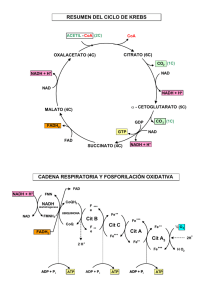
Resumen-met-glucidico-y-lipidico... Guadip Bioquímica y Biología Molecular 2º Medicina Facultad de Ciencias Médicas Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. UNLP, Facultad de Ciencias Médicas Cursada 2022 Guadalupe Palumbo Bioquímica Metabolismo de glúcidos P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 Glucolisis . . ● Vía catabólica en el citosol de todas las células; oxidación de glucosa en 10 pasos secuenciales que dan lugar a la . formación de 4 moléculas de ATP y 2 moléculas de NADH con la previa inversión de 2 ATP en la etapa de pérdida . de esta vía. Reacciones reguladas: muy exergónicas y por lo tanto irreversibles! ○ Glucosa + ATP → Glucosa-6P + ADP | glucoquinasa - hexoquinasas ○ Fructosa -6P + ATP → Fructosa 1,6 P 2 + ADP | PFK-1 ○ PEP → piruvato + ATP | Piruvato quinasa ● Regulaciones ○ Hexoquinasas: inhibidas alostéricamente Glucosa 6P ○ Glucoquinasa: ■ Inhibida indirectamente por Fructosa 6P que esta en igual [Glucosa 6P] y promueve la compartimentalización de la enzima hacia el núcleo por una proteína reguladora. ■ Activada La glucosa compite con la fructosa para la translocación de la enzima hacia el citosol, altas concentraciones de glucosa permiten que la enzima esté activa en este compartimento fomentando su disociación de la proteína reguladora. ○ PFK-1 ■ Regulación energética ● Inhibida por ATP y citrato Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. Resúmen para final! ● ■ Estimulada por AMP Regulación por metabolito ( es la reg más potente!) → Modulada positivamente por la Fructosa 2,6 P2 ● Fosforilación de F 6P | Enzima bifuncional, desfosforilada por proteínas Pasas està activa su función quinasa, fosforilada por disminuye la [F 2,6 P2] y la enzima es menos activa). ○ Piruvato quinasa: ■ Regulación alostérica: modulado + por Fructosa 1,6 P2 ■ Regulación covalente en hígado: ■ ● Inhibida por fosforilación por PKA (glucagón) ● Activa desfosforilada por proteínas Pasas (insulina) Regulación covalente en músculo: ● activa fosforilada por PKA vía adrenalina! P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 Fase de inversión: Glucosa + ATP → glucoquinasa→ Glucosa 6 P + ADP Se invierten 2 ATP! Fructosa 6P + ATP →PFK-1→ Fructosa 1,6 biP + ADP Fase de recuperación: 2. (1,3 Bifosfoglicerato + ADP) →fosfoglicerato quinasa ← 2.(3 P glicerato + ATP) Se generan 4 ATP por fosforilación 2. (PEP + ADP) → piruvato quinasa→ 2. (piruvato + ATP) a nivel del sustrato! Glucolisis entera …… …………………... Glucosa → Glucosa 6P → hexosas P isomerasa← Fructosa 6P →PFK 1→ F 1,6 P2 → aldolasa← DHAP + GA 3 P DHAP →triosa P isomerasa← GA 3P 2(GA-3 P) + 2 Pi + 2 NAD+→GA 3 P DH← 2( 1,3- P 2 glicerato) +2 NADH + 2 ADP → fosfoglicerato quinasa ← 2(3-P glicerato) + 2 ATP → glicerato mutasa← 2.(2 -P glicerato) →enolasa← 2 PEP + 2 ADP → piruvato → 2 Piruvato + 2 ATP quinasa Fermentación láctica Vía citosólica. Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. PKA esta se inactiva y se reactiva el dominio fosfatasa ( así Los eritrocitos tienen como única fuente de energía (ATP) la glucólisis al no presentar sistemas de endomembrambranas -orgánulos- incluídos la mitocondrias, donde ocurre la respiración celular mediante fosforilación oxidativa a través de un sistema de transporte de electrones, que en los demás tipos celulares es la principal fuente de energía (genera entre 30 y 32 ATP). Es por esto que los eritrocitos, y el músculo en contracción vigorosa e hipoxia, presentan una glucólisis para esto es necesario reoxidar el NADH a NAD+ para que pueda ser reutilizado constantemente en la reacción catalizada por la GA 3P DH, que en otros tejidos se logra por un sistema de lanzadera hacia la mitocondria; para esto los glóbulos rojos y los miocitos llevan a cabo la fermentación láctica, donde el piruvato se reduce a lactato, reoxidado así el NADH NAD+. Piruvato + NADH→ Lactato Deshidrogenasa← Lactato + NAD+ En diferentes situaciones metabólicas el exceso de lactato genera acidosis láctica, esto también sucede en el cáncer donde las células neoplásicas crecen en situaciones de hipoxia, abasteciendo sus necesidades energéticas por una alta glucólisis y acidificando su microambiente para que su proliferación anómala sea favorable! ➔ Formación de acetil-Coa| Complejo Piruvato Deshidrogenasa (PDH)..... P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 ◆ PDH: complejo multienzimático ubicado en la mitocondria! ● E1: piruvato descarboxilasa |TPP cofactor ● E2: dihidrolipoil transacetilasa ● E3: dihidrolipoil DH ◆ Descarboxilación oxidativa del piruvato (3C) a Acetil-CoA (2C) ● 1) Descarboxilación del piruvato en E1 unido a tiamina pirofosfato (TPP) ● 2) Oxidación del intermediario por trasferencia al grupo S-S del ácido lipoico en E2! ● 3) transferencia del grupo acetilo que se encontraba unido como tioéster del ac lipoico a una Coenzima A ● 4) oxidación del ácido lipoico por reducción de FAD en E3, y trasferencia del poder reductor del FADH a un NADH → cadena mitocondrial electrones!, de esta manera se reoxidó todo lo que había reducido para un nuevo ciclo de producción de acetil CoA ➔ Regulación: ◆ COVALENTE ● PDH quinasa la fosforila e inactiva. Esta enzima es modulada alostéricamente ● ○ +: Sus productos → Acetil Coa, NADH, ATP ○ - : su sustrato: Piruvato. PDH fosfatasa: desfosforila la E1 y activa la enzima. ○ +: Ca2 ◆ ALOSTÉRICAMENTE Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. elevada para suplir sus necesidades energéticas, ya que esta vía genera netamente sólo dos ATP, ● Inhibido por sus productos : NADH y Ac CoA. Gluconeogénesis! Algunos tejidos como el cerebro, los eritrocitos, testículos médula renal dependen de un suministro constante de glucosa, e spor esto que a glucemia (concentración sanguínea de sangre) está altamente regulado por el hígado, principalmente, y los riñones. LA glucemia debe mantenerse en un rango entre 90 mg/dl y 60 mg/dl En situación de ayuno el primer suministro de glucosa parte de la degradación de glucógeno hepático, sin embargo una vez este se termina comienza la formación de glucosa a partir de precursores diferentes a los hidratos de carbono, precursores gluconeogénicos! P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 Los principales precursores son el lactato, piruvato, los alfa ceto ácidos como el alfa kg proveniente de la degradación de prot tisulares, y el glicerol, otorgado por la degradación de TAG en tej adiposo (ingresa al hígado transportado por sangre) ● Glicerol: proviene de la degradación de TAG en tejido adiposos y lleva vía sangre a los hepatocitos donde es fosforilado por la glicerol quinasa a G3P que pasa por la G3P DH para oxidarse generando NADH y convertirse en DHAP (intermediario de la gluconeogénesis) ○ 👉Recordar que la degradación de TAG y obtener AG que ingresen en Beta oxidación es necesaria para generar energía y poder llevar a cabo la gluconeogénesis, que es un proceso sumamente costoso al utilizar 6 enlaces de alta energía y 2 NADH. ○ 💡además como está muy activa la B-ox se comienzan a acumular Acetil CoA en la mitocondria, inhibiendo a la PDH (para que no se use el piruvato) y activado la Piruvato carboxilasa para formar OAA, de esta manera el Acetil CoA tampoco puede avanzar por el CK pq el oxalacetato del mismo está desplazado a la formación de PEP y no queda disponible para ser sustrato de la citrato sintasa! Por todo esto el hígado decide aprovechar este exceso de Ac CoA para generar cuerpos cetónicos, una fuente alternativa de energía para tejidos extrahepáticos menos para el eritrocito y el hígado (éste no tiene la B-cetoacil transferasa para utilizarlos). ● Lactato: proviene del músculo en ejercicio y de los eritrocitos. Se genera el Ciclo de Cori → Se degrada la glucosa a piruvato, y éste se reduce a lactato por la lactato DH oxidando NADH a NAD+; el lactato pasa a sangre hacia el hígado y es oxidado a piruvato (produciendo NADH) quien es sustrato de la piruvato Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. Sólo funciona en el hígado! → regulador de la glucemia! carboxilasa para generar OAA, para generar glucosa que pasa al torrente sanguíneo que nuevamente es captado por el músculo y el eritrocito. ● Aminoácidos: el catabolismo de prot tisulares en ayuno dan lugar alfa cetoácidos con el alfa kg, quien puede ingresar al CK y desviarse a la formación de OAA. kg generando Glutamato y Piruvato → TGP: transaminasa Glutamato-Piruvato ⚕️importancia clínica: presencia en sangre sirve para determinar daño hepático, de localización citosólica. ○ Aspartato se transamina con alfa kg a Glutamato y OAA → TGO: transaminasa Glutamato- Oxalacetato ⚕️ importancia clínica: presencia en sangre revela daño hepático grave, localización mitocondrial y citosólica ■ 🩸 Índice de Ritis: TGO/ TGP Enzimas gluconeogénicas- reacciones de rodeo (reguladas) P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 ➔ Piruvato carboxilasa - mitocondrial piruvato + HCO3 + ATP → oxalacetato + ADP ➢ Regulación alostérica ○ Inhibida por ADP ○ Activada por Acetil Coa de Box ➔ PEP carboxiquinasa - mitocondrial y citosólica Oxalacetato + ADP → PEP + CO2 + ADP ➢ Regulación alostérica ○ Inhibida por ADP ○ Activada por AMPc (vía glucagón) ➔ Fructosa 1,6 BiFosfatasa - citosólica Fructosa 1,6 P2 → Fructosa 6 P + Pi ➢ Regulación- Alostérica ○ Inhibida por Fructosa 2,6 BiP, ADP ○ Activada por citrato ➔ Glucosa 6 Fosfatasa - en el Re de hepatocitos, cara luminal. Glucosa 6P → Glucosa + Pi Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. ○ la Ala es un precursor gluconeogénico al transferir su grupo NH2 al alfa Algunas personas solo expresan la PEP carboxiquinasa citosólica y para esto deben transportar el OAA hacia este compartimento, pero éste no tiene un transportador específico para salir de mitocondrias por lo que se reduce a malato, quien sale a citosol y se reoxida a OAA aportando un NADH que luego es utilizado por la Gliceraldehído Ciclo de Krebs! Ciclo Anfibólico → según las necesidades celulares actúa como vía catabólica hacia la producción de NADH , FADH y GTP oxidando a 2 (CO2) Acetil CoA o como vía anabólica cediendo sus intermediarios para la producción de nuevas moléculas! Los intermediarios luego son repuestos a través de reacciones anapleróticas P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 Piruvato + HCO3 ·+ ATP←→ oxalacetato + ADP + Pi | Pir carboxilasa (mitocondrial) Oxalacetato + GTP ←→ PEP + GDP + HCO3 | PEP carboxiquinasa (mitocondrial y citosólico) Piruvato + NADPH + HCO3 ← → Malato + NADP + H2O | enzima málica ➔ Regulación: Alostérica: por carga energética! ➔ Citrato Sintasa ◆ - : Citrato, succinil Coa, NADH, ATP ◆ +: ADP ★ en hígado y tej adiposos la citrato sintasa no se inhibe alostéricamente por ATP ya que se busca la sobreproducción de citrato para que este pueda salir de la mit hacía citosol y así transportar Acetil Coa para que puedan ser sustrato de la Acetil Coa carboxilasa que forma MAlonyl Coa → síntesis de AG ➔ Isocitrato DH: ◆ -: NADH, ATP ◆ +: ADP, Ca+2 ➔ Alfa kg DH: ◆ - NADH , ATP, succinil Coa ◆ + Ca+2 Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. 3P DH para reducir el 1,3 BiP glicerato a GA 3P. 1. Se condensa el Ac coa con el oxalacetato por enz citrato sintasa se forma citrato 2. citrato se deshidrata y rehidrata isomerizándose por la enzima aconitasa a isocitrato 3. isocitrato se descarboxila oxidativamente por la Isocit DH liberando CO2 formando alfa ketoglutarato → NADH 4. alfa kg pasa por la alfa kg DH y por otra descarboxilación oxidativa forma succinil coa utilizando una SH- CoA, liberando CO2 → NADH succinil coa pasa por la succinil coa sintetasa generando GTP por fosforilación nivel del sustrato y liberando CoA, dando lugar al succinato 6. succinato pasa por la succinato DH dando fumarato y FADH 7. fumarato se hidrata por la enzima fumarasa dando malato 8. malato se oxida a oxalacetato → NADH Lanzaderas de poder reductor! → transportan el poder reductor de NADH citosólico, reoxidándolos, a NADH mitocondrial para que pasen a la cadena transportadora de electrones. P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 👉 ¿Pq son importantes ? es necesario re-oxidar el NADH producido en la glucólisis en la reacción catalizada por la GA 3P DH donde Gliceraldehído 3 P + Pi + NAD → 1,3 BiP glicerato + NADH, para que esté disponible constantemente y poder llevar la glucólisis a cabo! LANZADERA MALATO- ASPARTATO hígado, corazón Oxalacetato en mitocondria se transamina gracias al glutamato a aspartato quien sale de la mitocondria hacia el citosol, donde transfiere su grupo NH2 a un alfa kg (generando glutamato), de manera que el oxalacetato ahora en citosol utiliza un NADH, reoxidandolo, para reducirse a malato quien vuelve a ingresar a la mitocondria por transportadores específicos en la membrana mit interna, una vez dentro se oxida por la malato DH nuevamente a oxalacetato transfiriendo estos electrones a un NAD mitocondrial → cadena transportadora de electrones para reoxidarse. El oxalacetato utiliza un glutamato que le transfiere el grupo NH2 para formar aspartato y nuevamente repetir el ciclo. LANZADERA DHAP - G3P musc esq y cerebro DHAP se reduce a Glicerol 3 P oxidando un NADH citosólico por la G3P DH citosólica; este G3P pasa a mitocondria y nuevamente es sustrato de la G3P DH mitocondrial oxidándose a DHAP para transferir el poder reductor a un FAD→ FADH2 transfiere electrones a ubiquinona quien los transporta al complejo II. Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. 5. DHAP sale a citosol y se repite el ciclo. Metabolismo del glucógeno Glucógeno: polisacárido ramificado formado por unidades de glucosas unidas en alfa 1,4 y almacena en gránulos intracelulares mayormente en hígado y músculo esquelético como fuente de glucosa instantánea para suplir demandas energéticas - músculo- y regular la glucemia basal -hígado-. ¿Por qué se almacena la glucosa excedente en forma de glucógeno y no como glucosa en sí misma? La glucosa es osmóticamente activa, por lo que almacenarla significaria la lisis celular, además de que escaparía por los glut si no es fosforilada. El glucógeno resuelve los problemas osmóticos de esta molécula, sin embargo ocupa mucho lugar intracelularmente, por lo que su almacenamiento es limitado (por esto también se guarda energía en forma de TAG, pero éste no es fuente de glucosa ni precursores gluconeogénicos! no sirve para controlar la glucemia, pero P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 otorga energía para llevarse a cabo la gluconeogénesis en hígado) Síntesis de glucógeno: vía anabólica que requiere de ATP y UTP 1) Glucosa + ATP → Glucosa 6P | glucoquinasa/ hexoquinasa 2) Glucosa 6P → Glucosa 1P |Fosfoglucomutasa 3) Glucosa 1P + UTP → UDP-glucosa + PPi → 2 Pi | UDP glucosa pirofosforilasa 4) Glucogenina es una proteína autoglucosilante que genera un cebador de una pequeña cadena de glucosas utilizando UDP glucosa para que pueda actuar a continuación la glucógeno sintasa quien alarga la cadena en alfa 1,4 utilizando el mismo sustrato y libreando UDP (quien se fosforila a UTP nuevamente) 5) Glucógeno sintasa: Regulada Glucógeno n + UDP glucosa → Glucógeno n + 1 + UDP ➢ Regulación hormonal - covalente ○ Inactiva: Hígado: glucagón (hipoglucemia) y adrenalina (estrés fisiológico) = PKA fosforila. En músculo solo por adrenalina. ○ Activa: Hígado y músculo: insulina (hiperglucemia).; se desfosforila proteínas inhibitorias de PP 1 y esta desfosforila a GS ➢ Regulación alostérica: ○ +: Glucosa 6P 6) Enzima ramificante: retira una cadena pequeña de glucosas de un extremo no reductor y lo une nuevamente por medio de un enlace alfa 1,6. Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. ramificaciones en alfa 1,6, contiene muchos extremos no reductores y uno reductor. Se Degradación del glucógeno: aporta glucosa para glucemia (hígado) y para energía (musc) 1) Glucógeno fosforilasa: Regulada Glucógeno (n + 1) → Glucógeno (n -1) + Glucosa 1P | Fosforólisis de enlaces alfa 1,4 ○ Activa: Hígado: glucagón y adrenalina, músculo adrenalina→ PKA fosforila la Glucógeno fosforilasa quinasa y la activan, permitiendo que esta fosforila a GF ○ Inactiva: Insulina → PP 1 desfosforila todo, se activan fosfodiesterasa que hidrolizan AMPc ➢ Regulación alostérica ○ -: Glucosa 6P y ATP ○ + : Ca+ se une a subunidad Calmodulina de Glucogeno Fosforilasa Quinasa activandola | Hígado y músculo. ○ +: AMP| Músculo (tiene sentido pq en el músculo el glucógeno sólo aporta glucosa para obtener energía) P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 2) Enzima desramificante: transferasa y glucosidasa Transfiere residuos unidos por alfa 1,4 de una ramificación a un extremo no reductor para que actúe la glucógeno fosforilasa, y al residuo restante en enlace alfa 1,6 lo hidroliza liberando glucosa. Vía Pentosas Fosfato Ocurre en el citosol de todas las células, siendo más activa en tejidos con alta actividad biosintética de lípidos y eritrocitos. Consiste en una etapa irreversible y reguladora donde se da la oxidación y descarboxilación oxidativa de Glucosa 6P a ribulosa 5P generando 2 NADPH, y otra reversible no regulada de conversión de monosacáridos. → Fase oxidativa y reguladora ➢ Glucosa 6P DH, inhibida alostéricamente por altas [NADPH] Glucosa 6 P + NADP+ → 6P gluconato + NADPH ➢ 6P gluconato DH, descarboxilación oxidativa. 6P gluconato + NADP + → Ribulosa 5P + NADPH + CO2 Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. ➢ Regulación hormonal- covalente 👉 La ribulosa 5P puede isomerizarse a Ribosa 5P si la célula requiere sintetizar nucleótidos o epimerizarse a Xilulosa 5P para regular el metabolismo glucídico. → Fase no oxidativa ➢ Transaldolasas: transfieren unidades de 3 C ➢ Transcetolasas: transfieren unidades de 2 C Funciones de la vía de las PP: 1) Generar NADPH para síntesis de lípidos y regenerar glutatión | Fase ox 2) Síntesis de nucleótidos | Fase no ox 3) Generación de intermediarios glucolíticos y gluconeogénicos.| Fase no ox Integración con otras vías según necesidades celulares P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 ➔ Célula en división activa: requiere sintetizar nucleótidos ◆ La F6P y el GA3P de la glucólisis se interconvierten en ribosa 5P por las reacciones reversibles de la rama no oxidativa (transaldolasas y transcetolasas) ➔ Célula en división y crecimiento activo: requiere sintetizar nucleótidos y lípidos membranas- (NADPH) ◆ Fase oxidativa → 2 NADPH + Ribulosa 5P ◆ isomerasa → Ribosa 5P ➔ Célula con alta actividad biosintética de lípidos: requiere mucho NADPH ◆ FAse oxidativa muy activa junto con fase no oxidativa para dar F6P y GA3P como intermediarios gluconeogénicos y regenerar Glucosa 6P quien vuelve a ser sustrato de la fase oxidativa dando más NADPH ➔ Célula que requiere de NADPH y ATP ◆ Fase Ox y no oxidativa para que la F6P y el GA3P ingresen a glucólisis dando piruvato, ac coa, CK. Funciones del NADPH: - Biosíntesis de lípidos, detoxificación de xenobióticos , reducción del Glutatión - Las ROS como el H2O2 dañan proteínas, fosfolípidos y el ADN, por lo que las células tienen mecanismos para evitar que esto suceda, el Glutatión es un tripéptido (glutamil cistein glicina) que es capaz de reducir el H2O2 en 2 moléculas de agua → Glutatión peroxidasa Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. Interconversión de 3 (C5P) a dos hexosas (2 Fructosas 6P) y una triosa ( GA 3P) H2O2 + Glutatión reducido → 2 H2O + glutatión oxidado → Glutatión reductasa Glutatión oxidado + NADPH → Glutatión reducido + NADP+ da lugar a la anemia hemolítica ya que la célula no puede protegerse ante los radicales libres y demás ROS que terminan dañando su membrana plasmática (los glóbulos rojos pierden su núcleo en la diferenciación celular por esto es que no tienen recambio de proteínas). Metabolismo de lípidos Los lípidos son moléculas diversas que comparten en común la característica de ser hidrofóbicos, en las células forman parte de las membranas. P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 El 90% de los lípidos que consumimos son TAG, el 10% restante está conformado por colesterol, AG libres, etc; su absorción se da en el duodeno gracias a liberación de ácidos biliares desde la vesícula y lipasas desde el páncreas, quien también libera bicarbonato para neutralizar el quimo ácido. Con la llegada del quimo desde el estómago las células de la mucosa intestinal liberan secretina, estimulando la secreción de bicarbonato, y CKK, que promueve la liberación de las enzimas pancreáticas y la bilis. ● BILIS → se genera en el hígado y almacena en la vesícula, contiene sales y ácidos biliares –poseen una cara polar y otra apolar– encargadas de emulsionar las grasas evitando que coalescan y formen estructuras como gotas que dificultan la actividad de las lipasas. Las sales o ácidos biliares (son los mismos compuestos pero en diferentes estados de ionización, si tienen COOH → ácidos, y si tienen COO- → sales). Se generan a partir de colesterol. ● ENZIMAS PANCREÁTICAS → se liberan desde el páncreas exocrino junto con el bicarbonato (liberado por células intercalares del acino pancreático) por el conducto pancreático principal. 1) Lipasa pancreática: hidroliza enlace éster entre C 1 y 2 de la molécula del glicerol con los AG de un TAG liberando un 2-Monoacilglicérido + dos AG libres Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. ⚕️La carencia de Glucosa 6P DH (que cataliza la formación del primer NADPH) en eritrocitos pancreática 3) Colipasa: facilita la unión de la lipasa con la gota lipídica 4) Fosfolipasa A2: hidroliza el enlace éster entre AG y glicerol en posición 2 en un fosfolípido generando un lisofosfolípido P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 5) Colesterol esterasa: hidroliza ésteres de colesterol ➢ Ingreso de lipidos diatarios a las células ○ Una vez los lípidos son digeridos pasan al interior de los enterocitos en forma de micelas donde a nivel del REL van a servir para síntesis y resíntesis de TAAG, colesterol, etc. En el interior del enterocito se sintetiza también los quilomicrones una apolipoproteína encargada de transportar lípidos exógenos. → Quilomicrones :apolipoproteína, Apo B48, lípidos exógenos |palabras clave Se forman en los enterocitos y son exocitados hacia la linfa por el vaso quilífero de las vellosidades intestinales para pasar a sangre a nivel de la vena subclavia. Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. 2) Isomerasa: moviliza el AG en posición 2 del 2-MAG hacia el C3 para que actúe la lipasa Cuando recién son sintetizados presentan a Apo B48 (su apoproteína característica) y un núcleo repleto de TAG, resguardado por una cubierta polar de proteínas. Una vez alcanza el torrente sanguíneo recibe Apo E y Apo C 2 de las HDL. Apo C2 es un activador de la Lipoproteinlipasa (LPL) , enzima que hidroliza los TAG en AG y 👉La LPL es regulada a nivel de su expresión en T adiposo y muscular por estado nutricional y niveles de insulina. Cuando la insulina desciende por ayuno aumenta la expresión de la LPL en músculo (que utiliza los AG como energía) y baja en T ad Cuando aumenta la insulina en situación postprandial , aumenta exp en T Ad (para que este almacene los AG) y disminuye en músculo (utiliza glucosa). P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 A medida que los quilomicrones transcurren por los capilares la LPL va degradando su contenido, de manera que disminuye su tamaño y aumenta cada vez más su densidad. → Quilomicrón remanente: transfiere Apo C a HDL nuevamente y sufre una endocitosis mediada por receptores (que reconocen a la Apo E) en el hígado para ser degradado totalmente en lisosomas. Lipólisis| movilización de TAG en tejido adiposo por necesidad energética Sabemos que el exceso de energía proveniente del metabolismo de carbohidratos y proteínas se almacena en forma de grasa en los adipocitos para ser utilizada en próximas demandas energéticas. Es un proceso regulado hormonalmente por el glucagón que indica hipoglucemia y por lo tanto el hígado va a llevar a cabo la glucogenolisis y luego la gluconeogénesis, este último requiere de mucha energía que le va a ser aportada por la beta oxidación . ¿Pero como trascurre todo este proceso? ADIPOCITO| Glucagón (hipoglucemia) /adrenalia (estress) → receptor acoplato a Gs → adenilato ciclasa → AMPc → PKA PKA fosforila: Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. glicerol presente en el endotelio de los capilares. 1) PERILIPINAS, proteínas que recubren la gota lipídica impidiendo el contacto de las enzimas →, una vez fosforiladas sufren cambio conformacional , permitiendo el paso de las enzimas a la gota. 2) Lipasas sensibles a hormonas, que hidrolizan los TAG en AG libres (van por albúminas a diferentes tejidos para aportar energía como beta ox) y glicerol, que va hacia el hígado a) Glicerol + ATP→ glicerol quinasa → Glicerol 3P + ADP Glicerol 3P + NADH → glicerol 3P DH → DHAP + NAD+ Beta Oxidación| en mitocondrias de hígado, corazón, músculo esq, t ad. Oxidación del C beta de los AG librando 2 C por cada ciclo en forma de acetil CoA El paso limitante de la beta oxidación es el ingreso de los AG a mitocondria, una vez se encuentran en la matriz están destinados a beta oxidarse, por esto es que uno de las etapas reguladas a través del transportador CPT ! (acil carnitina transferasa 1); luego en la mitocondria P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 dos de las enzimas de esta vía también están moduladas alostéricamente por sus productos, la B-HidroxiacilCoA DH y la tiolasa. INGRESO A MITOCONDRIA 1º) Activación del AG | ➔ Acil Coa Sintetasa : se activa el grupo COOH del AG con ATP, liberando PPi, que es hidrolizado por una pirofosfatasa. Se obtiene R 1COOH- AMP que reacciona con grupo tiol (SH) de una SH-CoA (coenzima A ) desplazando el AMP, generando un Acil-Coa 2º) Transporte del Acil Coa a la mitocondria Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. por sangre como precursor gluconeogénico ➔ CPT1 : transfiere el grupo acilo del Acil Coa a una molécula de Carnitina liberando la CoA. La Acil-carnitina pasa a través de acilcarnitina translocasa hacia matriz mitocondrial ◆ Regulación| alostérica síntesis de AG) ➔ CPT2: intercambia acil carnitina por carnitina libre y acil Coa. La carnitina sale por la translocasa nuevamente para cumplir otro ciclo de transporte. Una vez el Acil-CoA ingresa a la mitocondria inmediatamente es atacado por las enzimas de la B Ox ➔ Acil-CoA DH: oxidación del acil-coa en carbono beta (se genera un doble enlace entre C alfa y beta) ◆ Acil-Coa + FAD+ → enoil-Coa + FADH ➔ Enoil- CoA hidratasa: se genera un grupo hidroxi en carbono B P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 ◆ Enoil-CoA + H2O → β-hidroxiacil-CoA ➔ β-hidroxiacil-CoA DH : oxidación del grupo hidroxi a ceto| REGULADA ◆ β-hidroxiacil-CoA + NAD+ → β-cetoacil-CoA + NADH ➢ Inhibida por alta concentración NADH respecto NAD+ ➔ Tiolasa: tiolisis, se escinde el carbono beta y alfa en forma de acetil-CoA ◆ β-cetoacil-CoA + SH-CoA → Acil-CoA (n-2) + Acetil-CoA ➢ Inhibida por altas [Acetil-CoA] (su producto) Síntesis de cuerpos cetónicos. En mitocondria de hígado y riñón por exceso… Cetogénesis| . de Acetil-Coa generado en B-ox (no puede ingresar a CK pq el OAA está .. . desviado hacia la gluconeogénesis) Puntos importantes ● Hígado y riñón generan cuerpos cetónicos pero no los utilizan por falta de una enzima (β-cetoacil CoA transferasa) ● Los cuerpos cetónicos son una manera de utilizar el Acetil CoA que se acumula en las mitocondrias de estos tejidos cuando está activa la gluconeogénesis ● Se basa en la condensación de moléculas de Acetil-CoA. ● Son fuentes de energía alternativa para músculos y otros tejidos no dependiente de glucosa (así no usan la poca glucosa en sangre de la que dependen los eritrocitos, cerebro y otros), proporcionan Acetil-CoA que ingresa en CK. ● El cerebro tiene como principal fuente de energía la glucosa, pero en caso de ayuno muy prolongado o inanición se adapta para utilizar estos cuerpos cetónicos y no morir. Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. ● Inhibida fuertemente por el Malonyl CoA (intermediario de ● En la diabetes no tratada los cuerpos cetónicos son prácticamente la fuente de energía para músculo y otros tejidos ya que no pueden usar glucosa ( problema en la secreción de insulina o en su vía e transducción) Se generan 3 cuerpos cetónicos: 1º) acetoacetato | 2º) β- hidroxibutirato| 3º) acetato Vía de síntesis ➔ Tiolasa ◆ 2 Acetil-CoA ←→ Acetoacetil-CoA + HS- CoA ➔ HMG CoA sintasa ◆ Acetoacetil-CoA + Acetil-CoA → HMG- CoA + HS- CoA ➔ HMG coa Liasa ◆ HMG-CoA → Acetil-CoA + Acetoacetato P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 ➔ 👉1º cuerpo cetónico! β- hidroxibutirato DesHidrogenasa ◆ Acetoacetato + NADH←→ NAD+ + β- hidroxibutirato ➔ Acetoacetato descarboxilasa 👉2º cuerpo cetónico! 👉3º cuerpo cetónico! ◆ Aceto acetato → acetato + CO2 Utilización de cuerpos cetónicos: reconversión a acetil-CoA 1) β- hidroxibutirato + NAD+→ β- hidroxibutirato DH 2) Acetoacetato + Succinil-CoA → ← Acetoacetato + NADH β-cetoacil CoA transferasa 3) Acetoacetil-CoA + HS-CoA → tiolasa → acetoacetil CoA + succinato ← 2 Acetil-CoA reductora en el citosol de todas las células, especialmente en . Síntesis de AG|vía … tejido adiposo, glándula mamaria e hígado, ante exceso de energía .. de glúcidos y aa → Puntos importantes ● Requiere de poder reductor → NADPH (se usa en la etapa de elongación con la ER en la reducción del doble enlace) , fuente de carbonos → Acetil-CoA (proviene de la activa glucólisis en hígado para disminuir glucemia y en demás tejidos donde la señal de insulina promueve la captación de glucosa como t adiposo) y energía! (que obvio tenemos pq estamos en una situación donde todas las necesidades energéticas ya están saciadas y queremos llevar a cabo procesos anabólicos para almacenar la energía sobrante) Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. ● ● El acetil-CoA se produce y acumula en mitocondria → esta vez la acumulación de esta molécula se da pq el CK está enlentecido ya que sus enzimas reguladas se encuentran disminuídas por carga energética (la isocitrato DH y la akg) ● La síntesis de AG tiene dos etapas: ○ Formación de Malonyl Coa por la ACC (Acetil CoA Carboxilasa), la enzima ○ Alargamiento de la cadena carbonada por la AG sintasa (que utiliza una vez acetil-CoA y luego alarga la cadena con Malonyl-CoA) ¿Cómo sale el Acetil-CoA de la | sistema de lanzadera Citrato: el hígado y tejido adiposo mitocondria hacia el citosol? .. | presentan una citrato sintasa que no se inhibe ante mucho ATP, pero si las otras enzimas reguladas del CK Para salir de la mitocondria el acetil coa presenta un sistema de lanzadera: citrato-OAA. → Acetil-CoA se condensa con OAA = citrato por la citrato sintasa! P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 👉 esta enzima por carga energética en otros tejidos se ve inhibida pero en hígado y tejido adiposo se presenta una isoforma donde esta reg no ocurre, permitiendo que se forme tanto citrato como acetil coA y OAA haya disponible y así poder salir a citosol por un transportador específico. →En citosol: se hidroliza por la Citrato Liasa en OAA y Acetil-CoA nuevamente utilizando ATP; el OAA se reduce a malato quien ingresa de nuevo a mitocondria y vuelve a oxidarse OAA (ingresa en otro ciclo de condensación con Acetil-CoA) o bien puede pasar por la Enzima Málica y oxidarse a piruvato generando NADPH! → esencial para la síntesis de AG (no es la principal vía de producción de NADPH como sí lo es la vía de las PP), el piruvato ingresa a mitocondria y es sustrato de la piruvato carboxilasa (activa por la cantidad de Ac CoA mitocondrial) para regenerar OAA. SÍNTESIS DE MALONIL-COA| ACC con actividad carboxilasa y transferasa usa biotina como cofactor y acetil-CoA como sustrato ………….. Etapa regulada de la vía, genera Malonyl-CoA, de 3 carbonos a partir de Acetil-CoA, de 2 carbonos. 🚨Malonyl-CoA es inhibidor de la CPT1 → coordinación entre síntesis y degradación! ➔ ACC → Acetil-Coa Carboxilasa: Carboxila el Acetil CoA transfiriendo un grupo Carboxilo (proveniente del HCO3 –bicarbonato–) que previamente fue activado por la enzima gastando ATP ◆ Acetil- CoA + HCO3 + ATP → Malonil-CoA + ADP + Pi Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. reguladora de toda esta vía → Regulación : Covalente ➢ Activa: Proteínas Pasas de vía Insulina ➢ Inactiva : FOSFORILADA por PKA (glucagón = hipoglucemia, ❓para qué AMPK → responde a baja carga energética ( si no me sobra energía?) ❓pq voy a queres sintetizar lípidos → Regulación : Alostérica ➢ Activa: citrato fomenta la polimerización de las cadenas polipéptidos de la enzima que inactiva es dimérica ➢ Inactiva :Acil-CoA de cadena larga (son sus productos) → Regulación : Expresión P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 ➢ Activa: dietas ricas en glúcidos ➢ Inactiva : Dietas ricas en AG poliinsaturados ALARGAMIENTO |Ciclos donde se alarga en 2 carbonos la cadena| AGS tiene 7 dominios diferentes donde 6 DE LA CADENA |tienen actividad catalítica y uno funciona como brazo flexible. Dos etapas: Una de carga y otra de elongación 1) CARGA de la proteína: sucede una vez por síntesis. a) Porción acetilo de Ac CoA se carga en ACP —porción trasp de acilos-– y luego MAT —porción Malonyl/ Ac CoA transferasa— lo transfiere a KS — beta ketoacil ACP sintasa– dejando nuevamente la ACP libre para que cargue malonilo (del malonil-coa) 2) ELONGACIÓN: en cada ciclo se condensa un malonilo (que esta en ACP) con el acilo cargado en KS y luego de las diferentes reacciones (donde los carbonos alcanzan todos los sitios activos gracias a estar unidos a ACP) la cadena se transfiere nuevamente a KS para que ACP cargue otro Malonilo y se repita el ciclo a) Condensación: el malonilo unido a ACP se descarboxila (perdiendo el carboxilo que le añadió la ACC) en simultáneo que el acetilo unido a KS se libera por ruptura del enlace tiol que los unía para condensarse con el ahora acilo-ACP —> se genera el β cetobutiril-ACP b) Reducción del grupo β-ceto: Actúa KR (reductasa) que lo reduce utilizando NADH Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. queremos síntesis de lípidos si el hígado necesita energía para gluconeo) y —> β hidroxibutiril-ACP c) Deshidratación por una DesHidratasa generando un enlace doble —> trans Δ2- butenoyl ACP → Butiril ACP El grupo butirilo es transferido a KS para que la ACP cargue otro malonilo y se repite todo. La ACP tiene actividad tioesterasa liberando así los AG Para obtener un palmitato se llevan a cabo 7 ciclos. SÍNTESIS DE TAG Y| Ambos tienen como precursor al Ac fosfatídico FOSFOLÍPIDOS P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 Síntesis de Ácido Fosfatídico está compuesto por una molécula de glicerol …. …………………………………………. esterificado con dos AG en sus C1 y C2 y un grupo Fosfato en su C3 ¿Pero cómo se sintetiza este intermediario? 1) Primero tenemos que obtener glicerol 3 fosfato! Hay tres fuentes de glicerol 3P, según la glucemia se sigue una u otra! ● Glicerogénesis (ayuno) ● Glucólisis (post prandial) ● Fosforilación del glicerol → Glicerogénesis: es la vía gluconeogénica pero desviada a la formación de Glicerol 3P. Tejido Adiposo e hígado. Piruvato + HCO3 + ATP→ piruvato carboxilasa→ OAA + ADP + Pi Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. d) Reducción del doble enlace por ER (reductasa) dependiente de NADPH OAA + GTP→ PEP carboxiquinasa→ PEP + GDP + CO2 PEP → → → → → DHAP + NADH → Glicerol 3P DH→ Glicerol 3 P + NAD+ → Glucolisis: tejido adiposo e hígado (recordar que aunque en otros tejidos por carga energética esta vía esta disminuida, en t ad e hígado continúa para generar Ac CoA p/ Glucosa → → → → DAHP DHAP + NADH → Glicerol 3P DH→ Glicerol 3P + NAD+ → Fosforilación del Glicerol: sólo en HÍGADO! es el único que presenta la glicerol quinasa 👉viene de B ox por sangre!) + ATP → Glicerol ( → Glicerol 3P Glicerol quinasa 2) Luego actúan unas aciltransferasas que tal cual su nombre transfieren acil-CoA en los P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 C1 y 2 del G3P a) Glicerol 3P+ Acil-CoA → Glicerol 3P aciltransferasa → Lisofosfatidato + CoA b) Lisofosfatidato + Acil-CoA → Acilglicerol 3P aciltransferasa → Ac Fosfatídico Síntesis de TAG Se elimina el grupo fosfato x fosfatasa específica y el C3 libre se esterifica con otro ……………………. Acil-CoA nuevamente gracias a una acil transferasa. CICLO TRIACILGLICEROL ¿Por qué se forma triacilglicerol en situación de ayuno si es un lípido asociado a la reserva energética? Por el ciclo del triacilglicerol: todo el tiempo hay síntesis y degradación de TAG, hay lipolisis y alrededor del 75% de AG generados por la lipolisis son reesterificados a TAG. Parte de esta resíntesis se da en el TA y otra en el hígado. Gasta energía degradando y sintetizando TAG, ¿es un ciclo fútil? SI, a primera vista es pointless. Pero sirve para tener una reserva energética extra en la sangre de rápido acceso en forma de AG o TAG unidos a VLDL. Síntesis de Fosfolípidos | Glicerofosfolípidos Se hidroliza el grupo P del Ac fosfatídico dando lugar a un DAG (diacilglicerol), y se activa un OH con CTP para poder unir la cabeza polar con la molecula de DAG y así formar el fosfoglicérido! Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. síntesis de AG y TAG —> almacenar energía) 👉 Si el OH que se activa con CTP pertenece a la molécula de DAG se trata de la estrategia 1 : eucariotas y procariotas 👉Si el OH pertenece a la cabeza polar se trata de la estrategia 2: sólo eucariotas ● Se hidroliza el Ac Fosfatídico y se lo condensa con CTP quedando DAG-CDP liberando PPi ● DAG-CDP reacciona con Glicerol 3P quien desplaza el CMP → Fosfatidil glicerol 3P ● Fosfatidil Glicerol 3P → fosfatasa→ Fosfatidilglicerol ● Fosfatidilglicerol + DAG-CDP → Cardiolipina + CMP ● DAG-CDP + inositol → Fosfatidilinositol + CDP Estrategia 2 P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 Síntesis de fosfatidilcolina y fosfatidiletanolamina Colina + ATP → quinasa→ Fosfocolina + ADP Fsfocolina + CTP → transferasa→ CDP-colina + PPi | alcohol activado CDP-colina + DAG → CMP + Fosfatidilcolina básicamente se fosforila la colina o etanolamina y lego el fosfato se desplaza por adición de CDP (q viene de CTP) y luego éste es desplazado por un DAG! 👉SIntesis de Fcolina en Hígado: a partir de serina el hígado exporta gran cantida de fosfatidilcolina hacia la bilis y lipoproteínas (VLDL) Fosfatiilserina → descarboxilasa → CO2 + fosfatidiletanolamina Fosfatidiletanolamina → metiltransferasa→ Fosfatidilcolina 👉 La fosfatidilserina se forma por intercambio de grupo polar Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. Estrategia 1 Fosfatidiletanolamina + serina → fosfatidilserina + colina SÍNTESIS DE COLESTEROL| Colesterol: molécula anfipática, participa en la regulación Está formado por 4 anillos hidrocarbonados unidos entre sí, una cadena carbonada y un OH que le otorga polaridad a la molécula.Procede de la dieta transportados por quilomicrones y también de la síntesis de novo. ● La mayor parte del colesterol plasmático se haya esterificado con un AG, lo que lo vuelve más hidrofóbico. ● Síntesis en citosol de todos los tejidos, reserva en hígado, suprarrenales, org reproductores ● Todos sus carbonos vienen del acetato → ac CoA, y el NADPH facilita equivalentes de reducción P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 ● Requiere de enzimas citosólicas y de la membrana del REL. ● 4 etapas generales ○ 1) síntesis de mevalonato (6 C) ○ 2)conversión a isoprenos activados ○ 2) síntesis de escualeno (30 C) ○ 3) Formación de colesterol (27 C) 1) Síntesis de mevalonato a partir de Acetil-CoA Condensación de 2 acetil-CoA por tiolasa → Acetoacetil CoA (4c) Se condensa otra molécula de Acetil-CoA liberando nuevamente la Coenzima por la HMG CoA sintasa → HMG CoA ( 6C) quien es reducido usando 2 NADPH por la HMG Reductasa → mevalonato (6C) ⚕️HMG CoA es inhibida competitivamente (al ser estructuralmente parecido al HMG coA) por las estatinas, lo que permite disminuir la síntesis de colesterol en las hipercolesterolemias La HMG CoA reductasa es una enzima de la membrana del REL! Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. de la permeabilidad y fluidez de las membranas. ¡Proceso caro! requiere 18 ATP y Ac CoA y 14 NADPH 2) Conversión en 2 isoprenos activados: Tres fosforilaciónes consecutivas donde formamos primero un 5-P-mevalonato, luego un 5-Pirofosfomevalonato y por último un 3-P-5-pirofosfomevalonato quien pasa por una decarboxilasa liberando COO- y el P en posición 3 como CO2 y Pi generando así 3) Condensación de 6 isoprenos activados con liberación de PPi para formar escualeno ● Isopentenyl PPi + Dimetilalil PPi → prenil transferasa → geranyl PPi (10C) + PPi ● Gernyl PPi + isopentenyl PPi → Farnesyl PPi (15C) +PPi ● Farnesil PPi + Farnesil PPi + NADPH → escualeno sintasa → escualeno (30 C) + NADP + 2PPi 4) Ciclación del escualeno en el núcleo esteroideo Escualeno + NADPH + O2→ Colesterol P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 → Lanosterol (30C ciclados) → monooxigenasa y luego pasa por ciclasas Formación de ésteres de colesterol → ACAT: acil CoA Colesterol Acil Transferasa | en células almacenadoras de colesterol Colesterol + Acil CoA → éster de coleserol+ CoA SH → LCAT : lecitina ( colina) colesterol acil transferasa | en HDL Colesterol + Fosfatidilcolina → éster de colesterol + lisolecitina Regulación de la vía: → HMG CoA Reductasa Regulación de su expresión según la concentración de colesterol 1.Expresión controlada x el factor de transcripción SREBP que se une al ADN que codifica para el elemento regulador de esteroles (ERE). SREBP es una una proteína integral de membrana del RE que se asocia con otra proteína, SCAP (prot activadora de clivaje) Ante [colesterol] baja el complejo SREBP- SCAP se desplazan a Golgi para clivarse por proteasas y dar un fragmento soluble que ingresa al núcleo, se une al ERE y actúa como factor de transcripción aumentando la síntesis de HMG CoA reductasa y por lo tanto aumentando la síntesis de colesterol Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. isoprenos activados como el isopentenyl PPi y dimetilalil PPi (de 5 C cada uno) Si el colesterol es abundante, se une a SCAP e inducen su unión con otras proteínas, Insig (insulin induced gene) que provocan la retención de SREBP-SCAP en el RE, provocando una reducción en la síntesis de colesterol 2. Degradación enzimática acelerada por esteroles: la reductasa es una proteína integral del RE ubiquitinización de la misma. Regulación covalente: hormonal y por estado energético → Inactiva: Fosforilada x PKA y AMPK ( responde a glucemia y a estado energético) → Activa: desfosforilada x Prot Pasa LIPOPROTEÍNAS: VLDL |SINTETIZADAS EN EL HÍGADO, TRANSPORTAN LÍPIDOS ENDÓGENOS ……. …… CARGAN APO B 100 Y SE TRANSFORMAN EN LDL P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 Se sintetizan y pasan directamente a la sangre — donde recinben Apo C y Apo E d HDL—transportando TAG endógenos para que sean hidrolizados por LPL en los diferentes tejidos. Una vez los TAG son degradados y por tanto la lipoproteína disminuyó su tamaño aumentando su densidad, devuelve Apo E y C a HDL con quien también intercambia TAG por ésteres de colesterol transformándose en IDL (lipoproteína de densidad intermedia) para luego dar lugar a una LDL LIPOPROTEÍNAS: LDL| SUMINISTRA COLESTEROL A LOS TEJIDOS Y SI NO ES CAPTADO POR ESTOS DJDHJHSDFJHSDF LO DEVUELVE AL HÍGADO Contiene mucho menos TAG y más colesterol libre y esterificado. → Endocitosis mediada por receptor: los receptores son glucoproteínas que reconocen Apo B100 y se encuentran en cavidades cubiertas citosólicamente por una cubierta de clatrina en las membranas de los hepatocitos. Una vez se da la unión receptor -LDL estos son endocitosis en vesículas que rápidamente pierden la clatrina; la vesícula se une con otra formando un endosoma que disminuye su pH gracias a una bomba de H+ para generar la disociación del receptor y que este pueda ser reciclado hacia la membrana; los componentes de la LDL se degradan en el lisosoma. Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. que detecta esteroles, ante elevados niveles la enzima se une a las Insig que provocan la —> Efecto del colesterol endocitado en la homeostasis celular: Colesterol proveniente de quilomicrones,IDL y LDL inhibe la exp de HMG CoA reductasa y acelera su degradación, así como disminuye la síntesis de receptores de LDL —> Si las LDL circulan mucho tiempo por sangre los lípidos que transporta o Apo B100 concentración de colesterol por lo que endocitan toda LDL oxidada que encuentren llenándose así de ésteres de colesterol transformándose en células espumosas → participan en la formación de placas ateroscleróticas. LIPOPROTEÍNAS: HDL| se forman en sangre por adición de lípidos a Apo A1 que se sintetiza en hígado e jsndjf intestinos. Son fuente de Apo C 2 y Apo E Las HDL nacientes contienen sobre todo fosfolípidos y apoproteínas. Trasportan colesterol excedente al hígado P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 Transporte inverso del colesterol: el colesterol excedente en tejido extrahepáticos es presentados por las células vía transportadores ABC, reconocidos por HDL que los captan para esterificados por LCAT activada gracias a Apo A1 A medida que va acumulando ésteres de colesterol se transforma en HDL3 y luego en HDL 2 ( con la mayor cantidad de colesterol en su interior) quien le transfiere al hígado los ésteres para eliminarlo vía bilis Síntesis de Eicosanoides| derivan del ácido araquidónico desde fosfolípidos de membrana por acción de fosfolipasas específicas Las prostaglandinas y tromboxanos forman parte de los eicosanoides. Tienen vida corta y no se almacenan. Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. pueden ser oxidados y reconocidos por receptores de macrófago que no son sensibles a la -Cox1 = prostaglandinas de regulan secreción gastrica P i d a u G a64b0469ff35958ef4ab887a898bd50bdfbbe91a-6698103 competitivo Cox 2 = prostaglandinas del dolor inflamación y fiebre competitivo ❌ Aspirina | irreversible ❌Ibup| ❌ Aspirina | irreversible ❌Ibup| Leucotrienos| a partir de ácido araquidónico por lipooxigenasas Glucolípidos| a partir de ceramida → componente lipídico + un glúcido activado por UDP. Síntesis en golgi. En …… mm de todas las células principalmente en tejido nervioso! Degradación en lisosomas Neutro: cerebrósidos → ceramida + monosacárido activado con UTP (actúan glicosiltransferasas) , ej: galactocerebrósidos Ácidos: gangliósidos, carga - por contener ácido siálico (monosacárido ácido) unido a la ceramida + oligosacáridos. Reservados todos los derechos. No se permite la explotación económica ni la transformación de esta obra. Queda permitida la impresión en su totalidad. Las prostaglandinas se sintetizan mediantes los Cox (ciclooxigenasas)