eje córtico-límbico hipotálamo-hipófiso-tiroideo
Anuncio

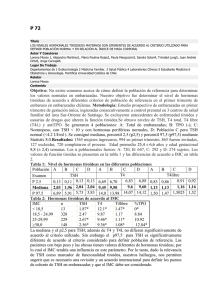
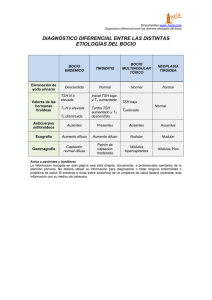
EJE CÓRTICO-LÍMBICO HIPOTÁLAMO-HIPÓFISO-TIROIDEO (C-L-H-H-T) La importancia de la función tiroidea en la actividad psíquica se destaca desde hace décadas tanto en la bibliografía endocrinológica como en la psiquiátrica, a tal punto que suele decirse que un funcionamiento tiroideo correcto es necesario para mantener la salud mental. Como dice Boullosa, el conocimiento de la implicancia psiquiátrica del eje tiroideo nos ayudará a comprender mejor los componentes biológicos de distintas entidades clínicas, el por qué de la asociación de hormonas tiroideas como coadyuvantes de algunos tratamientos psicofarmacológicos y a su vez los efectos colaterales de ciertos fármacos sobre su función. 1 Este eje se encuentra íntimamente ligado a la actividad noradrenérgica. La noradrenalina ejerce una función de estimulación sobre el mismo, favoreciendo la liberación hipotalámica de la hormona liberadora de tirotrofina (TRH). Como es habitual comenzaremos la descripción por los productos finales para ir luego a la categorización de los centrales y su regulación. HORMONAS TIROIDEAS La T3 es considerada como la verdadera hormona, comportándose la T4 como una prohormona, que podrá metabolizarse a T3 de acuerdo a las necesidades del organismo. La glándula tiroides segrega principalmente T4 y secundariamente T3 y T3 reversa (T3r), en casos especiales donde debe evitar la catabólisis. Ambas ejercen un mecanismo de retroalimentación negativa sobre el eje La T4 es la principal iodotironina de la circulación, que mantiene concentraciones séricas muy estables a pesar de las importantes variaciones tanto diarias como estacionales en la demanda de iodotironinas, para responder a las vicisitudes de la vida diaria como ingesta de carbohidratos, ejercicio, deprivación de sueño, exposición al frío y cambios lumínicos. La marcada estabilidad se debe a la sensibilidad y eficiencia de los mecanismos de feedback. En condiciones fisiológicas la proteína thyroxin-binding globulin (TGB) está unida al 70% de la T4 circulante, niveles que se mantienen estables durante toda la vida. El hígado genera la forma más activa, que es la T3, a través de un proceso de monodeionización (5’D) de la T4. La T3 es liberada a la circulación para actuar en los diversos tejidos. Los receptores nucleares para T3 fueron detectados en casi todos los tejidos, incluyendo corazón, cerebro, hígado, riñones y gónadas. Son receptores múltiples, codificados por distintos genes, y que varían según la región o el órgano en que se encuentren. Así mismo, se han descrito al menos tres sitios de unión de la T3 a la membrana plasmática Resumiendo: la hormona tiroidea es sintetizada ( a partir de 150 microgramos diarios de yodo inorgánico proveniente de la comida, incluida la sal de mesa que ha sido enriquecida con este mineral por la escasez en algunos suelos del país y la bebida) en las células epiteliales como tiroglobulina o T4, que como ya vimos es una prohormona. La tiroides tiene una enorme capacidad de almacenarla y una relativamente lenta capacidad para liberarla con una reserva suficiente para casi tres meses. No es habitual que describamos el metabolismo químico de las hormonas de cada eje, pero lo realizaremos aquí para comprender las diferencias entre los distintos tipos de hormonas tiroideas y el paso 2 de una a otra forma. El yodo inorgánico es oxidado a iodine o hipoiodito, dependiendo de una peroxidasa. El iodine es incorporado en residuos tirosínicos en la molécula de tiroglobulina. La iodinación de la tiroglobulina requiere de cuatro componentes: tiroglobulina, iodine, peroxidasa y peróxido de hidrógeno. De este proceso resultan la monoiodotirosina (MIT) y la diiodotirosina (DIT), a partir de las cuales se forman la T3 y la T4. Dijimos ya que el hígado genera la forma más activa de T3 ( a través de un proceso de monodeionización (5’D)) que es liberada a la circulación para actuar en los diversos tejidos. Sin embargo, muchos tejidos tienen cantidades propias de enzimas deiodinizantes de T4. La enzima formadora de T3 en los tejidos extrahepáticos es similar a la enzima 5’D hepática. La enzima hepática se conoce como “tipo I” y la extrahepática como “tipo II”. Una dieta deficiente en selenio (oligoelemento imprescindible para esta síntesis) provoca una disminución importante de la síntesis de T3. La enzima 5’D de tipo II tiene diferentes propiedades que la de tipo I. Se localiza en hipófisis, en cerebro (solo en hipotiroideos) y en tejidos que reciben fuerte inervación adrenérgica, como la grasa parda, la pineal y la médula adrenal. Siendo mucho menos abundante que la de tipo I, es muy activa en el hipotiroidismo y es inhibida por la T4 y por la T3r. Por lo tanto, en los tejidos que convierten la T4 en T3, la cantidad de T4 que ingresa en esos tejidos y la forma de enzima 5’D que es activada determinarán la habilidad del tejido para obtener la cantidad de T3 requerida para sus necesidades propias. Sin embargo, la cantidad de T4 disponible para cada tejido está también determinada por la actividad de otra enzima deiodinizante, denominada 5D o de “tipo III”, formadora de T3 reversa. La T3r, que podría ser inhibidora de la enzima 5’D, contribuye a la homeostasis, ya que puede determinar cuanta T4 pasará a T3. Por lo tanto existe la posibilidad de que la T3r sea el antagonista natural de la T3. Pero muchos investigadores dudan de la inactividad de la T3r, ya que es la hormona tiroidea predominante en el feto, donde la actividad tiroidea es fundamental para la maduración cerebral. Si se administra en animales de experimentación T3r, ésta es rápidamente destruida en varios tejidos. La T3 es un sustrato pobre para las enzimas que intervienen en su formación (5’D, tipo I y tipo II), pero es un excelente sustrato para la enzima formadora de T3r (5D, tipo III). La T3r, en cambio, es un excelente sustrato para la 5’D, especialmente la de tipo I. En cerebros de individuos hipotiroideos, donde la 5D está disminuida y la 5’D está estimulada, la limitada disponibilidad de T4 es máximamente utilizada para aumentar la formación de T3 y limitar su deionización. Estas moléculas de T3 son entonces útiles como ligandos para receptores de T3 nucleares o putativos o como sustrato para la activación de más T3. Así, si se protege a la T3 de la deionización, y se facilita la formación de ácido triiodotiroacético (triac), a partir de la T3. El triac tiene alta afinidad por el receptor nuclear de T3. Cuando se administra triac, éste se comporta como un débil agonista T3. Tiene vida media corta de manera que la oportunidad de unirse al núcleo es poca y la acción biológica es débil. Parece haber diferencias entre el triac endógeno y el exógeno. El endógeno podría sintetizarse cerca del núcleo. Triac es sólo uno de los posibles 3 productos con la cadena lateral de T3 alterada. Se comercializa como coadyuvante de tratamientos para perder peso, ya que su acción periférica es más importante que la central. Los receptores para T3 son nucleares y fueron detectados en casi todos los tejidos, incluyendo corazón, cerebro, hígado, riñones y gónadas. Son receptores múltiples, codificados por distintos genes, y que varían según la región o el órgano en que se encuentren. Son el producto del proto-oncogen c-erb A, el homólogo celular del oncogen viral conocido como v-erb A. Se reconocieron entonces, al menos dos clases de genes c-erb A, que producen los llamados receptores alfa y beta.. Existen a su vez las subclases alfa 1 y alfa 2 y beta 1 y beta 2, que tienen diferente distribución tisular, como así también diferentes características estructurales y de unión. Los alfa 1, beta 1 y beta 2 permiten la unión de la T3 pero el alfa 2 no lo hace. Este último tiene un sitio de unión trunco y no existe un ligando descubierto aún para el mismo. El beta 2 se encuentra principalmente en la hipófisis y es fuertemente regulado a la baja por la T3. Cuando la T3 llega al receptor nuclear, afecta la transcripción de ADN. Esto resulta en un aumento en la producción total de ARNm. Las hormonas tiroideas actúan entonces directamente en el código genético para modular la función de numerosos sistemas fisiológicos. Se han descripto al menos tres sitios de unión de la T3 a la membrana plasmática. Uno de ellos, tiene gran afinidad por la T3 y se asocia con cambios inmediatos en la glucosa celular, el calcio y las funciones de transporte de aminoácidos. Estas respuestas se modifican en presencia de noradrenalina. La transducción de la información se realiza a través del calcio. Otra categoría de receptores median la endocitosis de T3 en los fibroblastos a través de mecanismos reminiscentes de aquellos descriptos para la lipoproteína de baja densidad (LDL). El tercer tipo de receptores de membrana está unido a la familia calcio ATPasa de proteínas transmembrana y es estimulado por la unión de T4 y de sus análogos. HORMONA LIBERADORA HIPOFISARIA O TSH Aunque la TRH y los niveles circulantes de T4 son los principales coordinadores de la síntesis y liberación de TSH, existen numerosos mecanismos que regulan de forma inhibitoria la secreción de TSH. Entre ellos debemos mencionar al cortisol, la serotonina, la testosterona, la somatostatina o "paninhibina" y la dopamina. Los efectos de la TSH están influenciados por la transducción del mensaje noradrenérgico vía receptores beta y alfa de las paredes de los vasos sanguíneos y las membranas plasmáticas de las células foliculares tiroideas. La TSH presenta una secreción pulsátil cuyo pico nocturno se produce entre las 0 y las 3 AM, aunque algunos autores niegan la mencionada pulsatilidad. El pico nocturno de liberación de TSH en individuos 4 sanos es independiente del cambio de hormonas circulantes o de sustancias centrales activas conocidas y requiere un ciclo de sueño-vigilia normal. Si el estado de vigilia se mantiene por 24 horas, el pico de TSH se modifica. La resultante final de la acción de la TSH sobre la glándula tiroidea es la secreción de T3 (tri-yodotironina) y de T4. HORMONA LIBERADORA HIPOTALÁMICA O TRH La TRH es un tripéptido que en animales inferiores ejerce funciones noradrenérgicas y que tiene un rol importante en el despertar de animales que hibernan. Se encuentra distribuido en todo el SNC habiéndose descripto una amplia distribución de receptores. Es liberada desde el terminal sináptico en la eminencia media en forma pulsátil, pasa a la circulación portal hipotálamo-hipofisaria y alcanza las células hipofisarias tirotropas. Los receptores hipofisarios para TRH transmiten el constante cambio de TRH hacia el aparato sintetizador de TSH, vía un segundo mensajero calcio-fosfoinositol, lo que resulta en un apropiado aumento o disminución de liberación de tirotrofina (TSH) hipofisaria. Además de esta acción liberadora de TSH, la TRH presenta un efímero efecto antidepresivo per se; también se comporta como factor liberador de prolactina (PRF) y tiene una acción directa, estimulando la liberación de la prohormona tiroidea levotiroxina o T4. En ciertas circunstancias también estimula la liberación (y posiblemente la síntesis) de otras hormonas hipofisarias, como la hormona de crecimiento (GH). El efecto sobre la prolactina es totalmente independiente del efecto sobre la TSH. La hiperprolactinemia del hipotiroidismo primario probablemente se deba al TRH elevado. La TRH tiene efectos directos sobre la función cerebral, independientes de su acción sobre el eje tiroideo. Estos incluyen el revertir de la acción de drogas sedativas o anestésicas, la estimulación de la actividad locomotora y efectos sobre las funciones cardiorrespiratoria, gastrointestinal y neurológica. Tiene también un efecto antidepresivo transitorio. EJE INMUNO-TIROIDEO La TRH regula en forma positiva a la prolactina y estimula la síntesis linfocitaria tanto de TSH como de su receptor. Es decir, que tiene capacidades inmuno-protectoras. A su vez, las IL-1 e IL-6) y el factor de necrosis tumoral (FNT) producen inhibición de la secreción de TSH por aumentar la síntesis de somatostatina. La IL-1alfa y la IL-1beta inhiben la incorporación y liberación de iodo. Concentraciones altas de FNT correlacionan con bajas de T3. El interferón gamma (IFN) también inhibe la función 5 tiroidea, situación que es concentración-dependiente. Todas las citoquinas antes mencionadas actúan sinérgicamente para inhibir la función tiroidea. Consignemos que los ejes CLHHT y prolactínico son inmunoestimulantes, en contraposición al eje CLHHA que es inmunosupresor. En la entrega N° 11, al detallar el sistema inmune comprenderemos mejor esta interrelación. ESTUDIO FUNCIONAL DEL EJE C-L-H-H-T El dosaje de hormonas básales y de TSH es clásico en endocrinología. Con estas determinaciones se diagnostican diferentes tipos de hipo e hipertiroidismo, cuyas características clínicas describiremos más adelante. El hipotiroidismo primario puede dividirse en tres grados. El de grado I presenta niveles de T4 libre basal disminuidos, TSH basal aumentada, test de TRH/TSH con hiperrespuesta y numerosos signos y síntomas clínicos clásicos.. En el de grado II, la T4 libre basal es normal, la TSH basal se encuentra aumentada, el test de TRH/TSH presenta hiperrespuesta y existe por lo menos un síntoma o signo de hipotiroidismo. El de grado III presenta hormonas tiroideas y TSH básales normales, no tiene síntomas ni signos claros y sólo la hiperrespuesta en el test de TRH/TSH permite hacer el diagnóstico (la describiremos en breve) Finalmente, algunos autores refieren otro subtipo de hipotiroidismo subclínico, sin síntomas ni signos clínicos, con hormonas basales normales y prueba TRH/TSH también normal pero con la presencia de anticuerpos antitiroideos. Esto podría involucrar una vulnerabilidad inmunológica. La funcionalidad del eje se estudia mediante la prueba de estimulación TRH/TSH. Consiste en la medición basal de TSH por radioinmunoensayo, la aplicación de 200 a 500 microgramos de TRH vía endovenosa y la posterior medición de TRH a los 25 y a los 90 minutos. En condiciones normales el valor basal de TSH no debe ser superior a 6.5 microUI/ml. A los 25 minutos de aplicada la TRH debe, por lo menos, duplicar el valor basal y no superar el valor de 25 microUI/ml (estandarizado para población argentina) para volver, a los 90 minutos, a valores cercanos a los básales. Se considera hiperrespuesta cuando el valor a los 25 minutos es superior a 25 microUI/ml, observándose en pacientes con hipofunción tiroidea clínica o subclínica. Para determinar si una curva es plana se calcula el delta TSH. Se trata del valor resultante de restar el valor basal de TSH al valor máximo de la misma. Normalmente, el resultado debe ser superior a 5 o a 7, según los distintos autores. Si los valores son inferiores, se considera que la curva es plana o “blunted”, probablemente debido a un hipotono noradrenérgico o disbalance 5-HT/NA, a un aumento de cortisol secundario al aumento de CRH (que 6 inhibe a la hipófisis), a un aumento de somatostatina (que también inhibe a la hipófisis), o a una "downregulation" de receptores hipofisarios para TRH secundario al aumento de TRH. Muchos factores alteran la interpretación del TRH/TSH: la edad avanzada y el sexo masculino pueden disminuir la respuesta; también el ayuno, fallas renales y hepáticas y el síndrome de Klinefelter. La dopamina, los glucocorticoides, la neurotensina y la somatostatina pueden aplanar la curva. También pueden alterar el resultado del test la administración de litio y de algunos antipsicóticos y antirrecurrenciales. Si el valor del delta TSH es inferior a 1, existe un alto riesgo de conducta suicida. Algunos autores rescatan también el concepto de “delta-delta TSH”, que es la diferencia entre el delta TSH en el momento de la recuperación menos el valor de delta TSH en el momento de inicio de la enfermedad depresiva. Cuando este valor es inferior a 2 predice la posibilidad de recaída depresiva temprana. Para otro grupo de autores, el delta-delta TSH corresponde a la diferencia entre el delta TSH de las 11 PM y el delta TSH de las 8 AM. Con respecto a la aplicación endovenosa de TRH, debemos tener en cuenta que se trata de una sustancia que posee una vida media de aproximadamente 6 minutos y que puede producir, coincidiendo con el pico plasmático, náuseas, urgencia miccional, rush facial, molestias abdominales, embotamiento e hipertensión arterial. La prueba está contraindicada en hipertensión arterial, cardiopatías, daño cerebral, hemiplejía y epilepsia. Esto último se debe a la posibilidad de la TRH de desencadenar convulsiones. APLICACIÓN CLÍNICA DE LAS ALTERACIONES DEL EJE CLHHT La hiperactividad del eje límbico-hipotálamo-hipófiso-adrenal en los desórdenes afectivos ha sido bien documentada en la literatura y esbozada en las entregas anteriores. Con respecto al eje tiroideo, es bien sabido que los glucocorticoides inhiben a la TRH y a la TSH y disminuyen la producción de T3, por lo cual se dedujo, en principio y se probó luego la alteración de este eje en patología psiquiátrica. La respuesta de alteración tiroidea más reproducida es la presencia de curvas de respuesta plana de TSH tras la estimulación con TRH en depresión. Durante muchos años se describió que entre el 25 y el 30% de los pacientes depresivos unipolares presentaban la mencionada alteración comportándose como marcador de rasgo y expresando claramente una vulnerabilidad hacia la depresión. Los sujetos con conducta violenta o suicida tienden a presentar los niveles más altos de TRH dentro del grupo de los depresivos. Una disminución en la variación circadiana de TSH se asocia generalmente con depresión primaria. También, cuanto más severo es el cuadro depresivo, se ve mayor disminución de TSH nocturna. 7 En la depresión psicótica, la disminución de T3 es más frecuente acompañada a menudo con aumento de T4, como mecanismo compensatorio en el marco de un hipertiroidismo relativo. Según algunos autores, el aumento de T4 que se observa en la depresión podría ser un mecanismo compensatorio del eje para mantener la homeostasis afectiva y puede predecir la respuesta a los antidepresivos. Pero existe una explicación alternativa: la depresión es un estado de relativo hipertiroidismo, asociándose con aumentos relativos de los niveles circulantes de T4. Se requieren sustanciales pero limitadas disminuciones de T4 para obtener la respuesta antidepresiva. En otras palabras, los aumentos relativos de T4 en la depresión se interpretan como compensatorios en la primer teoría expuesta, y como patológicos en la segunda. La T3r sérica está disminuida en la depresión y normaliza con la recuperación clínica. Los pacientes depresivos pueden presentar, además de la curva TRH/TSH plana, un aumento de la producción de T4. Los niveles de T4 después del tratamiento antidepresivo disminuyen, pero dentro del rango normal. La buena respuesta a antidepresivos o antirrecurrenciales correlacionan con disminución de la T4 dentro del rango normal, produciéndose un cambio significativo en la relación T4-T3 después de la recuperación clínica. Este cambio no implica un efecto consistente sobre la T3 o sobre la TSH. La mayor parte de los trabajos de la literatura reportan curvas planas en depresión mayor con hormonas basales tiroideas normales que no normalizan con tratamiento antidepresivo solamente . En los estados depresivos con excesiva anorexia, los niveles de T4 suelen encontrarse dentro de límites normales pero con niveles subnormales de T3, secundario a una disminución de la conversión periférica de T4 a T3. Tanto los fármacos antidepresivos como la terapia electroconvulsivante (TEC) producen modificaciones en el eje tiroideo. El TEC produce disminución tanto de T4 como de T3. El litio también afecta la producción de hormonas tiroideas, comportándose como un potente inhibidor de la conversión de T4 a T3 además de presentar efectos inhibitorios sobre los niveles de TRH y de TSH. La carbamacepina puede disminuir tanto los niveles de T3 como los de T4 sin modificar los básales de TSH. La T3 acelera la respuesta a los antidepresivos, sobre todo en mujeres, pudiendo convertir en respondedores a pacientes no respondedores; es mucho más efectiva que la T4. La T4 sería de elección en el tratamiento de pacientes bipolares. Se sugiere que altas dosis de T4, suficientes para inducir un estado de hipertiroidismo químico, pueden disminuir la frecuencia y la severidad de los ciclos, particularmente en los cicladores rápidos. Las hormonas tiroideas potencian a las catecolaminas porque aumentan la sensibilidad de los receptores beta adrenérgicos; los beta bloqueantes tienden a su vez a suprimir la función tiroidea. La T3 puede aumentar la cardiotoxicidad de los antidepresivos tricíclicos. Repitamos que la respuesta plana a la prueba de TRH-TSH es el marcador biológico más fidedigno del compromiso de este eje en desordenes afectivos o en depresiones comórbidas a otras patologías. Esta reacción refleja anormalidades en la función cerebral, anormalidades endócrinas no sólo del eje tiroideo 8 sino también del eje adrenal y se debe, al menos en parte, a la hipersecreción crónica de TRH. Existe correlación entre la curva plana y la cronicidad, la ansiedad y la conducta suicida violenta. En el lado opuesto, la presencia de una curva hiperrespondiente, en presencia de hormonas y TSH basales normales, pone de manifiesto la existencia de un hipotiroidismo subclínico frecuentemente asociado a la presencia de anticuerpos antitiroideos. La causa más común de hipotiroidismo subclínico es la tiroiditis autoinmune. El hipotiroidismo subclínico, en presencia de anticuerpos antitiroideos, progresa hacia el hipotiroidismo clínico en razón del 5 al 8% por año. Se reportó alta prevalencia de hipotiroidismo clínico en mujeres bipolares de rápida ciclación. Durante muchos años se describió que aproximadamente el 15% de los pacientes depresivos bipolares cicladores rápidos presentaban la mencionada alteración de la curva, comportándose como marcador de estado que aporta información sobre el diagnóstico, pronóstico, evolución y tratamiento del cuadro de base. Hace muchos años publicamos un estudio preliminar sobre 34 pacientes mostrando 305 de curvas de hiperrespuesta en pacientes depresivos con evolución a la bipolaridad y presencia elevada de anticuerpos antiperoxidasa en los mismos. Años después, en 1000 pacientes, Flores Amargos obtiene la misma respuesta al tratamiento antidepresivo en unipolares y bipolares, objetivando a las hiperrespuestas TRH-TSH como predictores de switching. Ambos conceptos están hoy controvertidos. Tal vez, donde cobre mayor interés la toma de la respuesta a la estimulación TRH-TSH es en los "debuts" depresivos de población infanto-juvenil donde la probabilidad de intuir un cuadro bipolar es crucial para la administración temprana de antirrecurrenciales. En un trabajo con 15 adolescentes obtuvimos curvas de hiperrespuestas en 3 pacientes coincidiendo con antecedentes genéticos de bipolaridad y su normalización con tratamiento con carbamacepina, 5 con respuesta plana y 7 con respuesta normal. Siempre, la no normalización de la curva de TSH por estimulación con TRH predice una recaída temprana e indica la necesidad de tratamiento de mantenimiento. Pueden tener también curvas TRH/TSH planas los trastornos de personalidad borderline y los síndromes de dolor crónico Con respecto a los trastornos de ansiedad, la disfunción tiroidea puede estar directamente relacionada con síntomas de esta vertiente. Pero pacientes con ataques de pánico suelen tener función tiroidea normal y aquéllos que presentan curva plana son los que se acompañan de sintomatología depresiva concomitante. El agregado de T3 no produce mejoría de los síntomas de ansiedad. En los trastornos de la alimentación este eje presenta respuestas particulares. Numerosos autores han reportado una disminución o falta de cambios de la TSH basal o incluso curvas planas o normales durante la starvation. Por otra parte, la sobrenutrición resulta en un aumento de la concentración y producción de 9 T3 y no la esperada respuesta plana a la TRH. La sensibilidad intrahipofisaria a la T3 está normal en el ayuno, lo que implica que el mecanismo de feed-back de T3 se conserva intacto. La aparente falta de respuesta de la TSH a niveles periféricos bajos de T3 durante el ayuno puede explicarse por un aumento de la conversión intrahipofisaria de T4 a T3. La disminución de la glucemia que se encuentra en la desnutrición provoca un aumento en la secreción de hormona de crecimiento (GH), que a su vez permite el aumento de somatostatina. El aumento de somatostatina provoca, a su vez, disminución de la secreción de TSH. También el efecto alfa adrenérgico, la serotonina y la insulina están implicados en la regulación de la TSH. Si bien ninguno de ellos es suficiente para explicar los cambios en el estado de nutrición alterado, el mismo probablemente se trate de una combinación de varios factores neuroendócrinos. Es decir que debemos conceptualizar que con respecto a los trastornos de la alimentación, la restricción calórica a la que se someten los pacientes con anorexia nerviosa resulta en una disminución de los niveles de T4 (pero dentro del rango normal) y de T3 circulantes y en un aumento de los niveles de T3r. Estos cambios transitorios forman parte del denominado patrón de “enfermedad eutiroidea” y pueden normalizarse luego de un corto período. Los patrones tiroideos referidos se deberían simplemente a mecanismos fisiológicos de adaptación a la starvation, relacionado siempre con condiciones en las cuales existe una rápida disminución de peso. Se produce entonces un paso preferencial de T4 a T3r como mecanismo para conservar energía, mientras los niveles de TSH se encuentran en rango normal bajo. La concentración de T3 y de T4 puede relacionarse con el nivel circulante de sustancias tales como carotenos, asociándose la hipercarotinemia con niveles descendidos de T3 y de T4. Los cuadros de bulimia nerviosa pueden cursar con alteraciones que no necesariamente dependen de la starvation ni del aumento de peso. Así, es frecuente detectar niveles plasmáticos elevados de ácido beta hidroxibutírico y de ácidos grasos libres, como así también bajos de T3 y de glucosa y disminución de la respuesta de la noradrenalina a un cambio ortostático. Las bulímicas presentan un estado nutricional más estable que las anorécticas, con menor necesidad de disminuir la conversión periférica de T4 a T3. Pueden encontrase también bajos niveles séricos de TSH. Las bulímicas en estado de remisión pueden presentar un estado químico de hipotiroidismo debido a una anormalidad hipotalámica en la neurona TRH y a que durante las conductas de atracones y de purgas puede haber activación del eje tiroideo, causando una aparente normalización de los índices tiroideos. Una respuesta anormal a la TRH es más frecuente en el grupo de pacientes restrictivas en bajo peso que en el de peso normal. Las anormalidades tiroideas se relacionan más comúnmente con el estado de semi-starvation de la paciente bulímica. Recordemos que una ingesta baja en carbohidratos puede estar seguida por una curva plana en los pacientes con bulimia nerviosa. La disminución de la respuesta de la prolactina a la TRH se debe también al bajo consumo de hidratos de carbono. El abuso de hormonas tiroideas exógenas como forma de aumentar el metabolismo para bajar de peso también deben tenerse en cuenta en toda paciente con desórdenes de la alimentación. 10 En el alcoholismo, los datos reportados en la literatura son escasos y frecuentemente influenciados por el estado nutricional o afectivo concomitante. De todas formas es importante consignar que el etanol no afecta en forma aguda al eje tiroideo. Los niveles de T4 están disminuidos en el alcoholista, si se trata de pacientes que recayeron, sugiriendo que el etanol produce supresión de T4. Durante la fase aguda de abstinencia alcohólica, existe una activación tiroidea puesta de manifiesto a través de una curva TRH/TSH plana, aumento de TSH y de T4 básales, acompañados también de disminución de hormona de crecimiento y de prolactina debidos al aumento de la actividad dopaminérgica central. Los niveles de T3 están disminuidos en pacientes alcoholistas que presentan daño hepático, debido a una disminución de la conversión de T4 a T3. En estos casos, el aumento de T3 se asocia con mejoría clínica, mientras que su disminución se relaciona con riesgo de muerte. Se ha reportado también la disminución del volumen de la glándula tiroidea, acompañado de un aumento de la fibrosis de la misma. La TRH produce disminución del efecto depresivo causado por el etanol y probablemente aumenta sus acciones ansiolíticas. En las esquizofrenias, los estudios publicados son muy limitados e incluso contradictorios. Algunos autores reportan una elevación transitoria de T4 en pacientes psicóticos agudos recientemente internados. También, antiguos trabajos no replicados mostraban aumento de LATS (long acting thyroid stimulator) en esquizofrénicos aunque carezcan de los signos clínicos de exoftalmos que conlleva la presencia patológica de esta sustancia. En otros subtipos de esquizofrénicos se ha observado un aplanamiento de la curva tiroidea. Un número importante de pacientes presenta disminución de su función tiroidea. Se cree que la curva plana en el paciente esquizofrénico predice una mejor respuesta a los antipsicóticos y se asocia con una más rápida respuesta. Es necesario dejar consignado que el hipertiroidismo aumenta y el hipotiroidismo disminuye la sensibilidad a los antagonistas dopaminérgicos En cuanto a las enfermedades endocrinológicas (a las cuales no nos dedicamos en otras entregas sobre otros ejes PNIE, pero sí aquí, por ser productoras "per se" de sintomatología psiquiátrica), consignemos brevemente las características clínicas de la hipofunción y la hiperfunción glandular. El Hipertiroidismo puede deberse a un tumor hipofisario hipersecretor de TSH, a una falta de regulación hormonal, debida a la producción de anticuerpos antitiroideoas, que provoca un aumento de T3 y T4 (enfermedad de Graves), a la existencia de tejidos tiroideos ectópicos o a un origen medicamentoso (T3 y/o T4 exógenas). El hipertiroidismo primario es aquél que, debido a una hipersecreción de hormona tiroidea, cursa con niveles disminuidos de TSH debido al freno que ejerce el mecanismo de retroalimentación negativa. El cerebro, en colaboración con el hígado, también hace ajustes homeostáticos en el hipertiroidismo ya que la captación y el turn-over de T3 y de T4 en distintos tejidos están marcadamente aumentados. 11 La clínica del hipertiroidismo incluye, además de un cuadro de agitación motora, sintomatología cognitiva similar a la de la depresión (dificultad para concentrarse, fallas de memoria y despersonalización). Es decir, psiquiátricamente se corresponden a cuadros de depresión ansiosa sintomática y no de mania. El diagnóstico diferencial con manía obliga a detectar, en el caso del hipertiroidismo, la disminución de energía, el insomnio pero con necesidad de dormir, la ansiedad y/o agitación, la labilidad, la pérdida de peso y la disforia e irritabilidad similares a lo que se encuentra en algunos pacientes depresivos agitados, que en caso de ancianos pueden ser apáticos y que poco tienen que ver con auténticos episodios maníacos. Es de capital importancia destacar que, a la inversa del cuadro maníaco, el hipertiroidismo es un cuadro básicamente displacentero. Muchos signos periféricos de tirotoxicosis son consistentes con hiperfunción noradrenérgica y mejoran con antagonistas beta adrenérgicos. La tirotoxicosis se asocia con aumento del binding del receptor beta adrenérgico, aumento del binding del receptor alfa-2 adrenérgico y disminución del turn-over de noradrenalina. Las hormonas tiroideas inhiben la captación de GABA, facilitando la transmisión gabaérgica. En el Hipotiroidismo secundario o terciario (la disfunción primaria se encuentra a nivel de hipotálamo o de hipófisis) el laboratorio muestra valores basales de TSH y de hormonas tiroideas disminuidos, debido a una falta de estímulo. En cambio, si la disfunción primaria es a nivel tiroideo, la TSH basal se encuentra aumentada debido a la falta de la retroalimentación negativa que debería ejercer la hormona tiroidea sobre el eje. Simultáneamente, los niveles elevados de TRH son los responsables del aumento de prolactina, típico del hipotiroidismo. El hipotiroidismo subclínico está presente en el 5% de la población, sobre todo en mujeres mayores de 60 años. Se describe una relación inversa entre la búsqueda de sensaciones y elevados niveles de TSH. Pueden presentar disminución de energía y lasitud y confundirse con depresiones refractaria. Si este hipotiroidismo se prolonga crónicamente puede producir disfunción cognitiva irreversible. Es típico encontrar enlentecimiento paroxismal en el EEG. Es necesario dejar consignado aquí que, ante la hipofunción tiroidea la disponibilidad de hormona prioriza al cerebro, debido a que la hipofunción es responsable de anormalidades regionales en el crecimiento de células nerviosas y de alteraciones en la sinaptogénesis. Se cree que los efectos de la hormona tiroidea en el cerebro son bien evidentes después del parto. El feto hipotiroideo parece estar protegido por hormonas placentarias si la madre tiene función tiroidea normal y no hay deficiencia de yodo. Luego del parto, el feto sólo recibe pequeñas cantidades de hormona, 12 provenientes de la leche. De manera que las peores consecuencias de este hipotiroidismo pueden prevenirse haciendo el diagnóstico inmediatamente después del parto. A partir de esto, las hormonas tiroideas están consideradas como el mayor regulador de la maduración cerebral normal. Así, el hipotiroidismo perinatal tiene severas consecuencias para la maduración del SNA. La respuesta adrenocortical al stress y el desarrollo de los receptores hipocampales de glucocorticoides están marcadamente influenciados por la concentración de hormona tiroidea durante la vida postnatal temprana. La función tiroidea fetal puede ser afectada por factores no específicos como fumar durante el embarazo o por la ingesta materna de alcohol, como así también por la dieta o por factores genéticos. Nuevamente omitiremos la descripción de los síntomas y signos típicos del hipotiroidismo por estar fuera de los objetivos del presente capítulo, pero es sabido que pueden originarse cuadros que comprenden desde alteraciones de la personalidad hasta psicosis franca. Luego de una tiroidectomía, el hígado desiste de convertir la T4 en T3, permitiendo que lo que está disponible sea usado por otros tejidos con capacidad 5´D. Esto parecería favorecer aún más el hipotiroidismo. Sin embargo, la razón biológica de este hecho es favorecer al cerebro para la formación de T3 a partir de T4. Estos mecanismos ubican al cerebro entre los tejidos más activos para formar y utilizar T3 durante la hipofunción tiroidea. Por esta razón, en el hipotiroidismo los niveles de T3 y T4 cerebrales están sólo levemente disminuidos. El hipotiroidismo es un factor de riesgo para el desarrollo del trastorno bipolar ciclador rápido. Un relativo déficit central de hormona tiroidea predispone a la rápida ciclación, aun en presencia de T4 sérica normal. En las enfermedades autoinmunes, las células foliculares tiroideas son capaces de presentar antígenos en la superficie celular, permitiéndole a la peroxidasa generar la producción de anticuerpos. Anticuerpos antiperoxidasa, antimicrosomal, antitiroglobulina y anti-receptor TSH están implicados en tiroiditis autoinmunes. La tiroiditis autoinmune de Hashimoto puede presentarse con hiper o hipotiroidismo. En algunos casos cursa con síntomas psiquiátricos en ausencia de hipotiroidismo clínico o subclínico. Los más comunes incluyen depresión, dificultades para concentrarse e irritabilidad. No todos los pacientes presentan anticuerpos antitiroideos. Aun si los niveles de T4 son normales, resulta apropiado estudiar anticuerpos antimicrosomales. La enfermedad autoinmune tiroidea puede darse también como una enfermedad de Graves, que suele ir precedida de un período de hipotiroidismo, indistinguible del de la tiroiditis de Hashimoto. 13 BIBLIOGRAFÍA COMPLEMENTARIA SUGERIDA Balhs Sc, Amaral de Carvalho G. The thyroid fuction and depression. A review. University of Curitiba. Brazil. Publicado en www.scielo.br. 2005 Bauer M.S., Droba M., Whybrow P.C: Disorders of the Thyroid and Parathyroid. Chap. 2. In: Nemeroff Ch.B., Loosen E.T.: Handbook of Clinical Psychoneuroendocrinology. Ed. Wiley J & Sons. New York. USA. 1987. Boullosa O. Eje cortico-limbico-hipotálamo-hipófiso-tiroideo en: Lopez Mato. Psiconeuroinmunoendocrinologia . Editorial Polemos., Buenos Aires, Argentina. 2002. Boullosa Oscar; Márquez Coral; López-Mato Andrea et al. TRH/TSH en población infantojuvenil. Revista de Comunicaciones. AAP. IV, (7). 1997. Cody V. Thyroid hormone interactions: molecular conformation, protein binding, and hormone action. Endocr Rev. 1. 1980. Esposito S, Prange AJ Jr, Golden RN. The Thyroid Axis and Mood Disorders: Overview and Future Prospects. Psychopharmacology Bulletin. 1997 Haddow JE, Palomaki GE, Allan WC, Williams JR et al Maternal thyroid deficiency during pregnancy and subsequent neuropsychological development of the child. N Engl J Med. 1999. Jackson IMD, Asamoah EO. Thyroid Function in Clinical Depression: Insights and Uncertainties. Thyroid Today 22(2). 1999. Joffe R., Levitt A.: The Thyroid Axis and Psychiatric Illness. Ed. American Psychiatric Press. Washington, USA. 1993. Kirkegaard C, Faber J. Influence of free thyroid hormone levels on the TSH response to TRH in endogenous depression. Psychoneuroendocrinol 11. 1986 Kirkegaard C, The role of thyroid hormones in depression. Eur J Endoc.138. 1998 Kirkegaard C. The thyrotropin response to TRH in endogenous depression. Psychoneuroendocrinol .1981. Kjellman BF, Beck-Friss J, Ljunggren J-G, Wetterberg L. 24 serum levels of TSH in affective disorders. Acta Psychiatr Scand 69. 1984. Kraus RP, Phoenix E, Edmonds MW, Nicholson IR et al. Exaggerated TSH responses to TRH in depressed patients with ''normal'' baseline TSH. J Clin Psychiatry 58. 1997. Lechan R.M., Toni R.: Thyrotropin-Releasing Hormone Neural Systems in the Central Nervous System. (Chap. 10) In: Nemeroff Ch.B.. Neuroendocrinology. Ed. CRC Press. Florida. USA. 1992. 14 Linnoila M, Lamberg B-A, Potter WZ, et al High reverse T3 levels in manic and unipolar depressed women. Psychiatry Res. 6. 1982 Loosen P.T.: The TRH Test Stimulation Test in Psychiatric Disorders: A Review Chap. 15. In: Nemeroff Ch.B., Loosen E,T.: Handbook of Clinical Psychoneuroendocrinology. Ed. Wiley J & Sons. New York, USA. 1987. López-Mato A; Boullosa O; Segretin, G.El Test TRH- TSH en Depresión Endógena. Implicancias Diagnósticas y Terapéuticas. Alcmeon, Año VI vol. 4 1996. McEwen B.S.: Effects of the Steroid/Thyroid Hormone Family on Neural and Behavioral Plasticity. Chap. 11. In: Nemeroff Ch.B.: Neuroendocrinology.. Ed. CRC Press. Florida. USA. 1992. Moller HG; Flores Amargos D. Respuesta a antidepresivos en pacientes unipolares y bipolares y el test TRH-TSH como predictor de switching (no textual) presentado en Congreso Internacional dela WFSBP. Berlin. 2001 Nemeroff CB, Simon JS, Haggerty JJ, Evans DL. Antithyroid antibodies in depressed patients. Am J Psychiatry. 142. 1985 Reed Larsen P., Davies T.F., Hay I.D.: The Thyroid Gland. Chap. 11. In: Wilson J.D., Foster D.W., Kronenberg H.M., Reed Larsen P.: Williams Textbook of Endocrinology (9th Edition). Ed. W.B. Sanders. Philadelphia, USA. 1998. Shelton RC, Winn S, Ekhatore N, Loosen PT. The effects of antidepressants on the thyroid axis in depression. Biol Psychiatry. 33. 1993. Targum SD, Sullivan AC, Byrnes SM. Compensatory pituitary-thyroid mechanisms in major depressive disorder. Psychiatr Res. 6. 1982. Winokur A.: The Relevance of Thyrotropin-Releasing Hormone to Psychiatric Disorders. (Chap. 2). In: Nemeroff Ch.B.: Neuropeptides and Psychiatric Disorders. Ed. American Psychiatric Press. Washington DC, USA. 1991. 15