ES (2)")
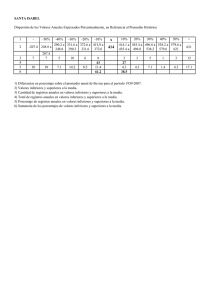
Suscríbete a DeepL Pro para poder editar este documento. Entra en www.DeepL.com/pro para más información. BIOLOGÍA DE LA PLoS Dispersión de Semillas y Patrón Espacial en Árboles Tropicales Tristram G. Seidler1, Joshua B. Plotkin2* 1 Natural Environment Research Council Centre for Population Biology, Imperial College, Ascot, Berkshire, Reino Unido, 2 Department of Biology, University of Pennsylvania, Philadelphia, Pennsylvania, Estados Unidos de América Las teorías de la diversidad de árboles tropicales enfatizan la limitación de la dispersión como un mecanismo potencial para separar especies en el espacio y reducir la exclusión competitiva. Comparamos las morfologías de dispersión, los tamaños de los frutos y las distribuciones espaciales de 561 especies de árboles dentro de una parcela de 50 hectáreas de bosque tropical primario en Malasia peninsular. Demostramos aquí que el alcance y la escala de la agregación espacial concomitante está correlacionada con el modo de dispersión de las semillas. Esta relación es válida tanto para los arbolitos como para los árboles maduros. Los contrastes filogenéticamente independientes confirman que la relación entre la dispersión y el patrón espacial es significativa incluso después de controlar la ascendencia común entre especies. Encontramos los mismos resultados cualitativos para una parcela de bosque tropical de 50 hectáreas en Panamá. Nuestros resultados proporcionan una amplia evidencia empírica de la importancia del modo de dispersión en el establecimiento de la estructura comunitaria a largo plazo de los bosques tropicales. Citación: Seidler TG, Plotkin JB (2006) Dispersión de semillas y patrón espacial en árboles tropicales. PLoS Biol 4(11): e344. DOI: 10.1371/journal.pbio. 0040344 Introducción Las comunidades de árboles de los bosques tropicales se encuentran entre las más ricas en especies de la Tierra. Aunque el mantenimiento de la diversidad sigue siendo un problema central en la ecología[1], el trabajo teórico ha puesto de relieve la agregación de conspecific como un mecanismo para reducir la exclusión competitiva y promover la diversidad[2,3]. De hecho, los bosques tropicales exhiben una amplia agregación de árboles a escalas que van desde unos pocos metros hasta unos pocos cientos de metros[4-6]. La causa de la agrupación de conspecific sigue siendo poco clara[7], y se ha atribuido de diversas maneras a la variación desigual del hábitat[5,8], a la dispersión limitada de las semillas[9], o a procesos neutros que no tienen en cuenta las características de las especies-specific[10]. Aquí demostramos que las morfologías de dispersión están fuertemente correlacionadas con las distribuciones espaciales de cientos de especies de árboles y , por lo tanto, con la estructura comunitaria de los bosques tropicales. Las especies de árboles tropicales varían en su capacidad de dispersar semillas. Se sabe que la dispersión limitada causa agregación espacial entre semillas y plántulas de árboles pioneros[11]. Si los patrones espaciales producidos por la dispersión limitada persisten o no más allá de la etapa de las plántulas es menos conocido, aparte de la evidencia anecdótica o los estudios limitados a unas pocas especies[4,6,7]. Establecer un vínculo entre los mecanismos de dispersión y los patrones espaciales a nivel de la comunidad ayudaría a cerrar la brecha en el ' 'bucle demográfico'' que separa las observaciones de la dispersión limitada de semillas de las consecuencias a largo plazo de la dispersión para las poblaciones de árboles[7,12,13]. La agregación espacial inducida por la dispersión local podría PLoS Biología | www.plosbiology.org 2132 verse reforzada por asociaciones con hábitats irregulares[14] o podría verse perturbada por una mortalidad dependiente de la densidad debida a la depredación[15,16]. Sin embargo, planteamos la hipótesis de que los árboles de una especie con dispersión limitada de semillas se agruparán estrechamente en el espacio, mientras que una especie con un mecanismo para la dispersión de semillas a larga distancia exhibirá menos agrupamiento o incluso aleatoriedad espacial. Para examinar esta hipótesis, analizamos los mecanismos de dispersión y las distribuciones espaciales de los árboles dentro de una parcela de 50 hectáreas de bosque tropical de tierras bajas en Malasia peninsular (ver Materiales y Métodos). El censo incluye todos los Noviembre 2006 | Volumen 4 | Número 11 | e344 árboles de más de 1 cm de diámetro a la altura del pecho, mapeados con una precisión de 1 m[17]. Dividimos estas especies en síndromes de dispersión primaria, basados en su anatomía y morfología del fruto, y preguntamos si los síndromes de dispersión se correlacionan con las distribuciones espaciales. Resulta dos La variación en la morfología de las semillas y los frutos se debe en parte a la selección de las capacidades de dispersión[18]. Alas y plumas, así como fleshy, tejidos jugosos y nutritivos, han surgido muchas veces a través de un amplio rango taxonómico[19], y se originan en una variedad de capas histológicas de la testa, el pericarpio y tejidos adyacentes. Asignamos cada una de las 561 especies estudiadas a uno de los siete síndromes de dispersión, sobre la base de los datos de las colecciones de field, los especímenes de herbario y las descripciones publicadas en flora[20-24]. Los siete síndromes de dispersión son: balístico , gravedad, giro, viento, animal (tamaño de fruta pequeño), animal (fruta mediana) y animal (fruta grande) (Tabla 1). Con el fin de cuantificar el grado global de agregación espacial para cada especie, se realizó un proceso de puntos de conglomerados de Poisson para la distribución observada de los individuos de conspecific en la parcela (ver Materiales y Métodos). El proceso del cluster de Poisson, y este método de los parámetros de fitting en particular, reproduce fielmente los patrones espaciales cualitativos de la mayoría de las especies[5]. Como resultado de fitting un proceso de cluster de Poisson, para cada especie obtuvimos un promedio ''tamaño del cluster,'' r, que quantifies el diámetro típico de un conspecific Editor Académico: Ran Nathan, Universidad Hebrea de Jerusalén, Israel Recibida el 10 de abril de 2006; aceptada el 14 de agosto de 2006; publicada el 17 de octubre de 2006 DOI: 10.1371/journal.pbio.0040344 Derechos de autor: 2006 Seidler y Plotkin. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia de Reconocimiento Creative Commons, que permite el uso, distribución y reproducción sin restricciones en cualquier medio, siempre y cuando se acredite el autor y la fuente originales. Abreviaturas: BCI, Isla Barro Colorado; df, grados de libertad * A quién debe dirigirse jplotkin@sas.upenn.edu la correspondencia. PLoS Biología | www.plosbiology.org Correo electrónico: 2133 Noviembre 2006 | Volumen 4 | Número 11 | e344 racimo de árboles. Los valores pequeños de r indican agrupaciones espaciales estrechas; grandes valores indican conglomerados más difusos (Figura 1). La Figura 2 muestra l a relación entre la dispersión de sindroma y el tamaño del conglomerado r para nuestras 561 especies de estudio. Las especies dispersas balísticamente exhiben los valores r más pequeños, es decir, las distribuciones espaciales más agregadas, seguidas de las especies dispersas por gravedad, giro, viento y animales. Entre las especies dispersas por los animales, el grado de agrupación espacial depende del diámetro del fruto. Las especies con frutos de menos de 2 cm de diámetro son más agregadas que aquellas con frutos de 2-5 cm de diámetro; y las especies con frutos de más de 5 cm de diámetro muestran la menor concentración espacial (Figura 2). En general, existe una gran relación entre el tamaño de los cúmulos espaciales y el síndrome de dispersión en nuestras 561 especies estudiadas (Kruskal-Wallis, grados de libertad[df] ¼ 6, v2 ¼ 64.3, p , 10 6). Además de este análisis general de varianza, se compararon los tamaños de conglomerados espaciales entre pares de síndromes o entre grupos de síndromes. Cada una de estas comparaciones se realizó bajo dos supuestos diferentes sobre las relaciones entre especies: first, tratamos a la especie en estudio como independiente, y en segundo lugar, controlamos las relaciones filogenéticas entre las especies [25] (ver Materiales y Métodos). Al controlar la filogenia, podemos interrogar la relación entre el patrón de agregación y el mecanismo de dispersión, a la vez que evitamos el problema potencial de la pseudo-replicación entre especies que comparten un antepasado común. Aunque los contrastes filogenéticos no pueden descartar la posibilidad de que un tercer rasgo (como la estatura) explique la variación tanto en el síndrome de dispersión como en el tamaño de los conglomerados espaciales, dicho análisis controla los rasgos de evolución neutra determinados por el patrón de ascendencia entre especies. Las especies dispersas por animales exhiben tamaños de conglomerados más grandes que las especies no dispersas por los animales (Wilcoxon signrank test, p............................................................................................. ..................................................................................................... .......... Esta relación es significant incluso después de controlar las relaciones filogenéticas entre las especies estudiadas (p ¼ 0.002). Especies animales dispersas con frutos . Los racimos de 5 cm de diámetro son más grandes que los de las especies dispersas de animales con frutos de ,2 cm de diámetro (p ¼ 0,0007), una relación que también se conserva bajo contrastes filogenéticos independientes (p ¼ 0,001). Las especies dispersas por gravedad exhiben tamaños de racimo más grandes que las especies dispersas balísticamente (p ¼ 0.03), pero esta tendencia no pudo ser probada cuando se controló la filogenia debido a la escasez de contrastes independientes. El tamaño de los frutos también está relacionado con la distribución espacial de los árboles. Entre las especies dispersas por animales, existe una correlación débil pero positiva entre el diámetro del fruto y el tamaño del racimo r (n ¼ 425 especies, Spearman r ¼ 0.13, p ¼ 0.022). Esta correlación permanece significant incluso después de controlar las relaciones filogenéticas (n ¼ 143 contrastes, r ¼ 0.18, p ¼ 0.032). Entre las especies no dispersas por los animales, el diámetro del fruto también está correlacionado con el tamaño del racimo (n ¼ 78 especies, r ¼ 0.33, p ¼ PLoS Biología | www.plosbiology.org 2134 0,0039), y esta correlación sigue siendo significant después del control de la filogenia (n ¼ 39 contrastes, r ¼ 0,39, p ¼ 0,014). Además del tamaño medio del cluster r, podemos cuantificar la agregación sobre un rango de escalas espaciales usando la medida del segundo momento llamada k[26] de Ripley. Para cada especie, la estadística k(d) quantifies el número medio de conspecifics dentro de una distancia d de un árbol focal, dividido por la densidad de conspecifics en l a parcela. Si los individuos se distribuyen al azar, entonces el valor esperado de k(d) es pd2. Informamos de la Noviembre 2006 | Volumen 4 | Número 11 | e344 estadística sin dimensiones K(d) ¼ k(d)/(pd2). K(d) excede la unidad cuando una especie es más agregada que aleatoria en la escala de distancia d; K(d) es menor que la unidad cuando una especie es más regular que aleatoria. En comparación con el tamaño medio del conglomerado r, la estadística K(d) puede proporcionar una descripción más detallada de la relación entre el mecanismo de dispersión y el patrón espacial. Las curvas K(d) dentro de cada síndrome de dispersión (Figura 3) son consistentes con los resultados obtenidos usando el tamaño medio del conglomerado r (Figura 2). La figura 3 revela además que la agregación a pequeña escala (d 75 m) está más fuertemente correlacionada con el síndrome de dispersión que la agregación a gran escala. A escalas espaciales muy grandes (d 200 m), los patrones espaciales ya no están correlacionados con los síndromes de dispersión. Este resultado sugiere que cualquiera que sea la agregación a gran escala que se presente, es probable que sea causada por factores distintos de la dispersión, como las asociaciones con hábitats irregulares. Este resultado es consistente con la escala de variación topográfica de nuestra parcela de estudio[5]. Para examinar la generalidad de nuestros resultados en los trópicos del nuevo y viejo mundo, también hemos analizado los patrones espaciales y los modos de dispersión en la parcela de 50 hectáreas en la Isla de Barro Colorado (BCI), Panamá[27-29]. Existen diferencias sustanciales en el floral y en los ensamblajes faunísticos de ICB y Pasoh, lo que podría inducir diferencias en la relación entre la dispersión y los patrones espaciales. Sin embargo, encontramos resultados cualitativamente similares en el ICB: las especies dispersas balísticamente son las más agregadas (media r ¼ 49,0 m), seguidas por las especies dispersas por el viento (media r ¼ 65,2 m), las especies dispersadas por mamíferos (media r ¼ 112,6 m), y especies dispersas de aves y murciélagos (media r ¼ 146,8 m). La relación entre el modo de dispersión y el patrón espacial en BCI es significant (Kruskal-Wallis, df ¼ 3, v2 9.32, p ¼ 0.025) pero más débil que en Pasoh, lo que se debe en parte al reducido número de especies. Al igual que en Pasoh, las especies dispersas en el ICB son más difusas que las especies que carecen de dispersión animal primaria (p ¼ 0.003). dispersión creciente (Figura 1) ilustra varios puntos importantes relacionados con la historia natural de la dispersión de semillas en árboles tropicales. Las especies dispersas que producen frutos pequeños exhiben racimos más apretados que las que producen frutos más grandes, un resultado que apoya la hipótesis de que las especies más grandes. ¼ Discusión Aunque estudios anteriores han utilizado datos de caída de semillas y censos repetidos para examinar las consecuencias de la dispersión para la distribución de las plántulas[11,30], tal enfoque no es factible en las escalas de tiempo necesarias para estudiar poblaciones de árboles más maduros. En contraste, aquí hemos aprovechado la variación en los mecanismos de dispersión sobre una amplia gama de especies filogenéticas para interrogar la relación entre la capacidad de dispersión y la agrupación espacial . Los tamaños estimados de los racimos, apoyados por las curvas k de Ripley, indican que los síndromes de dispersión con u n mayor potencial para el transporte de semillas a larga distancia resultan en tamaños de racimos más grandes. La mayoría de estas correlaciones permanecen en significant incluso cuando se controla la filogenia, lo que indica que la relación entre la dispersión y el patrón espacial no es probablemente un efecto secundario de la variación en otros rasgos de especies en evolución neutral. Además, es poco probable que esta relación sea causada por la covariación de la estatura del árbol, porque la estatura no muestra correlación con el patrón espacial (Figuras S2 y S3). La secuencia de patrones espaciales a lo largo del eje de PLoS Biología | www.plosbiology.org 2135 Noviembre 2006 | Volumen 4 | Número 11 | e344 Dispersión y Patrón Espacial Tabla 1. La definición de los síndromes de dispersión utilizados en este estudio y las características de las especies dentro de cada síndrome de dispersión Dispersión Síndrome de Morfología de la fruta Fruta o Semilla Número Número Rasgo Relevante de especies de Genera a la Dispersión Balística Gravedad Cápsula explosiva Tuerca, alas ausentes Giros Tuerca de mariposa Viento Cápsula, cápsula, tuerca de mariposa Berry, drupe, o cápsula Berry, drupe, o cápsula Berry, drupe, o cápsula oArilo ausente reducido 28 dispersión por dispersor de semillas 28 dispersión por dispersor de semillas Semilla o fruta diminuta; con plumas o alas Pulpa o arilo comestible Pulpa o arilo comestible Pulpa o arilo comestible Animal ,2 cm Animal 2-5 cm Animal 5 cm Número de Familias Diámetro del fruto, número de árboles Tamaño del racimo Eje largo (mm) por Especie sigma; (m) 16 24 9 14 1 7 19.5 6 3.5 82.0 6 23.4 636 6 155 1,144 6 424 31.1 6 4.7 47.4 6 4.9 29 10 5 107.6 6 15.2 872 6 228 54.5 6 6.2 19 17 11 80.4 6 16.3 300 6 99 64.5 6 13.4 209 97 43 13.3 6 0.35 506 6 56 99.3 6 7.7 177 84 35 32.7 6 0.72 527 6 74 120.6 6 10.6 87 41 20 96.9 6 6.0 312 6 53 157.8 6 17.0 Las especies dispersas por los animales se subdividen en tres grupos en función del diámetro del fruto. Los síndromes de balística, gravedad, giro y viento dependen de medios mecánicos para la dispersión primaria. Muchas especies dispersas por gravedad y giro son dispersadas por los dispersores de semillas de animales, ya sea incidentalmente o a través del almacenamiento en caché. La tabla indica el número de especies, géneros y familias dentro de cada categoría de dispersión primaria; la media de 6 1 error estándar de diámetros de fruta; la media de 6 1 error estándar de abundancia de especies; y la media de 6 1 tamaño de conglomerado de error estándar. DOI: 10.1371/journal.pbio.0040344.t001 los pájaros y mamíferos corporales comen frutos más grandes[18,31], tienen un rango de distribución más grande[32], y pueden llevar semillas a distancias más largas que los pájaros y mamíferos pequeños. Hemos visto que las especies dispersas por el viento típicamente exhiben conglomerados más apretados que las especies dispersas por los animales (Figura 1). Esta tendencia puede ser explicado en parte por el influence del dosel del bosque PLoS Biología | www.plosbiology.org 2136 en la velocidad del viento. Aunque los vientos fuertes pueden ocurrir en Pasoh, la mayoría de las semillas caen a través del dosel del bosque , donde el movimiento del aire está restringido[33]. Las especies dispersas por giro se dispersan ocasionalmente a lo largo de largas distancias debido a los fuertes vientos. Pero tales frutas típicamente tienen un levantamiento limitado, debido a la alta masa. con respecto a la superficie de trabajo[34] (figura S1). Las especies en nuestro Noviembre 2006 | Volumen 4 | Número 11 | e344 Dispersión y Patrón Espacial Figura 1. Ejemplos de poblaciones arbóreas mapeadas de cuatro especies en la parcela forestal de Pasoh de 50 ha Arriba a la izquierda, Baccaurea racemosa (animal dispersado; n ¼ 1,228, r ¼ 146.5 m); abajo a la izquierda, Neobalanocarpus heimii (dispersión por gravedad; n ¼ 3,334, r ¼ 86.7 m); arriba a la derecha, Shorea leprosula (giro disperso; n ¼ 2,154, r ¼ 33.1 m); abajo a la derecha, Croton argyratus (dispersión balística; n ¼ 1,248, r ¼ 27.9 m). DOI: 10.1371/journal.pbio.0040344.g001 PLoS Biología | www.plosbiology.org 2137 Noviembre 2006 | Volumen 4 | Número 11 | e344 Dispersión y Patrón Espacial inconvenientes. El más importante de ellos es nuestra tosca caracterización del"mecanismo de dispersión" en unas cuantas categorías simples. Los mecanismos de dispersión, por no hablar de los núcleos, son ciertamente más variados de lo que nuestras diversas categorías pueden cuantificar. Además, el importante concepto de limitación del reclutamiento[9] es más complejo que el mecanismo de dispersión por sí solo, e incluye las limitaciones de la especie -specific en fecundidad y establecimiento que nuestro análisis ha ignorado. Por lo tanto, nuestros resultados se limitan a la relación entre los modos morfológicos de distribución de propágulos y los patrones espaciales resultantes de los árboles. La posibilidad sigue siendo, por supuesto, que otros rasgos covariables con el modo de dispersión y pueden ser responsables de las correlaciones que hemos observado. Aunque no podemos separar los efectos de la distribución diferencial de propágulos de los de los establecimientos diferenciales, podemos al menos analizar las distribuciones espaciales de árboles maduros bien establecidos. El conjunto de datos completo de Pasoh, analizado anteriormente, incluye árboles jóvenes de muchas especies. Sin embargo, cuando repetimos nuestro análisis de patrones espaciales restringidos sólo a árboles maduros -es decir, tallos de más de 5 cm de diámetro[42]- nos encontramos con la misma relación cualitativa ¼ entre los síndromes de dispersión y las distribuciones espaciales (Figura 4), que todavía es altamente significant (Kruskal-Wallis, df ¼ 6, 46.7, p , 10 6). v2 Figura 2. La Relación entre el Síndrome de Dispersión y el Síndrome Espacial Agregación de 561 especies de árboles en Pasoh, Malasia La figura muestra el error estándar medio de 6 1 del tamaño del conglomerado espacial r) para las especies arbóreas de cada uno de los siete síndromes de dispersión. agregación espacial, r, que es el grado general de agrupamiento[5] en lugar de sólo el patrón local[4]. Sin embargo, nuestro análisis confirms La observación de Hubbell de que las especies en el BCI dispersas por aves o murciélagos exhiben las más difusas patrones. En comparación con los estudios de datos de caída de semillas para un número limitado de especies tratables, nuestro estudio se ve reforzado por el gran número y amplitud filogenética de especies analizadas. Sin embargo, la escala de nuestro análisis implica varios aspectos importantes DOI: 10.1371/journal.pbio.0040344.g002 La categoría de dispersión por gravedad a menudo experimenta una dispersión secundaria desde el suelo después de la caída de la fruta. ¿Por qué, entonces, son estas especies más estrechamente agregadas que las especies primarias dispersas por animales? La respuesta está probablemente relacionada con el hecho de que la dispersión secundaria de semillas que carecen de recompensas secundarias es típicamente realizada por mamíferos, tales como los roedores que depositan las semillas[35-37], que transportan las semillas a distancias relativamente cortas[38]. Las especies dispersas por gravedad y giro son los mástiles dominantes en los bosques tropicales asiáticos[39]. Durante los eventos de fructificación de mástiles, las semillas escapan abrumadoramente a la depredación, ya sea por saciedad de los depredadores[40] o por germinación rápida antes de la llegada de los depredadores migratorios[41]. Por lo tanto, el impacto en la agregación de la dispersión secundaria por parte de los depredadores de semillas puede ser limitado, una hipótesis que se apoya en nuestro análisis de los patrones espaciales en dichas especies. En parte, nuestro findings contradice los primeros estudios sobre el modo de dispersión y el patrón espacial realizados por Hubbell[4]. Al comparar 30 especies de árboles en el ICB, Hubbell encontró que las especies dispersas por los mamíferos eran las más agregadas y reducidas por las especies dispersadas por el viento. En contraste, tanto en el ICB como en Pasoh, encontramos que las especies dispersas por el viento son más agregadas que las dispersas por los mamíferos. Hay varias explicaciones metodológicas para la discrepancia entre nuestros resultados y los de Hubbell. Aparte del hecho de que hemos usado categorías de dispersión potencialmente diferentes y analizado muchas más especies que Hubbell, también hemos usado una métrica diferente de PLoS Biología | www.plosbiology.org 2138 Noviembre 2006 | Volumen 4 | Número 11 | e344 Dispersión y Patrón Espacial La concordancia de nuestros resultados en las diferentes etapas de la vida sugiere que el modo de dispersión en sí mismo juega un papel en la causa de las correlaciones observadas. A pesar de sus limitaciones, nuestro s resultados ayudan a proporcionar una base empírica para las teorías de la asamblea comunitaria y la diversidad mediada por la variación en el modo de dispersión. De acuerdo con modelos teóricos, la agrupación espacial es reconocida como un mecanismo crítico para reducir la exclusión competitiva y promover la diversidad. El efecto de la agrupación en la diversidad depende de la escala. En escalas espaciales pequeñas, la agrupación tiende a reducir la diversidad local (alfa), mientras que en escalas más grandes, la dispersión limitada conduce a una mayor rotación de especies. Figura 3. La Estadística de Agregación Espacial, K(d), Evaluada en un rango de Distancias para especies de árboles en cuatro síndromes de dispersión En cada síndrome de dispersión, el gráfico muestra el valor medio K(d) 6 1 error estándar. Una especie se agrega a la distancia d si K(d) supera unidad. Las líneas de puntos indican K(d) para una distribución espacial aleatoria de Poisson. Todas las especies están fuertemente agregadas a pequeñas escalas espaciales y débilmente agregadas a grandes escalas. En las escalas espaciales d 75 m, cada uno de los tiene un valor K(d) medio significativamente diferente (Wilcoxon p , 0.008). A escalas espaciales mayores (d . 200m), la agregación espacial no es significativamente correlacionado con el síndrome de dispersión. (Los otros tres se omiten los síndromes de dispersión para mayor claridad.) DOI: 10.1371/journal.pbio.0040344.g003 PLoS Biología | www.plosbiology.org 2139 Noviembre 2006 | Volumen 4 | Número 11 | e344 Dispersión y Patrón Espacial las grandes aves y los grandes mamíferos son los probables dispersores de las aves de fruto grande. especie (. 5 cm de diámetro). Debido a que no usamos el tamaño de la semilla como criterio, los frutos grandes con semillas grandes no se distinguen de los frutos grandes con semillas pequeñas, aunque estos últimos a veces pueden ser dispersados por aves o mamíferos pequeños incapaces de tragar frutos. todo. Figura 4. La relación entre el síndrome de dispersión y la agregación espacial entre 425 especies de árboles tropicales, restringida sólo a árboles maduros (diámetro del tallo....................................................................................................... ........................................................................................................) La figura muestra el tamaño medio del conglomerado espacial r para las especies arbóreas en cada uno de los siete síndromes de dispersión. Los síndromes de dispersión están asociados significativamente con la agregación espacial entre árboles maduros (Kruskal-Wallis, df ¼ 6, v2 ¼ 46.7, p. 10 6). DOI: 10.1371/journal.pbio.0040344.g004 aumentar la diversidad total (beta y gamma)[3]. La escala típica de agrupamiento entre las especies de nuestro estudio (r ; 100 m) es significantly menor que la escala de toda la parcela de Pasoh, y por lo tanto esperamos que el agrupamiento actúe generalmente para aumentar la diversidad de especies dentro de la parcela. Además de separar especies en el espacio, la variación en el síndrome de dispersión también puede mantener la diversidad al imponer una compensación entre competencia y colonización, como sugiere un gran cuerpo de literatura teórica[3,43,44]. Materiales y métodos Data. Se estudiaron especies arbóreas dentro de la parcela de 50 hectáreas de dinámica forestal permanente en la Reserva Forestal de Pasoh, ubicada en la Malasia peninsular (28589N, 1028179E). La reserva contiene 2.450 ha de bosque protegido de tierras bajas dominado por dipterocarpos, con unos 1.700 mm de precipitaciones al año. Todos los árboles y arbustos independientes que excedían 1 cm de diámetro a la altura del pecho fueron mapeados, medidos, y identified a la especie[17]. 814 especies en 290 géneros y 78 familias fueron identified, que comprenden un total de casi 340.000 tallos. Las familias más abundantes fueron Euphorbiaceae, Dipterocarpaceae y Annonaceae. Los censos se llevaron a cabo en 1986-1987, 1990, 1995 y 1995. 2000. En el censo de 1995, que se utilizó en este estudio, 637 especies estaban representadas por al menos 20 individuos en la parcela. No se consideraron las especies con menos individuos. Pudimo s asignar síndromes de dispersión a 561 de estas 637 especies. Asignación de síndromes de dispersión. Las especies dispersas por los animales son defined por tener partes comestibles que favorecen la deglución o el consumo. transporte de semillas durante o después de la alimentación de los vertebrados. Los dispersores de animales varían en anchura de abertura y otras características, afectando su elección de frutos[31] y las distancias a las que transportan las semillas[45]. Dependiendo del análisis, utilizamos el diámetro de la fruta en el eje más largo como una variable continua, o dividimos las especies dispersas en animales en tres grupos basados en el diámetro de la fruta (Tabla 1). Los paseriformes son los dispersores más probables de frutos de la clase más pequeña (2,2 cm de diámetro, a lo largo del eje más largo); las aves y los mamíferos pequeños y medianos son los dispersores más probables de frutos medianos (2-5 cm de diámetro); y PLoS Biología | www.plosbiology.org 2140 Noviembre 2006 | Volumen 4 | Número 11 | e344 Dispersión y Patrón Espacial Entre las especies no dispersas por los animales, se encuentran cuatro síndromes de dispersión: dispersión por viento, dispersión por giro, dispersión por gravedad y dispersión balística (Tabla 1). Las especies que tienen alas membranosas se dividen en dispersión por viento y giro, basándose en la relación entre el volumen de propágulos (cm3) y el área del ala (cm2), que es una estimación estándar de la carga del ala[34,46]. Las especies cuya carga alar inversa excede 100 veces el volumen de propágulos son classified como dispersas por el viento; las demás especies aladas son classified como dispersas por giro (Figura S1). Esta distinción se aproxima a la distancia de dispersión basada en la carga alar y el volumen del propágulo[46,47]. Todas las especies emplumadas en este estudio tienen pequeños propágulos y se encuentran dispersas por el viento. Las especies dispersas balísticamente poseen cápsulas explosivamente deshidratantes que lanzan las semillas a cierta distancia de la planta madre. Las especies dispersas por gravedad son defined como aquellas que carecen de cualquier mecanismo obvio de dispersión o recompensa por dispersión. Aunque muchas especies están sujetas a dispersión secundaria por los animales o el agua, para el propósito de estos análisis examinamos sólo la fase primaria de dispersión. Estimación del tamaño espacial de los cúmulos. Un proceso de conglomerados de Poisson consiste en centros de conglomerados localizados al azar; alrededor de cada centro de conglomerados, los árboles se colocan de acuerdo a una distribución gaussiana radialmente simétrica. Para cada especie, el modelo de conglomerado es defined por dos parámetros: q, la densidad de los centros de conglomerados, y r, donde 2r2 denota la distancia media cuadrada de un árbol desde el centro de su conglomerado. Para una distribución espacial observada de una especie, se eligieron q y r para mejorar la curva k de Ripley, como se describió anteriormente[5]. Cuarenta y seis especies exhiben distribuciones espaciales casi al azar, resultando en valores r mayores que el ancho de la parcela de 50 ha, o valores q de fitted mayores que la densidad de árboles. A estas especies se les asignó r ¼ 500 m, aunque la elección del límite superior de r no afecta a los resultados de nuestros análisis (no paramétricos). Los tamaños de los clusters de fitted r no son normales ni homosexuales, por lo que utilizamos métodos no paramétricos en todos nuestros análisis estadísticos. Regresión filogenética. Las relaciones filogenéticas entre las 561 especies estudiadas se estimaron utilizando la base de datos Phylomatic y las herramientas de ensamblaje[48]. Utilizamos el reciente árbol de angiospermas de Soltis et al.[49], al que se han adherido árboles de consenso estricto. El programa Phylomatic utiliza este árbol esqueleto para construir un árbol completo de taxones de entrada, uniendo especies no reconocidas a un nodo de género politómico y géneros no reconocidos a un nodo de familia politómica. La filogenia utilizada aquí representa, por lo tanto, la mejor estimación de la relación de los taxones basada en un gran número de estudios de datos moleculares y morfología, con sus clados terminales siendo sus consensos estrictos. Una versión NEXUS de nuestro árbol taxonómico 561 está disponible bajo petición. Se realizaron contrastes filogenéticamente independientes según el método de Pagel[25] implementado en el análisis comparativo por el programa de contrastes independientes[50]. Los grados de libertad para las politomías no resueltas se calcularon según el método de Purvis[51], resultando en pruebas conservadoras de la hipótesis evolutiva. En todos los casos, la variable dependiente fue el tamaño del conglomerado r. Los contrastes continuos se calcularon utilizando la longitud del fruto como variable independiente. Los contrastes binarios se calcularon utilizando pares de categorías de dispersión. En lugar de calcular los contrastes para todos los pares de síndromes de dispersión, comparamos grupos de síndromes; por ejemplo, ''animal- dispersado'' versus ' 'no animal-dispersado''. ’’ Datos del ICB. También estudiamos especies arbóreas dentro de la parcela de 50 hectáreas de dinámica forestal permanente en el ICB[2729], censada en 1995. Nuestro análisis incluye 209 especies representadas por al menos 20 individuos en la parcela, cuyos síndromes de dispersión han sido caracterizados recientemente[52,53]. Los síndromes de dispersión de las especies de ICB son classified en las categorías balística, de viento, de pájaro/murciélago o de mamífero. (El mamífero tiene prioridad en los casos de mecanismos múltiples, aunque esta elección no afecta a nuestros resultados.) Estos classifications no incluyen información sobre el diámetro del fruto o la carga alar. según la relación entre la elevación producida por el tamaño relativo de las alas y el tamaño absoluto del propágulo. El eje y estima la carga recíproca de las alas. Nosotros define Dispersión por giro (triángulos sólidos invertidos) como y , 100x (elevación débil y un propágulo grande), y dispersión por viento (triángulos abiertos) cuando y... 100x (elevación fuerte y un pequeño propágulo; línea defines y ¼ 100x). Varias especies con penachos o penachos en lugar de alas no fueron incluidas en este análisis y fueron categorizadas como dispersas por el viento. Información de apoyo Figura S1. Área de Ala y Volumen de Propágulo para Tropical Alado Especies arbóreas El volumen se utiliza aquí como un proxy para la masa. Las especies con propágulos alados son classified en forma de viento o giro-dispersados PLoS Biología | www.plosbiology.org 2141 Noviembre 2006 | Volumen 4 | Número 11 | e344 Dispersión y Patrón Espacial DOI: 10.1371/journal.pbio.0040344.sg001 (456 KB EPS). Figura S2. El tamaño espacial del conglomerado r no está correlacionado con la estatura de los árboles en todas las especies estudiadas (p ¼ 0.9146) DOI: 10.1371/journal.pbio.0040344.sg002 (585 KB EPS). Figura S3. El tamaño de los grupos espaciales r no está correlacionado con la estatura de los árboles entre especies dispersas por mecanismos bióticos o abióticos (Dispersión biótica, p ¼ 0.5098; Dispersión abiótica, p ¼ 0.2324). DOI: 10.1371/journal.pbio.0040344.sg003 (541 KB EPS). Agradecimientos Forest y el Smithsonian Institution's Center for Tropical Forest Science para acceder a los conjuntos de datos de Pasoh y BCI. Agradecemos a Joe Wright por compartir datos sobre los modos de dispersión en BCI. Estamos en deuda con Peter Ashton y William Bossert por sus aportes conceptuales. También agradecemos a Andy Purvis, Ran Nathan, Charles Godfray, Mark Rees, Jack Putz, Kristina Stinson y dos revisores anónimos por sus comentarios sobre el manuscrito. Contribuciones de los autores. TGS y JBP concibieron y diseñaron los experimentos. TGS y JBP realizaron los experimentos. TGS y JBP analizaron los datos. TGS y JBP escribieron el artículo. Financiación. TGS fue apoyado en parte por la Conservation, Food and Health Foundation; JBP fue apoyado en parte por el Burroughs Wellcome Fund y el William F. Milton Fund. Intereses en pugna. Los autores han declarado que no existen intereses opuestos. Agradecemos a Abdul-Razak Mohammed Ali, de Forest Research Instituto de Malasia para obtener permiso para recolectar especímenes en Pasoh Referencias 1. Hubbell SP (1997) A unified theory of biogeography and relative species abundance and its application to tropical rain forests and coral reefs. Arrecifes de Coral 16: S9-S21. 2. Hubbell SP, Foster RB (1986) Biología, oportunidad e historia y estructura de las comunidades de árboles de la selva tropical. In: Cody ML, Diamond J, editores. Ecología comunitaria. Nueva York: Harper & Row. pp. 314–329. 3. Chave J, Muller-Landau HC, Levin SA (2002) Comparación de modelos comunitarios clásicos: Consecuencias teóricas para los patrones de diversidad. Am Nat 159: 1-23. 4. Hubbell SP (1979) Dispersión, abundancia y diversidad de árboles en un bosque tropical seco. Science 203: 1299-1309. 5. Plotkin JB, Potts MD, Leslie N, Manokaran N, LaFrankie J, et al (2000) Speciesarea curves, spatial aggregation, and habitat specialization in tropical forests. J Theor Biol 207: 81-99. 6. Condit R, Ashton PS, Baker P, Bunyavejchewin S, Gunatilleke S, et al (2000) Spatial patterns in the distribution of tropical tree species. Ciencia 288: 1414–1418. 7. Levine JM, Murrell DJ (2003) The community-level consequences of seed dispersal patterns. Annu Rev Ecol Evol Syst 34: 549-574. 8. Harms KE, Condit R, Hubbell SP, Foster RB (2001) Asociaciones de hábitats de árboles y arbustos en una parcela de bosque neotropical de 50 ha. J Ecol 89: 947959. 9. Nathan R., Muller-Landau HC (2000) Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Tendencias Ecol Evol 15: 278–285. 10. Hubbell SP (2001) The unified neutral theory of biodiversity and biogeography. In: Levin SA, Horn HS, editores. Monografías en biología de poblaciones. Princeton (Nueva Jersey): Princeton University Press. 448 p. 11. Dalling JW, Muller-Landau HC, Wright SJ, Hubbell SP (2002) Papel de la dispersión en la limitación del reclutamiento de especies pioneras neotropicales. J Ecol 90: 714-727. 12. Schupp EW, Fuentes M (1995) Spatial patterns of seed dispersal and the unification of plant population ecology. Ecoscience 2: 267-275. 13. Wang BC, Smith TB (2002) Closing the seed dispersal loop. Tendencias Ecol Evol 17: 379-385. 14. Webb CO, Peart DR (2000) Asociaciones de hábitats de árboles y plántulas en una Bosque lluvioso de Borneo. J Ecol 88: 464-478. 15. Janzen DH (1970) Herbívoros y el número de especies arbóreas en los bosques tropicales. Am Nat 104: 501-528. 16. Connell JH (1971) On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. In: Den Boer PJ, Gradwell GR, editores. Dinámica de las poblaciones: Actas del instituto de estudios avanzados sobre la dinámica del número de poblaciones. Wageningen (Países Bajos): Center for Agricultural Publishing and Documentation. pp. 298-310. 17. Manokaran N, LaFrankie JV, Kochummen KM, Quah ES, Klahn J, et al (1990) Methodology for the fifty hectare research plot at Pasoh Forest Reserve. Kepong (Malasia): Instituto de Investigación Forestal de Malasia. 69 p. 18. Janson CH (1983) Adaptación de la morfología de las frutas a los agentes de dispersión en un bosque neotropical. Science 219: 187-189. 19. Janson CH (1992) Measuring evolutionary constraints: Un modelo de Markov para las transiciones filogenéticas entre síndromes de dispersión de semillas. Evolución 46: 136–158. 20. Whitmore TC (1972) Tree flora de Malaya: Un manual para silvicultores. Registro forestal malayo no. 26. Londres: Longman. 471 p. 21. Symington CF (1974) Manual de dipterocarpos de Foresters. Kuala Lumpur (Malasia): Universidad Penerbit de Malaya. 520 p. 22. Ng FSP (1978) Tree flora de Malaya, Volumen 3. Kuala Lumpur (Malasia): Longman. 339 p. PLoS Biología | www.plosbiology.org 2142 23. Ng FSP (1989) Tree flora de Malasia, Volumen 4. Kuala Lumpur (Malasia): Longman. 549 p. 24. Ng FSP (1991) Manual de frutos, semillas y plántulas forestales. Registro forestal malayo no. 34. Kepong (Malasia): Instituto de Investigación Forestal de Malasia. 996 p. 25. Pagel MD (1992) Un método para el análisis de datos comparativos. J Teoría Biol 156: 431–442. Noviembre 2006 | Volumen 4 | Número 11 | e344 Dispersión y Patrón Espacial 26. Ripley BD (1976) The second order analysis of stationary processes. J Aplicar Prob 13: 255-266. 27. Condit R (1998) Parcelas para el censo de bosques tropicales: Métodos y resultados de la Isla de Barro Colorado, Panamá, y una comparación con otras parcelas. Berlín: Springer Verlag. 211 p. 28. Hubbell SP, Foster RB, O'Brien ST, Harms KE, Condit R, et al. (1999) Lightperturbaciones de la brecha, limitación del reclutamiento y diversidad de árboles en un bosque neo-tropical. Science 283: 554-557. 29. Condit R, Hubbell SP, Foster RB (2005) Barro Colorado forest census plot data. Disponible: http://ctfs.si.edu/datasets/bci. Consultado el 6 de septiembre de 2006. 30. Augspurger CK, Kitajima K (1992) Estudios experimentales de plántulas reclutamiento a partir de distribuciones contrastantes de semillas. Ecology 73: 1270-1284. 31. Wheelwright NT (1985) Tamaño de la fruta, anchura de la abertura, y las dietas de las aves que comen fruta. Ecology 66: 808-818. 32. Kelt DA, Van Vuren DH (2001) The ecology and macroecology of área de distribución de los mamíferos. Soy Nat 157: 637–645. 33. Nathan R., Katul GG (2005) El desprendimiento de hojas en los bosques caducifolios eleva la dispersión de semillas a larga distancia por el viento. Proc Natl Acad Sci U.S.A. 102: 8251-8256. 34. Suzuki E, Ashton PS (1996) Relación de tamaño de sépalo y nuez de frutas de Asia. Dipterocarpaceae y sus implicaciones para la dispersión. J Trop Ecol 12: 853870. 35. Brewer SW, Rejmanek M (1999) Small roents as significant dispersers of tree seeds in a neotropical forest. J Veg Sci 10: 165-174. 36. Yasuda M, Miura S, Nor Azman H (2000) Evidence for food hoarding comportamiento en roedores terrestres en la Reserva Forestal de Pasoh, un bosque lluvioso de tierras bajas de Malasia. J Trop For Sci 12: 164-173. 37. Vander Wall SB, Kuhn KM, Beck MJ (2005) Seed removal, seed predation, and secondary dispersal. Ecology 86: 801-806. 38. Olvidar PM (1990) Dispersión de semillas de Vouacapoua americana (Caesalpiniaceae) por roedores caviomorfos en la Guayana Francesa. J Trop Ecol 6: 459-468. 39. Ashton PS, Givnish TJ, Appanah S (1988). Dipterocarpaceae: New insights into floral induction and the evolution of mast fruiting in the aseasonal tropics. Am Nat 132: 44-66. 40. Janzen DH (1974) Tropical blackwater rivers, animals and mast fruiting by the Dipterocarpaceae. Biotropica 6: 69-103. 41. Curran LM, Leighton M (2000) Respuestas de los vertebrados al espacio temporal variación en la producción de semillas de Dipterocarpaceae fructíferas en mástil. Ecol Mono 70: 101–128. 42. Uriarte M, Condit R, Canham CD, Hubbell SP (2004) Un modelo espacialmente explícito de crecimiento de árboles jóvenes en un bosque tropical: ¿Importa la identidad de los vecinos? J Ecol 92: 348-360. 43. Levins R, Culver D (1971) Convivencia regional y competencia entre especies raras. Proc Natl Acad Sci U.S.A. 68: 1246-1248. 44. Tilman D (1994) Competition and biodiversity in spatially structured habitats. Ecología 75: 2-16. 45. Schupp EW (1993) Cantidad, calidad y eficacia de la dispersión de semillas por los animales. Vegetatio 108: 15-29. 46. Augspurger CK (1986) Morfología y potencial de dispersión del viento. diásporas dispersas de árboles neotropicales. Am J Bot 73: 353-363. 47. Greene DF, Johnson EA (1993) Masa de semillas y capacidad de dispersión en diásporas dispersas por el viento. Oikos 67: 69-74. 48. Webb CO, Donoghue MJ (2002) Phylomatic. Revisión de la base de datos L20020928. 49. Soltis DE, Soltis PS, Chase MW, Mort ME, Albach DC, et al (2000) Filogenia angiospermica inferida a partir de secuencias de 18S rDNA, rbcL y atpB. Bot J Linnean Soc 133: 381-461. 50. Purvis A, Rambaut A (1995) Análisis comparativo por contrastes independientes (CAIC): Una aplicación de Apple-Macintosh para analizar datos comparativos. Comp Appl Biosci 11: 247-251. 51. Purvis A, Garland T (1993) Politomías en análisis comparativos de caracteres continuos. Syst Biol 42: 569-575. 52. Muller-Landau HC, Hardesty BD (2005) Dispersión de semillas de plantas leñosas en bosques tropicales: Conceptos, ejemplos y direcciones futuras. In: Bolsas DFRP, Pinard MA, Hartley S, editores. Interacciones bióticas en los trópicos. Cambridge: Cambridge University Press. 580 p. 53. Wright SJ, Hernandez A, Condit R (2007) La cosecha de carne de monte altera bancos de plántulas favoreciendo a las lianas, semillas grandes y semillas dispersas por los murciélagos, las aves y el viento. Biotropica: En prensa. PLoS Biología | www.plosbiology.org 2143 Noviembre 2006 | Volumen 4 | Número 11 | e344