer de células no pequeñas estadíos IIIb y IV (Diaz et al., 2003; Alfonso et al., 2007; Neninger et al., 2007). En todos los ensayos clínicos fase I la mayoría de los pacientes tratados con Racotumumab en Alúmina desarrollaron Ac IgM e IgG anti-NGcGM3. Particularmente, en el ensayo con los pacientes de cáncer de células no pequeñas estadíos IIIb y IV, se encontraron diferencias estadísticamente significativas en la mediana de supervivencia de los pacientes que generaron respuesta de Ac IgM e IgG anti-NGcGM3 y los que no (Hernandez et al., 2008). En el ensayo clínico fase II, en pacientes de cáncer de pulmón de células no pequeñas, .la administración de Racotumumab en Alúmina demostró un beneficio en la supervivencia de los pacientes tratados respecto a los controles que recibieron placebo (Vazquez et al., 2012). 40 Revisión bibliográfica 2.5.3 El preparado vacunal NGcGM3/VSSP El segundo candidato dirigido a este mismo blanco, conjuga al gangliósido NGcGM3 en VSSP, obtenido a partir de las VME de la Nm (Estevez et al., 1999). Estévez y cols. (1999) describieron que la inmunización con preparaciones de los gangliósidos NAcGM3, NGcGM3 y GD3, en VSSP y adyuvante oleoso, indujo la respuesta de Ac anti-gangliósidos tanto tipo IgM como IgG (Estevez et al., 1999). En los ratones, la fracción IgG de Ac anti-gangliósidos no se restringió isotipo IgG3, típico de las respuestas a Ag T-independentes. De manera inesperada, esos autores encontraron Ac anti-gangliósidos de los isotipos IgG1, IgG2a e IgG2b. Resultados similares se alcanzaron cuando las mismas preparaciones se administraron en pollos y monos (Estevez et al., 1999). Con el preparado vacunal NGcGM3/VSSP adyuvado en Montanide ISA 51 se han realizado varios ensayos clínicos en Cuba, en pacientes de melanoma (Osorio et al., 2008; Osorio et al., 2012; Perez et al., 2013) y cáncer de mama (Carr et al., 2003; Mulens et al., 2010). Los eventos adversos producidos por la vacunación se clasificaron como leves. Todos los pacientes tratados con NGcGM3/VSSP desarrollaron títulos de Ac anti-NGcGM3, y el suero hiperinmune reconoció las líneas tumorales P3X63 y L1210, ambas NGcGM3 positivas (Carr et al., 2003; Mulens et al., 2010). Los ensayos clínicos fase III que se encuentran en curso, validarán el beneficio en supervivencia. 41 3. MATERÍALES Y METODOS 3.1 Animales Se utilizaron dos modelos animales: pollos Leghorn, de 10-12 semanas, mantenidos en el Centro Nacional de Salud Animal (CENSA, Cuba); y ratones hembras C57BL/6, de 8-12 semanas de edad, del Centro Nacional para la Producción de Animales de Laboratorio (CENPALAB, Cuba) que se mantuvieron en el bioterio del CIM con alimentación ad libitum. Las manipulaciones y los experimentos se realizaron de acuerdo con las directrices establecidas por este centro para el manejo de los animales de laboratorio. 3.2 Líneas celulares y medios de cultivo La línea celular murina del clon metastásico del carcinoma pulmonar de Lewis 3LL-D122 (Eisenbach et al., 1984), la línea celular de mieloma murino P3X63 (Kearney et al., 1979) y los hibridomas de rata, productores de inmunoglobulinas específicos por las moléculas murinas NK 1.1 (PK136), CD4 (YTS191) y CD8 (YTS169) se obtuvieron de la Colección Americana de Tipos Celulares (ATCC, del inglés: American Type Culture Collection). Para los cultivos celulares se utlilizó el medio Dulbecco Modificado (DMEM-F12, Hyclone), suplementado al 10% con suero fetal de ternera inactivado (SFT, HyClone), 2 mM de Lglutamina, 10 mM de HEPES, 20 µM de β-mercaptoetanol, 100 U/mL de penicilina y 100 U/mL de estreptomicina. 3.3 AcM utilizados para la eliminación de poblaciones linfocitarias Para los experimentos de eliminación in vivo de poblaciones linfocitarias se utilizaron preparaciones de AcM específicos por las moléculas NK1.1, CD8 y CD4, purificadas mediante precipitación con NH4SO4, a partir del sobrenadante de cultivo de los hibridomas PK136, YTS169 y YTS 191, respectivamente. 43 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 3.4 AcM utilizados en la citometría de flujo La determinación de moléculas en la superficie celular se realizó a través de ensayos de citometría de flujo (FACS, del inglés: fluorescence activated cell sorter). Todos los AcM conjugados a los fluoróforos se emplearon a una dilución final de 1:200 en solución salina tamponada con fosfatos (SSTF) con SFT al 1%. 3.4.1 Para la detección del gangliósido NGcGM3 en la membrana plasmática de las células tumorales Se utilizó el AcM 14F7 humanizado (Fernandez-Marrero et al., 2011), específico por el gangliósido NGcGM3. En el revelado de este Ac se empleó un suero de conejo anti-IgG humana (F0056, Dako) conjugado a isotiocianato de fluoresceína (FITC, del inglés: fluorescein iso-thiocyanate). Se empleó el AcM irrelevante hR3 como control de isotipo. 3.4.2 Para la determinación de moléculas de superficie en las DC Se utilizaron los siguientes AcM, generados en hámster o rata, adquiridos de la firma BD Pharmingen, conjugados a FITC o a ficoeritrina (PE, del inglés: phycoerythrin), específicos por las moléculas murinas CD11c (HL3) conjugado a PE; CD40 (3/23) conjugado a FITC; CD80 (16-10A1) conjugado a FITC; CD86 (GL1) conjugado a FITC; CD11c (HL3) conjugado a FITC; CD1d (1B1) conjugado a PE; B220 (RA3-6B2) conjugado a PE. 3.4.3 Para la detección de linfocitos infiltrantes en pulmón Se utilizaron los siguientes AcM, generados en rata o hámster, adquiridos de la firma eBioscience, conjugados a FITC o a PE, específicos por las moléculas murinas NK 1.1 (PK136) conjugado a PE; CD3 (145-2C11) conjugado a FITC; CD45 (30-F11) conjugado a ficoeritrina-cianina (PECy5, del inglés: phycoerythrin and cyanine), CD3 (145-2C11) conjugado a PECy5; CD4 (RM4-5) conjugado a FITC; CD8 (53-6.7) conjugado a PE; FoxP3 44 Materiales y Métodos (NRRF-30) conjugado a PE, CD25 (PC61.5) conjugado a PECy5; Gr1 (RB6-8C5) conjugado a PE, CD11b (Mac-1a) conjugado a PECy5. 3.5 Gangliósidos NGcGM3 y NAcGM3 Ambos gangliósidos los suministró el Centro de Química Biomolecular (CQB, Cuba). El gangliósido NGcGM3 de fuente natural se obtuvo a partir de eritrocitos de sangre de caballo (Estevez et al., 1999) y el sintético, mediante síntesis química por el método descrito por Duclos (Duclos, 2000) y Sherman (Sherman et al., 2002). El gangliósido NAcGM3 se obtuvo de eritrocitos de perro (Estevez et al., 1999). La pureza se monitoreó por cromatografía de alta resolución en placa delgada (HPTLC, del inglés: high performance thin layer chromatography). 3.6 Los preparados vacunales NGcGM3/VSSP y NAcGM3/VSSP Todos los preparados vacunales fueron proveídos por la Dirección de Desarrollo del CIM. Los preparados NGcGM3/VSSP y NAcGM3/VSSP se obtuvieron a partir de la incorporación hidrofóbica del gangliósido purificado de eritrocitos de caballo (NGcGM3) o de perro (NAcGM3) respectivamente, a las VME de la Nm, según el método descrito por Estévez y cols. (Estevez et al., 1999). Para los experimentos en pollos se construyó, por el mismo método, un preparado NGcGM3/VSSP con el gangliósido NGcGM3 sintético (SAg/VSSP). En el resto de los experimentos, tanto in vivo como in vitro, se utilizó el preparado vacunal NGcGM3/VSSP con Ag natural. La relación de masa entre gangliósidos y proteínas de cada preparado osciló entre 0,6-0,8. Los mismos se dosificaron a partir del contenido de gangliósidos. La preparación vacunal NGcGM3/VSSP emulsionada con Montanide se preparó con el adyuvante oleoso Montanide ISA 51 (Seppic, Francia), al unir iguales volúmenes de vacuna 45 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP y adyuvante. Para lograr la emulsión se agitó la mezcla en un vórtex (Hettich, Alemania) durante 5 min. 3.7 Desarrollo de tumores primarios del carcinoma de pulmón de Lewis 3LL-D122 Los tumores primarios utilizados para la determinación del contenido del gangliósido NGcGM3 se obtuvieron a partir de la inoculación subcutánea (SC) de línea tumoral 3LLD122 (5x105 células) en el flanco de ratones C57BL/6. Veinticuatro días después de crecido el tumor los animales se sometieron a sacrificio humanitario y se extrajeron los tumores primarios crecidos en el flanco de los mismos. 3.8 Modelo de metástasis espontánea del carcinoma de pulmón de Lewis 3LL-D122 La actividad antitumoral del preparado vacunal NGcGM3/VSSP se evaluó en el modelo murino de metástasis espontánea con la línea tumoral 3LL-D122 descrito por Eisenbach y cols. (Eisenbach et al., 1983). El día cero del experimento se realizó la inoculación de las células de la línea tumoral 3LL-D122 (2x105) en la almohadilla plantal de ratones C57BL/6. El momento de aparición de los tumores se monitoreó por palpación. Se siguió el crecimiento de los tumores primarios mediante la medición del diámetro del tumor con pie de rey. Entre los 26-30 días el diámetro del tumor crecido en la pata de los animales alcanzó 0,8-0,9 mm, momento en el cual estos tumores fueron removidos por cirugía, luego de anestesiar a los animales. Veintiún días después los animales se sometieron a sacrificio humanitario y se extrajeron los pulmones. Para la determinación del efecto antimetastásico del preparado vacunal los pulmones se pesaron en una balanza analítica. En este modelo el peso del pulmón correlaciona directamente con la talla y el número de las metástasis (Gorelik et al., 1980; Eisenbach et al., 1983). 46 Materiales y Métodos Las metástasis pulmonares para la determinación del contenido del gangliósido NGcGM3 se obtuvieron por separación de la masa tumoral metastásica con pinzas y tijeras bajo microscopio estereoscopio, con el cuidado de no incluir tejido pulmonar. 3.9 Extracción de gangliósidos de las muestras tumorales Las muestras tumorales del carcinoma pulmonar 3LL-D122: línea celular, tumor primario y metástasis, se procesaron para la extracción de gangliósidos según se describe a continuación. Primeramente se homogenizaron en agua y se extrajeron los lípidos totales en cloroformo:metanol:agua en proporción 4:8:3 (v:v:v) durante una hora. El homogenado obtenido de cada muestra tumoral se centrifugó a 3 000 rpm, 15 minutos y la fracción lipídica se recuperó en el sobrenadante (SN). La mezcla de solventes se evaporó por rotoevaporación (Büchi, Suiza), los lípidos secos se disolvieron en cloroformo:metanol en proporción 1:1 (v:v) y se mantuvieron a 4°C durante toda la noche. A continuación se centrifugaron a 3 000 rpm, 15 minutos y se recuperó en el SN la mezcla de lípidos totales. Los solventes contenidos en los SN se rotoevaporaron, los lípidos secos se disolvieron en cloroformo:metanol:agua en proporción 30:60:8 (v:v:v) y se aplicaron en una columna de DEAE-Sefarosa A-25, con vistas a obtener los gangliósidos monosialilados. La fracción de los monosialo-gangliósidos se despegó de la columna con acetato de sodio 0,02 M en metanol. Las mezclas de gangliósidos obtenidas se utilizaron para la identificación del gangliósido NGcGM3. 3.10 Identificación química del gangliósido NGcGM3 Para la identificación química del gangliósido NGcGM3, las mezclas de gangliósidos se sometieron a HPTLC en placas de sílica gel (Merck) (Estevez et al., 1999). Las placas se corrieron en cloroformo:metanol:2,5 M NH3 en proporción 65:35:8 (v:v:v). Los patrones de los gangliósidos NAcGM3 y NGcGM3 se corrieron en paralelo con las muestras de 47 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP gangliósidos procedentes de las muestras tumorales. El revelado químico de los gangliósidos se realizó con el reactivo de resorcinol-HCl (Svennerholm, 1963). Se calculó el valor del factor de retención (Rf) relativo a los controles de gangliósidos empleados para determinar la banda correspondiente a los gangliósidos NGcGM3 y NAcGM3. Las imágenes de la HPTLC se capturaron con un escáner (Powerlook III, Amersham Biosciences). El porcentaje de los gangliósidos NGcGM3 y NAcGM3 del total de los monosialio-gangliósidos obtenidos se calculó por análisis densitométrico con el programa TotalLab120 (Nonlinear Dynamics). 3.11 Identificación inmuno-química del gangliósido NGcGM3 en HPTLC Las mezclas de gangliósidos se sometieron a una inmuno-tinción de las placas HPTLC, con el AcM 14F7 murino (Carr et al., 2000), de acuerdo al protocolo descrito por Saito y cols. (Saito et al., 1985) con pequeñas modificaciones. Brevemente, las mezclas de gangliósidos se separaron por HPTLC, las placas se sumergieron en una solución al 0,1% de poliisobutilmetacrilato (Aldrich Chemical) en hexano durante 75 segundos y se pusieron a secar al aire. Posteriormente, las placas bloqueadas con SSTF conteniendo 1% de BSA (Sigma), se incubaron a 4°C (toda la noche) con 10 μg/ml del AcM 14F7 murino. Después de los lavados, se adicionó el Ac anti-IgG de ratón conjugado a fosfatasa alcalina (Jackson). Las manchas se visualizaron con una solución de 5-bromo-4-cloro-3-indolil fosfato a 1 mg/ml en tampón glicina 0,1 M (pH 10,4). La reacción se detuvo lavando con agua. 3.12 Identificación inmuno-histoquímica del gangliósido NGcGM3 Para la detección del gangliósido mediante inmunohistoquímica (IHQ) con el AcM 14F7 murino se utilizaron cortes de tumor primario y de metástasis pulmonares de la línea 3LLD122. Para la detección se empleó el ensayo Vectastain ABC (Vector Laboratories) y se siguieron las instrucciones del fabricante. Brevemente, las muestras de los tumores 48 Materiales y Métodos subcutáneos y de las metástasis pulmonares se fijaron en formalina y se procesaron usando la técnica de rutina de parafina (Alonso et al., 1999). Secciones de los tumores (5 μm) se incubaron se durante 1h con el AcM 14F7 murino (20 μg/ml en SSTF). A continuación se incubaron primero con un suero de caballo anti-IgG de ratón por 30 min y después con un complejo avidina-biotina conjugado a peroxidasa durante 30 min. Luego se añadió el sustrato de la peroxidasa (diaminobencidina) y las secciones de tumor se contrastaron con hematoxilina. Se realizaron los controles pertinentes de especificidad, que incluyeron el bloqueo de la peroxidasa endógena, la omisión del primer Ac y la utilización de un Ac no relacionado (AcM 1E10). 3.13 Detección del gangliósido NGcGM3 por espectrometría de masa La detección del gangliósido NGcGM3 se realizó a través de un MALDI/MS en un espectrómetro de masa (Shimadzu Biotech Axima). El ácido gentísico (ácido 2,5dihidroxibenzoico) 10 mM (Sigma) se usó como matriz. Las muestras (5 µg/ 1 µl) se depositaron en el soporte, cubiertas con 1 µl de la matriz en solución acuosa y se secaron. Los iones analitos se desorbieron de la matriz con pulsos de un láser de nitrógeno de 337 nm. Los espectros se obtuvieron en el modo de ión reflectivo positivo con un promedio de 128 pulsos, la ventana de iones se fijó en 1 000-1 500 Da. Se calculó el promedio de la masa obtenida en dos determinaciones. 3.14 Procedimientos de inmunización en los experimentos con animales 3.14.1 Experimentos de inmunogenicidad en pollos Se utilizaron dos variantes del preparado vacunal NGcGM3/VSSP construidas con el gangliósido NGcGM3 de fuente natural (NAg/VSSP) y con el gangliósido sintético (SAg/VSSP). Ambas preparaciones vacunales se emuslificaron con Montanide ISA 51 (Seppic). Grupos de 10 pollos recibieron 200 µg de cada preparado vacunal con Montanide, 49 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP por vía intramuscular (IM), los días cero y 14. Los sueros inmunes se colectaron el día 21 para el análisis de los Ac anti-NGcGM3. La extracción de sangre se realizó a partir de las venas de las alas. 3.14.2 Experimentos de inmunogenicidad en ratones Los ratones recibieron 200 µg del preparado vacunal NGcGM3/VSSP, con Ag natural, sin emulsión por vía subcutánea (SC). Los ratones sanos (n=10) recibieron las inmunizaciones los días cero y 14, y las extracciones de sangre se realizaron el día cero (suero preinmune) y el día 21. Los ratones portadores del tumor 3LL-D122 (n=4) recibieron las inmunizaciones los días siete y 21. Las extracciones de sangre se realizaron el día cero (suero preinmune) y los días 24, 28 y 35 después de implantadas las células tumorales. Las extracciones se realizaron por punción submandibular (Golde et al., 2005). 3.14.3 Experimentos de evaluación del efecto del preparado vacunal en el modelo murino de metástasis espontánea a pulmón del carcinoma de Lewis 3LL-D122 Para evaluar el efecto del preparado vacunal NGcGM3/VSSP en el modelo murino de metástasis espontánea del carcinoma pulmonar de Lewis 3LL-D122, treinta ratones C57BL/6 se inocularon con células tumorales 3LL-D122 (2x105/ratón) en la almohadilla plantal y se aleatorizaron en grupos de 15 animales. Un grupo recibió dos dosis de 200 µg del preparado vacunal NGcGM3/VSSP no emulsivo, por vía SC, siete y 21 días después de la inoculación del tumor, mientras el otro recibió SSTF (grupo control). En los experimentos de evaluación de las vías IM e intraparitoneal (IP) se empleó el mismo esquema de dosis y frecuencia. Para la comparación del efecto del preparado vacunal NGcGM3/VSSP no emulsionado, y emulsionado con Montanide, grupos aleatorizados de 7-10 ratones C57BL/6 inoculados con las células tumorales 3LL-D122 (2x105/ratón) en la almohadilla plantal, se trataron con dos 50 Materiales y Métodos dosis de 200 µg del preparado vacunal NGcGM3/VSSP sin adyuvante oleoso (por vía SC), o adyuvado en Montanide ISA 51 (por vía IM), siete y 21 días después de la inoculación del tumor. Los ratones del grupo control recibieron SSTF. Para el estudio de la relevancia del gangliósido NGcGM3 en la actividad de la vacuna NGcGM3/VSSP, grupos de ratones portadores del tumor de Lewis 3LL-D122 en la almohadilla plantal (aleatorizados a razón de 10 animales por grupo) recibieron por vía SC, dos dosis de 200 µg del preparado vacunal NGcGM3/VSSP, 200 µg del preparado NAcGM3/VSSP y la dosis equivalente de VME, los días siete y 21 del experimento. Los ratones del grupo control recibieron SSTF. 3.14.4 Experimentos de medición in vivo de maduración de DC Ratones sanos (n=3) se vacunaron con 200 μg del preparado vacunal NGcGM3/VSSP por vía SC (en el lomo) y luego de tres días se aislaron las células de los ganglios linfáticos aferentes (axilares) mediante maceración, para evaluar mediante FACS, la expresión de marcadores de maduración en las DC. Los ratones del grupo control recibieron SSTF. 3.14.5 Experimentos de medición de citocinas en suero Ratones sanos se vacunaron (n=3) con 200 μg del preparado vacunal NGcGM3/VSSP por vía IP y luego de 2, 4, 6 y 24 h se colectó el suero por punción submandibular (Golde et al., 2005). Grupos controles, de tres animales cada uno, recibieron LPS (15 µg), poli I:C (200 µg) y SSTF, todos por vía IP. 3.15 Determinación de anticuerpos séricos Los Ac séricos contra los gangliósidos NGcGM3 y NAcGM3 se determinaron mediante un ensayo inmunoenzimático colorimétrico (ELISA) indirecto, que se describe brevemente. Una parte de la placa de 96 pozos Polysorp (Nunc,) se recubrió con el gangliósido NGcGM3 o 51 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP NAcGM3 (4 µg/ml) disuelto en metanol, y la otra con el solvente orgánico sin gangliósido. Los pozos se secaron a 37ºC y se bloquearon con el tampón de bloqueo que contenía 1% de BSA en SSTF. Posterior a la incubación con las muestras de los sueros de los animales tratados, se realizó la incubación con el conjugado adecuado unido a la fosfatasa alcalina. Después de lavar las placas, se adicionaron 100 μl por pozo de la solución de sustrato que contenía 1 mg/ml de p-nitrofenilfosfato en tampón dietanolamina pH 9,8. La absorbancia del producto de la reacción se midió a los 30 min en un lector de placas (Organon Teknika) a 405 nm. Los sueros se consideraron positivos cuando la absorbancia fue mayor de 0,1. Estos valores se obtuvieron luego de sustraer al valor medio de las absorbancias experimentales obtenida para cada dilución en los pozos recubiertos con el Ag, los valores obtenidos al enfrentar los sueros a los pozos tratados de igual forma pero donde no se añadió el gangliósido. 3.15.1 Preparación de las muestras para la titulación de los Ac anti-gangliósido en los pollos Los sueros se diluyeron de forma seriada desde 1:80 hasta 1:5 120, se adicionaron a los pozos (50 μl por pozo) por duplicado y las placas se incubaron toda la noche a 4ºC. Después de lavar los pozos con 0,05% Tween 20-SSTF, se añadieron 50 μl de suero antiinmunoglobulinas IgY de pollo conjugado a fosfatasa alcalina (Sigma) diluido 1: 1 000. 3.15.2 Preparación de las muestras para la evaluación de los Ac anti-gangliósido en los ratones Los sueros de los ratones sanos vacunados se evaluaron a una dilución fija de 1:80. En el caso de la estimación de los Ac anti-gangliósido en los ratones portadores del tumor 3LLD122, los sueros se evaluaron en una dilución fija (1:100) y se titularon a partir de diluciones seriadas desde 1:100 hasta 1:1 600. En todos los casos se añadieron 50 μl por 52 Materiales y Métodos pozo, por duplicado y se incubaron toda la noche a 4ºC. A continuación los pozos se lavaron con 0,05% Tween 20-SSTF, y se añadieron 50 μl de suero anti-IgM de ratón conjugado a fosfatasa alcalina (Sigma) diluido 1: 5 000; ó 50 μl de suero anti-IgG de ratón conjugado a fosfatasa alcalina (Sigma) diluido 1: 5 000. 3.16 Reconocimiento de la línea tumoral P3X63 por los sueros de los animales vacunados Las células P3X63 se lavaron con SSTF, a continuación con solución de lavado para FACS (SLC: 0,1% BSA y 0,1% NaN3 en SSTF) y después se ajustaron a 1x105 células por muestra. Las células se incubaron por 30 min a 4°C con los sueros de los pollos, todos diluidos 1/10 en SLC. Posteriormente, se lavaron las muestras con SLC. A continuación se adicionó el Ac anti-IgY pollo conjugado a biotina (diluido 1/200). Luego de 15 minutos de incubación a 4°C y dos lavados con SLC, se adicionó el complejo estreptavidina-FITC, diluido 1/200, durante otros 15 min y a 4°C. Finalmente, las células se lavaron con SSTF, y se adicionó 200 µl de SSFT por muestra para el análisis en el citómetro de flujo. 3.17 Obtención de las células dendríticas murinas a partir de la médula ósea Las DC murinas se obtuvieron a partir de precursores aislados de la médula ósea (bm-DC, del inglés: bone marrow DC). Con este objetivo se extrajeron, en condiciones estériles, los huesos fémur y tibia de los ratones recién sacrificados por dislocación cervical. Los extremos de los huesos se cortaron y la médula se aisló mediante el paso de un flujo de medio de cultivo por el interior del hueso con una aguja 23Gx1. Las células de la médula ósea se centrifugaron y se resuspendieron en cloruro de amonio al 0,83% (a temperatura ambiente por 2 minutos) para lisar los eritrocitos. 3.17.1 Obtención de las DC convencionales Para obtener las DC convencionales (cDC) las células extraídas se sembraron en placas de cultivo de seis pozos, a una densidad de 6x105 células/ml en 3 ml de medio RPMI 1640 53 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP suplementado con 20 ng/mL del factor estimulador de colonias de granulocitos y macrófagos (GM-CSF, del inglés: granulocyte-macrophage colony stimulating factor) (R&D Systems). Entre las 72 y 90 h de cultivo, se añadieron otros 3 ml de medio de cultivo suplementado con GM-CSF (Inaba et al., 1992). A los siete días de cultivo las células se colectaron y se utilizaron frescas para los experimentos celulares. 3.17.2 Obtención de las DC plasmacitoides Para obtener las DC plasmacitoides (pDC), las células extraídas se sembraron en placas de cultivo de 24 pozos, a una densidad de 2x106 células/ml en 1 ml de medio IMDM (Gibco) suplementado al 15% con ligando de tirosina cinasa Fms-3 (FLt3-L, del inglés: Fms-related tyrosine kinase 3 ligand), obtenido en el SN del cultivo de las células Sp2/0 secretoras de FLT3-L (Brasel et al., 2000). Cada 24 h a las placas se le cambiaron el 50% del medio metabolizado por medio fresco, siempre suplementado con 15% de FLt3-L. A los siete días del cultivo, las bm-pDC se purificaron mediante selección positiva con perlas magnéticas conjugadas a AcM anti-B220+ (Miltenyi Biotec). Las bm-pDC se utilizaron frescas para los experimentos celulares. Para determinar la pureza de cada población se estudió la expresión de los marcadores por citometría de flujo. La pureza de las células estuvo por encima del 80%. 3.18 Maduración de células dendríticas con NGcGM3/VSSP Las bm-cDC inmaduras se cultivaron durante toda la noche (18h) en medio (RPMI 1640, Gibco), y en presencia de LPS (1 µg/ml, Sigma) y NGcGM3/VSSP (1 µg/ml). Las células obtenidas se utilizaron para evaluar mediante FACS, la expresión de marcadores de maduración. Los SN del cultivo se colectaron para la determinación de IL-6 e IL12. 54 Materiales y Métodos Las bm-pDC inmaduras se cultivaron cultivadas toda la noche (18 h) en medio (IMDM, Gibco), y en presencia de LPS (0,5 µg/ml, Sigma), CpG-A (5 µg/ml, InVivoGen) y NGcGM3/VSSP (0,1 µg/ml). Las células obtenidas se utilizaron para evaluar mediante FACS, la expresión de marcadores de maduración. Los SN del cultivo se colectaron 18 h después para la determinación de IFNα. 3.19 Determinación de la expresión de moléculas de superficie por FACS Las DC y de linfocitos se estudiaron a través del marcaje de moléculas en la membrana plasmática. Con este objetivo, las células se incubaron durante 15 min en SSTF, azida sódica 0.01% y SFT al 1%, para minimizar las uniones inespecíficas. A continuación, 2x105 células por cada muestra se incubaron durante 15 min a 4 ºC, en la oscuridad, con los diferentes AcM conjugados a fluoróforos diluidos en una solución de SSTF, azida sódica 0,01% y SFT al 1%. Una vez marcadas las células se lavaron con la misma solución centrifugando 2 min a 2 000 rpm y se adquirieron en el citómetro de flujo. 3.20 Determinación de la frecuencia de células inmunes en pulmones El infiltrado inmune pulmonar se determinó por FACS. La frecuencia de células NK1.1+ se midió en grupos de 3 ratones C57BL/6, en el modelo de metástasis espontánea con la línea 3LL-D122, a los 3 días después de la segunda dosis de vacuna. La frecuencia de células efectoras de la inmunidad adaptativa y de células reguladoras se evaluó en grupos de 4-6 ratones C57BL/6, en el mismo modelo, a los 21 días de operados los tumores primarios. En todos los casos se administraron dos dosis de 200 µg de NGcGM3/VSSP por vía SC, los días siete y 21 después del reto tumoral. Los leucocitos del pulmón se obtuvieron por digestión del órgano en DMEM-F12 (Hyclone) conteniendo colagenasa D (1 mg/ml; Sigma) y DNasa I (1 mg/ml; Sigma), durante 30 min a 37°C. 55 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 3.21 Medición de citocinas producidas por las DC Se analizó el contenido de IFNα, IL-6 e IL-12 mediante ensayos ELISA comerciales, procedentes de PBL Biomedical Laboratories (IFNα) y Thermo Fischer Scientific (IL-12 y IL6). Se siguieron las instrucciones del fabricante en cada caso. Para la determinación de IFNα las muestras se evaluaron sin diluir. Para la evaluación de IL-6 los SN se diluyeron 1/5 y los sueros 1/10. Para la estimación de IL-12 los SN se diluyeron 1/20 y los sueros 1/10. 3.22 Experimentos de eliminación de poblaciones celulares in vivo Para evaluar la relevancia de las poblaciones celulares en la respuesta inmune, grupos de ratones C57BL/6, en el modelo de metástasis espontánea con la línea 3LL-D122, vacunados con dos dosis de 200 μg del preparado vacunal NGcGM3/VSSP y controles (SSTF), recibieron además por vía IP 1 mg de AcM específicos por las moléculas NK1.1, CD8 y CD4, un día después de la administración del preparado vacunal o de la SSTF. Esta dosis de los AcM demostró, en experimentos previos, eliminar más del 98% de la población celular. 3.23 Análisis estadístico Para calcular los valores de los coeficientes r de Pearson y de probabilidad de la correlación, así como para los demás análisis estadísticos realizados se utilizó el editor de gráficos GraphPad Prism versión 5.03. En todas las variables se evaluó la normalidad mediante la prueba de Kolmogorov-Smirnov y la homogeneidad de varianza mediante la prueba de Levene. Para el análisis de los datos de la población celulares infiltrantes de pulmón se empleó la prueba no paramétrica de comparaciones pareadas U de Mann-Whitney, con un nivel de significación de 0,01. 56 Materiales y Métodos Para la evaluación de las diferencias de la variable peso de pulmón en los ensayos de actividad anti-metastásica del preparado vacunal NGcGM3/VSSP se utilizó la prueba t de Student para muestras no pareadas cuando se compararon sólo dos grupos; mientras que cuando el experimento tenía tres o más grupos se realizó un análisis de varianza de clasificación simple (ANOVA, del inglés: Analysis of Variance) y se aplicó la prueba de Tuckey. Las comparaciones del crecimiento de los diámetros tumorales en el modelo de metástasis espontánea se realizaron mediante un ANOVA de dos vías. Las diferencias se consideraron significativas cuando p<0,05. 57 4. RESULTADOS 4.1 La expresión del gangliósido NGcGM3 en el modelo de carcinoma pulmonar de Lewis 3LL-D122 se incrementa en las metástasis respecto a los tumores primarios La relevancia del modelo de metástasis espontánea a pulmón con la línea 3LL-D122 y el hecho de que no se ha descrito anteriormente la expresión del gangliósido NGcGM3 en este tipo de tumor, motivó determinar la presencia del mismo en las diferentes etapas por las que transita el modelo (línea tumoral 3LL-D122, tumor primario y metástasis). Las muestras tumorales se procesaron para la extracción química y la purificación de monosialo-gangliósidos de forma individual, según se describe en Materiales y Métodos. La presencia del gangliósido NGcGM3 en las diferentes fracciones de los monosialogangliósidos (Figura 1), derivadas de las tres etapas del modelo tumoral, se hizo evidente a partir de los valores del Rf relativo a los controles de gangliósidos empleados en la técnica HPTLC. El componente principal de la fracción de gangliósidos derivados de la línea tumoral 3LL-D122 en cultivo fue el gangliósido NAcGM3, representando el 87.4% del total de los ácidos siálicos unidos a lípidos, mientras que su variante N-glicolilada constituyó sólo el 12.6% (Figura 1A). Por otra parte, en los tumores primarios, la diferencia en la expresión de ambas especies fue menor. El gangliósido NAcGM3 constituyó el 56.1% del total de los monosialo-gangliósidos y el gangliósido NGcGM3 el 32.2% (Figura 1B). En las lesiones metastásicas (Figura 1C) esta situación se invirtió, pues el gangliósido NGcGM3 constituyó el 73.4% de los monosialo-gangliósidos, mientras que la variante N-acetilada representó sólo el 11.2%. La presencia del gangliósido NGcGM3 en las diferentes fracciones se corroboró además mediante inmuno-tinción de las placas de HPTLC con el AcM 14F7 y mediante espectrometría de masa. 59 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP Los experimentos de inmuno-tinción con el AcM 14F7 confirmaron la presencia de esta sialil-lactosil-ceramida en las fracciones de los monosialo-gangliósidos derivadas de las células 3LL-D122, ya fueran cultivadas in vitro, o provenientes de los tumores subcutáneos o de las lesiones metastásicas de los pulmones (Figura 1D). Adicionalmente en las Figuras 1B y 1C puede observarse que gangliósido NAcGM3 se desdobló en dos bandas, hallazgo que indica heterogeneidad en la estructura de la ceramida. Esto no se observó para el gangliósido NGcGM3. Figura 1. Análisis químico e inmunoquímico del contenido gangliósidos extraídos de las diferentes etapas del modelo tumoral murino de metástasis espontánea con 3LL-D122. A-C: HPTLC de gangliósidos provenientes de (a) la línea tumoral 3LL-D122, (b) de los tumores primarios, y (c): de las metástasis pulmonares. D: Identificación inmuno-química del gangliósido NGcGM3 con el AcM 14F7. Leyenda: 1) patrón del gangliósido NGcGM3 (fuente: eritrocitos de caballo), 2) patrón del gangliósido NAcGM3 (fuente: eritrocitos de perro). De manera similar el análisis de los espectros de masa MALDI evidenciaron el proceso típico de decarboxilación (m/z 1274) visto frecuentemente como parte del proceso de ionización de estas moléculas, previo a sufrir la completa pérdida de los motivos de AS, en el espectro correspondiente de las mezclas de tumores subcutáneos (Figura 2A) y las metástasis (Figura 2B). Una vez que se estableció inequívocamente la presencia de la entidad química en los extractos lipídicos de las diferentes muestras celulares, se evaluó si el gangliósido NGcGM3 podría ser detectado en el material biológico intacto. Para esto se analizó la línea tumoral 3LL-D122 por citometría de flujo usando el AcM 14F7. 60 Resultados Como se puede apreciar en la Figura 3, si bien se obtuvo una tinción marcada de las células de mieloma P3X63 (NGcGM3-positivas, Figura 3A), fue nulo el reconocimiento de la línea de carcinoma 3LL-D122 (Figura 3B) por el AcM específico, lo cual confirmó los resultados químicos previos. Figura 2. Espectros de masa por MALDI-TOF de los gangliósidos extraídos del tumor primario (A) y de las lesiones metastásicas (B). La comparación de los patrones de expresión del gangliósido NGcGM3 entre los tumores primarios y las lesiones metastásicas se realizó mediante inmuno-histoquímica con el AcM 14F7. El resultado fue similar al obtenido con el HPTLC, es decir, los tumores primarios mostraron una tinción positiva y menos intensa para el gangliósido NGcGM3 (Figura 3C), que las correspondientes lesiones metastásicas (Figura 3D). En este último caso se 61 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP observó un patrón heterogéneo de inmunoreactividad con áreas de células del carcinoma de pulmón mostrando una tinción citoplasmática difusa. Figura 3. Detección del gangliósido NGcGM3 en las diferentes etapas del modelo de metástasis espontánea con el carcinoma pulmonar de Lewis 3LL-D122. A-B: Identificación del gangliósido NGcGM3, mediante citometría de flujo con el AcM 14F7, en la línea tumoral P3X63 (A) y en la línea tumoral 3LL-D122 (B). Leyenda: histograma relleno y gris: células sin marcar, histograma gris claro sin relleno: marcaje de un AcM control de isotipo, histograma negro sin relleno: marcaje con AcM 14F7. C-F: Identificación del gangliósido NGcGM3 en cortes del tumor primario (C y E) y en cortes de metástasis pulmonares espontáneas (D y F) del carcinoma pulmonar de Lewis 3LL-D122, mediante inmunohistoquímica con el AcM 14F7 (C-D) o el AcM irrelevante 1E10 (E-F). En conjunto, estos resultados apoyan la idea de la expresión del gangliósido NGcGM3 como marcador de malignidad en este modelo experimental. 4.2 El preparado vacunal NGcGM3/VSSP es inmunogénico en modelos animales Para realizar la evaluación de la inmunogenicidad del preparado vacunal NGcGM3/VSSP se utilizaron dos modelos animales. En un primer grupo de experimentos se utilizaron pollos, pues son el modelo experimental más apropiado ya que, al igual que el hombre 62 Resultados (Ozawa et al., 1992a), carecen de Ag N-glicolilados en los tejidos normales (Kawai et al., 1991). Para los experimentos de inmunogenicidad en pollos se utilizaron las variantes del preparado vacunal NGcGM3/VSSP construidas con el gangliósido NGcGM3 aislado de eritrocitos de caballo (NAg) y con el gangliósido NGcGM3 obtenido mediante síntesis química (SAg). Previo a la evaluación de la inmunogenicidad de las dos variantes vacunales (NAg/VSSP y SAg/VSSP) se corroboró la similitud antigénica de los dos gangliósidos Nglicolilados de diferentes orígenes. En la Figura 4A se puede apreciar que hubo diferencias en los Rf de ambos gangliósidos, sin embargo, tanto el gangliósido NAg como el SAg reaccionaron con al AcM 14F7 en un experimento de inmuno-tinción sobre placas de HPTLC (Figura 4B), confirmando la antigenicidad similar de ambos gangliósidos NGcGM3. Figura 4. Pureza e identidad del gangliósido NGcGM3 utilizado en el preparado vacunal NGcGM3/VSSP. A: HPTLC teñidas con orcinol, B: Inmunotinción de los gangliósidos corridos en HPTLC con el AcM 14F7. Leyenda: 1) patrón del gangliósido NGcGM3 (fuente: eritrocitos de caballo, Rf: 0,5), 2) NAg (Rf: 0,5) y 3) SAg (Rf: 0,4). Las dos composiciones del preparado vacunal NGcGM3/VSSP emulsionadas en Montanide ISA51 (Seppic) y administradas de manera independiente a grupos de 10 pollos por vía IM, generaron importantes niveles de Ac IgY contra el gangliósido NGcGM3 en el suero de la totalidad de los animales (Figura 5A). Los valores de absorbancia a 405 nm de los sueros pre-inmunes fueron inferiores a 0,1. La media de los títulos de Ac anti-NGcGM3 detectados en los pollos inmunizados fue 1 280, independientemente de cuál Ag se usó para la 63 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP obtención del preparado administrado (p > 0,05, prueba t de Student para muestras no pareadas), lo que indica que ambas preparaciones vacunales estimularon de igual manera al sistema inmunitario. Se observó además, mediante experimentos de FACS, que ambos anti-sueros reconocieron de forma silimar al gangliósido NGcGM3 expresado en las membranas de la línea tumoral P3X63 (Figura 5B-C). Figura 5. Respuesta de Ac IgY anti-NGcGM3 en pollos tratados con los preparados NAg/VSSP o SAg/VSSP, emulsionados en Montanide. A: Títulos de Ac IgY anti-NGcGM3 de 10 animales por grupo inmunizados por vía IM (columna: media de los valores, barra de error: desviación estándar). Se aplicó la prueba t de Student para muestras no pareadas, con un nivel de significación de 0,05 (p>0,05). B-C: Reconocimiento de la línea tumoral P3X63 por los sueros inmunes obtenidos por la vacunación con NAg/VSSP/ Montanide (B), o con SAg/VSSP/ Montanide (C). Leyenda: histograma negro relleno: células P3X63 sin marcar, histograma gris claro: suero preimmune, histograma gris oscuro: Ac anti-IgYbiotina, histograma negro: suero inmune. IMF: intensidad media de fluorescencia. En un segundo grupo de experimentos se evaluó la inmunogenicidad del preparado vacunal NGcGM3/VSSP (con Ag natural) sin adyuvante oleoso, en ratones C57BL/6, que es la cepa de origen de la línea tumoral 3LL-D122. En los ratones el gangliósido NGcGM3 constituye un Ag propio, por lo que evaluar la inmunogenicidad en este modelo es crítico para demostrar que el preparado tiene la capacidad de movilizar la rama humoral de la respuesta inmune. 64 Resultados La evaluación de los niveles de Ac anti-NGcGM3 generados a partir de la administración SC no emulsiva del preparado vacunal NGcGM3/VSSP en grupos de 10 ratones sanos se realizó siete días después de la segunda dosis. Como se puede observar en las Figura 6, el 80% de los animales vacunados seroconvirtieron y desarrollaron respuestas de Ac antiNGcGM3 de tipo IgM e IgG, medidas como la absorbacia a 405 nm a una dilución fija del suero (1/80), en el ensayo tipo ELISA descrito. Los valores de absorbancia a 405 nm de los sueros pre-inmunes fueron inferiores a 0,1. Figura 6. Inmunogenicidad del preparado vacunal NGcGM3/VSSP en ratones sanos. Diez animales fueron inmunizados con 200 µg del preparado vacunal NGcGM3/VSSP (NAg/VSSP) por vía SC los días cero y 14. La evaluación de los niveles de Ac antiNGcGM3 se realizó mediante ELISA con los sueros extraídos el día 21. Las figuras muestran los valores de absorbancia a 405 nm de la dilución 1/80 del suero de los animales. Por otra parte, la evaluación de los niveles de Ac anti-NGcGM3 y anti-NAcGM3 en grupos de cuatro ratones portadores del tumor 3LL-D122 evidenció que la presencia del tumor no fue suficiente para generar una respuesta inmune humoral anti-gangliósidos (Figura 7) que, como se mostró anteriormente en el acápite 4.1, se encuentran expresados en los tumores del carcinoma de Lewis 3LL-D122. Los ratones no vacunados, ya fueran sanos o portadores del tumor, no presentaron Ac antiNGcGM3 ni anti-NAcGM3. Después de la administración de dos dosis del preparado vacunal NGcGM3/VSSP, los ratones portadores del tumor de Lewis 3LL-D122 generaron 65 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP Ac anti-NGcGM3 de tipo IgM e IgG (Figura 7A-B). Estos Ac no mostraron reactividad cruzada con la variante N-acetilada del gangliósido GM3 (Figura 7C-D). Figura 7. Inmunogenicidad del preparado vacunal NGcGM3/VSSP en ratones portadores del tumor 3LL-D122. Los animales fueron inoculados con las células tumorales en la almohadilla plantal el día cero, y posteriormente inmunizados con 200 µg del preparado vacunal NGcGM3/VSSP por vía SC los días 7 y 21. La evaluación de los niveles de Ac antiNGcGM3 se realizó mediante ELISA con los sueros extraídos los días 24, 28 y 35, de cuatro ratones por grupo cada día. Las figuras A-D muestran los valores de absorbancia a 405nm de la dilución 1/100 del suero de los animales, para Ac anti-NGcGM3 tipo IgM (A) e IgG (B), y para Ac anti-NAcGM3 tipo IgM (C) e IgG (D). E: muestra las medianas de los títulos de Ac IgM e IgG anti-NGcGM3 de los ratones portadores del tumor y vacunados. Tres días después de la segunda dosis (día 24 en el gráfico de la Figura 7E) la mediana de los títulos de Ac IgM anti-NGcGM3 fue 1/600, respuesta que en los siguientes 14 días disminuyó hasta 1/200 (día 35 en el gráfico de la Figura 7E). Para la respuesta de Ac IgG anti-NGcGM3, se observó que los títulos se elevaron desde 1/200 tres días después de terminar el esquema de vacunación, hasta 1/600 luego de siete días, volviendo a alcanzar niveles medios de 1/200 a los 14 días de concluido el tratamiento (Figura 7E). 66 Resultados 4.3 El preparado vacunal NGcGM3/VSSP disminuye las metástasis pulmonares en el modelo murino estudiado La actividad antitumoral del preparado vacunal NGcGM3/VSSP se evaluó en el modelo de metástasis espontánea del carcinoma pulmonar de Lewis 3LL-D122. Con este propósito, grupos de 15 ratones C57BL/6 retados con las células tumorales 3LLD122 en la almohadilla plantal, fueron tratados con dos dosis del preparado vacunal NGcGM3/VSSP sin emulsión, o con SSTF (control), por vía SC, siete y 21 días después de la inoculación del tumor. El monitoreo del crecimiento de los tumores primarios en la pata de los animales no mostró ningún efecto del preparado estudiado sobre la velocidad de crecimiento de los mismos, no observándose diferencias estadísticamente significativas en los diámetros de los tumores al comparar las curvas de crecimiento en los animales vacunados y los controles (p>0,05). Como puede distinguirse en la Figura 8A, los tumores aparecieron en los dos grupos a la misma vez, no observándose tampoco diferencias en los tiempos de prendimiento (p>0,05). A los 18 días de implantado el tumor en la almohadilla plantal los tumores tenían un diámetro promedio de 0,4 cm en todos los grupos. La remoción quirúrgica de los mismos se realizó a los 26 días del implante tumoral, cuando el diámetro promedio de los tumores en el grupo control fue superior a 0,8 cm. La incapacidad del preparado vacunal NGcGM3/VSSP de impedir el crecimiento de los tumores primarios se manifestó de forma similar en todas las réplicas experimentales. Para evaluar el efecto del tratamiento sobre las metástasis, los ratones se sacrificaron 21 días después de la cirugía del tumor primario. Como se observa en la Figura 8B, el grupo control tratado con SSTF desarrolló metástasis en pulmones, por los elevados valores de peso obtenidos (media 0,65 g). Los animales vacunados con NGcGM3/VSSP, tuvieron 67 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP significativamente menos metástasis pulmonares (p<0,05), según el peso de los pulmones (media de 0,39 g), comparado con el grupo control. Figura 8. Efecto del preparado vacunal NGcGM3/VSSP sobre las metástasis en pulmón, en el modelo de metástasis espontánea del carcinoma pulmonar de Lewis 3LL-D122. Treinta ratones C57BL/6 se inocularon con células tumorales 3LL-D122 (2x105/ratón) en la almohadilla plantal, de los cuales 15 se trataron con dos dosis del preparado vacunal NGcGM3/VSSP no emulsivo, por vía SC, siete y 21 días después de la inoculación del tumor. Los otros 15 ratones recibieron SSTF (grupo control). A: Cinética de crecimiento de los tumores primarios. Cada punto corresponde con la media de las observaciones y las barras de error corresponden a los errores estándar. Se aplicó la prueba ANOVA de dos vías, con un nivel de significación de p<0,05. B: Evaluación de la metastización a pulmón. Las barras representan los valores de la media y las barras de error corresponden a los errores estándar. Se aplicó la prueba t de Student, con un nivel de significación de p<0,05. La Figura 9 muestra el meta-análisis realizado reuniendo los resultados de actividad antimetastásica de la vacuna NGcGM3/VSSP en 10 experimentos independientes con el modelo murino de metástasis espontánea con 3LL-D122. La agrupación de los experimentos permitió comparar el peso de los pulmones de 72 animales controles (media del peso de pulmón: 0,75 g) y 98 vacunados (media del peso de pulmón: 0,42 g). La aplicación de la prueba t de Student dió un valor de probabilidad inferior a 0,0001. Con el meta-análisis se demostró la solidez del efecto antimetastásico del preparado vacunal NGcGM3/VSSP en el modelo murino de metástasis espontánea con 3LL-D122. 68 Resultados Figura 9. Solidez del efecto antimetastásico del preparado vacunal NGcGM3/VSSP en el modelo murino de metástasis espontánea con 3LLD122. Meta-análisis de los resultados de 10 experimentos independientes. Grupos de 7-10 ratones C57BL/6 se incularon con células tumorales 3LLD122 (2x105/ratón) en la almohadilla plantal y se trataron con dos dosis del preparado en estudio, por vía SC, siete y 21 días después del reto tumoral. Los ratones control recibieron SSTF. Las barras horizontales representan los valores de la media y las barras de error corresponden a los errores estándar. Se aplicó la prueba t de Student con un nivel de significación de 0,0001. En otro grupo de experimentos nos propusimos comparar el efecto obtenido por la administración SC no emulsiva del preparado vacunal NGcGM3/VSSP, con la administración emulsiva en Montanide ISA 51 (Seppic), ya que el mismo ha sido empleado como adyuvante en esquemas de vacunación en cáncer. Luego de la inoculación de las células tumorales 3LL-D122 en la almohadilla plantal de los ratones, éstos fueron tratados con dos dosis del preparado vacunal NGcGM3/VSSP sin adyuvante oleoso (por vía SC), o adyuvado en Montanide ISA 51 (por vía IM), siete y 21 días después de la inoculación del tumor. Los ratones control recibieron SSTF. Como se puede observar en la Figura 10, ambas formulaciones del preparado vacunal NGcGM3/VSSP lograron la disminución de la metastización respecto al grupo control (media peso pulmón de 0,64 g), no observándose diferencias estadísticamente significativas entre los valores obtenidos con la formulación no emulsiva (media 0,37 g) y la emulsiva (media 0,37 g). 69 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP Figura 10. Comparación del efecto del preparado vacunal NGcGM3/VSSP no emulsionado, y emulsionado con Montanide. Ratones C57BL/6 (7-10 por grupo) se inocularon con células tumorales 3LL-D122 (2x105/ratón) en la almohadilla plantal, y se trataron con dos dosis del preparado vacunal NGcGM3/VSSP sin adyuvante oleoso (por vía SC), o adyuvado en Montanide ISA 51 (por vía IM), siete y 21 días después de la inoculación del tumor. Los ratones control recibieron SSTF. Las barras representan los valores de la media y las barras de error corresponden a los errores estándar. Se aplicó el ANOVA y el ensayo de comparaciones múltiples de Tukey, con un nivel de significación de 0,01. Letras diferentes indican diferencias estadísticamente significativas. Se muestra un resultado representativo de 2 experimentos realizados. 4.4 La presencia del gangliósido NGcGM3 en la vacuna es necesaria para la actividad antitumoral del preparado NGcGM3/VSSP Para estudiar la relevancia del gangliósido NGcGM3 en la actividad antitumoral del preparado vacunal NGcGM3/VSSP, se realizaron experimentos antitumorales en el modelo murino de metástasis espontánea de células 3LL-D122, utilizando como controles una preparación construida con el gangliósido NAcGM3 (NAcGM3/VSSP) y otra a partir de las proteínas solubilizadas de las VME de la Nm. Todos los preparados fueron administrados a los animales por vía SC y sin el empleo de adyuvante oleoso. Como se puede observar en la Figura 11, los tratamientos que no incluyeron al gangliósido NGcGM3, dígase VME o NAcGM3/VSSP, no mostraron disminución significativa (p>0,05) 70 Resultados de los pesos de los pulmones al compararlos con el grupo control. Figura 11. Estudio de la relevancia del gangliósido NGcGM3 para la actividad antimetastásica de la vacuna. Tratamiento de ratones C57BL/6 (10 por grupo) en el modelo murino de metástasis espontánea con el preparado vacunal NGcGM3/VSSP, con las VME de Nm y con el preparado NAcGM3/VSSP. Se aplicó el ANOVA y el ensayo de comparaciones múltiples de Tukey, con un nivel de significación de 0,01. Letras diferentes indican diferencias estadísticamente significativas. Se muestra un resultado representativo de 2 experimentos realizados. Los valores de las medias de los pesos de pulmón fueron: 0,67 g para el grupo control, 0,60 g para el tratado con NAcGM3/VSSP y 0,60 g para VME; mientras que para el grupo al que se le administró NGcGM3/VSSP la media de peso pulmonar fue de 0,40 g, valor inferior y estadísticamente diferente a los anteriores. 4.5 La actividad antimetastásica del preparado vacunal NGcGM3/VSSP es independiente de la vía de administración Para evaluar si el efecto antimetastásico del preparado vacunal, en el modelo estudiado depende de la vía de administración, se procedió a inmunizar con NGcGM3/VSSP por vía SC, IM o IP a ratones C57BL/6 que fueron retados con las células tumorales según se ha descrito anteriormente. La evaluación del efecto antimetastásico se realizó a través del pesaje de los pulmones 21 días después de realizar la cirugía de los tumores primarios. En este experimento el grupo control (tratado con SSTF) tuvo una media de peso de pulmón de 0,73 g (Figura 12). 71 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP Figura 12. Efecto del preparado vacunal NGcGM3/VSSP administrado por vía SC, IM o IP, a ratones C57BL/6 (15-16 por grupo) retados con las células tumorales 3LL-D122 en la almohadilla plantal. Las barras representan los valores de la media y las barras de error corresponden a los errores estándar. Se aplicó el ANOVA y el ensayo de comparaciones múltiples de Tukey, con un nivel de significación de 0,0001. Letras diferentes indican diferencias estadísticamente significativas. Se muestra un resultado representativo de 2 experimentos realizados. Como se puede observar, las medias de peso pulmonar de los grupos vacunados fueron de 0,41 g (SC), 0,48 g (IM) y 0,40 g (IP); no existiendo diferencias significativas entre ellos, por lo que el preparado vacunal NGcGM3/VSSP disminuyó la metastización a pulmón independientemente de la vía por la que fue administrado. Los tres grupos vacunados fueron estadísticamente inferiores al grupo control. Este resultado podría indicar la capacidad del preparado vacunal de interactuar con poblaciones de DC residentes en diferentes sitios. 4.6 La vacunación con NGcGM3/VSSP induce la maduración de células dendríticas En el presente trabajo se evaluó in vitro e in vivo el efecto del preparado vacunal NGcGM3/VSSP sobre las DC. En los experimentos in vitro, precursores de médula ósea de ratones C57BL/6 se diferenciaron a cDC con medio rico en GM-CSF. Las cDC obtenidas se estimularon con LPS y con el preparado vacunal NGcGM3/VSSP, y posteriormente se realizó la evaluación 72 Resultados de la expresión de marcadores de maduración en la membrana celular por citometría de flujo con Ac específicos. Como se puede observar en la Figura 13A, el preparado vacunal NGcGM3/VSSP estimuló un 50% de células CD11c+CD40+, similar a como lo hizo el LPS (45%), un inductor bien conocido de maduración de DC. La inducción de células expresando la molécula coestimuladora CD86 por el preparado vacunal NGcGM3/VSSP fue incluso superior (43%) que el provocado por el LPS (28%). La expresión de la molécula CD1d también se examinó en las cDC obtenidas in vitro, debido al papel de esta molécula en la presentación de Ag glicolipídicos a las células NKT (MacDonald, 2002; Speak et al., 2008). Como se observa en la Figura 13A, sólo el 4,5% de las cDC inmaduras fueron CD1d+, mientras que luego del tratamiento con el preparado NGcGM3/VSSP, las DC CD1d+ aumentaron hasta el 27%. A partir del conocimiento de que la IL-6 e IL-12 son citocinas importantes en la atracción de otros mediadores celulares y la direccionalización hacia el patrón de respuesta Th1, respectivamente, se midió la liberación de ambas citocinas en el sobrenadante de las cDC diferenciadas e incubadas con cantidades crecientes del preparado vacunal NGcGM3/VSSP. Como se muestra en la Figura 13B las cDC secretaron altos niveles de IL-6 e IL-12, de manera dosis dependiente y similar a las tratadas con LPS (control positivo). Para confirmar estas evidencias en un escenario in vivo, tres ratones ratones C57BL/6 se vacunaron por vía SC con NGcGM3/VSSP y luego de tres días se extrajeron los ganglios linfáticos aferentes al sitio de inoculación (axilares), para evaluar por citometría de flujo la expresión de las moléculas marcadoras de maduración en las células DC. Este análisis mostró que el porcentaje de las DC en el ganglio linfático de los animales vacunados aumentó tres veces respecto al control, así como también se incrementó la frecuencia de 73 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP DC maduras, lo cual se evidenció al analizar los porcentajes de células expresando las moléculas CD40 (aumentó 10 veces), CD80 (4 veces) y CD86 (4 veces, Figura 13C). Figura 13. Efecto del preparado vacunal NGcGM3/VSSP sobre DC convencionales. A-B: bm-cDC se cultivaron toda la noche con LPS (1µg/ml), NGcGM3/VSSP (1µg/ml) o medio (inmaduras) y se midió la frecuencia de células CD11c+ maduras (A) y la secreción de IL-6 e IL-12p40 (B). C: Ratones C57BL/6 (n=3) se inyectaron por vía SC con NGcGM3/VSSP. El diagrama de barras muestra el porcentaje de células maduras CD11c+ en el linfonodo aferente, tres días después del tratamiento con NGcGM3/VSSP o SSTF (control). Los datos son representativos de dos experimentos independientes. 74 Resultados A partir del resultado de actividad antitumoral por las diferentes vías de administración, se estudió la capacidad de la vacuna de activar y madurar pDC, las cuales son residentes en sangre y esenciales para la secreción de citocinas, como el IFNα que media la activación de otras células del sistema inmunitario. Para ello, se purificaron las pDC del cultivo (en medio enriquecido con FLT3-L) de precursores de médula ósea de ratones C57BL/6. Las pDC obtenidas fueron cultivadas durante toda la noche con LPS, CpG-A y el preparado vacunal NGcGM3/VSSP. Como se puede observar en la Figura 14A, la adición del preparado NGcGM3/VSSP provocó la maduración de las pDC inmaduras obtenidas in vitro, de manera similar a como lo hizo el control positivo LPS y el CpG-A. Sin embargo, cuando se analizó la presencia de IFNα en el sobrenadante del cultivo, sólo el CpG-A fue capaz de inducir la secreción de dicha citocina (Figura 14B). Adicionalmente se administró por vía IP el preparado NGcGM3/VSSP a tres ratones C57BL/6 y se evaluaron los niveles de IFNα, IL-6 e IL-12 en el suero de los animales, en intervalos de 2 h, mediante ELISA. Como se puede observar en la Figura 14C los niveles de IFNα en suero se elevaron rápidamente en los animales tratados con el preparado vacunal, obteniéndose un pico de secreción a las 2h, anterior incluso al pico de IFNα estimulado por el Poli I:C. No se detectaron niveles de IFNγ en los sueros evaluados. La detección in vivo de las citocinas IL-6 e IL-12 confirmaron el hallazgo in vitro del efecto modulador que ejerce la vacuna sobre la funcionalidad de las DC. Estos datos, en conjunto, muestran que el preparado vacunal NGcGM3/VSSP indujo una fuerte respuesta de secreción de citocinas in vivo y la maduración in vitro de las DC convencionales y las plasmacitoides. 75 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP Figura 14. Efecto del preparado vacunal NGcGM3/VSSP sobre DC plasmacitoides. A-B: pDC purificadas a partir de bm-pDC, se cultivaron toda la noche con LPS (0,5 µg/ml), CpGA (5 µg/ml), NGcGM3/VSSP (0,1 µg/ml) o medio (inmaduras) y se midió la frequencia de células B220+ maduras (A) y la secreción de IFNα (B). C: Tres ratones C57BL/6 fueron inyectados por vía IP con NGcGM3/VSSP y se midieron los niveles de IFNα, IL6 e IL12 en intervalos de dos horas en el suero por ELISA. Los datos son representativos de dos experimentos independientes. 4.7 Las células NK y NKT están involucradas en la actividad anti-metastásica del preparado vacunal NGcGM3/VSSP Debido al gran potencial que tiene la respuesta inmune mediada por células NK1.1 para erradicar células tumorales en varios modelos de metástasis en pulmón y teniendo en cuenta la naturaleza glicolipídica del blanco seleccionado, se evaluó el papel de las células NK y las células NKT (específicas por Ag no proteicos), en la actividad antimetastásica demostrada para el preparado vacunal NGcGM3/VSSP en el modelo estudiado. 76 Resultados Se realizó un primer acercamiento mediante experimentos de eliminación de dichas poblaciones celulares, utilizando un AcM que elimina la población NK1.1+ in vivo. Los controles de este experimento fueron el grupo tratado con SSTF (animales que desarrollan metástasis en pulmones) y el grupo vacunado por vía SC con el preparado NGcGM3/VSSP no emulsionado (control de que la vacuna disminuye la metastización), ambos sin administrar el AcM que elimina la población a estudiar. Como se esperaba, el preparado vacunal disminuyó la metastización a pulmón, pues se observaron diferencias estadísticamente significativas entre los animales vacunados (media peso pulmonar: 0,36 g) y los controles (media: 0,64 g) (Figura 15). El papel de las células NK1.1+ se analizó a partir de la comparación entre los grupos control y NGcGM3/VSSP, cuando a los mismos además, se les administró el AcM que eliminó la población NK1.1+ (Figura 15). Cuando los animales se trataron con una dosis de 1 mg de AcM anti-NK1.1 un día después de la administración del preparado vacunal, se observó que el preparado vacunal fue incapaz de disminuir la metastización a pulmón. Los valores de la media del peso de pulmón corresponden a 0,93 g y 1,01 g, para control+anti-NK1.1 y NGcGM3/VSSP+antiNK1.1 respectivamente, valores que compararlos estadísticamente no mostraron diferencias significativas (p > 0,05). Este resultado sugiere que las células con el marcador NK1.1 (tanto NK como NKT) están involucradas en la respuesta inmune contra las metástasis de 3LL-D122, generada a partir de la vacunación con NGcGM3/VSSP, pues al eliminar esta población se pierde el efecto anti-metastásico. A partir de este resultado y con el objetivo de comprobar la movilización de las células con el marcador NK1.1 al pulmón de los animales vacunados, se realizó otro experimento donde animales del modelo de metástasis espontánea a pulmón con las células 3LL-D122 77 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP se trataron con el preparado vacunal NGcGM3/VSSP y tres días después se analizó el infiltrado pulmonar por citometría con el objetivo de detectar la presencia de células NK (NK1.1+CD3-) y NKT (NK1.1+CD3+). Figura 15. Papel de las células NK1.1+ en la actividad antimetastásica del preparado vacunal NGcGM3/VSSP. La figura representa los valores de los pesos de los pulmones en función del tratamiento recibido. Grupos de 15 ratones C57BL/6 portadores del tumor 3LL-D122 en la almohadilla plantal, fueron tratados con el AcM que elimina las células NK1.1+, un día después de cada dosis del preparado vacunal NGcGM3/VSSP. Los grupos controles recibieron SSTF o SSTF+anti-NK1.1. Las barras de columnas representan los valores de la media y las barras de error, la desviación estándar. Se aplicó el ANOVA y el ensayo de comparaciones múltiples de Tukey, con un nivel de significación de 0,05. Letras diferentes indican diferencias estadísticamente significativas. Se muestra un resultado representativo de 2 experimentos realizados. Como se puede observar en la Figura 16, en los animales controles el porcentaje de células NK que infiltraron los pulmones fue el 9,5% y se incrementó 1,8 veces en los animales vacunados (alcanzó el 16.8%). Un resultado similar se obtuvo para la población NKT (NK1.1+CD3+), las cuales fueron el 2,7% de las células de los pulmones en los controles, mientras que en los vacunados alcanzaron el 6,2%, mostrando un incremento de 2,3 veces. 78 Resultados Figura 16. Frecuencia de células NK1.1+ en los pulmones de los ratones vacunados portadores del tumor. Ratones C57BL/6 (n=3) portadores del tumor 3LL-D122, tratados con NGcGM3/VSSP o SSTF (Control), se sacrificaron 3 días después de la segunda administración de vacuna y se midió la frecuencia de células NK and NKT en los pulmones por citometría de flujo. A: Gráficos de puntos de un animal representativo por grupo. B: Gráfico de las frecuencias de células NK o NKT en el infiltrado pulmonar (porcentaje) en tres animales de cada grupo. Los datos son representativos de dos experimentos independientes. Se aplicó la prueba no paramétrica de comparaciones pareadas U de Mann-Whitney (p<0.05). 4.8 Las células CD8 y CD4 participan en la actividad anti-metastásica del preparado vacunal NGcGM3/VSSP El papel de las células de la inmunidad adquirida, CD8 y CD4, sobre la actividad antitumoral del preparado vacunal NGcGM3/VSSP, administrado por vía SC sin emulsión, se valoró también en el modelo murino 3LL-D122 metastásico. En esta dirección, se procedió de manera similar al análisis realizado para las poblaciones efectoras de la inmunidad innata. En un primer grupo de experimentos se analizó el papel de las células CD8. La comparación entre el grupo tratado con SSTF (media peso pulmón 0,67 g) y el grupo vacunado con NGcGM3/VSSP (0,35 g), en ausencia del AcM que elimina la población CD8 (Figura 17A), mostró que el preparado vacunal inhibió la metastización a pulmón. Cuando a ambos grupos de animales se les administró además el AcM que elimina la población CD8, se observó la pérdidad del efecto antimetastásico obtenido con el preparado vacunal NGcGM3/VSSP. En este caso, la media del peso de los pulmones de los 79 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP animales controles fue 0,47 g, y la de los animales vacunados fue 0,53 g, valores que no mostraron diferencias estadísticamente significativas (p > 0,05). Este resultado sugiere que las células CD8 participan en la actividad antimetastásica observada al tratar a los animales con el preparado vacunal NGcGM3/VSSP. Figura 17. Papel de las células T CD8 y T CD4 en la actividad antimetastásica del preparado vacunal NGcGM3/VSSP. La figura representa los valores de los pesos de los pulmones en función del tratamiento recibido. Ratones C57BL/6 portadores del tumor 3LLD122 en la almohadilla planta, fueron tratados con el AcM que elimina las células CD8+ (A) o ,con el AcM que elimina las células CD4+ (B), un día después de cada dosis del preparado vacunal. Los grupos controles recibieron SSTF o SSTF + AcM anti-CD8 o SSTF + AcM antiCD4. Las barras de columnas representan los valores de la media y las barras de error, la desviación estándar. Se aplicó el ANOVA y el ensayo de comparaciones múltiples de Tukey, con un nivel de significación de 0,05. Letras diferentes indican diferencias estadísticamente significativas. Se muestran resultados representativos de 2 experimentos realizados. Para evaluar la participación de las células T CD4 se realizó en un experimento similar, pero esta vez se administró un AcM que elimina la población CD4. Nuevamente la disminución de la metastización del preparado vacunal, en ausencia del AcM que elimina la población CD4, se verificó al observarse diferencias significativas entre el grupo tratado con SSTF (media peso pulmón 0,69 g) y el grupo vacunado con NGcGM3/VSSP (0,36 g) (Figura 17B). Los animales inmunizados con el preparado NGcGM3/VSSP y tratados con el AcM anti-CD4, por el contrario, no disminuyeron significativamente el peso de los pulmones (0,73 g) al compararlos con el grupo control al que también se le inyectó el AcM anti-CD4 (0,97 g, p > 0,05). Este resultado sugiere que las células CD4 están involucradas en la 80 Resultados actividad antimetastásica observada al inmunizar a los animales con el preparado vacunal NGcGM3/VSSP, ya que su ausencia implica la pérdida del efecto. 4.9 La vacunación con NGcGM3/VSSP promueve la infiltración de células efectoras de la inmunidad adquirida al sitio metastásico A partir de la observación de que el efecto anti-metastásico del preparado vacunal NGcGM3/VSSP en el modelo de cáncer de pulmón de Lewis dependía tanto de células T CD4+ como de T CD8+, se procedió a determinar la frecuencia de estas poblaciones celulares en los pulmones de ratones portadores del tumor 3LL-D122, tratados con SSTF o inmunizados por vía SC con NGcGM3/VSSP (sin emulsión). Los animales (4-7 por grupo) se sacrificaron veintiún días después de operados los tumores primarios. Los pulmones se pesaron para comprobar el efecto antimetastásico del preparado vacunal, y se procesaron para determinar la frecuencia de leucocitos mediante citometría de flujo. La mediana de peso de pulmón del grupo control fue de 0,94 g, mientras que la del grupo vacunado fue de 0,34 g, diferencia que resultó estadísticamente significativa al aplicar la prueba de MannWhitney (p < 0,005). El porcentaje de células CD45+ infiltrantes del pulmón fue significativamente superior en los pulmones de los ratones vacunados (74,7%) comparado con los controles (28,5%) (Figura 18A). Este incremento también se encontró en las poblaciones de células T, identificadas como CD3+CD45+, CD3+CD4+ y CD3+CD8+, siendo en este orden: el 5,1%, el 1,0% y el 0,6% de las células infiltrantes del pulmón de los animales controles, para alcanzar en el mismo orden: el 23,1%, el 8,0% y el 2,8%, de las células infiltrantes del pulmón de los animales vacunados (Figura 18A). Se analizó adicionalmente el porcentaje de células T reguladoras (Tregs: CD4+Foxp3+) y de células supresoras de origen mieloide (CD11b+Gr1+). 81 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP Figura 18. Frecuencia en pulmón de efectores celulares de la inmunidad adaptativa inducidos por la vacunación SC con NGcGM3/VSSP en animales portadores de tumor de Lewis 3LL-D122. Grupos de 4-6 ratones C57BL/6, retados con las células 3LL-D122, controles o vacunados (Vac) fueron sacrificados 21 días después de operados los tumores primarios. La frecuencia de células efectoras y reguladoras fue determinada en los pulmones de los animales por citometría de flujo. A: Frecuencias de células CD45+, CD3+, CD4+ o CD8+ en el total de células infiltrantes (porcentaje) en ratones vacunados y controles. B: Frecuencia de poblaciones regulatorias, arriba: células T reguladoras (CD4+Foxp3+) y abajo: células supresoras de origen mieloide (CD11b+Gr1+). Se empleó la prueba no paramétrica de comparaciones pareadas U de Mann-Whitney, con un nivel de significación de 0,01. Se muestran resultados representativos de 2 experimentos realizados. La frecuencia de estas poblaciones reguladoras, a diferencia de las anteriores, fue similar en ambos grupos de animales, no mostrando diferencias estadísticamente significativas en ninguno de los dos casos. Las células Tregs representaron el 2,1% y el 4,4% de las células infiltrantes del pulmón de los animales controles y vacunados, respectivamente. Los porcentajes de células MDSC infiltrantes de pulmón fueron 2,3% y 5,3%, en animales controles y vacunados, respectivamente (Figura 18B). 82 Resultados 4.10 El infiltrado de leucocitos correlaciona con la inhibición de la metastización provocada por la vacunación con NGcGM3/VSSP Para verificar el posible vínculo entre el infiltrado inmune inducido por la vacunación y el efecto anti-metastásico, se correlacionaron los porcentajes de células CD45+, CD3+CD45+, CD3+CD4+ y CD3+CD8+ obtenidos y los correspondientes pesos de los pulmones (como subrogado de la metastización) de los ratones individuales. Mientras que los ratones portadores de tumor no vacunados mostraron elevados pesos de pulmones (alta metastización, mediana 0,94 g) con un bajo porcentaje de leucocitos y células T infiltrantes, en los ratones vacunados los bajos pesos de pulmones (pocas metástasis, mediana 0,34 g) estuvo asociado con un incremento en el porcentaje de éstas poblaciones (Figura 19A-B). El análisis estadístico de estas dos variables reveló coeficientes r de correlación de Pearson negativos, oscilando entre 0,855 y 0,498 (Figura 19C-D, Tabla 2), lo cual establece una relación inversa entre el infiltrado celular inducido por la vacunación y la metastización del tumor. La significación de la correlación obtenida (p < 0,05) para las células infiltrantes CD45+, CD3+CD45+y CD3+CD4+ sostiene la idea de que estas células, correctamente movilizadas a los pulmones en los animales vacunados, fueron responsables de controlar la metastización. No se alcanzó significación para la correlación con las células CD3+CD8+. Tabla 2. Coeficientes r de Pearson y valores de probabilidad de la correlación entre el infiltrado de células efectoras de la inmunidad adaptativa, y la inhibición de la metastización inducida por la vacunación SC con NGcGM3/VSSP. % CD45+ % CD3+CD45+ %CD3+CD4+ %CD3+CD8+ Coeficiente r de Pearson -0,8553 -0,7489 -0,6609 -0,4975 Valor de probabilidad 0,0002 0,0032 0,0139 0,0837 83 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP Figura 19. Correlación entre el infiltrado de células efectoras de la inmunidad adaptativa, y la inhibición de la metastización inducida por la vacunación SC con NGcGM3/VSSP. Ratones C57BL/6 controles o tratados con NGcGM3/VSSP por vía SC, retados con las células 3LL-D122, se sacrificaron 21 días después de operar los tumores primarios y se correlacionó la frecuencia de células (A-C) CD45+, CD3+, (B-D) CD4+ y CD8+ presentes en los pulmones de los animales, con la metastización en pulmón (cuantificado como peso del pulmón). A-B: representación de la relación entre el porcentaje de células infiltrantes y el valor del peso de pulmón por grupo de animales (control o vacunado), C-D: correlación entre el porcentaje de células infiltrantes y el peso de pulmón por población celular en el total de los animales (control + vacunado). Se muestran resultados representativos de 2 experimentos realizados. 84 5. DÍSCUSÍON Muchas estrategias se han llevado a cabo a lo largo de los años con el objetivo de controlar el cáncer, debido al problema de salud que representa esta enfermedad a nivel mundial. Entre ellas está la IT activa de cáncer, la cual se basa en el empleo de vacunas que induzcan una respuesta inmune específica al Ag tumoral, capaz de eliminar el tumor y proteger al paciente de la aparición de metástasis, con un beneficio en la supervivencia. Dentro de estas, las vacunas basadas en gangliósidos han sido objeto de diferentes estudios al tener en cuenta la expresión alterada de dichas moléculas en las células tumorales (Ravindranath et al., 2003; Kavanagh et al., 2005) y a su baja inmunogenicidad (Livingston, 1995a). En el presente trabajo se estudió el mecanismo de acción del preparado vacunal NGcGM3/VSSP, cuyo blanco es el gangliósido NGcGM3 expresado específicamente en varios tipos de tumores en humanos (Carr et al., 2000; Oliva et al., 2006; Blanco et al., 2011a; Scursoni et al., 2011; Blanco et al., 2012; Zhong et al., 2012; Blanco et al., 2013). Para este estudio se empleó el modelo murino de metástasis espontánea a pulmón con la línea 3LL-D122 (Eisenbach et al., 1984). El modelo tiene un comportamiento muy similar a la enfermedad metastásica que se presenta en los pacientes de cáncer, donde en muchos casos las lesiones metastásicas se hacen evidentes un tiempo después de la cirugía de los tumores primarios, y son las metástasis las que causan la muerte del paciente. En este modelo tumoral no se había descrito la expresión del gangliósido NGcGM3. Por lo tanto, primeramente se definió la presencia del gangliósido NGcGM3 en las etapas por las que transita el modelo: línea celular, tumor primario y metástasis pulmonares. Como primer resultado se encontró que el modelo fue coherente con el incremento de la expresión del gangliósido NGcGM3 desde el tumor primario hasta la lesión metastásica en 85 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP pulmón (Figura 1), como se ha observado en muestras de pacientes de cáncer (Blanco et al., 2011b; Hayashi et al., 2013). Este resultado permite su uso como modelo relevante para evaluar el efecto de terapias antitumorales basadas en el gangliósido NGcGM3. Los resultados obtenidos, relativos al cambio del patrón de sialilación, coinciden con los estudios realizados por El-Abbadi y cols. (2001) en un modelo tumoral de metastización espontánea a cerebro. Estos autores mostraron resultados análogos con respecto al contenido total de AS en los gangliósidos de tumores que crecían subcutáneos (70% del AS estaba en la forma de ácido NGcNeu) en relación con las células tumorales mantenidas en cultivo (casi exclusivamente AS NAcNeu) (El-Abbadi et al., 2001). Además, otros autores han asociado la capacidad metastásica de las células tumorales con el patrón de expresión de gangliósidos y con cambios en la sialilación (Alessandri et al., 1987; Coulombe et al., 1993; Makatsori et al., 1998). Yin y cols. (2010) propusieron que los cambios en la sialilación se debían a la hipoxia que se genera en los tumores localmente avanzados y demostraron que en condiciones de hipoxia se sobre-expresa el gen de la sialina, lo que provoca el aumento de la expresión de gangliósidos N-acetilados y N-glicolilados en las células tumorales (Yin et al., 2010). La aparición de gangliósidos N-glicolilados en ratones está favorecida además por la acción de la enzima CMAH, que convierte el AS N-acetilado en N-glicolilado (Hedlund et al., 2007; Chandrasekharan et al., 2010), mientras que en pacientes se ha propuesto que es la dieta la fuente de AS N-glicolilado (Tangvoranuntakul et al., 2003). La hipoxia afecta además la transcripción de genes asociados con la síntesis de la ceramida (Yin et al., 2010), lo que explicaría los cambios en la movilidad de las bandas en HPTLC del gangliósido NAcGM3 presente en los tumores SC y en las metástasis (Figura 1B-C). 86 Discusión El resultado obtenido en esta tesis, referente al incremento de la expresión del gangliósido NGcGM3 en el modelo de metastización espontánea a pulmón con el carcinoma de Lewis 3LL-D122, apoya los resultados demostrados por otros autores acerca del papel del gangliósido NGcGM3 en la progresión tumoral (de Leon et al., 2006; de Leon et al., 2008; Gabri et al., 2009; Segatori et al., 2012a; Segatori et al., 2012b; Casadesus et al., 2013). El empleo del gangliósido NGcGM3 sintético en el preparado vacunal confirmó el valor de esta molécula como blanco de la IT de cáncer. Los resultados alcanzados demostraron que los perfiles de inmunogenicidad contra el gangliósido NGcGM3 fueron equivalentes independientemente del origen del Ag (Figura 5). Este resultado permite el desarrollo de un producto vacunal más consistente desde un enfoque regulatorio. Estudios previos de inmunización de ratones con otros candidatos vacunales basados en gangliósidos también han sido exitosos en la generación de respuesta humoral contra estas moléculas (Livingston et al., 1987; Livingston et al., 1989a; Livingston et al., 1989b; Livingston et al., 1993; Dumontet et al., 1997; Dickler et al., 1999). La respuesta de Ac antigangliósidos ha sido fundamentalmente de isotipo IgM, y estos Ac han mediado la protección en modelos animales retados con tumores que expresaban el gangliósido blanco (Zhang et al., 1998). Los resultados obtenidos en la presente tesis, relativos a la inmunogenicidad del preparado en pollos y ratones, coinciden con los resultados mostrados por Estévez y cols. (1999), quienes describieron la generación de respuestas IgM e IgG con preparaciones de los gangliósidos NAcGM3, NGcGM3 y GD3, en VSSP y adyuvante oleoso (Estevez et al., 1999). Adicionalmente, los resultados del actual trabajo de tesis mostraron como los Ac de los sueros inmunes de los pollos vacunados con el preparado reconocieron a las células de mieloma P3X63, que son gangliósido NGcGM3 positivas (Figura 5B). El ensayo clínico fase 87 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP I realizado con el preparado vacunal NGcGM3/VSSP /Montanide en pacientes de cáncer de mama avanzado evidenció como todos los pacientes tratados desarrollan Ac anti-NGcGM3 (IgM e IgG) que median la citotoxicidad dependiente de complemento contra la línea tumoral de mieloma P3X63, así como una marcada reactividad frente a muestras del carcinoma ductal de mama (Carr et al., 2003). Estudios clínicos posteriores con el preparado vacunal NGcGM3/VSSP /Montanide demostraron que los Ac anti-NGcGM3 generados por la vacunación, también son capaces de prevenir la disminución de la expresión de la molécula CD4 en los linfocitos T, provocado por el gangliósido NGcGM3 (de Leon et al., 2006; Mulens et al., 2010). Adicionalmente, el hallazgo de que AcM antiNGcGM3 median la muerte de las células tumorales por mecanismos independientes de la acción del complemento (Roque-Navarro et al., 2008; Casadesus et al., 2013) permite especular que los Ac anti-NGcGM3 generados por la administración del preparado vacunal NGcGM3/VSSP pudieran iniciar eventos de MCI que favorezcan la respuesta antitumoral. Estudios futuros permitirán profundizar en el papel de los Ac generados con la vacunación. En el presente trabajo se demostró el beneficio terapéutico de la administración del preparado vacunal NGcGM3/VSSP en los ratones portadores del tumor de Lewis 3LL-D122, en el modelo de metastización espontánea a pulmón. Dos administraciones subcutáneas del preparado vacunal en el escenario neoadyuvante (antes de la cirugía del tumor) sin adyuvante oleoso, inhibieron las metástasis espontáneas en los pulmones de los animales tratados (Figura 9). Conocer que el preparado NGcGM3/VSSP tiene una potente actividad antimetastásica en régimen de administración terapéutico y sin adyuvante oleoso es el principal aporte práctico de este trabajo. En las modalidades de vacunación de pacientes con cáncer donde se ha usado un adyuvante oleoso, se ha preferido el Montanide ISA 51 (Aucouturier et al., 2002; Aucouturier et al., 2006). Sin embargo, el Montanide ISA 51 ha dado lugar a la aparición de induraciones, enrojecimiento y dolor en el sitio de inoculación, 88 Discusión además de entorpecer el movimiento de los efectores de células T CD8 + generados por vacunación hacia el microambiente tumoral (Hailemichael et al., 2013). Por todo esto una preparación vacunal que no contenga este componente y que además mantenga sus propiedades antitumorales sería un mejor candidato a considerar para ser aplicado en pacientes de cáncer. Otro grupo de experimentos realizados en el modelo metastásico demostraron la relevancia de la presencia del gangliósido NGcGM3 en el preparado vacunal, para lograr el efecto antitumoral observado. En este sentido, los experimentos con el preparado NAcGM3/VSSP y con las VME de la Nm mostraron que la respuesta antimetastásica frente a la terapia fue dependiente de la inclusión del Ag en el preparado (Figura 11). Estos resultados, unidos al incremento de la expresión del gangliósido NGcGM3 en las metástasis (73.4% de los monosialo-gangliósidos) con respecto a los tumores primarios (32.2%), demostraron que la presencia del gangliósido NGcGM3 en el tumor es crítica para la actividad antitumoral del preparado vacunal NGcGM3/VSSP. Un resultado comparable fue descrito por Segatori y cols. (Segatori et al., 2012a), al clonar el ARN mensajero murino del gen de la enzima CMAH, que cataliza la síntesis del AS N-glicolil neuroamínico, y transfectar la línea celular murina de melanoma B16. Al usar las células transfectadas (B16-H) como modelo tumoral NGcGM3-positivo para la evaluación de la actividad terapéutica del preparado vacunal NGcGM3/VSSP, estos autores demostraron que la presencia del gangliósido NGcGM3 es crítica para la actividad antitumoral, ya que sólo observaron el efecto en aquellos animales que desarrollaron los tumores de la línea B16-H y no en aquellos donde se implantaron las células tumorales parentales B16 (NGcGM3 negativas, NAcGM3 positivas) o las B16-H que habían perdido la expresión del gangliósido NGcGM3. Estos resultados sustentan la necesidad de pre-definir la presencia del gangliósisdo NGcGM3 en las muestras de 89 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP biopsisas de los tumores de los pacientes que eventualmente serán sometidos a tratamiento. La inmunidad protectiva es el resultado de la interrelación de dos sistemas fundamentales: la inmunidad innata no Ag específica y la inmunidad adquirida Ag específica. Un vínculo esencial entre la inmunidad innata y la adaptativa es suministrado por las células presentadoras de Ag, entre las cuales las DC son las más eficientes inductoras de las respuestas inmunes. Por otra parte, se conoce que la ruta de administración es un factor crítico que influye la respuesta inmune asociada a la vacunación. En este trabajo, se describió que el efecto antimetastásico obtenido con el preparado vacunal NGcGM3/VSSP sobre el modelo de metástasis espontánea utilizado, es independiente de la vía de administración (Figura 12). Este resultado inesperado, nos permitió sugerir que el preparado vacunal sería capaz de activar poblaciones de DC residentes en diferentes sitios. En este sentido, se conoce que las rutas SC e IM involucran típicamente a las cDC y que la vía IP implica a las pDC. Nuestros resultados mostraron que el preparado vacunal NGcGM3/VSSP promovió el incremento, la maduración y la secreción de IL-12 e IL-6 por las cDC. La IL-12 producida por las DC es crucial para el desarrollo de respuestas tipo Th1 (Chamilos et al., 2008; Gilliet et al., 2008b). Por otra parte, se conoce que la producción de IL-6 por las DC es crítica para la activación de células T y que da lugar a células T Agespecíficas, que son refractarias a la actividad supresora de las células T (Kmieciak et al., 2011). Nuestros resultados están de acuerdo con los publicados por Mesa y cols. (2006), quienes con la variante acetilada del gangliósido NAcGM3 incluido en las VME de Nm (NAcGM3/VSSP), demostraron una potente activación y secreción de citocinas por las DC después del tratamiento con el preparado (Mesa et al., 2006b). En conjunto, estos resultados sugieren que el componente bacteriano del preparado vacunal es importante para la activación y la funcionalidad de las APC. 90 Discusión En contraste con las cDC, las pDC han sido descritas como células secretoras de IFN tipo I luego de la estimulación a través de sus receptores endosomales TLR, lo cual constituye la línea inicial de defensa del hospedero frente a las infecciones virales (Gilliet et al., 2008a; Swiecki et al., 2010). Varios estudios han indicado que las pDC activadas, asociadas al tumor, son importantes para la regresión de tumores en modelos murinos (Vermi et al., 2011). En esta dirección, un resultado significativo fue el obtenido al demostrar que el preparado vacunal NGcGM3/VSSP promovió la maduración de las pDC diferenciadas a partir de precursores de médula ósea (Figura 14). Sin embargo, esas pDC maduras no produjeron niveles detectables de IFNα en el sobrenadante del cultivo. El rápido incremento de los niveles de IFNα en el suero de los animales luego de la administración IP del preparado vacunal, sugiere la activación in vivo de las pDC por una vía indirecta, o la participación de otras poblaciones celulares, como los macrófagos, que son capaces de secretar IFNα en respuesta a determinados estímulos. Otro resultado importante es el aumento de la expresión de la molécula CD1d en las cDC derivadas de precursores de médula ósea, luego de la incubación con NGcGM3/VSSP, al considerar la naturaleza glicolipídica del gangliósido NGcGM3 (Figura 13A). La expresión de la molécula CD1d es esencial para la selección y activación de las células NKT (Moodycliffe et al., 2000; MacDonald, 2002). En ratones, la molécula CD1d ha sido detectada en APC profesionales, incluyendo monocitos, macrófagos y algunas poblaciones de DC (Spada et al., 2000; Kmieciak et al., 2011). En la literatura se ha descrito que los niveles de expresión de la molécula CD1d pueden ser modulados durante procesos infecciosos (Lin et al., 2005). Skold y cols. (2005) revelaron que la expresión de la molécula CD1d en la superficie celular de las APC es regulada y afecta la activación de células T bajo determinadas condiciones fisiológicas (Skold et al., 2005). El aumento de la expresión de la molécula CD1d depende de dos señales, una dada por el IFNγ y la otra mediada por 91 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP productos microbianos o por la citocina proinflamatoria TNF. Asimismo, Raghuraman y cols. (Raghuraman et al., 2006) demostraron que la infección intracelular con el patógeno Listeria monocitogenes conduce a la sobre-expresión de la molécula CD1d, y que además el IFNβ mediaba esta sobre-expresión. Los cambios en la expresión de la molécula CD1d en la superficie de las DC soportan la idea de la activación de células NKT durante la inmunización con el preparado vacunal NGcGM3/VSSP. En nuestro trabajo se comprobó mediante el experimento de eliminación de poblaciones celulares, que la actividad antimetastásica del preparado vacunal NGcGM3/VSSP en ratones C57BL/6 portadores del tumor de Lewis 3LL-D122, dependía de las células NK1.1 positivas (Figura 15). Los resultados obtenidos mostraron un aumento de la población celular NK en los pulmones de estos animales, lo que constituye la primera evidencia de movilización de células efectoras del sistema inmunitario innato por esta vacuna, al órgano donde se efectúa la respuesta antimetastásica. Una gran cantidad de estudios clínicos y pre-clínicos se han enfocado a la inducción de respuesta celular contra el tumor, la cual incluye la generación de células NK y linfocitos CTL específicos. Sin embargo, existen pocos estudios que demuestren la capacidad que tienen las vacunas, ya sean celulares o moleculares, para movilizar células NK. En este sentido, se informó por Chaput y cols. (Chaput et al., 2006) que un preparado vacunal compuesto por exosomas derivados de DCs expande células NK activadas en pacientes con melanoma y cáncer de pulmón. Otros autores con la vacuna Oncophage, compuesta por la proteína de estrés térmico gp-96 derivada de melanoma o colon, lograron la activación de células NK en los pacientes tratados, lo cual contribuye al desarrollo de una respuesta primaria de células T y a una disminución del tumor (Pilla et al., 2006). 92 Discusión Para el caso particular de las vacunas que contienen como entidad molecular a los gangliósidos no existen estudios relacionados con la movilización y activación de células NK. En la literatura consultada se encontró un trabajo que demostró el aumento de células NK activadas en ratones transgénicos para Ac IgM contra el GD2, retados con el tumor EL4 que expresa dicho gangliósido. Debido a este hallazgo, los autores especularon que las células NK pudieran ser activadas por Ac anti-gangliósidos o que dichos Ac, al neutralizar el GD2 secretado por los tumores, pudieran revertir la supresión de las células NK (Kawashima et al., 2003). Teniendo en cuenta que la vacuna NGcGM3/VSSP, contiene como entidad molecular un gangliósido, el cual pudiera ser presentado en la molécula CD1d y producir activación de las células NKT, se decidió estudiar el comportamiento de esta población linfocitaria una vez vacunados los animales portadores del tumor 3LL-D122. En nuestro trabajo se obtuvo un aumento estadísticamente significativo de la frecuencia de las células NKT en los pulmones de los animales tratados que fueron vacunados con NGcGM3/VSSP en comparación con el grupo control (Figura 16). En ratones las células NKT son capaces de reconocer al gangliósido GD3 (Wu et al., 2003) y al NAcGM3 (Park et al., 2008), presentados en la molécula CD1d y dar una respuesta contra estos Ag. Por otra parte, las células NKT desempeñan un importante papel en la respuesta contra tumores (Zhang et al., 2006). Estudios sucesivos serían necesarios para profundizar la relación entre el gangliósido NGcGM3 y la biología de las células NKT. Los linfocitos CTL no están relacionados con la respuesta immune a los gangliósidos debido a la naturaleza glicolipídica de estas moléculas (Livingston, 1995b). Sin embargo, el hallazgo de vitiligo en tres de 12 pacientes de melanoma cutáneo avanzado tratados con el preparado vacunal NGcGM3/VSSP (Osorio et al., 2008), propició la búsqueda de esta población en los animales vacunados portadores del tumor de Lewis. Los resultados 93 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP obtenidos con el modelo murino empleado, sugieren la contribución de una respuesta inmune celular CTL en el efecto antimetastásico generado por la vacunación con NGcGM3/VSSP. Luego de utilizar experimentos de eliminación específica de poblaciones celulares in vivo, en el presente trabajo de tesis se demostró la participación de las células T CD8+ y T CD4+ en la capacidad del preparado vacunal de inhibir la metastización de las células 3LL-D122 a pulmón. El efecto antimetastásico del preparado vacunal NGcGM3/VSSP se abrogó en los animales deficientes de las células T CD8+ o T CD4+ (Figura 17). A partir de este resultado, se evaluó la frecuencia de estas poblaciones en los pulmones de los animales portadores del tumor 3LL-D122, veintiún días después de eliminar mediante cirugía los tumores primarios. Los resultados alcanzados mostraron un incremento del infiltrado de células CD45+ en los pulmones de los animales vacunados con respecto a los animales controles, así como también se observó el incrementó del porcentaje de los linfocitos T CD4+ y T CD8+ (Figura 18). Mazorra y cols. (2008) realizaron experimentos similares de eliminación in vivo de poblaciones inmunes, en el modelo tumoral de melanoma, mediante la administración preventiva de un preparado análogo basado en el gangliósido NAcGM3, y demostraron la contribución de las células T CD8+, pero no de células T CD4+, ni de células NK1.1+ en la fase efectora de la respuesta inmune antitumoral (Mazorra et al., 2008). Se ha descrito que el análisis, localización y orientación funcional de las diferentes poblaciones celulares inmunes infiltrando los tumores humanos, conocido como el inmuno contexto (Galon et al., 2007), ha permitido la identificación de componentes del inmuno contexto cuya presencia puede ser utilizada como marcador pronóstico para los pacientes. Los resultados obtenidos en esta tesis constituyen las primeras evidencias de la movilización de células efectoras del sistema inmunitario innato y adaptativo hacia el sitio 94 Discusión metastásico en ratones portadores de tumor, por una vacuna basada en un gangliósido. De la misma forma, el infiltrado de leucocitos en los pulmones metastásicos, así como de células TCD4+, correlacionó con la inhibición de la metastización provocada por la vacunación con NGcGM3/VSSP (Figura 19). Varios autores han encontrado una fuerte asociación entre el infiltrado linfocítico en los tumores y el beneficio clínico, en diferentes tipos de cáncer, donde se incluye el cáncer de pulmón. Recientemente, en tumores provenientes de pacientes con cáncer colo-rectal se demostró una clara asociación entre la alta densidad de células totales T CD3+, T CD8+ citotóxicas y células T CD45RO+ de memoria, con una mayor supervivencia libre de enfermedad y supervivencia total (Galon et al., 2006; Fridman et al., 2012). No obstante, el análisis del efecto de las poblaciones de células T CD4+ en la respuesta clínica ha derivado en aparentes resultados contradictorios, por lo que la participación de estas células o sus efectos sobre las respuestas antitumorales continúa siendo materia de debate en la actualidad. Estudios futuros serán necesarios para estudiar la actividad funcional y la especificidad de la respuesta generada. El análisis conjunto de los resultados obtenidos en esta tesis permiten pensar que en el mecanismo de acción del preparado vacunal NGcGM3/VSSP los Ac anti-NGcGM3 serían los efectores primarios que facilitarían la interacción de las DC con los tumores y desencadenarían una respuesta efectora de las células NK1.1+ y de las células de la inmunidad adaptativa. En este sentido se propone que los Ac anti-NGcGM3 generados por la administración del preparado NGcGM3/VSSP en individuos con tumores que porten el gangliósido NGcGM3, secuestren al gangliósido y ayuden a restaurar la inmunidad del hospedero, a la vez que desencadenen la lisis de las células tumorales por mecanismos diversos que incluyan la MCI. Los Ac anti-NGcGM3 generados pudieran además ayudar a las células NK1.1+ estimuladas por la vacunación a destruir las células tumorales, lo que liberaría nuevos Ag, así como mediadores solubles o de superficie, que constribuirán a la 95 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP atracción y estimulación de las DC activadas por la vacunación. Este ambiente inflamatorio propiciará la interacción entre las células de la inmunidad innata y el desencadenamiento de la respuesta de células efectoras de la inmunidad adaptativa (ver esquema en Anexo 1). 96 6. CONCLUSÍONES 1. El carcinoma pulmonar de Lewis 3LL-D122 aumenta la expresión del gangliósido NGcGM3 en la misma medida que se incrementa el grado de malignidad, lo que lo hace un modelo apropiado para el estudio de terapias contra el cáncer basadas en este Ag tumoral. 2. El preparado vacunal NGcGM3/VSSP es inmunogénico en animales y los perfiles de inmunogenicidad son similares para el gangliósido NGcGM3 de origen natural y sintético, lo cual propicia su desarrollo como vacuna de cáncer. 3. La vacuna terapéutica NGcGM3/VSSP posee un efecto anti-metastásico en el modelo de carcinoma pulmonar de Lewis 3LL, independientemente del carácter emulsivo de la formulación empleada y de la ruta de administración, y dependiente de la presencia del Ag en el preparado vacunal, lo que refuerza su valía como medio para la ITAE de cáncer. 4. La actividad anti-metastásica de la vacuna NGcGM3/VSSP depende de la participación de efectores celulares de la inmunidad adaptativa y de la inmunidad innata. 5. El efecto anti-metastásico de la vacuna NGcGM3/VSSP depende significativamente de la movilización de células TCD4+ en el órgano diana de la metastización, lo cual le incorpora un nuevo atributo relativo a su capacidad inmunomoduladora. 97 7. RECOMENDACÍONES 1. Estudiar el papel de los anticuerpos en la actividad antitumoral del preparado vacunal NGcGM3/VSSP. 2. Identificar si el preparado vacunal NGcGM3/VSSP moviliza células NKT con especificidad por el gangliósido NGcGM3. 3. Estudiar la especificidad de las células de la inmunidad adquirida involucradas. 4. Estudiar la participación de otras poblaciones de la inmunidad innata, como los macrófagos. 99 8. BÍBLÍOGRAFÍA 1. (1978). "The nomenclature of lipids (Recommendations 1976) IUPAC-IUB Commission on Biochemical Nomenclature." Biochem J 171(1): 21-35. 2. Aebischer, T., Wolfram, M., Patzer, S. I., Ilg, T., Wiese, M. and Overath, P. (2000). "Subunit vaccination of mice against new world cutaneous leishmaniasis: comparison of three proteins expressed in amastigotes and six adjuvants." Infect Immun 68(3): 1328-1336. 3. Aguilar, F. F., Barranco, J. J., Fuentes, E. B., Aguilera, L. C., Saez, Y. L., Santana, M. D., Vazquez, E. P., Baker, R. B., Acosta, O. R., Perez, H. G. and Nieto, G. G. (2012). "Very small size proteoliposomes (VSSP) and Montanide combination enhance the humoral immuno response in a GnRH based vaccine directed to prostate cancer." Vaccine 30(46): 6595-6599. 4. Akiyama, Y., Tanosaki, R., Inoue, N., Shimada, M., Hotate, Y., Yamamoto, A., Yamazaki, N., Kawashima, I., Nukaya, I., Takesako, K., Maruyama, K., Takaue, Y. and Yamaguchi, K. (2005). "Clinical response in Japanese metastatic melanoma patients treated with peptide cocktailpulsed dendritic cells." J Transl Med 3(1): 4. 5. Al-Bader, T., Jolley, K. A., Humphries, H. E., Holloway, J., Heckels, J. E., Semper, A. E., Friedmann, P. S. and Christodoulides, M. (2004). "Activation of human dendritic cells by the PorA protein of Neisseria meningitidis." Cell Microbiol 6(7): 651-662. 6. Alessandri, G., Filippeschi, S., Sinibaldi, P., Mornet, F., Passera, P., Spreafico, F., Cappa, P. M. and Gullino, P. M. (1987). "Influence of gangliosides on primary and metastatic neoplastic growth in human and murine cells." Cancer Res 47(16): 4243-4247. 7. Alexandroff, A. B., Jackson, A. M., O'Donnell, M. A. and James, K. (1999). "BCG immunotherapy of bladder cancer: 20 years on." Lancet 353(9165): 1689-1694. 8. Alfonso, M. and Zeuthen, J. (1996). "Generation of human monoclonal antibodies against ganglioside antigens and their applications in the diagnosis and therapy of cancer." Acta Oncol 35(3): 287-295. 9. Alfonso, S., Diaz, R. M., de la Torre, A., Santiesteban, E., Aguirre, F., Perez, K., Rodriguez, J. L., Barroso Mdel, C., Hernandez, A. M., Toledo, D., Gabri, M. R., Alonso, D. F., Viada, C., Gomez, R. E., Suarez, E., Vazquez, A. M., Perez, R. and Macias, A. E. (2007). "1E10 anti-idiotype vaccine in non-small cell lung cancer: experience in stage IIIb/IV patients." Cancer Biol Ther 6(12): 18471852. 10. Alfonso, S., Valdes-Zayas, A., Santiesteban, E. R., Flores, Y. I., Areces, F., Hernandez, M., Viada, C. E., Mendoza, I. C., Guerra, P. P., Garcia, E., Ortiz, R. A., de la Torre, A. V., Cepeda, M., Perez, K., Chong, E., Hernandez, A. M., Toledo, D., Gonzalez, Z., Mazorra, Z., Crombet, T., Perez, R., Vazquez, A. M. and Macias, A. E. (2014). "A randomized, multicenter, placebo-controlled clinical trial of racotumomab-alum vaccine as switch maintenance therapy in advanced non-small cell lung cancer patients." Clin Cancer Res 20(14): 3660-3671. 101 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 11. Alonso, D. F., Gabri, M. R., Guthmann, M. D., Fainboim, L. and Gomez, D. E. (1999). "A novel hydrophobized GM3 ganglioside/Neisseria meningitidis outer-membrane-protein complex vaccine induces tumor protection in B16 murine melanoma." Int J Oncol 15(1): 59-66. 12. Alves, F., Borchers, U., Keim, H., Fortte, R., Olschimke, J., Vogel, W. F., Halfter, H. and Tietze, L. F. (2002). "Inhibition of EGF-mediated receptor activity and cell proliferation by HK1-ceramide, a stable analog of the ganglioside GM3-lactone." Glycobiology 12(8): 517-522. 13. Ambrosino, D. M., Bolon, D., Collard, H., Van Etten, R., Kanchana, M. V. and Finberg, R. W. (1992). "Effect of Haemophilus influenzae polysaccharide outer membrane protein complex conjugate vaccine on macrophages." J Immunol 149(12): 3978-3983. 14. Amon, R., Reuven, E. M., Leviatan Ben-Arye, S. and Padler-Karavani, V. (2014). "Glycans in immune recognition and response." Carbohydr Res. 15. Andrade, C. M., Ziulkoski, A. L., dos Santos, A. X., Boldrini, L. C., Trindade, V. M., Borojevic, R. and Guma, F. C. (2006). "Gangliosides of the stroma layer participate in the interferon-gamma receptor-dependent controls of myelopoiesis." Biochim Biophys Acta 1761(9): 1030-1040. 16. Angata, T., Varki, N. M. and Varki, A. (2001). "A second uniquely human mutation affecting sialic acid biology." J Biol Chem 276(43): 40282-40287. 17. Arnold, D., Faath, S., Rammensee, H. and Schild, H. (1995). "Cross-priming of minor histocompatibility antigen-specific cytotoxic T cells upon immunization with the heat shock protein gp96." J Exp Med 182(3): 885-889. 18. Aucouturier, J., Ascarateil, S. and Dupuis, L. (2006). "The use of oil adjuvants in therapeutic vaccines." Vaccine 24 Suppl 2: S2-44-45. 19. Aucouturier, J., Dupuis, L., Deville, S., Ascarateil, S. and Ganne, V. (2002). "Montanide ISA 720 and 51: a new generation of water in oil emulsions as adjuvants for human vaccines." Expert Rev Vaccines 1(1): 111-118. 20. Basu, S., Binder, R. J., Suto, R., Anderson, K. M. and Srivastava, P. K. (2000). "Necrotic but not apoptotic cell death releases heat shock proteins, which deliver a partial maturation signal to dendritic cells and activate the NF-kappa B pathway." Int Immunol 12(11): 1539-1546. 21. Bennaceur, K., Popa, I., Portoukalian, J., Berthier-Vergnes, O. and Peguet-Navarro, J. (2006). "Melanoma-derived gangliosides impair migratory and antigen-presenting function of human epidermal Langerhans cells and induce their apoptosis." Int Immunol 18(6): 879-886. 22. Berenson, C. S. and Ryan, J. L. (1991). "Murine peritoneal macrophage gangliosides inhibit lymphocyte proliferation." J Leukoc Biol 50(4): 393-401. 23. Berzofsky, J. A., Terabe, M., Oh, S., Belyakov, I. M., Ahlers, J. D., Janik, J. E. and Morris, J. C. (2004). "Progress on new vaccine strategies for the immunotherapy and prevention of cancer." J Clin Invest 113(11): 1515-1525. 102 Bibliografía 24. Birkett, A., Lyons, K., Schmidt, A., Boyd, D., Oliveira, G. A., Siddique, A., Nussenzweig, R., CalvoCalle, J. M. and Nardin, E. (2002). "A modified hepatitis B virus core particle containing multiple epitopes of the Plasmodium falciparum circumsporozoite protein provides a highly immunogenic malaria vaccine in preclinical analyses in rodent and primate hosts." Infect Immun 70(12): 6860-6870. 25. Birkle, S., Zeng, G., Gao, L., Yu, R. K. and Aubry, J. (2003). "Role of tumor-associated gangliosides in cancer progression." Biochimie 85(3-4): 455-463. 26. Bjune, G., Hoiby, E. A., Gronnesby, J. K., Arnesen, O., Fredriksen, J. H., Halstensen, A., Holten, E., Lindbak, A. K., Nokleby, H., Rosenqvist, E. and et al. (1991). "Effect of outer membrane vesicle vaccine against group B meningococcal disease in Norway." Lancet 338(8775): 1093-1096. 27. Black, P. H. (1980). "Shedding from normal and cancer-cell surfaces." N Engl J Med 303(24): 1415-1416. 28. Blanco, R., Blanco, D., Quintana, Y., Escobar, X., Rengifo, C. E., Osorio, M., Gutierrez, Z., Lamadrid, J., Cedeno, M., Frometa, M., Carr, A. and Rengifo, E. (2013). "Immunoreactivity of the 14F7 Mab Raised against N-Glycolyl GM3 Ganglioside in Primary Lymphoid Tumors and Lymph Node Metastasis." Patholog Res Int 2013: 920972. 29. Blanco, R., Rengifo, C. E., Cedeno, M., Frometa, M., Rengifo, E. and Carr, A. (2012). "Immunoreactivity of the 14F7 Mab (Raised against N-Glycolyl GM3 Ganglioside) as a Positive Prognostic Factor in Non-Small-Cell Lung Cancer." Patholog Res Int 2012: 235418. 30. Blanco, R., Rengifo, E., Cedeno, M., Rengifo, C. E., Alonso, D. F. and Carr, A. (2011a). "Immunoreactivity of the 14F7 Mab Raised against N-Glycolyl GM3 Ganglioside in Epithelial Malignant Tumors from Digestive System." ISRN Gastroenterol 2011: 645641. 31. Blanco, R., Rengifo, E., Rengifo, C. E., Cedeno, M., Frometa, M. and Carr, A. (2011b). "Immunohistochemical Reactivity of the 14F7 Monoclonal Antibody Raised against N-Glycolyl GM3 Ganglioside in Some Benign and Malignant Skin Neoplasms." ISRN Dermatol 2011: 848909. 32. Brasel, K., De Smedt, T., Smith, J. L. and Maliszewski, C. R. (2000). "Generation of murine dendritic cells from flt3-ligand-supplemented bone marrow cultures." Blood 96(9): 3029-3039. 33. Brinkman-Van der Linden, E. C., Sjoberg, E. R., Juneja, L. R., Crocker, P. R., Varki, N. and Varki, A. (2000). "Loss of N-glycolylneuraminic acid in human evolution. Implications for sialic acid recognition by siglecs." J Biol Chem 275(12): 8633-8640. 34. Brunner, C., Seiderer, J., Schlamp, A., Bidlingmaier, M., Eigler, A., Haimerl, W., Lehr, H. A., Krieg, A. M., Hartmann, G. and Endres, S. (2000). "Enhanced dendritic cell maturation by TNF-alpha or cytidine-phosphate-guanosine DNA drives T cell activation in vitro and therapeutic anti-tumor immune responses in vivo." J Immunol 165(11): 6278-6286. 35. Burnet, F. M. (1970). "The concept of immunological surveillance." Prog Exp Tumor Res 13: 127. 103 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 36. Buskas, T., Thompson, P. and Boons, G. J. (2009). "Immunotherapy for cancer: synthetic carbohydrate-based vaccines." Chem Commun (Camb)(36): 5335-5349. 37. Cahan, L. D., Irie, R. F., Singh, R., Cassidenti, A. and Paulson, J. C. (1982). "Identification of a human neuroectodermal tumor antigen (OFA-I-2) as ganglioside GD2." Proc Natl Acad Sci U S A 79(24): 7629-7633. 38. Caldwell, S., Heitger, A., Shen, W., Liu, Y., Taylor, B. and Ladisch, S. (2003). "Mechanisms of ganglioside inhibition of APC function." J Immunol 171(4): 1676-1683. 39. Carnaud, C., Lee, D., Donnars, O., Park, S. H., Beavis, A., Koezuka, Y. and Bendelac, A. (1999). "Cutting edge: Cross-talk between cells of the innate immune system: NKT cells rapidly activate NK cells." J Immunol 163(9): 4647-4650. 40. Carr, A., Mazorra, Z., Alonso, D. F., Mesa, C., Valiente, O., Gomez, D. E., Perez, R. and Fernandez, L. E. (2001). "A purified GM3 ganglioside conjugated vaccine induces specific, adjuvantdependent and non-transient antitumour activity against B16 mouse melanoma in vitro and in vivo." Melanoma Res 11(3): 219-227. 41. Carr, A., Mullet, A., Mazorra, Z., Vazquez, A. M., Alfonso, M., Mesa, C., Rengifo, E., Perez, R. and Fernandez, L. E. (2000). "A mouse IgG1 monoclonal antibody specific for N-glycolyl GM3 ganglioside recognized breast and melanoma tumors." Hybridoma 19(3): 241-247. 42. Carr, A., Rodriguez, E., Arango Mdel, C., Camacho, R., Osorio, M., Gabri, M., Carrillo, G., Valdes, Z., Bebelagua, Y., Perez, R. and Fernandez, L. E. (2003). "Immunotherapy of advanced breast cancer with a heterophilic ganglioside (NeuGcGM3) cancer vaccine." J Clin Oncol 21(6): 10151021. 43. Casadesus, A. V., Fernandez-Marrero, Y., Clavell, M., Gomez, J. A., Hernandez, T., Moreno, E. and Lopez-Requena, A. (2013). "A shift from N-glycolyl- to N-acetyl-sialic acid in the GM3 ganglioside impairs tumor development in mouse lymphocytic leukemia cells." Glycoconj J 30(7): 687-699. 44. Casares, N., Pequignot, M. O., Tesniere, A., Ghiringhelli, F., Roux, S., Chaput, N., Schmitt, E., Hamai, A., Hervas-Stubbs, S., Obeid, M., Coutant, F., Metivier, D., Pichard, E., Aucouturier, P., Pierron, G., Garrido, C., Zitvogel, L. and Kroemer, G. (2005). "Caspase-dependent immunogenicity of doxorubicin-induced tumor cell death." J Exp Med 202(12): 1691-1701. 45. Colaco, C. A. (1998). "Towards a unified theory of immunity: dendritic cells, stress proteins and antigen capture." Cell Mol Biol (Noisy-le-grand) 44(6): 883-890. 46. Cottrez, F., Hurst, S. D., Coffman, R. L. and Groux, H. (2000). "T regulatory cells 1 inhibit a Th2specific response in vivo." J Immunol 165(9): 4848-4853. 47. Coulombe, J. and Pelletier, G. (1993). "Gangliosides and organ-specific metastatic colonization." Int J Cancer 53(1): 104-109. 104 Bibliografía 48. Chamilos, G., Lewis, R. E., Hu, J., Xiao, L., Zal, T., Gilliet, M., Halder, G. and Kontoyiannis, D. P. (2008). "Drosophila melanogaster as a model host to dissect the immunopathogenesis of zygomycosis." Proc Natl Acad Sci U S A 105(27): 9367-9372. 49. Chan, S. Y., Evan, G. I., Ritson, A., Watson, J., Wraight, P. and Sikora, K. (1986). "Localisation of lung cancer by a radiolabelled monoclonal antibody against the c-myc oncogene product." Br J Cancer 54(5): 761-769. 50. Chandrasekharan, K., Yoon, J. H., Xu, Y., deVries, S., Camboni, M., Janssen, P. M., Varki, A. and Martin, P. T. (2010). "A human-specific deletion in mouse Cmah increases disease severity in the mdx model of Duchenne muscular dystrophy." Sci Transl Med 2(42): 42ra54. 51. Chang, F., Li, R. and Ladisch, S. (1997). "Shedding of gangliosides by human medulloblastoma cells." Exp Cell Res 234(2): 341-346. 52. Chapman, P. B., Morrisey, D., Panageas, K. S., Williams, L., Lewis, J. J., Israel, R. J., Hamilton, W. B. and Livingston, P. O. (2000a). "Vaccination with a bivalent G(M2) and G(D2) ganglioside conjugate vaccine: a trial comparing doses of G(D2)-keyhole limpet hemocyanin." Clin Cancer Res 6(12): 4658-4662. 53. Chapman, P. B., Morrissey, D. M., Panageas, K. S., Hamilton, W. B., Zhan, C., Destro, A. N., Williams, L., Israel, R. J. and Livingston, P. O. (2000b). "Induction of antibodies against GM2 ganglioside by immunizing melanoma patients using GM2-keyhole limpet hemocyanin + QS21 vaccine: a dose-response study." Clin Cancer Res 6(3): 874-879. 54. Chapman, P. B., Wu, D., Ragupathi, G., Lu, S., Williams, L., Hwu, W. J., Johnson, D. and Livingston, P. O. (2004). "Sequential immunization of melanoma patients with GD3 ganglioside vaccine and anti-idiotypic monoclonal antibody that mimics GD3 ganglioside." Clin Cancer Res 10(14): 4717-4723. 55. Chaput, N., Flament, C., Viaud, S., Taieb, J., Roux, S., Spatz, A., Andre, F., LePecq, J. B., Boussac, M., Garin, J., Amigorena, S., Thery, C. and Zitvogel, L. (2006). "Dendritic cell derived-exosomes: biology and clinical implementations." J Leukoc Biol 80(3): 471-478. 56. Cheresh, D. A. (1985). "Structural and functional properties of ganglioside antigens on human tumors of neuroectodermal origin." Surv Synth Pathol Res 4(2): 97-109. 57. Chou, H. H., Hayakawa, T., Diaz, S., Krings, M., Indriati, E., Leakey, M., Paabo, S., Satta, Y., Takahata, N. and Varki, A. (2002). "Inactivation of CMP-N-acetylneuraminic acid hydroxylase occurred prior to brain expansion during human evolution." Proc Natl Acad Sci U S A 99(18): 11736-11741. 58. Chu, J. W. and Sharom, F. J. (1995). "Gangliosides interact with interleukin-4 and inhibit interleukin-4-stimulated helper T-cell proliferation." Immunology 84(3): 396-403. 59. Chung, M. H., Gupta, R. K., Hsueh, E., Essner, R., Ye, W., Yee, R. and Morton, D. L. (2003). "Humoral immune response to a therapeutic polyvalent cancer vaccine after complete resection of thick primary melanoma and sentinel lymphadenectomy." J Clin Oncol 21(2): 313319. 105 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 60. Chung, T. W., Kim, S. J., Choi, H. J., Kim, K. J., Kim, M. J., Kim, S. H., Lee, H. J., Ko, J. H., Lee, Y. C., Suzuki, A. and Kim, C. H. (2009). "Ganglioside GM3 inhibits VEGF/VEGFR-2-mediated angiogenesis: direct interaction of GM3 with VEGFR-2." Glycobiology 19(3): 229-239. 61. Daniotti, J. L. and Iglesias-Bartolome, R. (2011). "Metabolic pathways and intracellular trafficking of gangliosides." IUBMB Life 63(7): 513-520. 62. de Leon, J., Fernandez, A., Clavell, M., Labrada, M., Bebelagua, Y., Mesa, C. and Fernandez, L. E. (2008). "Differential influence of the tumour-specific non-human sialic acid containing GM3 ganglioside on CD4+CD25- effector and naturally occurring CD4+CD25+ regulatory T cells function." Int Immunol 20(4): 591-600. 63. de Leon, J., Fernandez, A., Mesa, C., Clavel, M. and Fernandez, L. E. (2006). "Role of tumourassociated N-glycolylated variant of GM3 ganglioside in cancer progression: effect over CD4 expression on T cells." Cancer Immunol Immunother 55(4): 443-450. 64. de Mello Coelho, V., Nguyen, D., Giri, B., Bunbury, A., Schaffer, E. and Taub, D. D. (2004). "Quantitative differences in lipid raft components between murine CD4+ and CD8+ T cells." BMC Immunol 5: 2. 65. Diaz, A., Alfonso, M., Alonso, R., Saurez, G., Troche, M., Catala, M., Diaz, R. M., Perez, R. and Vazquez, A. M. (2003). "Immune responses in breast cancer patients immunized with an antiidiotype antibody mimicking NeuGc-containing gangliosides." Clin Immunol 107(2): 80-89. 66. Diaz, S. L., Padler-Karavani, V., Ghaderi, D., Hurtado-Ziola, N., Yu, H., Chen, X., Brinkman-Van der Linden, E. C., Varki, A. and Varki, N. M. (2009). "Sensitive and specific detection of the nonhuman sialic Acid N-glycolylneuraminic acid in human tissues and biotherapeutic products." PLoS One 4(1): e4241. 67. Dickler, M. N., Ragupathi, G., Liu, N. X., Musselli, C., Martino, D. J., Miller, V. A., Kris, M. G., Brezicka, F. T., Livingston, P. O. and Grant, S. C. (1999). "Immunogenicity of a fucosyl-GM1keyhole limpet hemocyanin conjugate vaccine in patients with small cell lung cancer." Clin Cancer Res 5(10): 2773-2779. 68. Dixon, G. L., Newton, P. J., Chain, B. M., Katz, D., Andersen, S. R., Wong, S., van der Ley, P., Klein, N. and Callard, R. E. (2001). "Dendritic cell activation and cytokine production induced by group B Neisseria meningitidis: interleukin-12 production depends on lipopolysaccharide expression in intact bacteria." Infect Immun 69(7): 4351-4357. 69. Dolo, V., Li, R., Dillinger, M., Flati, S., Manela, J., Taylor, B. J., Pavan, A. and Ladisch, S. (2000). "Enrichment and localization of ganglioside G(D3) and caveolin-1 in shed tumor cell membrane vesicles." Biochim Biophys Acta 1486(2-3): 265-274. 70. Dubois, C., Magnani, J. L., Grunwald, G. B., Spitalnik, S. L., Trisler, G. D., Nirenberg, M. and Ginsburg, V. (1986). "Monoclonal antibody 18B8, which detects synapse-associated antigens, binds to ganglioside GT3 (II3 (NeuAc)3LacCer)." J Biol Chem 261(8): 3826-3830. 106 Bibliografía 71. Duclos, R. I., Jr. (2000). "The total synthesis of ganglioside GM3." Carbohydr Res 328(4): 489507. 72. Duda, R. B., Yang, H., Dooley, D. D. and Abu-Jawdeh, G. (1995). "Recombinant BCG therapy suppresses melanoma tumor growth." Ann Surg Oncol 2(6): 542-549. 73. Dumontet, C., Rebbaa, A. and Portoukalian, J. (1997). "Very low density lipoproteins and interleukin 2 enhance the immunogenicity of 9-O-acetyl-GD3 ganglioside in BALB/c mice." J Immunol Methods 206(1-2): 115-123. 74. Durand, V., Mackenzie, J., de Leon, J., Mesa, C., Quesniaux, V., Montoya, M., Le Bon, A. and Wong, S. Y. (2009). "Role of lipopolysaccharide in the induction of type I interferon-dependent cross-priming and IL-10 production in mice by meningococcal outer membrane vesicles." Vaccine 27(13): 1912-1922. 75. Durrant, L. G., Noble, P. and Spendlove, I. (2012). "Immunology in the clinic review series; focus on cancer: glycolipids as targets for tumour immunotherapy." Clin Exp Immunol 167(2): 206215. 76. Dutoit, V., Taub, R. N., Papadopoulos, K. P., Talbot, S., Keohan, M. L., Brehm, M., Gnjatic, S., Harris, P. E., Bisikirska, B., Guillaume, P., Cerottini, J. C., Hesdorffer, C. S., Old, L. J. and Valmori, D. (2002). "Multiepitope CD8(+) T cell response to a NY-ESO-1 peptide vaccine results in imprecise tumor targeting." J Clin Invest 110(12): 1813-1822. 77. Dyatlovitskaya, E. V. and Bergelson, L. D. (1987). "Glycosphingolipids and antitumor immunity." Biochim Biophys Acta 907(2): 125-143. 78. Eggermont, A. M., Suciu, S., Rutkowski, P., Marsden, J., Santinami, M., Corrie, P., Aamdal, S., Ascierto, P. A., Patel, P. M., Kruit, W. H., Bastholt, L., Borgognoni, L., Bernengo, M. G., Davidson, N., Polders, L., Praet, M. and Spatz, A. (2013). "Adjuvant ganglioside GM2-KLH/QS-21 vaccination versus observation after resection of primary tumor > 1.5 mm in patients with stage II melanoma: results of the EORTC 18961 randomized phase III trial." J Clin Oncol 31(30): 38313837. 79. Eisenbach, L., Hollander, N., Greenfeld, L., Yakor, H., Segal, S. and Feldman, M. (1984). "The differential expression of H-2K versus H-2D antigens, distinguishing high-metastatic from lowmetastatic clones, is correlated with the immunogenic properties of the tumor cells." Int J Cancer 34(4): 567-573. 80. Eisenbach, L., Segal, S. and Feldman, M. (1983). "MHC imbalance and metastatic spread in Lewis lung carcinoma clones." Int J Cancer 32(1): 113-120. 81. El-Abbadi, M., Seyfried, T. N., Yates, A. J., Orosz, C. and Lee, M. C. (2001). "Ganglioside composition and histology of a spontaneous metastatic brain tumour in the VM mouse." Br J Cancer 85(2): 285-292. 107 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 82. el-Shami, K., Tirosh, B., Bar-Haim, E., Carmon, L., Vadai, E., Fridkin, M., Feldman, M. and Eisenbach, L. (1999). "MHC class I-restricted epitope spreading in the context of tumor rejection following vaccination with a single immunodominant CTL epitope." Eur J Immunol 29(10): 32953301. 83. Estevez, F., Carr, A., Solorzano, L., Valiente, O., Mesa, C., Barroso, O., Sierra, G. V. and Fernandez, L. E. (1999). "Enhancement of the immune response to poorly immunogenic gangliosides after incorporation into very small size proteoliposomes (VSSP)." Vaccine 18(1-2): 190-197. 84. Feeney, R. E., Rhodes, M. B. and Anderson, J. S. (1960). "The distribution and role of sialic acid in chicken egg white." J Biol Chem 235: 2633-2637. 85. Fernandez-Marrero, Y., Roque-Navarro, L., Hernandez, T., Dorvignit, D., Molina-Perez, M., Gonzalez, A., Sosa, K., Lopez-Requena, A., Perez, R. and de Acosta, C. M. (2011). "A cytotoxic humanized anti-ganglioside antibody produced in a murine cell line defective of N-glycolylatedglycoconjugates." Immunobiology 216(12): 1239-1247. 86. Fernandez, A., Mesa, C., Marigo, I., Dolcetti, L., Clavell, M., Oliver, L., Fernandez, L. E. and Bronte, V. (2011). "Inhibition of tumor-induced myeloid-derived suppressor cell function by a nanoparticulated adjuvant." J Immunol 186(1): 264-274. 87. Fernández, A., Oliver, L., Alvarez, R., Fernández, L. E. and Mesa, C. (2014a). "GM3-containing nanoparticles in immunosuppressed hosts: Effect on myeloid-derived suppressor cells." World J Immunol 4(2): 98-106. 88. Fernández, A., Oliver, L., Alvarez, R., Hernández, A., Raymond, J., Fernández, L. E. and Mesa, C. (2014b). "Very small size proteoliposomes abrogate cross-presentation of tumor antigens by myeloidderived suppressor cells and induce their differentiation to dendritic cells." J Immunother Cancer 2: 5. 89. Fernandez, L. E., Gabri, M. R., Guthmann, M. D., Gomez, R. E., Gold, S., Fainboim, L., Gomez, D. E. and Alonso, D. F. (2010). "NGcGM3 ganglioside: a privileged target for cancer vaccines." Clin Dev Immunol 2010: 814397. 90. Fishman, P. H. (1986). "Recent advances in identifying the functions of gangliosides." Chem Phys Lipids 42(1-3): 137-151. 91. Fridman, W. H., Pages, F., Sautes-Fridman, C. and Galon, J. (2012). "The immune contexture in human tumours: impact on clinical outcome." Nat Rev Cancer 12(4): 298-306. 92. Frugoni, P., Secchi, E. F. and Chizzolini, C. (1994). "Effect of pure bovine brain-derived gangliosides on normal human B cell proliferation in vitro." J Neuroimmunol 49(1-2): 189-195. 93. Fuentes, D., Avellanet, J., Garcia, A., Iglesias, N., Gabri, M. R., Alonso, D. F., Vazquez, A. M., Perez, R. and Montero, E. (2010). "Combined therapeutic effect of a monoclonal anti-idiotype tumor vaccine against NeuGc-containing gangliosides with chemotherapy in a breast carcinoma model." Breast Cancer Res Treat 120(2): 379-389. 108 Bibliografía 94. Fuentes, D., Chacon, L., Casaco, A., Ledon, N., Fernandez, N., Iglesias, A., Hernandez, D. R., Sanchez, B. and Perez, R. (2012). "Effects of an epidermal growth factor receptor-based cancer vaccine in the wound healing and inflammation processes in murine experimental models." Int Wound J. 95. Gabri, M. R., Otero, L. L., Gomez, D. E. and Alonso, D. F. (2009). "Exogenous incorporation of neugc-rich mucin augments n-glycolyl sialic acid content and promotes malignant phenotype in mouse tumor cell lines." J Exp Clin Cancer Res 28: 146. 96. Galon, J., Costes, A., Sanchez-Cabo, F., Kirilovsky, A., Mlecnik, B., Lagorce-Pages, C., Tosolini, M., Camus, M., Berger, A., Wind, P., Zinzindohoue, F., Bruneval, P., Cugnenc, P. H., Trajanoski, Z., Fridman, W. H. and Pages, F. (2006). "Type, density, and location of immune cells within human colorectal tumors predict clinical outcome." Science 313(5795): 1960-1964. 97. Galon, J., Fridman, W. H. and Pages, F. (2007). "The adaptive immunologic microenvironment in colorectal cancer: a novel perspective." Cancer Res 67(5): 1883-1886. 98. Giaccone, G., Debruyne, C., Felip, E., Chapman, P. B., Grant, S. C., Millward, M., Thiberville, L., D'Addario, G., Coens, C., Rome, L. S., Zatloukal, P., Masso, O. and Legrand, C. (2005). "Phase III study of adjuvant vaccination with Bec2/bacille Calmette-Guerin in responding patients with limited-disease small-cell lung cancer (European Organisation for Research and Treatment of Cancer 08971-08971B; Silva Study)." J Clin Oncol 23(28): 6854-6864. 99. Gilliet, M., Cao, W. and Liu, Y. J. (2008a). "Plasmacytoid dendritic cells: sensing nucleic acids in viral infection and autoimmune diseases." Nat Rev Immunol 8(8): 594-606. 100. Gilliet, M. and Lande, R. (2008b). "Antimicrobial peptides and self-DNA in autoimmune skin inflammation." Curr Opin Immunol 20(4): 401-407. 101. Gogas, H., Ioannovich, J., Dafni, U., Stavropoulou-Giokas, C., Frangia, K., Tsoutsos, D., Panagiotou, P., Polyzos, A., Papadopoulos, O., Stratigos, A., Markopoulos, C., Bafaloukos, D., Pectasides, D., Fountzilas, G. and Kirkwood, J. M. (2006). "Prognostic significance of autoimmunity during treatment of melanoma with interferon." N Engl J Med 354(7): 709-718. 102. Golde, W. T., Gollobin, P. and Rodriguez, L. L. (2005). "A rapid, simple, and humane method for submandibular bleeding of mice using a lancet." Lab Anim (NY) 34(9): 39-43. 103. Gorelik, E., Segal, S. and Feldman, M. (1980). "Control of lung metastasis progression in mice: role of growth kinetics of 3LL Lewis lung carcinoma and host immune reactivity." J Natl Cancer Inst 65(6): 1257-1264. 104. Grayson, G. and Ladisch, S. (1992). "Immunosuppression by human gangliosides. II. Carbohydrate structure and inhibition of human NK activity." Cell Immunol 139(1): 18-29. 105. Guan, F., Handa, K. and Hakomori, S. I. (2011). "Regulation of epidermal growth factor receptor through interaction of ganglioside GM3 with GlcNAc of N-linked glycan of the receptor: demonstration in ldlD cells." Neurochem Res 36(9): 1645-1653. 109 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 106. Guthmann, M. D., Bitton, R. J., Carnero, A. J., Gabri, M. R., Cinat, G., Koliren, L., Lewi, D., Fernandez, L. E., Alonso, D. F., Gomez, D. E. and Fainboim, L. (2004). "Active specific immunotherapy of melanoma with a GM3 ganglioside-based vaccine: a report on safety and immunogenicity." J Immunother 27(6): 442-451. 107. Hailemichael, Y., Dai, Z., Jaffarzad, N., Ye, Y., Medina, M. A., Huang, X. F., Dorta-Estremera, S. M., Greeley, N. R., Nitti, G., Peng, W., Liu, C., Lou, Y., Wang, Z., Ma, W., Rabinovich, B., Sowell, R. T., Schluns, K. S., Davis, R. E., Hwu, P. and Overwijk, W. W. (2013). "Persistent antigen at vaccination sites induces tumor-specific CD8(+) T cell sequestration, dysfunction and deletion." Nat Med 19(4): 465-472. 108. Hakomori, S. (1985). "Aberrant glycosylation in cancer cell membranes as focused on glycolipids: overview and perspectives." Cancer Res 45(6): 2405-2414. 109. Hakomori, S. (1996). "Tumor malignancy defined by aberrant sphingo(glyco)lipid metabolism." Cancer Res 56(23): 5309-5318. glycosylation and 110. Hamilton, W. B., Helling, F., Lloyd, K. O. and Livingston, P. O. (1993). "Ganglioside expression on human malignant melanoma assessed by quantitative immune thin-layer chromatography." Int J Cancer 53(4): 566-573. 111. Hayashi, N., Chiba, H., Kuronuma, K., Go, S., Hasegawa, Y., Takahashi, M., Gasa, S., Watanabe, A., Hasegawa, T., Kuroki, Y., Inokuchi, J. and Takahashi, H. (2013). "Detection of N-glycolyated gangliosides in non-small-cell lung cancer using GMR8 monoclonal antibody." Cancer Sci 104(1): 43-47. 112. Heath, W. R. and Carbone, F. R. (1999). "Cytotoxic T lymphocyte activation by cross-priming." Curr Opin Immunol 11(3): 314-318. 113. Hedlund, M., Tangvoranuntakul, P., Takematsu, H., Long, J. M., Housley, G. D., Kozutsumi, Y., Suzuki, A., Wynshaw-Boris, A., Ryan, A. F., Gallo, R. L., Varki, N. and Varki, A. (2007). "Nglycolylneuraminic acid deficiency in mice: implications for human biology and evolution." Mol Cell Biol 27(12): 4340-4346. 114. Helling, F. and Livingston, P. O. (1994). "Ganglioside conjugate vaccines. Immunotherapy against tumors of neuroectodermal origin." Mol Chem Neuropathol 21(2-3): 299-309. 115. Helling, F., Zhang, S., Shang, A., Adluri, S., Calves, M., Koganty, R., Longenecker, B. M., Yao, T. J., Oettgen, H. F. and Livingston, P. O. (1995). "GM2-KLH conjugate vaccine: increased immunogenicity in melanoma patients after administration with immunological adjuvant QS21." Cancer Res 55(13): 2783-2788. 116. Hernandez, A. M., Rodriguez, M., Lopez-Requena, A., Beausoleil, I., Perez, R. and Vazquez, A. M. (2005). "Generation of anti-Neu-glycolyl-ganglioside antibodies by immunization with an antiidiotype monoclonal antibody: A self versus non-self-matter." Immunobiology 210(1): 11-21. 110 Bibliografía 117. Hernandez, A. M., Rodriguez, N., Gonzalez, J. E., Reyes, E., Rondon, T., Grinan, T., Macias, A., Alfonso, S., Vazquez, A. M. and Perez, R. (2011). "Anti-NeuGcGM3 antibodies, actively elicited by idiotypic vaccination in nonsmall cell lung cancer patients, induce tumor cell death by an oncosis-like mechanism." J Immunol 186(6): 3735-3744. 118. Hernandez, A. M., Toledo, D., Martinez, D., Grinan, T., Brito, V., Macias, A., Alfonso, S., Rondon, T., Suarez, E., Vazquez, A. M. and Perez, R. (2008). "Characterization of the antibody response against NeuGcGM3 ganglioside elicited in non-small cell lung cancer patients immunized with an anti-idiotype antibody." J Immunol 181(9): 6625-6634. 119. Higashi, H., Hirabayashi, Y., Hirota, M., Matsumoto, M. and Kato, S. (1987). "Detection of ganglioside GM2 in sera and tumor tissues of hepatoma patients." Jpn J Cancer Res 78(12): 1309-1313. 120. Higashi, H., Sasabe, T., Fukui, Y., Maru, M. and Kato, S. (1988). "Detection of gangliosides as Nglycolylneuraminic acid-specific tumor-associated Hanganutziu-Deicher antigen in human retinoblastoma cells." Jpn J Cancer Res 79(8): 952-956. 121. Hirabayashi, Y., Higashi, H., Kato, S., Taniguchi, M. and Matsumoto, M. (1987). "Occurrence of tumor-associated ganglioside antigens with Hanganutziu-Deicher antigenic activity on human melanomas." Jpn J Cancer Res 78(6): 614-620. 122. Hirabayashi, Y., Higashi, H. and Taniguchi, M. (1986). "[Biochemistry of melanoma-associated ganglioside antigens]." Gan To Kagaku Ryoho 13(3 Pt 2): 834-840. 123. Holmgren, J., Lonnroth, I. and Svennerholm, L. (1973). "Tissue receptor for cholera exotoxin: postulated structure from studies with GM1 ganglioside and related glycolipids." Infect Immun 8(2): 208-214. 124. Hoos, A. and Levey, D. L. (2003). "Vaccination with heat shock protein-peptide complexes: from basic science to clinical applications." Expert Rev Vaccines 2(3): 369-379. 125. Hsueh, E. C., Famatiga, E., Gupta, R. K., Qi, K. and Morton, D. L. (1998). "Enhancement of complement-dependent cytotoxicity by polyvalent melanoma cell vaccine (CancerVax): correlation with survival." Ann Surg Oncol 5(7): 595-602. 126. Huang, X., Li, Y., Zhang, J., Xu, Y., Tian, Y. and Ma, K. (2013). "Ganglioside GM3 inhibits hepatoma cell motility via down-regulating activity of EGFR and PI3K/AKT signaling pathway." J Cell Biochem 114(7): 1616-1624. 127. Inaba, K., Inaba, M., Romani, N., Aya, H., Deguchi, M., Ikehara, S., Muramatsu, S. and Steinman, R. M. (1992). "Generation of large numbers of dendritic cells from mouse bone marrow cultures supplemented with granulocyte/macrophage colony-stimulating factor." J Exp Med 176(6): 1693-1702. 128. Irani, D. N., Lin, K. I. and Griffin, D. E. (1996). "Brain-derived gangliosides regulate the cytokine production and proliferation of activated T cells." J Immunol 157(10): 4333-4340. 111 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 129. Itai, S., Nishikata, J., Yoneda, T., Ohmori, K., Yamabe, H., Arii, S., Tobe, T. and Kannagi, R. (1991). "Tissue distribution of 2-3 and 2-6 sialyl Lewis A antigens and significance of the ratio of two antigens for the differential diagnosis of malignant and benign disorders of the digestive tract." Cancer 67(6): 1576-1587. 130. Jager, D., Jager, E. and Knuth, A. (2001). "Vaccination for malignant melanoma: recent developments." Oncology 60(1): 1-7. 131. Jager, E., Jager, D. and Knuth, A. (1999). "CTL-defined cancer vaccines: perspectives for active immunotherapeutic interventions in minimal residual disease." Cancer Metastasis Rev 18(1): 143-150. 132. Jakob, T., Walker, P. S., Krieg, A. M., Udey, M. C. and Vogel, J. C. (1998). "Activation of cutaneous dendritic cells by CpG-containing oligodeoxynucleotides: a role for dendritic cells in the augmentation of Th1 responses by immunostimulatory DNA." J Immunol 161(6): 30423049. 133. Kannagi, R. (2004). "Molecular mechanism for cancer-associated induction of sialyl Lewis X and sialyl Lewis A expression-The Warburg effect revisited." Glycoconj J 20(5): 353-364. 134. Kannagi, R., Yin, J., Miyazaki, K. and Izawa, M. (2008). "Current relevance of incomplete synthesis and neo-synthesis for cancer-associated alteration of carbohydrate determinants-Hakomori's concepts revisited." Biochim Biophys Acta 1780(3): 525-531. 135. Kaplan, D. H., Shankaran, V., Dighe, A. S., Stockert, E., Aguet, M., Old, L. J. and Schreiber, R. D. (1998). "Demonstration of an interferon gamma-dependent tumor surveillance system in immunocompetent mice." Proc Natl Acad Sci U S A 95(13): 7556-7561. 136. Karlsson, K. A. (1989). "Animal glycosphingolipids as membrane attachment sites for bacteria." Annu Rev Biochem 58: 309-350. 137. Kavanagh, D., Hill, A. D., Djikstra, B., Kennelly, R., McDermott, E. M. and O'Higgins, N. J. (2005). "Adjuvant therapies in the treatment of stage II and III malignant melanoma." Surgeon 3(4): 245-256. 138. Kawai, T., Kato, A., Higashi, H., Kato, S. and Naiki, M. (1991). "Quantitative determination of Nglycolylneuraminic acid expression in human cancerous tissues and avian lymphoma cell lines as a tumor-associated sialic acid by gas chromatography-mass spectrometry." Cancer Res 51(4): 1242-1246. 139. Kawamura, Y. I., Toyota, M., Kawashima, R., Hagiwara, T., Suzuki, H., Imai, K., Shinomura, Y., Tokino, T., Kannagi, R. and Dohi, T. (2008). "DNA hypermethylation contributes to incomplete synthesis of carbohydrate determinants in gastrointestinal cancer." Gastroenterology 135(1): 142-151 e143. 140. Kawashima, I., Ozawa, H., Kotani, M., Suzuki, M., Kawano, T., Gomibuchi, M. and Tai, T. (1993). "Characterization of ganglioside expression in human melanoma cells: immunological and biochemical analysis." J Biochem 114(2): 186-193. 112 Bibliografía 141. Kawashima, I., Yoshida, Y., Taya, C., Shitara, H., Yonekawa, H., Karasuyama, H., Tada, N., Furukawa, K. and Tai, T. (2003). "Expansion of natural killer cells in mice transgenic for IgM antibody to ganglioside GD2: demonstration of prolonged survival after challenge with syngeneic tumor cells." Int J Oncol 23(2): 381-388. 142. Kearney, J. F., Radbruch, A., Liesegang, B. and Rajewsky, K. (1979). "A new mouse myeloma cell line that has lost immunoglobulin expression but permits the construction of antibodysecreting hybrid cell lines." J Immunol 123(4): 1548-1550. 143. Kelland, L. (2006). "Discontinued drugs in 2005: oncology drugs." Expert Opin Investig Drugs 15(11): 1309-1318. 144. Kikuta, K. (1985). "[Sialic acid in human whole saliva. Reliability of assay methods and correlation with periodontal disease]." Nichidai Koko Kagaku 11(1): 27-34. 145. Kim, S. J., Chung, T. W., Choi, H. J., Jin, U. H., Ha, K. T., Lee, Y. C. and Kim, C. H. (2014). "Monosialic ganglioside GM3 specifically suppresses the monocyte adhesion to endothelial cells for inflammation." Int J Biochem Cell Biol 46: 32-38. 146. Kimata, H. and Yoshida, A. (1996). "Inhibition of spontaneous immunoglobulin production by ganglioside GM2 in human B cells." Clin Immunol Immunopathol 79(2): 197-202. 147. Kitamura, K., Livingston, P. O., Fortunato, S. R., Stockert, E., Helling, F., Ritter, G., Oettgen, H. F. and Old, L. J. (1995). "Serological response patterns of melanoma patients immunized with a GM2 ganglioside conjugate vaccine." Proc Natl Acad Sci U S A 92(7): 2805-2809. 148. Kmieciak, M., Basu, D., Payne, K. K., Toor, A., Yacoub, A., Wang, X. Y., Smith, L., Bear, H. D. and Manjili, M. H. (2011). "Activated NKT cells and NK cells render T cells resistant to myeloidderived suppressor cells and result in an effective adoptive cellular therapy against breast cancer in the FVBN202 transgenic mouse." J Immunol 187(2): 708-717. 149. Ko, E. C., Wang, X. and Ferrone, S. (2003). "Immunotherapy of malignant diseases. Challenges and strategies." Int Arch Allergy Immunol 132(4): 294-309. 150. Koike, T., Kimura, N., Miyazaki, K., Yabuta, T., Kumamoto, K., Takenoshita, S., Chen, J., Kobayashi, M., Hosokawa, M., Taniguchi, A., Kojima, T., Ishida, N., Kawakita, M., Yamamoto, H., Takematsu, H., Suzuki, A., Kozutsumi, Y. and Kannagi, R. (2004). "Hypoxia induces adhesion molecules on cancer cells: A missing link between Warburg effect and induction of selectinligand carbohydrates." Proc Natl Acad Sci U S A 101(21): 8132-8137. 151. Kong, Y., Li, R. and Ladisch, S. (1998). "Natural forms of shed tumor gangliosides." Biochim Biophys Acta 1394(1): 43-56. 152. Kono, K., Tsuchida, T., Kern, D. H. and Irie, R. (1990). "Ganglioside composition of human melanoma and response to antitumor treatment." Cancer Invest 8(2): 161-167. 153. Kroemer, G., Galluzzi, L., Kepp, O. and Zitvogel, L. (2013). "Immunogenic cell death in cancer therapy." Annu Rev Immunol 31: 51-72. 113 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 154. Krug, L. M., Ragupathi, G., Hood, C., Kris, M. G., Miller, V. A., Allen, J. R., Keding, S. J., Danishefsky, S. J., Gomez, J., Tyson, L., Pizzo, B., Baez, V. and Livingston, P. O. (2004). "Vaccination of patients with small-cell lung cancer with synthetic fucosyl GM-1 conjugated to keyhole limpet hemocyanin." Clin Cancer Res 10(18 Pt 1): 6094-6100. 155. Kubler, K., tho Pesch, C., Gehrke, N., Riemann, S., Dassler, J., Coch, C., Landsberg, J., Wimmenauer, V., Polcher, M., Rudlowski, C., Tuting, T., Kuhn, W., Hartmann, G. and Barchet, W. (2011). "Immunogenic cell death of human ovarian cancer cells induced by cytosolic poly(I:C) leads to myeloid cell maturation and activates NK cells." Eur J Immunol 41(10): 30283039. 156. Ladisch, S., Becker, H. and Ulsh, L. (1992). "Immunosuppression by human gangliosides: I. Relationship of carbohydrate structure to the inhibition of T cell responses." Biochim Biophys Acta 1125(2): 180-188. 157. Ladisch, S., Ulsh, L., Gillard, B. and Wong, C. (1984). "Modulation of the immune response by gangliosides. Inhibition of adherent monocyte accessory function in vitro." J Clin Invest 74(6): 2074-2081. 158. Lage, A., Perez, R. and Fernandez, L. E. (2005). "Therapeutic cancer vaccines: at midway between immunology and pharmacology." Curr Cancer Drug Targets 5(8): 611-627. 159. Lee, S. C., Srivastava, R. M., Lopez-Albaitero, A., Ferrone, S. and Ferris, R. L. (2011). "Natural killer (NK): dendritic cell (DC) cross talk induced by therapeutic monoclonal antibody triggers tumor antigen-specific T cell immunity." Immunol Res 50(2-3): 248-254. 160. Li, F. and Pestronk, A. (1991). "Autoantibodies to GM1 ganglioside: different reactivity to GM1liposomes in amyotrophic lateral sclerosis and lower motor neuron disorders." J Neurol Sci 104(2): 209-214. 161. Li, R. and Ladisch, S. (1997). "Inhibition of endogenous ganglioside synthesis does not block neurite formation by retinoic acid-treated neuroblastoma cells." J Biol Chem 272(2): 1349-1354. 162. Lin, Y., Roberts, T. J., Spence, P. M. and Brutkiewicz, R. R. (2005). "Reduction in CD1d expression on dendritic cells and macrophages by an acute virus infection." J Leukoc Biol 77(2): 151-158. 163. Liu, Y., Su, Y., Wiznitzer, M., Epifano, O. and Ladisch, S. (2008). "Ganglioside depletion and EGF responses of human GM3 synthase-deficient fibroblasts." Glycobiology 18(8): 593-601. 164. Livingston, P. O. (1995a). "Approaches to augmenting the immunogenicity of melanoma gangliosides: from whole melanoma cells to ganglioside-KLH conjugate vaccines." Immunol Rev 145: 147-166. 165. Livingston, P. O. (1995b). "Augmenting the immunogenicity of carbohydrate tumor antigens." Semin Cancer Biol 6(6): 357-366. 114 Bibliografía 166. Livingston, P. O., Adluri, S., Helling, F., Yao, T. J., Kensil, C. R., Newman, M. J. and Marciani, D. (1994a). "Phase 1 trial of immunological adjuvant QS-21 with a GM2 ganglioside-keyhole limpet haemocyanin conjugate vaccine in patients with malignant melanoma." Vaccine 12(14): 12751280. 167. Livingston, P. O., Calves, M. J., Helling, F., Zollinger, W. D., Blake, M. S. and Lowell, G. H. (1993). "GD3/proteosome vaccines induce consistent IgM antibodies against the ganglioside GD3." Vaccine 11(12): 1199-1204. 168. Livingston, P. O., Calves, M. J. and Natoli, E. J., Jr. (1987). "Approaches to augmenting the immunogenicity of the ganglioside GM2 in mice: purified GM2 is superior to whole cells." J Immunol 138(5): 1524-1529. 169. Livingston, P. O. and Ragupathi, G. (1997a). "Carbohydrate vaccines that induce antibodies against cancer. 2. Previous experience and future plans." Cancer Immunol Immunother 45(1): 10-19. 170. Livingston, P. O., Ritter, G. and Calves, M. J. (1989a). "Antibody response after immunization with the gangliosides GM1, GM2, GM3, GD2 and GD3 in the mouse." Cancer Immunol Immunother 29(3): 179-184. 171. Livingston, P. O., Ritter, G., Srivastava, P., Padavan, M., Calves, M. J., Oettgen, H. F. and Old, L. J. (1989b). "Characterization of IgG and IgM antibodies induced in melanoma patients by immunization with purified GM2 ganglioside." Cancer Res 49(24 Pt 1): 7045-7050. 172. Livingston, P. O., Watanabe, T., Shiku, H., Houghton, A. N., Albino, A., Takahashi, T., Resnick, L. A., Michitsch, R., Pinsky, C. M., Oettgen, H. F. and Old, L. J. (1982). "Serological response of melanoma patients receiving melanoma cell vaccines. I. Autologous cultured melanoma cells." Int J Cancer 30(4): 413-422. 173. Livingston, P. O., Wong, G. Y., Adluri, S., Tao, Y., Padavan, M., Parente, R., Hanlon, C., Calves, M. J., Helling, F., Ritter, G. and et al. (1994b). "Improved survival in stage III melanoma patients with GM2 antibodies: a randomized trial of adjuvant vaccination with GM2 ganglioside." J Clin Oncol 12(5): 1036-1044. 174. Livingston, P. O., Zhang, S. and Lloyd, K. O. (1997b). "Carbohydrate vaccines that induce antibodies against cancer. 1. Rationale." Cancer Immunol Immunother 45(1): 1-9. 175. MacDonald, H. R. (2002). "Development and selection of NKT cells." Curr Opin Immunol 14(2): 250-254. 176. Makatsori, E., Fermani, K., Aletras, A., Karamanos, N. K. and Tsegenidis, T. (1998). "Screening of N-acylneuraminic acids in serum and tissue specimens of mouse C57BI with Lewis' lung cancer by high-performance liquid chromatography." J Chromatogr B Biomed Sci Appl 712(1-2): 23-29. 177. Malykh, Y. N., Schauer, R. and Shaw, L. (2001). "N-Glycolylneuraminic acid in human tumours." Biochimie 83(7): 623-634. 115 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 178. Marcus, D. M. (1984). "A review of the immunogenic and immuno-modulatory properties of glycosphingolipids." Mol Immunol 21(11): 1083-1091. 179. Martin-Orozco, E., Kobayashi, H., Van Uden, J., Nguyen, M. D., Kornbluth, R. S. and Raz, E. (1999). "Enhancement of antigen-presenting cell surface molecules involved in cognate interactions by immunostimulatory DNA sequences." Int Immunol 11(7): 1111-1118. 180. Matzinger, P. (1994). "Tolerance, danger, and the extended family." Annu Rev Immunol 12: 991-1045. 181. Mazorra, Z., Mesa, C., Fernandez, A. and Fernandez, L. E. (2008). "Immunization with a GM3 ganglioside nanoparticulated vaccine confers an effector CD8(+) T cells-mediated protection against melanoma B16 challenge." Cancer Immunol Immunother 57(12): 1771-1780. 182. Mazzaferro, V., Coppa, J., Carrabba, M. G., Rivoltini, L., Schiavo, M., Regalia, E., Mariani, L., Camerini, T., Marchiano, A., Andreola, S., Camerini, R., Corsi, M., Lewis, J. J., Srivastava, P. K. and Parmiani, G. (2003). "Vaccination with autologous tumor-derived heat-shock protein gp96 after liver resection for metastatic colorectal cancer." Clin Cancer Res 9(9): 3235-3245. 183. Mesa, C., de Leon, J. and Fernandez, L. E. (2006a). "Very small size proteoliposomes derived from Neisseria meningitidis: An effective adjuvant for generation of CTL responses to peptide and protein antigens." Vaccine 24(14): 2692-2699. 184. Mesa, C., De Leon, J., Rigley, K. and Fernandez, L. E. (2004). "Very small size proteoliposomes derived from Neisseria meningitidis: an effective adjuvant for Th1 induction and dendritic cell activation." Vaccine 22(23-24): 3045-3052. 185. Mesa, C., de Leon, J., Rigley, K. and Fernandez, L. E. (2006b). "Very small size proteoliposomes derived from Neisseria meningitidis: an effective adjuvant for dendritic cell activation." Vaccine 24 Suppl 2: S2-42-43. 186. Miescher, S., Whiteside, T. L., Carrel, S. and von Fliedner, V. (1986). "Functional properties of tumor-infiltrating and blood lymphocytes in patients with solid tumors: effects of tumor cells and their supernatants on proliferative responses of lymphocytes." J Immunol 136(5): 18991907. 187. MINSAP (2013) "Anuario Estadístico de Salud 2012." 188. Mitchell, M. S., Kan-Mitchell, J., Kempf, R. A., Harel, W., Shau, H. Y. and Lind, S. (1988). "Active specific immunotherapy for melanoma: phase I trial of allogeneic lysates and a novel adjuvant." Cancer Res 48(20): 5883-5893. 189. Miyoshi, E., Moriwaki, K. and Nakagawa, T. (2008). "Biological function of fucosylation in cancer biology." J Biochem 143(6): 725-729. 190. Moodycliffe, A. M., Nghiem, D., Clydesdale, G. and Ullrich, S. E. (2000). "Immune suppression and skin cancer development: regulation by NKT cells." Nat Immunol 1(6): 521-525. 116 Bibliografía 191. Moore, A., McCarthy, L. and Mills, K. H. (1999). "The adjuvant combination monophosphoryl lipid A and QS21 switches T cell responses induced with a soluble recombinant HIV protein from Th2 to Th1." Vaccine 17(20-21): 2517-2527. 192. Morera, Y., Bequet-Romero, M., Ayala, M., Velazco, J. C., Perez, P. P., Alba, J. S., Ancizar, J., Rodriguez, M., Cosme, K. and Gavilondo, J. V. (2010). "Immunogenicity and some safety features of a VEGF-based cancer therapeutic vaccine in rats, rabbits and non-human primates." Vaccine 28(19): 3453-3461. 193. Morton, D. L., Hsueh, E. C., Essner, R., Foshag, L. J., O'Day, S. J., Bilchik, A., Gupta, R. K., Hoon, D. S., Ravindranath, M., Nizze, J. A., Gammon, G., Wanek, L. A., Wang, H. J. and Elashoff, R. M. (2002). "Prolonged survival of patients receiving active immunotherapy with Canvaxin therapeutic polyvalent vaccine after complete resection of melanoma metastatic to regional lymph nodes." Ann Surg 236(4): 438-448; discussion 448-439. 194. Muchmore, E. A., Varki, N. M., Fukuda, M. and Varki, A. (1987). "Developmental regulation of sialic acid modifications in rat and human colon." FASEB J 1(3): 229-235. 195. Mujoo, K., Cheresh, D. A., Yang, H. M. and Reisfeld, R. A. (1987). "Disialoganglioside GD2 on human neuroblastoma cells: target antigen for monoclonal antibody-mediated cytolysis and suppression of tumor growth." Cancer Res 47(4): 1098-1104. 196. Mulens, V., de la Torre, A., Marinello, P., Rodriguez, R., Cardoso, J., Diaz, R., O'Farrill, M., Macias, A., Viada, C., Saurez, G., Carr, A., Crombet, T., Mazorra, Z., Perez, R. and Fernandez, L. E. (2010). "Immunogenicity and safety of a NeuGcGM3 based cancer vaccine: Results from a controlled study in metastatic breast cancer patients." Hum Vaccin 6(9). 197. Naftzger, C., Takechi, Y., Kohda, H., Hara, I., Vijayasaradhi, S. and Houghton, A. N. (1996). "Immune response to a differentiation antigen induced by altered antigen: a study of tumor rejection and autoimmunity." Proc Natl Acad Sci U S A 93(25): 14809-14814. 198. Naito, Y., Takematsu, H., Koyama, S., Miyake, S., Yamamoto, H., Fujinawa, R., Sugai, M., Okuno, Y., Tsujimoto, G., Yamaji, T., Hashimoto, Y., Itohara, S., Kawasaki, T., Suzuki, A. and Kozutsumi, Y. (2007). "Germinal center marker GL7 probes activation-dependent repression of Nglycolylneuraminic acid, a sialic acid species involved in the negative modulation of B-cell activation." Mol Cell Biol 27(8): 3008-3022. 199. Neninger, E., Diaz, R. M., de la Torre, A., Rives, R., Diaz, A., Saurez, G., Gabri, M. R., Alonso, D. F., Wilkinson, B., Alfonso, A. M., Combet, T., Perez, R. and Vazquez, A. M. (2007). "Active immunotherapy with 1E10 anti-idiotype vaccine in patients with small cell lung cancer: report of a phase I trial." Cancer Biol Ther 6(2): 145-150. 200. Nores, G. A., Dohi, T., Taniguchi, M. and Hakomori, S. (1987). "Density-dependent recognition of cell surface GM3 by a certain anti-melanoma antibody, and GM3 lactone as a possible immunogen: requirements for tumor-associated antigen and immunogen." J Immunol 139(9): 3171-3176. 201. Nowell, P. C. (1986). "Mechanisms of tumor progression." Cancer Res 46(5): 2203-2207. 117 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 202. Offner, H., Thieme, T. and Vandenbark, A. A. (1987). "Gangliosides induce selective modulation of CD4 from helper T lymphocytes." J Immunol 139(10): 3295-3305. 203. Ogawa, J., Inoue, H. and Koide, S. (1997). "Glucose-transporter-type-I-gene amplification correlates with sialyl-Lewis-X synthesis and proliferation in lung cancer." Int J Cancer 74(2): 189192. 204. Oliva, J. P., Valdes, Z., Casaco, A., Pimentel, G., Gonzalez, J., Alvarez, I., Osorio, M., Velazco, M., Figueroa, M., Ortiz, R., Escobar, X., Orozco, M., Cruz, J., Franco, S., Diaz, M., Roque, L., Carr, A., Vazquez, A. M., Mateos, C., Rubio, M. C., Perez, R. and Fernandez, L. E. (2006). "Clinical evidences of GM3 (NeuGc) ganglioside expression in human breast cancer using the 14F7 monoclonal antibody labelled with (99m)Tc." Breast Cancer Res Treat 96(2): 115-121. 205. Oliver, L., Fernandez, A., Raymond, J., Lopez-Requena, A., Fernandez, L. E. and Mesa, C. (2012). "Very small size proteoliposomes derived from Neisseria meningitidis: an effective adjuvant for antigen-specific cytotoxic T lymphocyte response stimulation under leukopenic conditions." Vaccine 30(19): 2963-2972. 206. Osorio, M., Gracia, E., Reigosa, E., Hernandez, J., de la Torre, A., Saurez, G., Perez, K., Viada, C., Cepeda, M., Carr, A., Avila, Y., Rodriguez, M. and Fernandez, L. E. (2012). "Effect of vaccination with N-glycolyl GM3/VSSP vaccine by subcutaneous injection in patients with advanced cutaneous melanoma." Cancer Manag Res 4: 341-345. 207. Osorio, M., Gracia, E., Rodriguez, E., Saurez, G., Arango Mdel, C., Noris, E., Torriella, A., Joan, A., Gomez, E., Anasagasti, L., Gonzalez, J. L., Melgares Mde, L., Torres, I., Gonzalez, J., Alonso, D., Rengifo, E., Carr, A., Perez, R. and Fernandez, L. E. (2008). "Heterophilic NeuGcGM3 ganglioside cancer vaccine in advanced melanoma patients: results of a Phase Ib/IIa study." Cancer Biol Ther 7(4): 488-495. 208. Ozawa, H., Kawashima, I. and Tai, T. (1992a). "Generation of murine monoclonal antibodies specific for N-glycolylneuraminic acid-containing gangliosides." Arch Biochem Biophys 294(2): 427-433. 209. Ozawa, H., Kotani, M., Kawashima, I. and Tai, T. (1992b). "Generation of one set of monoclonal antibodies specific for b-pathway ganglio-series gangliosides." Biochim Biophys Acta 1123(2): 184-190. 210. Park, J. E., Wu, D. Y., Prendes, M., Lu, S. X., Ragupathi, G., Schrantz, N. and Chapman, P. B. (2008). "Fine specificity of natural killer T cells against GD3 ganglioside and identification of GM3 as an inhibitory natural killer T-cell ligand." Immunology 123(1): 145-155. 211. Paulson, J. C., Macauley, M. S. and Kawasaki, N. (2012). "Siglecs as sensors of self in innate and adaptive immune responses." Ann N Y Acad Sci 1253: 37-48. 212. Pawelec, G. (2004). "Tumour escape from the immune response." Cancer Immunol Immunother 53(10): 843. 118 Bibliografía 213. Peguet-Navarro, J., Sportouch, M., Popa, I., Berthier, O., Schmitt, D. and Portoukalian, J. (2003). "Gangliosides from human melanoma tumors impair dendritic cell differentiation from monocytes and induce their apoptosis." J Immunol 170(7): 3488-3494. 214. Perez, C. A., Fu, A., Onishko, H., Hallahan, D. E. and Geng, L. (2009). "Radiation induces an antitumour immune response to mouse melanoma." Int J Radiat Biol 85(12): 1126-1136. 215. Perez, K., Osorio, M., Hernandez, J., Carr, A. and Fernandez, L. E. (2013). "NGcGM3/VSSP vaccine as treatment for melanoma patients." Hum Vaccin Immunother 9(6): 1237-1240. 216. Pilla, L., Patuzzo, R., Rivoltini, L., Maio, M., Pennacchioli, E., Lamaj, E., Maurichi, A., Massarut, S., Marchiano, A., Santantonio, C., Tosi, D., Arienti, F., Cova, A., Sovena, G., Piris, A., Nonaka, D., Bersani, I., Di Florio, A., Luigi, M., Srivastava, P. K., Hoos, A., Santinami, M. and Parmiani, G. (2006). "A phase II trial of vaccination with autologous, tumor-derived heat-shock protein peptide complexes Gp96, in combination with GM-CSF and interferon-alpha in metastatic melanoma patients." Cancer Immunol Immunother 55(8): 958-968. 217. Portoukalian, J., Carrel, S., Dore, J. F. and Rumke, P. (1991). "Humoral immune response in disease-free advanced melanoma patients after vaccination with melanoma-associated gangliosides. EORTC Cooperative Melanoma Group." Int J Cancer 49(6): 893-899. 218. Portoukalian, J., David, M. J., Gain, P. and Richard, M. (1993). "Shedding of GD2 ganglioside in patients with retinoblastoma." Int J Cancer 53(6): 948-951. 219. Portoukalian, J., Zwingelstein, G. and Dore, J. F. (1979). "Lipid composition of human malignant melanoma tumors at various levels of malignant growth." Eur J Biochem 94(1): 19-23. 220. Pukel, C. S., Lloyd, K. O., Travassos, L. R., Dippold, W. G., Oettgen, H. F. and Old, L. J. (1982). "GD3, a prominent ganglioside of human melanoma. Detection and characterisation by mouse monoclonal antibody." J Exp Med 155(4): 1133-1147. 221. Raghuraman, G., Geng, Y. and Wang, C. R. (2006). "IFN-beta-mediated up-regulation of CD1d in bacteria-infected APCs." J Immunol 177(11): 7841-7848. 222. Ragupathi, G., Koide, F., Sathyan, N., Kagan, E., Spassova, M., Bornmann, W., Gregor, P., Reis, C. A., Clausen, H., Danishefsky, S. J. and Livingston, P. O. (2003a). "A preclinical study comparing approaches for augmenting the immunogenicity of a heptavalent KLH-conjugate vaccine against epithelial cancers." Cancer Immunol Immunother 52(10): 608-616. 223. Ragupathi, G., Livingston, P. O., Hood, C., Gathuru, J., Krown, S. E., Chapman, P. B., Wolchok, J. D., Williams, L. J., Oldfield, R. C. and Hwu, W. J. (2003b). "Consistent antibody response against ganglioside GD2 induced in patients with melanoma by a GD2 lactone-keyhole limpet hemocyanin conjugate vaccine plus immunological adjuvant QS-21." Clin Cancer Res 9(14): 5214-5220. 224. Ramirez, B. S., Alpizar, Y. A., Fernandez, D. R., Hidalgo, G. G., Capote, A. R., Rodriguez, R. P. and Fernandez, L. E. (2008). "Anti-EGFR activation, anti-proliferative and pro-apoptotic effects of polyclonal antibodies induced by EGFR-based cancer vaccine." Vaccine 26(38): 4918-4926. 119 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 225. Ravindranath, M. H., Brazeau, S. M. and Morton, D. L. (1994). "Efficacy of tumor cell vaccine after incorporating monophosphoryl lipid A (MPL) in tumor cell membranes containing tumorassociated ganglioside." Experientia 50(7): 648-653. 226. Ravindranath, M. H., Hsueh, E. C., Verma, M., Ye, W. and Morton, D. L. (2003). "Serum total ganglioside level correlates with clinical course in melanoma patients after immunotherapy with therapeutic cancer vaccine." J Immunother 26(3): 277-285. 227. Ravindranath, M. H., Morton, D. L. and Irie, R. F. (1989). "An epitope common to gangliosides O-acetyl-GD3 and GD3 recognized by antibodies in melanoma patients after active specific immunotherapy." Cancer Res 49(14): 3891-3897. 228. Ravindranath, M. H., Tsuchida, T., Morton, D. L. and Irie, R. F. (1991). "Ganglioside GM3:GD3 ratio as an index for the management of melanoma." Cancer 67(12): 3029-3035. 229. Rebbaa, A., Hurh, J., Yamamoto, H., Kersey, D. S. and Bremer, E. G. (1996). "Ganglioside GM3 inhibition of EGF receptor mediated signal transduction." Glycobiology 6(4): 399-406. 230. Reglero, A., Hueso, P., Rodrigo, M., Garcia-Alonso, J., Llanillo, M. and Cabezas, J. A. (1982). "[Gangliosides of neural and extraneural tissues of various species of mammals]." Rev Esp Fisiol 38 Suppl: 37-43. 231. Reis, C. A., Osorio, H., Silva, L., Gomes, C. and David, L. (2010). "Alterations in glycosylation as biomarkers for cancer detection." J Clin Pathol 63(4): 322-329. 232. Ritter, G., Boosfeld, E., Adluri, R., Calves, M., Oettgen, H. F., Old, L. J. and Livingston, P. (1991). "Antibody response to immunization with ganglioside GD3 and GD3 congeners (lactones, amide and gangliosidol) in patients with malignant melanoma." Int J Cancer 48(3): 379-385. 233. Robinson, A. and Melling, J. (1993). "Envelope structure and the development of new vaccines." J Appl Bacteriol 74 Suppl: 43S-51S. 234. Rodriguez, T., Perez, O., Menager, N., Ugrinovic, S., Bracho, G. and Mastroeni, P. (2005). "Interactions of proteoliposomes from serogroup B Neisseria meningitidis with bone marrowderived dendritic cells and macrophages: adjuvant effects and antigen delivery." Vaccine 23(10): 1312-1321. 235. Rojas, G., Pupo, A., Gomez, S., Krengel, U. and Moreno, E. (2013). "Engineering the binding site of an antibody against N-glycolyl GM3: from functional mapping to novel anti-ganglioside specificities." ACS Chem Biol 8(2): 376-386. 236. Roque-Navarro, L., Chakrabandhu, K., de Leon, J., Rodriguez, S., Toledo, C., Carr, A., de Acosta, C. M., Hueber, A. O. and Perez, R. (2008). "Anti-ganglioside antibody-induced tumor cell death by loss of membrane integrity." Mol Cancer Ther 7(7): 2033-2041. 237. Rosenberg, S. A. (1999). "A new era for cancer immunotherapy based on the genes that encode cancer antigens." Immunity 10(3): 281-287. 120 Bibliografía 238. Rosenberg, S. A., Yang, J. C. and Restifo, N. P. (2004). "Cancer immunotherapy: moving beyond current vaccines." Nat Med 10(9): 909-915. 239. Rosner, H. (1975). "Changes in the contents of gangliosides and glycoproteins and in the ganglioside pattern of the chicken brain." J Neurochem 24(4): 815-816. 240. Sa, G., Das, T., Moon, C., Hilston, C. M., Rayman, P. A., Rini, B. I., Tannenbaum, C. S. and Finke, J. H. (2009). "GD3, an overexpressed tumor-derived ganglioside, mediates the apoptosis of activated but not resting T cells." Cancer Res 69(7): 3095-3104. 241. Sahin, U., Tureci, O., Schmitt, H., Cochlovius, B., Johannes, T., Schmits, R., Stenner, F., Luo, G., Schobert, I. and Pfreundschuh, M. (1995). "Human neoplasms elicit multiple specific immune responses in the autologous host." Proc Natl Acad Sci U S A 92(25): 11810-11813. 242. Saito, M. (1993). "Bioactive gangliosides: differentiation inducers for hematopoietic cells and their mechanism(s) of actions." Adv Lipid Res 25: 303-327. 243. Saito, M., Kasai, N. and Yu, R. K. (1985). "In situ immunological determination of basic carbohydrate structures of gangliosides on thin-layer plates." Anal Biochem 148(1): 54-58. 244. Scursoni, A. M., Galluzzo, L., Camarero, S., Lopez, J., Lubieniecki, F., Sampor, C., Segatori, V. I., Gabri, M. R., Alonso, D. F., Chantada, G. and de Davila, M. T. (2011). "Detection of N-glycolyl GM3 ganglioside in neuroectodermal tumors by immunohistochemistry: an attractive vaccine target for aggressive pediatric cancer." Clin Dev Immunol 2011: 245181. 245. Schiavoni, G., Sistigu, A., Valentini, M., Mattei, F., Sestili, P., Spadaro, F., Sanchez, M., Lorenzi, S., D'Urso, M. T., Belardelli, F., Gabriele, L., Proietti, E. and Bracci, L. (2011). "Cyclophosphamide synergizes with type I interferons through systemic dendritic cell reactivation and induction of immunogenic tumor apoptosis." Cancer Res 71(3): 768-778. 246. Schild, H., Arnold-Schild, D., Lammert, E. and Rammensee, H. G. (1999). "Stress proteins and immunity mediated by cytotoxic T lymphocytes." Curr Opin Immunol 11(1): 109-113. 247. Schnaar, R. L. (1991). "Glycosphingolipids in cell surface recognition." Glycobiology 1(5): 477485. 248. Segatori, V. I., Otero, L. L., Fernandez, L. E., Gomez, D. E., Alonso, D. F. and Gabri, M. R. (2012a). "Antitumor protection by NGcGM3/VSSP vaccine against transfected B16 mouse melanoma cells overexpressing N-glycolylated gangliosides." In Vivo 26(4): 609-617. 249. Segatori, V. I., Vazquez, A. M., Gomez, D. E., Gabri, M. R. and Alonso, D. F. (2012b). "Preclinical evaluation of racotumomab, an anti-idiotype monoclonal antibody to N-glycolyl-containing gangliosides, with or without chemotherapy in a mouse model of non-small cell lung cancer." Front Oncol 2: 160. 250. Shamshiev, A., Donda, A., Carena, I., Mori, L., Kappos, L. and De Libero, G. (1999). "Self glycolipids as T-cell autoantigens." Eur J Immunol 29(5): 1667-1675. 121 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 251. Sherman, A. A., Yudina, O. N., Shashkov, A. S., Menshov, V. M. and Nifantiev, N. E. (2002). "Preparative route to N-glycolylneuraminic acid phenyl 2-thioglycoside donor and synthesis of Neu5Gc-alpha-(2-->3')-lactosamine 3-aminopropyl glycoside." Carbohydr Res 337(5): 451-457. 252. Shida, K., Korekane, H., Misonou, Y., Noura, S., Ohue, M., Takahashi, H., Ohigashi, H., Ishikawa, O. and Miyamoto, Y. (2010). "Novel ganglioside found in adenocarcinoma cells of Lewisnegative patients." Glycobiology 20(12): 1594-1606. 253. Shurin, G. V., Shurin, M. R., Bykovskaia, S., Shogan, J., Lotze, M. T. and Barksdale, E. M., Jr. (2001). "Neuroblastoma-derived gangliosides inhibit dendritic cell generation and function." Cancer Res 61(1): 363-369. 254. Siebert, H. C., von der Lieth, C. W., Dong, X., Reuter, G., Schauer, R., Gabius, H. J. and Vliegenthart, J. F. (1996). "Molecular dynamics-derived conformation and intramolecular interaction analysis of the N-acetyl-9-O-acetylneuraminic acid-containing ganglioside GD1a and NMR-based analysis of its binding to a human polyclonal immunoglobulin G fraction with selectivity for O-acetylated sialic acids." Glycobiology 6(6): 561-572. 255. Sierra, G. V., Campa, H. C., Varcacel, N. M., Garcia, I. L., Izquierdo, P. L., Sotolongo, P. F., Casanueva, G. V., Rico, C. O., Rodriguez, C. R. and Terry, M. H. (1991). "Vaccine against group B Neisseria meningitidis: protection trial and mass vaccination results in Cuba." NIPH Ann 14(2): 195-207; discussion 208-110. 256. Skold, M., Xiong, X., Illarionov, P. A., Besra, G. S. and Behar, S. M. (2005). "Interplay of cytokines and microbial signals in regulation of CD1d expression and NKT cell activation." J Immunol 175(6): 3584-3593. 257. Slingluff, C. L., Jr., Petroni, G. R., Yamshchikov, G. V., Barnd, D. L., Eastham, S., Galavotti, H., Patterson, J. W., Deacon, D. H., Hibbitts, S., Teates, D., Neese, P. Y., Grosh, W. W., ChianeseBullock, K. A., Woodson, E. M., Wiernasz, C. J., Merrill, P., Gibson, J., Ross, M. and Engelhard, V. H. (2003). "Clinical and immunologic results of a randomized phase II trial of vaccination using four melanoma peptides either administered in granulocyte-macrophage colony-stimulating factor in adjuvant or pulsed on dendritic cells." J Clin Oncol 21(21): 4016-4026. 258. Slovin, S. F., Keding, S. J. and Ragupathi, G. (2005). "Carbohydrate vaccines as immunotherapy for cancer." Immunol Cell Biol 83(4): 418-428. 259. Sonnino, S., Mauri, L., Chigorno, V. and Prinetti, A. (2007). "Gangliosides as components of lipid membrane domains." Glycobiology 17(1): 1R-13R. 260. Spada, F. M., Borriello, F., Sugita, M., Watts, G. F., Koezuka, Y. and Porcelli, S. A. (2000). "Low expression level but potent antigen presenting function of CD1d on monocyte lineage cells." Eur J Immunol 30(12): 3468-3477. 261. Speak, A. O., Cerundolo, V. and Platt, F. M. (2008). "CD1d presentation of glycolipids." Immunol Cell Biol 86(7): 588-597. 122 Bibliografía 262. Spiegel, S. and Merrill, A. H., Jr. (1996). "Sphingolipid metabolism and cell growth regulation." Faseb J 10(12): 1388-1397. 263. St Hill, C. A., Baharo-Hassan, D. and Farooqui, M. (2011). "C2-O-sLeX glycoproteins are Eselectin ligands that regulate invasion of human colon and hepatic carcinoma cells." PLoS One 6(1): e16281. 264. Steeghs, L., Kuipers, B., Hamstra, H. J., Kersten, G., van Alphen, L. and van der Ley, P. (1999). "Immunogenicity of outer membrane proteins in a lipopolysaccharide-deficient mutant of Neisseria meningitidis: influence of adjuvants on the immune response." Infect Immun 67(10): 4988-4993. 265. Steitz, J., Bruck, J., Lenz, J., Buchs, S. and Tuting, T. (2005). "Peripheral CD8+ T cell tolerance against melanocytic self-antigens in the skin is regulated in two steps by CD4+ T cells and local inflammation: implications for the pathophysiology of vitiligo." J Invest Dermatol 124(1): 144150. 266. Svennerholm, L. (1963). "Chromatographic Separation of Human Brain Gangliosides." J Neurochem 10: 613-623. 267. Swiecki, M. and Colonna, M. (2010). "Unraveling the functions of plasmacytoid dendritic cells during viral infections, autoimmunity, and tolerance." Immunol Rev 234(1): 142-162. 268. Tai, T., Kawashima, I., Furukawa, K. and Lloyd, K. O. (1988). "Monoclonal antibody R24 distinguishes between different N-acetyl- and N-glycolylneuraminic acid derivatives of ganglioside GD3." Arch Biochem Biophys 260(1): 51-55. 269. Tai, T., Paulson, J. C., Cahan, L. D. and Irie, R. F. (1983). "Ganglioside GM2 as a human tumor antigen (OFA-I-1)." Proc Natl Acad Sci U S A 80(17): 5392-5396. 270. Takahashi, T., Johnson, T. D., Nishinaka, Y., Morton, D. L. and Irie, R. F. (1999). "IgM antiganglioside antibodies induced by melanoma cell vaccine correlate with survival of melanoma patients." J Invest Dermatol 112(2): 205-209. 271. Tangvoranuntakul, P., Gagneux, P., Diaz, S., Bardor, M., Varki, N., Varki, A. and Muchmore, E. (2003). "Human uptake and incorporation of an immunogenic nonhuman dietary sialic acid." Proc Natl Acad Sci U S A 100(21): 12045-12050. 272. Teneberg, S., Willemsen, P., de Graaf, F. K. and Karlsson, K. A. (1990). "Receptor-active glycolipids of epithelial cells of the small intestine of young and adult pigs in relation to susceptibility to infection with Escherichia coli K99." FEBS Lett 263(1): 10-14. 273. Thomas, P. D. and Brewer, G. J. (1990). "Gangliosides and synaptic transmission." Biochim Biophys Acta 1031(3): 277-289. 274. Torrens, I., Mendoza, O., Batte, A., Reyes, O., Fernandez, L. E., Mesa, C. and Guillen, G. (2005). "Immunotherapy with CTL peptide and VSSP eradicated established human papillomavirus (HPV) type 16 E7-expressing tumors." Vaccine 23(50): 5768-5774. 123 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 275. Tsuchida, T., Otsuka, H., Niimura, M., Inoue, Y., Kukita, A., Hashimoto, Y., Seyama, Y. and Yamakawa, T. (1984). "Biochemical study on gangliosides in neurofibromas and neurofibrosarcomas of Recklinghausen's disease." J Dermatol 11(2): 129-138. 276. Tsuchida, T., Saxton, R. E., Morton, D. L. and Irie, R. F. (1989). "Gangliosides of human melanoma." Cancer 63(6): 1166-1174. 277. Tsuji, S., Matsumoto, M., Takeuchi, O., Akira, S., Azuma, I., Hayashi, A., Toyoshima, K. and Seya, T. (2000). "Maturation of human dendritic cells by cell wall skeleton of Mycobacterium bovis bacillus Calmette-Guerin: involvement of toll-like receptors." Infect Immun 68(12): 6883-6890. 278. Uronen-Hansson, H., Steeghs, L., Allen, J., Dixon, G. L., Osman, M., van der Ley, P., Wong, S. Y., Callard, R. and Klein, N. (2004). "Human dendritic cell activation by Neisseria meningitidis: phagocytosis depends on expression of lipooligosaccharide (LOS) by the bacteria and is required for optimal cytokine production." Cell Microbiol 6(7): 625-637. 279. van Heyningen, S. (1980). "Tetanus toxin." Pharmacol Ther 11(1): 141-157. 280. Varki, A., Freeze, H. H. and Manzi, A. E. (2001). "Overview of glycoconjugate analysis." Curr Protoc Protein Sci Chapter 12: Unit 12 11. 281. Vazquez, A. M., Hernandez, A. M., Macias, A., Montero, E., Gomez, D. E., Alonso, D. F., Gabri, M. R. and Gomez, R. E. (2012). "Racotumomab: an anti-idiotype vaccine related to N-glycolylcontaining gangliosides - preclinical and clinical data." Front Oncol 2: 150. 282. Vedralova, E., Borovansky, J. and Hach, P. (1995). "Ganglioside profiles of experimental melanomas and of their melanosomal fractions." Melanoma Res 5(2): 87-92. 283. Venier, C., Guthmann, M. D., Fernandez, L. E. and Fainboim, L. (2007). "Innate-immunity cytokines induced by very small size proteoliposomes, a Neisseria-derived immunological adjuvant." Clin Exp Immunol 147(2): 379-388. 284. Vermi, W., Soncini, M., Melocchi, L., Sozzani, S. and Facchetti, F. (2011). "Plasmacytoid dendritic cells and cancer." J Leukoc Biol 90(4): 681-690. 285. Voet D, V. J. (1995). Lipids and membranas. Biochemistry: 282-284. 286. Wang, J. and Yu, R. K. (2013). "Interaction of ganglioside GD3 with an EGF receptor sustains the self-renewal ability of mouse neural stem cells in vitro." Proc Natl Acad Sci U S A 110(47): 19137-19142. 287. Wei, J., Cui, L., Liu, F., Fan, Y., Lang, R., Gu, F., Guo, X., Tang, P. and Fu, L. (2010). "E-selectin and Sialyl Lewis X expression is associated with lymph node metastasis of invasive micropapillary carcinoma of the breast." Int J Surg Pathol 18(3): 193-200. 288. Wiegandt, H. (1980). "Introductory remarks on chemical structures of gangliosides." Adv Exp Med Biol 125: 3-10. 124 Bibliografía 289. Wiegandt, H. and Schulze, B. (1969). "Spleen gangliosides: "the structure of ganglioside GLNnT1(NGNA)"." Z Naturforsch B 24(7): 945-946. 290. Wipfler, D., Srinivasan, G. V., Sadick, H., Kniep, B., Arming, S., Willhauck-Fleckenstein, M., Vlasak, R., Schauer, R. and Schwartz-Albiez, R. (2011). "Differentially regulated expression of 9O-acetyl GD3 (CD60b) and 7-O-acetyl-GD3 (CD60c) during differentiation and maturation of human T and B lymphocytes." Glycobiology 21(9): 1161-1172. 291. Wu, D. Y., Segal, N. H., Sidobre, S., Kronenberg, M. and Chapman, P. B. (2003). "Crosspresentation of disialoganglioside GD3 to natural killer T cells." J Exp Med 198(1): 173-181. 292. Wu, Z. L., Schwartz, E., Seeger, R. and Ladisch, S. (1986). "Expression of GD2 ganglioside by untreated primary human neuroblastomas." Cancer Res 46(1): 440-443. 293. Yin, J., Miyazaki, K., Shaner, R. L., Merrill, A. H., Jr. and Kannagi, R. (2010). "Altered sphingolipid metabolism induced by tumor hypoxia - new vistas in glycolipid tumor markers." FEBS Lett 584(9): 1872-1878. 294. Yoshida, S., Kawaguchi, H., Sato, S., Ueda, R. and Furukawa, K. (2002). "An anti-GD2 monoclonal antibody enhances apoptotic effects of anti-cancer drugs against small cell lung cancer cells via JNK (c-Jun terminal kinase) activation." Jpn J Cancer Res 93(7): 816-824. 295. Zhang, C., Zhang, J. and Tian, Z. (2006). "The regulatory effect of natural killer cells: do "NK-reg cells" exist?" Cell Mol Immunol 3(4): 241-254. 296. Zhang, H., Zhang, S., Cheung, N. K., Ragupathi, G. and Livingston, P. O. (1998). "Antibodies against GD2 ganglioside can eradicate syngeneic cancer micrometastases." Cancer Res 58(13): 2844-2849. 297. Zhang, S., Cordon-Cardo, C., Zhang, H. S., Reuter, V. E., Adluri, S., Hamilton, W. B., Lloyd, K. O. and Livingston, P. O. (1997). "Selection of tumor antigens as targets for immune attack using immunohistochemistry: I. Focus on gangliosides." Int J Cancer 73(1): 42-49. 298. Zhong, Y., Wu, Y., Li, C., Tang, J., Wang, X., Ren, G., Carr, A., Perez, R. and Guo, W. (2012). "NGlycolyl GM3 ganglioside immunoexpression in oral mucosal melanomas of Chinese." Oral Dis 18(8): 741-747. 299. Ziegler-Heitbrock, H. W., Kafferlein, E., Haas, J. G., Meyer, N., Strobel, M., Weber, C. and Flieger, D. (1992). "Gangliosides suppress tumor necrosis factor production in human monocytes." J Immunol 148(6): 1753-1758. 125 9. ANEXOS Anexo 1. Esquema del mecanismo de acción del preparado vacunal NGcGM3/VSSP. Leyenda: 127 AUTOBÍBLÍOGRAFÍA Publicaciones relacionadas con el tema de la tesis: 1. Labrada, M., Clavell, M., Bebelagua, Y., Leon, J., Alonso, D. F., Gabri, M. R., Veloso, R. C., Verez, V. and Fernandez, L. E. (2010). "Direct validation of NGcGM3 ganglioside as a new target for cancer immunotherapy." Expert Opin Biol Ther 10(2): 153-162. 2. Labrada, M., Pablos, I., Prete, F., Hevia, G., Clavell, M., Benvenuti, F., and Fernandez, L. E. (2014). "Induction of leukocyte infiltration at metastatic site mediates the protective effect of NGcGM3-based vaccine." Hum Vaccin Immunother 10(8). Presentaciones en eventos científicos relacionados con el tema de tesis: 1. International Workshop (IT2012), 2012. New insights of the relation of the NGcGM3-vaccine and the innate immunity. Cartel. 2. International Workshop (IT2010), 2010. Direct validation of NGcGM3 ganglioside as a new target for cancer immunotherapy. Cartel. 3. X Congreso de Oncología, 2009. NGcGM3/VSSP vaccine: Preclinical studies of the mechanism of action. Presentación Oral. 4. 1er Congreso Internacional de Farmacología de Vacunas (VacciPharma), 2009. Cellular immune response dependence for antimetastatic effect of a tumor associated gangliosidecontaining vaccine. Cartel. 5. Inmunología 2008. VI Congreso Nacional de Inmunología, 2008. Efecto antimetastásico de la vacuna NGcGM3/VSSP en un modelo de metástasis pulmonar espontánea. Cartel. 6. International Workshop (IT2008), 2008. Cellular immune response dependence for antimetastatic effect of a tumor associated ganglioside-containing vaccine. Presentación Oral. 7. International Workshop “Immunotherapy for the new century”, 2004. Inhibitory effect of NGcGM3/VSSP vaccine on metastasis in mice. Cartel. 8. International Workshop “Immunotherapy for the new century”, 2004. Antitumor activity of VSSP∕GM3 vaccine depends on CD4+ and CD8+ T lymphocytes. Cartel. Otras publicaciones de la autora: 1. Prete, F., Catucci, M., Labrada, M., Gobessi, S., Castiello, M. C., Bonomi, E., Aiuti, A., Vermi, W., Cancrini, C., Metin, A., Hambleton, S., Bredius, R., Notarangelo, L. D., van der Burg, M., Kalinke, U., Villa, A. and Benvenuti, F. (2013). "Wiskott-Aldrich syndrome protein-mediated actin dynamics control type-I interferon production in plasmacytoid dendritic cells." J Exp Med 210(2): 355-374. 129 Mecanismos inmunológicos asociados a la vacunación con NGcGM3/VSSP 2. Labrada, M., Alonso, D.F., Gabri, M., Clavell, M., Bebelagua, Y., de León, J., Fernández, L.E. “Cellular immunoresponse dependence for antimetastatic effect of a tumor associated ganglioside-containing vaccine”. VacciMonitor (2009); Vol. 18 Año 1 Suppl. 1 3. de León, J., Fernández, A., Clavel, M., López, A. , Labrada, M., Bebelagua, Y. , Mesa, C., Fernández, L.E. “The N-glycolylated variant of ganglioside GM3 in tumor biology: An attractive target for cancer immunotherapy”. Biotecnol Apl. (2008), vol.25, no.2, p.166-170. ISSN 10272852. 4. de Leon, J., Fernandez, A., Clavell, M., Labrada, M., Bebelagua, Y., Mesa, C. and Fernandez, L. E. (2008). "Differential influence of the tumour-specific non-human sialic acid containing GM3 ganglioside on CD4+CD25- effector and naturally occurring CD4+CD25+ regulatory T cells function." Int Immunol 20(4): 591-600 5. Puerta L., Labrada, A., Labrada, M., Uyema, K., Jiménez, S., Caraballo, L. “Blo t 13 Allergen from Blomia tropicalis Shows High Frequency of IgE Binding in Allergic Cuban Patients and Cross-reactivity with Dermatophagoides siboney Extract.” The Journal of Allergy and Clinical Immunology. (2003), part 2, Volume 111, Number 2, S1028. 6. Labrada, M., Uyema, K., Sewer, M., Labrada, A., Gonzalez, M., Caraballo, L. and Puerta, L. (2002). "Monoclonal antibodies against Blo t 13, a recombinant allergen from Blomia tropicalis." Int Arch Allergy Immunol 129(3): 212-218. 7. Sewer, M., Uyema, K., Labrada, M., Gonzalez, M. and Coca, M. (2000). "Monoclonal antibodies against Der s 1, a major allergen of Dermatophagoides siboney." Int Arch Allergy Immunol 123(3): 242-248. 130