GENETICA VEGETAL I a IV 2013
Anuncio

GENETICA
INTRODUCCIÓN
La Genética, llamada la ciencia de la herencia, es tema central entre las ciencias
biológicas. La mayoría de los principios genéticos pueden expresarse con
exactitud en términos matemáticos, sobre todo en la genética básica, lo cual es
un rasgo único entre las disciplinas biológicas.
La Genética, descubriendo los principios en que se basa la herencia en los seres
vivos y la variación entre ellos y sus descendientes, ha contribuido mucho al
progreso de las ciencias biológicas, aclarando muchos hechos relacionados con
el origen y evolución de las especies.
La Genética Vegetal, además de estudiar los métodos que pueden emplearse
para la obtención de nuevas variedades en las plantas cultivadas y para la
mejora de los tipos existentes, investiga el modo de manifestarse o modificarse
los caracteres de mayor interés agronómico o industrial en las plantas más
importantes y el modo de conducir la mejora de estas plantas en relación a cada
uno de ellos.
El estudio y la observación de los fenómenos de la herencia, nos permite
reconocer que hay variación entre organismos de la misma especie, raza o
variedad; pero, asimismo, nos ayuda a reconocer qué características varían
únicamente debido a la herencia y cuáles varían por causa de la acción
recíproca o interacción de la herencia y el medio ambiente. Las variaciones a
que hemos hecho referencia han ido apareciendo gradualmente a través de las
edades, debido a cambios súbitos en el material genético. A estos cambios
súbitos en el material genético se les denominan mutaciones, las cuales han
producido, gradualmente, el proceso de la evolución de las especies y por lo
tanto la inmensa variabilidad existente entre los distintos organismos que
componen el cosmos.
El conocimiento y diferenciación de los caracteres cualitativos y cuantitativos,
permiten a los genetistas elegir correctamente los factores a mejorar para el
mejor y mayor provecho de la humanidad.
El presente texto trata de ser una valiosa herramienta para estudiantes que por
primera vez se involucran en el conocimiento de la herencia permitiendo
ayudarles a desarrollar un alto grado de pensamiento lógico que esta ciencia
exige; sin embargo, en algunos casos requiere de conocimientos de aritmética y
álgebra elemental para resolver problemas diversos.
1
I.
LA GENETICA - DEFINICIÓN - FINES - IMPORTANCIA – REFERENCIA
HISTORICA
1.1 DEFINICIÓN.- La Genética es la ciencia, rama de la Biología que se
ocupa del estudio de los fenómenos de la herencia y las causas de las
variaciones entre organismos. La herencia es la tendencia de los seres a
reproducir fielmente las características de sus progenitores y la variación
es la tendencia que manifiestan los seres vivos para diferenciarse unos de
otros. La resultante de las acciones mutuas de estas dos tende ncias
opuestas viene a conformar el conjunto de caracteres que cada uno
posee.
La Genética es la ciencia que estudia los fenómenos relativos a la
herencia y a la variación en los seres vivos. Las unidades hereditarias que
se transmiten de una generación a la siguiente (heredadas) se llaman
genes, los cuales están ubicados en una molécula larga llamada ácido
desoxirribonucleico (ADN).
El estudio de las variaciones entre organismos, es lo que ha permitido
demostrar que no hay dos individuos idénticos, aún dentro de la misma
especie, raza o variedad, y de ello se valen los genetistas para hacer
selección en organismos de caracteres superiores, logrando incrementar
considerablemente la producción agrícola y pecuaria por unidad.
Cuando las variaciones de los organismos se deben solamente a la
herencia, se denominan características cualitativas que no son
modificadas por el medio ambiente. Ejm.: color de ojos o de pelo en
animales, ausencia o presencia de cuernos en ovinos, etc. En cambio,
cuando las variaciones de los organismos se deben a la herencia y a la
influencia ambiental, se denominan características cuantitativas. Ejm.:
estatura o altura, inteligencia, producción de leche de una vaca,
rendimiento de papa/há, etc.
1.1 FINES.- La finalidad de la Genética es el descubrimiento de las leyes que
rigen la transmisión de los caracteres que presentan los individuos a su
descendencia, determinar el mecanismo por el cual se produce la
transmisión hereditaria y asimismo, la determinación de las causas que
originan las diferencias que se observan entre los seres de esa
descendencia.
1.3 IMPORTANCIA.- La importancia de esta ciencia radica en la parte
aplicativa tanto en animales (incluyendo al hombre) como en plantas. La
agricultura y la ganadería no sólo necesitan producir más, sino más barato
y para ello es necesario que los costos de producción disminuyan, lo que
a su vez depende de los mayores rendimientos unitarios. Estos
rendimientos unitarios en plantas, se obtienen aplicando una serie de
factores favorables al cultivo como clima, suelo, fertilizantes, pesticidas,
etc., sobre los cuales el hombre tiene poca o nula influencia; sin embargo
sí puede lograr una nueva variedad con potencial genético de rendimiento
superior utilizando todas las herramientas que la ciencia y la tecnología
ponen a su alcance. La genética humana es una herramienta auxiliar
muy valiosa en la patología del hombre, así como en el estudio de
2
algunas taras físicas y morales que se transmiten de una generación a
otra.
En los últimos tiempos, ha habido un avance vertiginoso en la obtención
de nuevas variedades de plantas con mayor potencial de rendimiento, lo
que es más, con atributos que difícilmente hace poco tiempo atrás hubiera
sido posible obtener, así tenemos que la Biotecnología es la herramienta
que ha logrado avances agigantados tanto en animales como en plantas
(clonación); la Ingeniería Genética con la obtención de plantas
transgénicas ha logrado avances de gran utilidad para la humanidad.
Toda manipulación es posible cuando los conocimientos de la genética
general y aplicada están bien cimentados y son utilizados dentro de un
marco de código de ética que garantice el respeto a la vida y su
integridad.
1.4 BREVE REFERENCIA HISTORICA
Los antecedentes de la Genética se remontan a los tiempos en que los
científicos trataban de explicar el origen y formación de las especies,
emitiendo diversas teorías desde las que defendían la inmutabilidad” de
las especies, hasta las evolucionistas.
Linneo, genial botánico sueco y muchos otros hombres de ciencia
defendían la teoría de que las especies habían permanecido sin
variaciones a través de los tiempos desde su creación y así trasmitían los
caracteres a sus descendientes sin mayores cambios.
Después de Robert Hooke y Bufón quienes pusieron en duda la teoría de
la inmutabilidad, es Lamarck el primer investigador que dice que las
especies no son inmutables, sino que existe entre ellas una constante
variación que las hace evolucionar, modificándose de generación en
generación.
Esta teoría fue duramente combatida por muchos científicos de la época
(hasta 1858 aproximadamente) y surgieron nuevos conceptos que se
acercaban poco a poco a una real teoría evolucionista como la de Darwin
que revolucionó el mundo con su obra: “El Origen de las Especies”, que
estableció firmemente en la ciencia el concepto evolucionista sobre el
origen de vegetales y animales.
Desde el siglo XVII, en que se inventaron los diferentes tipos de
microscopios, instrumento valioso para los biólogos, hasta el siglo XX en
que se producen los microscopios electrónicos; el hombre de ciencia ha
podido descifrar los detalles más recónditos de la célula y al iniciar el siglo
XXI, se presenta un panorama que avanza a pasos agigantados en el
descubrimiento de nuevos métodos de investigación sobre la vida, su
continuidad e inclusive su transformación. Un breve resumen de la obra
de algunos de aquellos hombres de ciencia se ofrece a continuación:
Henri Dutrochet, en 1824, explicó que todo tejido animal está
compuesto por células, y en 1837, descubrió que la clorofila era
necesaria para la fotosíntesis de las plantas.
3
Schleiden y Schwann (biólogos alemanes), en 1838, postularon la
“Teoría Celular”, que en resumen decía que tanto las plantas como los
animales están compuestos por células.
Von Mohl, en 1846, llamó protoplasma a la sustancia viva que contiene
la célula.
Robert Virchow, en 1858, propuso la teoría que toda célula proviene de
otra célula.
Hasta el siglo XVIII se creía que las características adquiridas se
heredan y que se transmiten de generación en generación como
defendía J. B. Lamarck.
A fines del siglo XIX, Weissman propuso y demostró que sólo se
heredan y se transmiten de generación en generación los caracteres
que se encuentran en las “células reproductivas” o “plasma germinal”,
dijo además que los caracteres adquiridos no se heredan porque dejan
de estar presente cuando el factor o factores que los inducen dejan de
actuar.
Gregor Mendel (1822-1884), en 1866, sentó las bases de la Genética,
por lo que en forma póstuma se le considera “Padre de la Genética”,
en reconocimiento a su trabajo ordenado y científico sobre
cruzamientos en guisantes que le permitió postular lo que hoy son las
leyes mendelianas.
Fleming, en 1879 y 1882, estableció los términos: mitosis, aster,
cromatina, profase, metafase, anafase y telofase.
Waldeyer, en 1888, le puso nombre al cromosoma (cuerpo coloreado).
Hugo De Vries (holandés), Carl Correns (alemán) y Erich Von
Tschermak (austriaco), en 1900, redescubrieron las leyes de Mendel al
experimentar por separado con otras plantas además del guisante.
Hardy y Weimberg sentaron las bases de la genética poblacional
mediante la aplicación de ciertos conceptos matemáticos a las
poblaciones de individuos.
Bateson (inglés), en 1909, fue el primer catedrático de Genética,
cátedra a la que él puso nombre y se estableció por primera vez en la
Universidad de Cambridge. Aceptó los trabajos de Mendel y sucesores,
aplicando los principios mendelianos a animales y plantas.
Johannsen, naturalista danés, estableció los términos, gen, genotipo,
fenotipo. Formuló su teoría de la Línea Pura.
Morgan, biólogo contemporáneo norteamericano (1866-1945),
partiendo de los conceptos mendelianos, formuló la teoría
cromosómica de la herencia. Trabajó con el díptero Drosophila
melanogaster. También explicó la mutación génica y el mecanismo de
la herencia ligada al sexo. Publicó su libro “La Teoría del Gen” en el
que describe la técnica de construir mapas de genes.
Entre los discípulos de Morgan, destacaron Müller, Bridges, Dobszanky
y otros que han contribuido a completar y afianzar con nuevos
descubrimientos esta teoría que hoy se acepta para explicar la
herencia y la variación.
4
Galton, es considerado Padre de la Biometría, por explicar y aplicar
conceptos matemáticos a la herencia cuantitativa.
Yule (inglés), en 1907; Ehle (sueco), en 1908 y East (americano),
determinaron que la herencia cuantitativa puede explicarse en
términos mendelianos.
Fisher, Pearson y Waldon contribuyeron con la Biometría entre 1908 y
1920.
La genética molecular se originó con el descubrimiento de los ácidos
nucleicos.
Avery, Mc Lead y Mc Carty en 1944, demostraron que el ADN es el
componente químico de los genes y es el portador de la información
genética.
Hersey y Chase, en 1952, dejaron establecido que el ADN es el
componente químico de los genes.
Watson y Crick en 1953, formularon la teoría sobre la estructura
hipotética del ADN.
Jacob y Monod, en 1960, desarrollaron la teoría del “Operón” para
explicar el mecanismo del control genético en la regulación de síntesis
de proteínas.
Niremberg y colaboradores en 1968, descifraron la clave genética y el
papel de cada uno de los tres ARN en la transcripción de la
información por un gen (ADN) para la síntesis de proteína por la célula.
En 1982, se obtiene el primer producto de la Ingeniería Genética: la
insulina humana, pr la Compañía Lily&Compañía.
En 1988, se inicia el proyecto: Genoma Humano, con el objetivo de
secuenciar los 23 pares de cromosomas humanos que contienen
aproximadamente 100,000 genes.
En 1996, se obtiene la secuencia del genoma de la bacteria E. coli.
En 1997, se produce la clonación del primer animal a partir de una
célula adulta: la oveja Dolly.
En el 2003, se publica la primera secuenciación completa del Genoma
humano.
La Citogenética, ha venido a destruir viejos conceptos y explicar
muchos enigmas sobre el contenido cromosómico y genético de los
organismos.
La Biotecnología, la Ingeniería Genética, han rebasado toda
expectativa sobre la manipulación del material genético (ADN) al punto
de llegar a obtener organismos vivos, completos a partir de porciones
de ADN, o de transferir porciones de ADN animal a vegetales y
viceversa, lo cual si bien es cierto que es un gran avance de la ciencia,
llama a reflexión sobre cuáles deberían ser los límites permisibles para
realizar dichas investigaciones en animales y en humanos sobre todo;
por lo que se hace referencia a un necesario Código de Ética.
II. LAS BASES FISICAS DE LA HERENCIA
5
La teoría celular desarrollada durante el siglo pasado, estableció
definitivamente que la célula es la unidad estructural y funcional de todo
organismo, sea éste animal o vegetal. Es necesario entonces recordar qué es
la célula, cuál es su constitución y algunos otros aspectos que ayuden a
comprender su participación en la reproducción.
2.1 LA ESTRUCTURA CELULAR
En forma general, la célula está formada por tres partes fundamentales:
Membrana celular, Citoplasma y Núcleo.
Membrana celular.- Es la parte externa de la célula que contiene a las
demás, a través de ella se realizan los intercambios de sustancias que
ponen al citoplasma en conexión con el medio. Tiene la propiedad de
poseer permeabilidad diferencial, dando lugar a los fenómenos osmóticos,
tan importantes en la vida de la célula.
Citoplasma.- Es la parte del protoplasma celular que ocupa el espacio
entre la membrana y el núcleo. Es el lugar donde se efectúan los
fenómenos que determinan la nutrición celular. Las células vegetales
tienen pared celular que rodea la membrana plasmática, plastidios,
plasmodesmos, esferosomas y vacuolas que las células animales no
poseen; en cambio carecen de aster o centrosoma con el centriolo en su
interior que sólo tienen las células animales. En general, las células tienen
entre sus organelos al retículo endoplasmático, aparato de golgi,
ribosomas y mitocondrias.
Núcleo.- Es la parte vital de la célula que consta de: membrana nuclear,
jugo nuclear o cariolinfa, red de cromatina y nucléolo. La red de
cromatina está formada por unos filamentos llamados cromonemas, que
posteriormente serán los cromosomas, elemento fundamental en
Genética. Está demostrado que entre todas las partes de la célula, el
papel primordial corresponde al núcleo, el cual preside el metabolismo de
la célula, interviene en la elaboración de sustancias con destino a la
asimilación, en el crecimiento, en la formación de la membrana y en la
constitución y empleo de reservas.
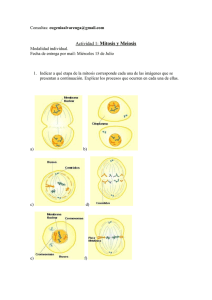
2.2 LA DIVISION CELULAR :
MITOSIS
Llamada también cariocinesis, es un proceso de división cuya función es
en primer lugar, construir una copia exacta de cada cromosoma y después
distribuir por medio de división de la célula original (célula madre) un
juego idéntico de cromosomas para cada una de las células hijas.
FASES
1. PROFASE.- Los cromosomas aparecen visibles al microscopio de luz,
las dos cromatidas hermanas pueden verse unidas a su centrómero
común, aparecen las fibras del huso acromático y comienza a
desaparecer la membrana nuclear y el nucleolo.
6
2. METAFASE.- La membrana nuclear y el nucleolo han desaparecido
por completo. Los cromosomas se ubican en la placa ecuatorial de la
célula. Las fibras del huso acromático están unidas a los cromosomas
por el centrómero.
3. ANAFASE.- Las cromátidas hermanas se separan en forma
longitudinal a partir del centrómero y se dirigen a polos opuestos de la
célula.
4. TELOFASE.- En cada polo de la célula, se han reunido un juego idéntico
de cromosomas. El huso acromático degenera, la membrana nuclear se
regenera y el citoplasma se divide por un proceso llamado CITOCINESIS,
que en las células vegetales se inicia con la formación del fragmoplasto
en la zona ecuatorial de la célula (de adentro hacia afuera); mientras que
en células animales se inicia con el estrangulamiento de esa zona (de
afuera hacia adentro).
FASES DE MITOSIS VEGETAL
FASES DE MITOSIS ANIMAL
7
MEIOSIS
La reproducción sexual requiere de la elaboración de gametos
(gametogénesis) y su unión (fertilización). La gametogénesis ocurre en las
células especializadas (línea germinal) de los órganos reproductivos. Los
gametos contienen el número haploide de cromosomas (n), pero se
originan en células diploides (2n). El proceso reductivo de la
gametogénesis es llamado meiosis, que implica dos divisiones:
1. MEIOSIS l O DIVISION REDUCTORA.- Es una división reductora
porque produce dos células haploides (n) a partir de una sola célula
diploide (2n).
2. MEIOSIS II O DIVISION ECUACIONAL.- Es una división equitativa que
separa y divide las cromátidas hermanas de las células haploides y
origina cuatro células haploides.
FASES :
1. MEIOSIS l :
A) PROFASE l .- Consta de cinco sub-fases:
Leptoteno.- Se observan las dos cromátidas unidas por el
centrómero.
Cigoteno .- Los cromosomas homólogos se colocan lado a lado
en un proceso de apareamiento (sinapsis).
Paquiteno.- Los cromosomas homólogos (bivalentes o tetradas)
se acortan, se condensan y se entrecruzan intercambiando
material genético entre cromátidas no hermanas.
Diploteno.- Los bivalentes se separan por las cromátidas pero
se mantienen unidos por el centrómero y por los puntos donde
intercambian material genético (quiasmas).
Diacinesis.- Las cuatro cromátidas de los dos cromosomas
homólogos forman configuraciones llamadas tetradas.
Al final de la Profase l, se ha formado el huso acromático y empiezan a
desaparecer la membrana nuclear y el nucleolo.
B) METAFASE l .- Los cromosomas homólogos (bivalentes) se alínean
en la zona ecuatorial de la célula.
C) ANAFASE l .- Los bivalentes se repelen entre sí y se mueven hacia
polos opuestos de la célula. Cada cromosoma (díada) va a un polo.
D) TELOFASE l.- El nuevo juego de cromosomas aún en estado
duplicado (díada) empieza a ser rodeado por la membrana nuclear
que se regenera y se divide el citoplasma (citocinesis), originando
dos células haploides (n).
2.
MEIOSIS II .-Empieza luego de una Interfase corta o después de
Telofase I
A) PROFASE ll.- Las díadas permanecen cortas y visibles, se forma
un nuevo huso acromático.
8
B) METAFASE II.- Las díadas o univalentes se alínean en la zona
ecuatorial de la célula.
C) ANAFASE II .- Los cromosomas univalentes se dividen por el
centrómero y las cromátidas se repelen y se dirigen a polos
opuestos de la célula.
D) TELOFASE II.- Se forma la membrana nuclear alrededor de los
cromosomas en número haploide (n) en las cuatro células hijas que
se han originado al finalizar la citocinesis.
Fases de meiosis en una célula vegetal diploide 2n = 6
Fases de la meiosis animal
9
2.3 GAMETOGÉNESIS
Es el proceso completo de producir gametos maduros, cuya parte
principal es la división meiótica. El producto de la meiosis no son
gametos, sino células haploides que requieren de un proceso de
crecimiento y maduración en animales y de varias divisiones mitótic as
en plantas.
2.3.1 GAMETOGÉNESIS EN ANIMALES.- La gametogénesis en
animales machos es la Espermatogénesis y en animales hembras es
la Ovogénesis.
ESPERMATOGENESIS.- Se origina en las células diploides
primordiales en las gónadas masculinas (testículos), las cuales por
crecimiento originan las espermatogonias (2n) y posteriormente un
espermatocito primario también (2n) que es el que va a realizar la
meiosis. La Meiosis l produce dos espermatocitos secundarios (n),
de los cuales, luego de Meiosis II se producen cuatro células
haploides llamadas espermátides (n), las que originarán por
maduración a los gametos funcionales llamados espermatozoides.
OVOGENESIS.- Es la gametogénesis en el animal hembra, que se
origina en las células primordiales diploides llamadas ovogonias (en
el ovario), las cuales por crecimiento dan lugar a los ovocitos
primarios (2n). Cada ovocito primario realiza la meiosis. La meiosis l
produce un ovocito secundario (n) y un corpúsculo polar primario. En
meiosis II, el ovocito secundario da origen a la ovótide (n) y a un
corpúsculo polar secundario; el corpúsculo polar primario se divide
en dos corpúsculos polares secundarios. La ovótide por maduración
se convierte en gameto funcional llamado óvulo.
ESPERMATOGENESIS Y OVOGENESIS
10
La unión de los gametos masculino (espermatozoide) y femenino
(óvulo), se llama fertilización, y de esta manera se restablece el número
diploide en la célula resultante que se llama cigoto. Subsecuentes
divisiones mitóticas producen las numerosas células del embrión, que se
organiza en tejidos y órganos del nuevo individuo.
2.3.2 GAMETOGÉNESIS EN PLANTAS (Angiospermas)
La gametogénesis en la parte masculina de la flor es la
microsporogénesis y la gametogénesis en la parte femenina de la flor
es la megasporogénesis.
MICROSPOROGÉNESIS.- Es la gametogénesis que ocurre en al
androceo de la planta. Una célula madre diploide
llamada
microsporocito se divide por meiosis. La Meiosis l forma un par de
células haploides (n) que permanencen unidas. La meiosis II produce
cuatro microsporas (n) que se mantienen unidas.
Después de la meiosis, cada microspora sufre una división mitótica
sin citocinesis, originando una célula con dos núcleos haploides (uno
generativo y el otro núcleo del tubo). El núcleo generativo se divide
nuevamente por mitosis sin citocinesis para formar dos núcleos
espermáticos. El otro núcleo que no se divide se convierte en el
núcleo tubular.
Microsporogénesis en Angiospermas
MEGASPOROGÉNESIS.- Es la gametogénesis que ocurre en el
gineceo de la planta. Una célula madre diploide llamada
megasporocito (2n), se divide en el ovario por meiosis, formando
después de la meiosis l, un par de células haploides (n). La meiosis
II produce un grupo lineal de cuatro megasporas haploides (n).
Después de la meiosis, tres megasporas degeneran y la otra sufre
tres divisiones mitóticas de los cromosomas sin citocinesis,
formando un saco inmaduro con ocho núcleos haploides (n). Tres
de los núcleos se orientan hacia el extremo micropilar, dos de los
cuales llamados sinérgidas, degeneran, el tercer núcleo será el
11
núcleo del huevo. Otro grupo de tres núcleos llamados antípodas,
se mueven hacia el extremo opuesto del saco y también
degeneran. Los dos núcleos restantes son los núcleos polares se
unen en el centro del saco formando un solo núcleo diploide de
fusión (2n). El saco embrionario maduro está listo para la
fecundación.
Megasporogénesis en Angiospermas
DOBLE FECUNDACION EN ANGIOSPERMAS.- Al caer el grano de
polen del androceo de una flor al gineceo de la misma o de otra
(polinización), empieza a germinar bajando por el tubo polínico del
estilo hasta hacer contacto con el óvulo en el ovario. Uno de los
núcleos espermáticos del grano de polen se funde con el huevo,
formando un cigote diploide (2n), que por mitosis dará origen al
embrión (2n). El otro núcleo espermático (n) se une con los núcleos
polares de fusión (2n) formando un núcleo triploide (3n) que por
mitosis dará lugar al endosperma de la semilla. Ocurre entonces un
doble proceso de fecundación para formar al embrión y al
endosperma de la semilla, porque participan dos núcleos
espermáticos.
Doble fecundación en angiospermas
12
III.- LA HERENCIA MENDELIANA
Conviene aclarar que Mendel, monge austriaco, profesor de Botánica,
ignoraba la existencia de los cromosomas y, en su tiempo, la teoría celular
se hallaba en sus inicios. Realizó numerosos experimentos de
cruzamientos sobre Pisum sativum y estudió escrupulosamente la
distribución de los caracteres hereditarios en los descendientes resultantes
de estos cruzamientos. Para explicar lo que comprobaba, es decir los
fenotipos que observaba, Mendel emitió la hipótesis de que los caracteres
eran determinados por unos factores específicos (los genes) como hoy se
dice, conforme a tres leyes, hoy llamadas Leyes de Mendel.
3.1 PRIMERA LEY DE MENDEL: LEY DE LA DOMINANCIA
Esta ley dice: “Si se cruzan dos individuos pertenecientes a razas puras,
homocigotas alternantes para un determinado carácter, todos los híbridos
de la primera generación (el 100%), son iguales a uno de los padres”.
Ejm 1: Mendel cruzó plantas de guisante de semilla amarilla (VV) con
plantas de semilla verde (vv) y en la primera generación filial (F1)
obtuvo solamente semillas amarillas.
P
:
Fenotipos:
Gametos :
diferentes
VV
X
semilla amarilla
{V
F1
:
Fenotipo :
vv
semilla verde
v}
V v --- 100%
semillas amarillas
Al expresarse solamente el color amarillo en la F1, se deduce que es el
caracter dominante al color verde, y éste vendría a ser el caracter
recesivo que permanece oculto en esta generación.
Ejem. 2: El carácter “ forma de la semilla” en guisantes puede ser: lisas
(R) y rugosas (r). Si se cruzan dos líneas puras alternantes,
tendríamos el siguiente resultado:
P
:
Fenotipos :
Gametos
diferentes :
F1
:
Fenotipo :
RR
x
semilla lisa
{
rr
se milla rugosa
R
r }
Rr ----
100%
semillas lisas
Entonces, el carácter dominante es semilla lisa y el recesivo es semilla
rugosa.
13
DEFINICIONES: FENOTIPO Y GENOTIPO
El conjunto de caracteres que presenta un individuo constituye lo que se
denomina el fenotipo (del griego Phainein = parecer) de este individuo. Si
tomamos como ejemplo el guisante con el que trabajó Mendel, podemos
mencionar las siguientes características fenotípicas: forma de semilla (lisa
o rugosa); color de la semilla (amarilla o verde); tamaño de plantas (altas o
bajas); color de las flores (púrpuras o blancas), etc.
El conjunto de todos los genes ( o factores hereditarios como los llamaba
Mendel) de un individuo, constituye su genotipo.
Pero, mientras que el fenotipo es relativamente fácil de describir (basta con
hacer la lista de todos los caracteres observables en un individuo), el
genotipo es muy difícil de descubrir, ya que no todos los genes se
manifiestan obligatoriamente en el fenotipo; algunos pueden hallarse
enmascarados, neutralizados por otros que los dominan y pueden aparecer
solo al cabo de varias generaciones. Por lo tanto, el genotipo debe ser
deducido mucho más que observado. El fenotipo ayuda a deducir en forma
relativa al genotipo.
3.2 SEGUNDA LEY DE MENDEL: LEY DE LA SEGREGACION
Esta ley dice: “Si se cruzan o se autofecundan dos individuos de la primera
generación filial (F1), en la segunda generación (F2) se obtendrá una
proporción fenotípica de 3:1 para el carácter dominante y recesivo
respectivamente”.
El carácter recesivo que aparentemente había desaparecido, vuelve a
manifestarse en esta segunda generación.
Retomando el primer caso anterior:
F1
:
Fenotipo
:
Gametos
Diferentes
:
F2
Vv
x
semillas amarillas
{V,v
Vv
semillas amarillas
V,v }
:
♂
V
v
VV
Vv
♀
V
s. amarilla s. amarilla
v
Genotipos
Proporción genotípica
Proporción fenotípica
Fenotipos
Vv
vv
s. amarilla
s. verde
:
VV , Vv ,
:
1 : 2 :
:
3
:
: s.amarillas,
vv
1
1
s. verdes
14
Del mismo modo, con las semillas lisas y rugosas:
F1
:
Rr
x
Rr
Fenotipo
:
semilla lisa
semilla lisa
Gametos
{R,r
R,r }
diferentes
:
F2
:
♂
R
r
♀
R
RR
Rr
r
Genotipos
:
Proporción genotípica:
s.lisa
s. lisa
Rr
rr
s. lisa
s. rugosa
RR, Rr , rr
1 : 2 : 1
Proporción fenotípica :
Fenotipos
3
:
:
1
lisas , rugosas
3.3 TERCERA LEY DE MENDEL: LEY DE LA INDEPENDENCIA DE LOS
CARACTERES
“Si se cruzan dos líneas puras con formas alternantes para dos caracteres
distintos, cada uno de ellos se transmite en forma independiente respecto
del otro. Los individuos que se formen en la segunda generación filial F 2
responden a todas las combinaciones posibles, con una proporción
fenotípica de 9:3:3:1”. Las dos leyes anteriores se cumplen para cada uno
de los caracteres alternantes involucrados.
Continuando con los mismos casos anteriores, se tiene que:
Si se cruza una línea pura de guisantes con semillas lisas y
amarillas(RRVV) con otra de semillas rugosas y verdes (rrvv) , en la
primera generación filial F 1 , se tendrá un dihíbrido (RrVv), cumpliéndose la
primera ley; luego al autofecundar dos plantas F 1 , en la F 2 , se tendrá una
descendencia con todas las formas de combinación posible, presentando
una proporción fenotípica de 9:3:3:1.
P
:
RRVV
x
rrvv
Fenotipos
:
semillas lisas
semillas rugosas
amarillas
verdes
Gametos
diferentes
:
F1
(Dihíbrido) :
P
(F1)
{
RV
rv
}
RrVv -
100%
semillas lisas, amarillas
Continuando con nuestro ejemplo: Cruce de plantas F1:
Gametos
diferentes
:
RrVv
:
{ RV, Rv, rV, rv
x
RrVv
RV, Rv, rV, rv }
15
F2:
♂
♀
RV
Rv
rV
rv
P.G
RV
Rv
rV
rv
RRVV
s.lisas
amarillas
RRVv
s. lisas
amarillas
RrVV
s. lisas
amarillas
RrVv
s. lisas
amarillas
RRVv
s. lisas
amarillas
RRvv
s. lisas
verdes
RrVv
s. lisas
amarillas
Rrvv
s. lisas
verdes
RrVV
s. lisas
amarillas
RrVv
s. lisas
amarillas
rrVV
s.rugosas
amarilla
rrVv
s.rugosas
amarillas
RrVv
s. lisas
amarillas
Rrvv
s. lisas
verdes
rrVv
s.rugosas
amarillas
rrvv
s.rugosas
verdes
Genotipos
1
2
1
2
4
2
1
2
1
RRVV
RRVv
RRvv
RrVV
RrVv
Rrvv
rrVV
rrVv
rrvv
P.F.
Fenotipos
9
s. lisas, amarillas
3
s. lisas, verdes
3
s. rugosas, amarillas
1
s. rugosas, verdes
3.4 CRUCE DE PRUEBA
Es un cruzamiento retrógrado o un retrocruzamiento que consiste en cruzar
individuos de la primera generación filial F 1 (híbridos, dihíbridos) con el
progenitor totalmente recesivo. Cuando no se conoce el genotipo de un
individuo, se recurre al cruce de prueba con la finalidad de identificar al otro
progenitor (dominante) para comprobar si se encuentra en estado
homocigota o heterocigota.
a) P (F1)
Fenotipo
:
Gametos dif.
:
Descendencia
:
P.G.
b) P (F1)
Fenotipo
Gametos dif.
:
Ee
planta alta
x
{ E, e
e }
Ee
50% plantas altas
ee
50% plantas enanas
1:1
:
EeBb
x
: pta.alta, flor roja
:
ee
planta enana
{EB, Eb, eB, eb
eebb
pta.enana, flor blanca
eb}
16
Descendencia
:
P.G.
Genotipos
1
1
1
1
EeBb
Eebb
eeBb
eebb
Fenotipos
planta alta, flor roja
planta alta, flor blanca
planta enana, flor roja
planta enana, flor blanca
3.5 SIMBOLOGIA EN POLIHÍBRIDOS
n
=
número de pares de alelos (genes) involucrados en el cruce.
(2)n
=
número de gametos diferentes producidos por un híbrido F1.
(2)n
=
número de clases fenotípicas de individuos con dominancia
completa.
(3)n
=
número de genotipos posibles.
(4)n
=
número de combinaciones posibles de gametos en F1.
(4)n
=
número mínimo de individuos en la F2 necesarios para
obtener proporciones fenotípicas correctas.
IV. MODIFICACIONES DE LA HERENCIA MENDELIANA
4.1 DOMINANCIA INCOMPLETA O HERENCIA INTERMEDIA
Se dice que hay dominancia incompleta o herencia intermedia cuando
ninguno de los genes involucrados enmascara o domina totalmente al otro;
por lo que, los híbridos formados presentan un fenotipo intermedio al que
producen los individuos homocigotas recíprocos. Cuando hay dominancia
incompleta entre dos alelos, las proporciones fenotípicas en la F2 son 1:2:1
y cada fenotipo describe a su respectivo genotipo.
Ejem. : En ganado de la raza Shorthorn, existen tres colores para el pelaje:
rojo, blanco y roano. El color rojo, es producido por el gen ( R ) en estado
homocigota, el blanco es producido por su alelo, el gen (R’) en estado
homocigota. El cruce entre un toro Shorthorn rojo (RR) y un hato de vacas
blancas (R’R’) de la misma raza, producirá crías de color roano (RR”) en la
F1. Si esquematizamos este ejemplo, tenemos:
P
Fenotipos
Gametos
diferentes
:
:
R’R’
blanco
:
{ R’
F1
:
x
RR
rojo
R}
RR’ -----> 100% roanos
Si se cruza dos animales de la F1 :
P (F1 x F1) :
Fenotipos
:
Gametos
diferentes
:
RR’
roano
{ R , R’
x
RR’
roano
R , R’ }
17
F2:
♂
♀
R
R’
PG
1
2
1
R
R’
RR
rojo
RR’
roano
RR’
roano
R’R’
blanco
Genotipos
RR
RR’
R’R’
P.F.
1
2
1
Fenotipos
rojos
roanos
blancos
Se deduce entonces, que el gen para el color rojo, es dominante
incompleto sobre el gen para el color blanco, y que cuando ambos alelos
están juntos producen un nuevo fenotipo diferente a los progenitores
homocigotas.
Del mismo modo, cuando se trata de herencia intermedia para dos pares de
genes, las proporciones mendelianas en la F2, se modifican de 9:3:3:1 a
1:2:1:2:4:2:1:2:1 ya que cada genotipo origina su propio fenotipo.
4.2 GENES LETALES
Se denomina así a cierto tipo de genes que producen la muerte del
individuo que los porta, ya sea durante el período pre-natal o entre el
nacimiento y el inicio de la madurez sexual. La mayoría de estos gen es,
manifiestan su acción letal en estado homocigota dominante o recesiva;
pero también puede haber dominancia incompleta del gen letal y el
individuo heterocigota manifiesta un fenotipo defectuoso. Las proporciones
fenotípicas mendelianas en F2, se modifican a 2:1.
Ejemplo: En aves de corral, existe un defecto llamado micromelia (las aves
casi se arrastran porque tanto las patas como las alas están afectadas). Se
ha determinado que el gen responsable (M) en estado homocigota, es letal;
que el genotipo heterocigota (Mm) produce el fenotipo de la micromelia y
solamente el homocigota recesivo (mm) es normal. Si esquematizamos la
cruza entre dos animales micromélicos, se tendrá:
P
Fenotipos
Gametos
Diferentes
:
:
Mm
micromélica
:
x
Mm
micromélico
{M,m
M,m}
Descendencia (F2):
♂
♀
M
m
M
m
MM
mueren
Mm
micromélica
Mm
Micromélica
mm
normal
18
P.G. Genotipos
1
MM
2
Mm
1
mm
P.F.
X
2
1
Fenotipos
--micromélicos
normal
4.3 ALELISMO, SERIES ALELICAS O ALELOS MULTIPLES
Se refiere a un grupo de genes (más de dos) que componen una serie
alélica; de la cual solamente dos alelos ocupan el mismo locus de
cromosomas homólogos a la vez, para determinar un fenotipo. Los
fenotipos que se observan se presentan de acuerdo a una jerarquía de
dominancia, según la cual un gen es dominante a todos los demás alelos
y otro es recesivo a todos los demás alelos.
Casos de alelismo o alelos múltiples, se presentan con frecuencia en el
color del pelaje y plumaje en animales, así como en el color de ojos de
Drosophila; también hay una serie alélica de esterilidad en plantas y en
humanos se presenta en determinados tipos o grupos de sangre.
4.3.1 Series alélicas o Alelos Múltiples en animales
El caso más clásico que siempre se menciona es el del color del pelaje
en conejos, que está gobernado por una serie de alelos múltiples que
producen cuatro fenotipos distintos con la siguiente jerarquía de
dominancia:
Agutí
Chinchilla
Himalaya
Albino
: color ancestral silvestre (gris uniforme)
: plomo plateado
: blancos con orejas, nariz, patas y cola de color negro.
: blanco
La serie alélica se puede resumir de la siguiente manera:
C > c ch > c h > c
La jerarquía de dominancia se expresa de la siguiente manera:
Dominantes
Genotipos
Fenotipos
Recesivo
CC, Cc ch , Cc h , Cc
c ch c ch , cch ch , cch c
ch ch , ch c
cc
Agutí (gris uniforme)
Chinchilla
himalaya
albino
Ejemplos:
1) El cruce entre una coneja (CC), con un conejo albino (cc), producirá
un híbrido (Cc), 100% de color agutí, cumpliendo la primera ley
mendeliana.
P
:
Fenotipos :
CC
agutí
Gametos
diferentes :
{C
x
cc
albino
c}
19
Descendencia (F1):
uniforme)
Cc
------->
100%
agutí
(gris
2) Si se cruzan dos conejos híbridos (Cc), se obtendrá la proporción
fenotípica clásica mendeliana de 3:1.
P
:
Cc
x
Cc
Fenotipos :
Agutí
Agutí
Gametos
diferentes :
{ C, c
Descendencia (F2):
C, c }
♂
♀
C
C
CC
agutí
Cc
agutí
c
P.G.
1
2
1
Genotipos
CC
Cc
cc
c
Cc
agutí
cc
albino
P.F.
3
Fenotipos
Agutí
1
albino
3) El cruce entre una coneja Cc ch y un conejo c ch c h , producirá la
siguiente descendencia:
P
:
Fenotipos :
Gametos
diferentes :
Cc ch
x
c ch c h
agutí
chinchilla
C
c ch
c ch
ch
Descendencia:
P.G.
1
1
1
1
Genotipos
C c ch
C ch
c ch c ch
cch c h
P.F.
2
Fenotipos
Agutí
2
chinchilla
4.3.2 Alelos Múltiples y Esterilidad en Plantas
Las plantas de Nicotiana sp. (tabaco), de trébol y de algunas otras
especies producen gametos fértiles pero incompatibles, debido a que
existe una serie multialélica que interfiere con la polinización,
produciendo esterilidad por autoincompatibilidad de ciertos genes
contenidos en el polen, que también están en el tejido esporofítico del
estilo, que reducen la velocidad del crecimiento del tubo polínico, hasta
el punto que se marchita la flor, antes que dicho tubo llegue al óvulo.
Cuando hay compatibilidad genética el polen se desarrolla a una
20
velocidad normal, realizándose a tiempo la polinización y la
fecundación. La serie alélica es: S 1 , S 2 , S 3 , S 4, . . . . . . . . . . . . , S 15
Cuando un grano de polen lleva el alelo S 1 , y en el genotipo de la
planta madre también se encuentra este alelo, entonces el tubo polínico
no se desarrolla en forma normal y la planta se comporta com o
autoestéril porque no hay autopolinización. Así, se tiene que no se
forman genotipos homocigotas.
Mecanismo de autoincompatibilidad gametofítica
Ejemplos:
1) Plantas S 1 S 2 , se cruzan sin problemas con plantas S 3 S 4 , porque
producen gametos diferentes.
P
:
Gametos
Diferentes
:
S1 S2
x
S3 S4
S1
S3
S2
S4
Descendencia:
P.G.
Genotipos
1
S1 S3
1
S1 S4
100% viables, heterocigotas
1
S 2 S3
1
S2 S4
En ningún caso se produce una autofecundación.
2) Cuando se cruzan plantas S 2 S 3 con plantas S 1 S 3 , la descendencia
que se produce es:
P
:
Gametos diferentes
S2 S3
:
x
S1 S3
S2
S1
S3
S3
Descendencia:
21
P.G.
1
1
1
1
Genotipos
S1 S2
S2 S3
75% viables
S1 S3
X
(No se forma el genotipo homocigota S 3 S 3 )
4.3.3 Alelos múltiples y los Tipos de Sangre en humanos
La membrana celular de los glóbulos rojos contiene en su superficie,
diferentes antígenos, los cuales son los responsables de los diferentes tipos
de sangre o grupos sanguíneos. Existen principalmente dos tipos de
antígenos que determinan el tipo de sangre: el Antígeno A y el B.
Según las diferentes combinaciones de los antígenos de la superficie de los
glóbulos rojos se obtienen como resultado los 4 grupos sanguíneos
existentes:
Grupo A
: Tiene Antígeno A en la superficie del glóbulo rojo.
Grupo B
: Tiene Antígeno B en la superficie del glóbulo rojo.
Grupo AB
: Tiene ambos Antígenos A y B.
Grupo O
: No tiene Antígeno (A ó B) en la superficie del glóbulo
rojo.
En el fenogrupo ABO, en humanos, de acuerdo a la reacción antígenoanticuerpo, la sangre puede presentar los siguientes genotipos:
Genotipos
(Grupo Fenotipo)
A
A
A
l l ,l i
A
B
B
B
l l ,l i
B
lA lB
AB
ii
O
Las transfusiones sanguíneas dependen de la compatibilidad entre los
grupos, que se puede resumir en el siguiente esquema:
O -------> Dador universal
A
B
AB -------> Receptor universal
Sistema ABO de grupo sanguíneo en humanos
Ejemplos:
1) Un matrimonio tiene los siguientes genotipos de grupos sanguíneos: l A lB
x lB i
Cuáles serán los grupos sanguíneos de los hijos que lleguen a tener?
Padres
:
Fenogrupos:
lA lB
AB
x
lB i
B
22
Gametos
diferentes :
IA
IB
lB
i
P.G.
Genotipos
PF
Fenogrupos
(fenotipos)
1
lA lB
1
AB
1
lA i
1
A
B B
1
l l
2
B
1
lB i
2) Cuál es la probabilidad de que cuando María (grupo A) se case con
Juan (Grupo B), tengan hijos del grupo sanguíneo O?.
María (Grupo A) -------> l A lA ó
lA i
B B
Juan (Grupo B) -------> l l ó
lB i
Razonamiento: para que un hijo sea del grupo O, tiene que recibir el gen
“i” de ambos padres.
P
Fenogrupos
:
:
Gametos
diferentes
:
P.G.
:
Genotipos :
P.F.
:
Fenogrupos:
1 :
lA lB
1 :
AB,
María lA i
A
lA
1 :
, lA i
1 :
A ,
i
1 : 1
, l B i , ii
1 : 1
B, O
x
Juan l B i
B
lB
i
Rpta.- existe un 25% de probabilidad de que tengan hijos del grupo O.
Factor Rh en humanos.El Rh es otro antígeno que puede estar o no presente en la superficie del
glóbulo rojo. Este antígeno es llamado Rh o antígeno D, debido a que las
primeras investigaciones se llevaron a cabo experimentando con un simio del
tipo Macaccus Rhesus, en el cual se observó que al inyectarle hematíes
humanos, estos producían un anticuerpo que era capaz de reaccionar
aglutinando los hematíes en el 85% de la población.
Dependiendo de la presencia o no del antígeno Rh en la superficie del glóbulo
rojo, este será positivo o negativo. Si está presente en la superficie del
glóbulo rojo será Rh positivo, y los hematíes son aglutinados por este
anticuerpo y si está ausente, es Rh negativo y no son aglutinados.
De esta forma, una persona tiene un grupo sanguíneo formado por un
antígeno A, B, las dos o ninguna y además será Rh positivo o negativo.
Este Factor se encuentra en la sangre del 85% de las personas, que se
denominan Rh positivas, mientras que el 15% restante que carece de este
factor, y se denominan Rh negativas.
23
De la misma manera que en el sistema ABO, en el sistema Rh no se puede
realizar transfusión de glóbulos rojos con el antígeno Rh a las personas que
no lo tienen, ya que podría originar la producción de anticuerpos anti-Rh en el
receptor. Los sujetos Rh negativos sólo podrán recibir sangre de donantes Rh
negativos.
Genotipos
RR, Rr
rr
Fenotipo (Factor)
Rh +
Rh -
Ejemplo 1: Determinar el factor sanguíneo de un niño cuyos padres son:
P
:
Rr
Gametos
diferentes
x
RR
{R
R}
r
P.G
1
1
Genotipos
RR
Rr
P.F.
Factor
Rh +
2
El factor sanguíneo del niño es Rh + , con 100% de certeza.
Ejemplo 2: Cuál es la probabilidad de cada uno de los factores sanguíneos
a lograrse en un matrimonio de los siguientes genotipos?:
Rr
x
Rr
Gametos
diferentes
R
R
r
P.G.
1
2
1
Genotipos
RR
Rr
rr
r
P.F.
Factor
Rh +
3
1
Rh
-
La probabilidad del factor Rh + es 3 veces superior al Rh - ; lo que equivale a
un 75% para el primero y 25% para el segundo.
4.4 INTERACCIONES GENICAS.
Se denomina así cuando alguna característica se produce por efecto de la
acción recíproca de dos o más pares de genes. Esta interacción puede ser
con epistasis o sin epistasis.
4.4.1 Interacción Génica sin Epistasis.Se produce cuando en la interacción génica no hay un efecto
epistático de ningún gen sobre otro, sino más bien una acción
complementaria entre genes dominantes o recesivos.
Existe un caso de interacción génica sin epistasis en aves de corral,
en las cuales existen diversos tipos de cresta. La raza Wyandotte
presenta cresta “tipo roseta”, la raza Brahma “tipo guisante” y la raza
24
Leghorn “tipo simple”. Al cruzar aves de la raza Wyandotte con
Brahma, toda la descendencia presentó cresta “tipo nuez”. En un
cruce F2, se observa la P.F. 9:3:3:1, ya conocida pero en la que se
involucran dos pares de genes para formar un solo carácter.
Se sabe que los diversos tipos de cresta se presentan por las
siguientes interacciones de los genes R y P y sus alelos:
Genotipos
R-PR - pp
rr P rr pp
Fenotipos
Nuez
Roseta
Guisante
Simple
Ejemplo: El cruce de aves de cresta Roseta (RRpp) con aves de
cresta Guisante (rrPP), produce en la F1, aves con cresta Nuez
(RrPp). Cómo será la descendencia en F2?
P:
Fenotipos
Gametos
diferentes
F1:
RR pp
roseta
x
Rp
F2: (F1 x F1)
rP
Rr Pp ----> 100% cresta tipo Nuez
Rr Pp
x
Gametos diferentes {RP, Rp, rP, rp
♂
♀
RP
Rp
rP
rp
P.G.
1
2
1
2
4
2
1
2
1
rr PP
guisante
RP
Rp
RRPP
nuez
RRPp
nuez
RrPP
nuez
RrPp
nuez
RRPp
nuez
RRpp
roseta
RrPp
nuez
Rrpp
roseta
Genotipos
RR PP
RR Pp
RR pp
Rr PP
Rr Pp
Rr pp
rr PP
rr Pp
rr pp
RP, Rp, rP, rp}
rP
RrPP
nuez
RrPp
nuez
rrPP
guisante
rrPp
guisante
P.F.
Rr Pp
rp
RrPp
nuez
Rrpp
Roseta
rrPp
guisante
rrpp
simple
Fenotipos
9
tipo “Nuez”
3
tipo “Roseta”
3
1
tipo “Guisante”
tipo “Simple”
La proporción fenotípica conocida 9:3:3:1, de un dihíbrido se
mantiene, pero con cuatro características diferentes producidas por la
interacción génica sin epistasis de los genes R y P ó r y p.
25
4.4.2 Interacciones Génicas con Epistasis.Es la interacción génica que se produce como resultado del e fecto
epistático que se produce entre genes no alélicos.
Epistasis.- Es la acción que se produce cuando un gen enmascara o
anula la manifestación de otro gen no alélico. Como consecuencia de
ello, hay un gen epistático que es el que se manifiesta inhibien do la
acción del otro no alélico y hay un gen hipostático, que es aquél gen
que no se manifiesta por acción del gen epistático. Estos genes no
alélicos pueden estar ubicados en un mismo cromosoma o en
cromosomas diferentes.
El concepto de epistasis, puede parecerse al concepto de dominancia;
sin embargo nótese que se trata de genes no alélicos, lo que los
diferencia completamente.
Entre las diversas epistasis conocidas que modifican la proporción
fenotípica en F2, se pueden mencionar:
1.- Epistasis Recesiva.9:3:4
2.- Epistasis Dominante.12:3:1
3.- Epistasis con efectos complementarios.9:7
4.- Epistasis con acción duplicada de genes.15:1
1.- Epistasis Recesiva.- Se produce cuando un gen recesivo es
epistático a un gen dominante y al alelo de éste.
Ejemplo.- En roedores, el gen (N) produce el color negro y su alelo
recesivo (n) produce color marrón en presencia del gen (C). El gen (c)
en estado homocigota es epistático a los genes N y n, produciendo
individuos albinos. Cómo se modificará la proporción fenotípica en
F2?. Demuestre.
Genotipos
C_ N_
C_ nn
cc N_
cc nn
Fenotipos
Negro
marrón
albino
albino
P:
CCnn
Fenotipos:
marrón
Gametos
diferentes:
{ Cn
F1
:
F2: (F1 x F1)
x
ccNN
albino
Cn}
Cc Nn -----> 100% negros
Cc Nn
Gametos
diferentes: {CN, Cn, cN, cn}
x
Cc Nn
{CN, Cn, cN, cn}
26
F2:
♂
♀
CN
Cn
cN
cn
PG.
1
2
1
2
4
2
1
2
1
CN
Cn
cN
cn
CCNN
negro
CCNn
negro
CcNN
negro
CcNn
negro
CCNn
negro
CCnn
negro
CcNn
negro
Ccnn
marrón
CcNN
negro
CcNn
negro
ccNN
marrón
ccNn
albino
CcNn
negro
Ccnn
marrón
ccNn
albino
ccnn
albino
Genotipos
CCNN
CCNn
CC nn
CcNN
CcNn
Ccnn
ccNN
ccNn
ccnn
P.F.
Fenotipos
9
negros
3
marrones
4
albinos
La proporción fenotípica 9:3:3:1 se modifica a 9:3:4
2.- Epistasis Dominante.- En este caso, un gen dominante es epistático a otro
dominante y a su alelo respectivo.
Ejemplo.- En perros, el gen ( l ) es epistático a los genes (N) para color
negro y (n) para color marrón, originando que el animal sea blanco. cómo
será la proporción fenotípica en F2?
Genotipos
Fenotipos
l_N_
blanco
l _ nn
blanco
ii N_
negro
ii nn
marrón
P
:
iinn
x
l l NN
Fenotipos:
marrón
blanco
Gametos
diferentes
(in
l N)
F1
:
F2
: (F1 x F1)
Gametos
diferentes:
Ii Nn -------> 100% blancos
Ii Nn
{IN , In, iN, in}
x
Ii Nn
{IN, In, iN, in}
27
F2:
♂
♀
IN
In
iN
in
P.G.
1
2
1
2
4
2
1
2
1
IN
IINN
blanco
IINn
blanco
IiNN
blanco
IiNn
blanco
Genotipos
IINN
IINn
Ilnn
IiNN
IiNn
Iinn
iiNN
iiNn
iinn
In
iN
IINn
blanco
IInn
blanco
IiNn
blanco
Iinn
blanco
IiNN
blanco
IiNn
blanco
iiNN
negro
iiNn
negro
in
IiNn
blanco
Iinn
blanco
iiNn
negro
iinn
marrón
P.F.
Fenotipos
12
blancos
3
negros
1
marrón
La proporción fenotípica se modifica a 12:3:1
28