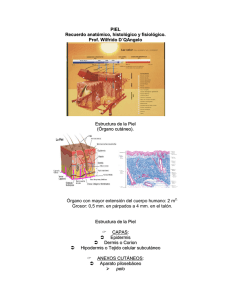
Células de Langerhans: detectar el medio ambiente en la salud y la enfermedad En las últimas décadas, nuestra comprensión de las células de Langerhans (LC) ha cambiado drásticamente en base a los hallazgos novedosos con respecto al origen del desarrollo y las funciones biológicas de estas células inmunes residentes específicas de la epidermis. Se ha hecho evidente que las LC no solo desempeñan un papel fundamental en la vigilancia inmune y la homeostasis, sino que también tienen un impacto en la patología ya sea induciendo tolerancia o mediando inflamación. Sus capacidades únicas de autorenovarse dentro de la epidermis, a la vez que pueden migrar a los ganglios linfáticos para presentar el antígeno, colocan las LC en una posición clave para muestrear el entorno local y decidir la respuesta inmune cutánea apropiada. Los nuevos datos emocionantes que distinguen las LC de las células dendríticas dérmicas Langerin + (DC) en un nivel funcional y ontogénico revelan roles cruciales para las LC en trauma y diversas patologías de la piel, que serán discutidas a fondo aquí. Sin embargo, a pesar del rápido progreso en el campo, el papel exacto de las LC durante las respuestas inmunes no se ha dilucidado completamente.Esta revisión se centra en los modelos de ratón que se han desarrollado para permitir el estudio de las LC murinas y otras DC que expresan Langerin, y nos han enseñado sobre el desarrollo y la función de LC. Palabras clave: células de Langerhans, inmunología de la piel, microambiente tumoral, marcha atópica, modelos de ratón INTRODUCCION Paul Langerhans describió por primera vez las células de Langerhans (LC) en 1868. Estas células se identificaron en la epidermis y recuerdan a las neuronas debido a su morfología dendrítica. Más de un siglo después, Ralph Steinman descubrió la presentación del antígeno por las células dendríticas (CD) ( 1 ). Las CL se clasificaron posteriormente como CD ( 2 ) que se localizan en la capa más externa de la piel, concretamente la epidermis, donde residen en estrecha asociación con queratinocitos, el tipo de célula epidérmica principal. Las LC también están presentes en otros epitelios estratificados, como el epitelio oral y vaginal de la mucosa. Estudios recientes muestran que estas LC de la mucosa actúan funcionalmente como LC genuinas, aunque se originan a partir de diferentes precursores ( 3 ). En esta revisión, nos centramos exclusivamente en las LC presentes en el epitelio cutáneo. Los queratinocitos se someten a una forma especializada de muerte celular, lo que resulta en la formación de las capas cornificadas de la piel que son cruciales en el establecimiento de una barrera de permeabilidad epidérmica ajustada. Establecer una barrera cutánea adecuada es un requisito previo para la vida terrestre, ya que protege al cuerpo de la deshidratación y la invasión de patógenos. Además de las células estructurales como los queratinocitos, la epidermis alberga diferentes tipos de células inmunitarias, a saber, LC, células T residentes de tejido y células T γδ (células T epidérmicas dendríticas, solo en ratones) que constituyen la barrera inmunológica de la piel. Tanto los queratinocitos como las células inmunes son esenciales para detectar el entorno y funcionar como una primera línea de defensa contra los insultos externos ( 4 ). Como tal, las LC han demostrado ser cruciales en diversas respuestas antimicrobianas, que se revisan en detalle por West et al. en este tema de investigación ( 5 ). Durante mucho tiempo, las LC se consideraron como las células presentadoras de antígeno exclusivas en la piel y un cuerpo de la literatura atribuyó un papel a las LC en la patogénesis de diversas enfermedades de la piel. Sin embargo, otros subtipos de DC residen en la dermis y una plétora de evidencia reciente desafió el paradigma que centra las LC como el único tipo de célula responsable del cebado de las células T a los antígenos de la piel. Las LC difieren ontogénicamente de las CD dérmicas, ya que las LC comparten un precursor común con los macrófagos, mientras que las DC dérmicas están más relacionadas con los subconjuntos convencionales de DC (cDC) presentes en los tejidos linfoides ( 6 ). Funcionalmente, parece haber un alto nivel de redundancia entre las LC y las DC dérmicas, cuyos fenotipos y funciones específicas se revisan en otra parte ( 7 , 8 ). Las LC pueden migrar al ganglio linfático que drena la piel, que se requiere para el cebado de células T clásico. Las LC han sido recientemente implicadas en reacciones cutáneas inmunosupresoras locales y en la activación de las células T de memoria residente en la piel, lo que indica un papel importante de las CL en la mediación de la fase adaptativa de la inmunidad de la piel ( 9 , 10 ). Además, a diferencia de las CDC, las LC tienen un origen embrionario y se mantienen en gran medida mediante la autorrenovación. En los últimos años, ha habido un gran debate sobre la clasificación de las LC. Utilizando el origen del desarrollo como un factor de categorización, que se fomenta en el campo complejo del sistema de fagocitos mononucleares ( 11 ), es razonable clasificar CL en el linaje de los macrófagos. La expresión del factor de transcripción Zbtb46 refuerza la identidad de cDC ( 12 ).Recientemente, un estudio de rastreo de linaje altamente elegante corroboró la identidad dual de las LC ya que estos autores muestran que las LC expresan Zbtb46, mientras que se originan a partir de un progenitor que expresa Mafb, lo que indica el origen de los macrófagos ( 6 ). Se demostró que MafB controla una red de genes de autorrenovación en proliferantes macrófagos residentes ( 13 ). Al igual que los macrófagos residentes de tejidos, las LC proliferan en un estado diferenciado, y estos hallazgos implican que MafB probablemente esté involucrado en la proliferación de LC. Sin embargo, las señales moleculares específicas que conducen la proliferación de LC in situ aún no se han dilucidado. Estos resultados resumen muy bien la persuasión actual de las LC que son macrófagos con funciones DC. Fenotípicamente, los LC comparten muchas características con CDCs, mientras que su ontología los relaciona con macrófagos residentes de tejido. En conjunto, esto muestra que las LC representan un tipo de célula altamente único, compartiendo características con los países en desarrollo, pero que surgen de un origen diferente y ejercen funciones únicas que los distinguen de los CDC, tal como se resume en la Figura 1 . Figura 1 Representación esquemática de las propiedades de la célula de Langerhans (LC) compartida por células dendríticas (DC) o macrófagos. Las LC muestran una mezcla de propiedades que comparten con los macrófagos (indicados en verde), como el auto mantenimiento y la ontogenia. Sin embargo, las LC son capaces de presentar antígeno y migrar activamente a los ganglios linfáticos que lo drenan, lo que los califica como CD (las propiedades compartidas entre LC y DC están indicadas en rojo). FENOTIPO Y ONTOGENIA Las células de Langerhans constituyen el 3-5% de todas las células nucleadas en la epidermis y ocupan el estrato espinoso , una capa epidérmica suprabasal que se caracteriza por queratinocitos "espinosos" que se someten a un refuerzo de su citoesqueleto. Las LC son células estrelladas que sobresalen de sus dendritas a través de uniones estrechas hacia el estrato córneo y, como tales, pueden detectar antígenos a través de varias capas de la epidermis sin alterar la barrera de permeabilidad. Las LC adquieren un fenotipo de DC típico después del nacimiento expresando moléculas del complejo mayor de histocompatibilidad II (MHCII) y la cadena de integrina αX (CD11c). A diferencia de las LC de ratón, las LC humanas expresan niveles bajos de CD11c y no de F4 / 80, mientras que expresan CD1a y CD1c, dos moléculas relacionadas con MHCI implicadas en la presentación de antígenos lipídicos ( 7 ). La molécula de adhesión de células epiteliales (EpCAM) regula la adhesión de LC a los queratinocitos y promueve la migración de LC a los ganglios linfáticos de drenaje ( 14 ). Las LC expresan la lectina tipo C Langerin (CD207), que está involucrada en la formación de gránulos de Birbeck, orgánulos endosómicos que son típicos para las LC.Se ha demostrado que estos gránulos de Birbeck internalizan virus ( 15 ) y se cree que son parte del complejo de reciclaje endosomal ( 16 ). La presencia de Langerin sola no es confiable para definir inequívocamente las LC, ya que la expresión de Langerin también se ha demostrado en cDC en la dermis, los pulmones y el bazo. Del mismo modo, las LC humanas y de ratón expresan SIRPα (CD172α), CD11b y CX3CR1, que también se expresa en la mayoría de las CDC de tipo 2. Dado que los marcadores mencionados anteriormente son compartidos por diferentes subconjuntos de células mieloides, se requiere una citometría de flujo multicolor con un conjunto mínimo de marcadores impresos de linaje para la identificación inequívoca de las LC en la piel y los ganglios linfáticos que drenan la piel ( 17 ). Langerhans células claramente comparten similitudes con DC y macrófagos (Figura 1 ]. Doebel et al.recientemente resumió la evidencia emergente de la identidad dual de las LC, ya que surgen de precursores de macrófagos y adquieren las propiedades únicas de las DC en la epidermis ( 18 ). De hecho, las LC son células resistentes a la radio de origen embrionario, que se mantienen mediante la autorrenovación. Con una vida media estimada de 2 meses ( 19 ), las LC exhiben una tasa de proliferación lenta en condiciones homeostáticas para reemplazar las células que mueren y emigran. En ese aspecto, las LC se parecen más a los macrófagos residentes en tejido que las CD clásicas, que se derivan de precursores de médula ósea y dependen de progenitores circulantes para retener su alta velocidad de rotación ( 18 , 20 ). A diferencia de los precursores de cDC que requieren señalización de tirosina quinasa-3 (Flt3) tipo FMS para diferenciarse, las LC se desarrollan independientemente del ligando Flt3 y Fltl3 (Flt3L). Es de destacar que las LC de migración regulan positivamente la expresión de Flt3 , lo que sugiere una función de Flt3L en la activación de las CL ( 21 ). Al igual que los macrófagos residentes de tejidos, las LC proliferan en un estado diferenciado y, por lo tanto, expresan un conjunto de genes de autorrenovación. Sin embargo, las señales moleculares específicas que conducen la proliferación de LC in situ aún no se han dilucidado. A pesar de que las LC comparten similitudes ontogénicas con los macrófagos que viven en los tejidos, a diferencia de los macrófagos, son capaces de migrar a los nódulos linfáticos de drenaje. Las LC pueden absorber y procesar antígenos extraños en condiciones de estado estacionario e inflamación, que pueden presentar a las células del sistema inmune adaptativo en los ganglios linfáticos. La migración de LC es un proceso de varios pasos que implica una plétora de cambios. En primer lugar, las LC necesitan debilitar sus conexiones intercelulares con los queratinocitos circundantes, que está parcialmente regulado por Ecadherina ( 2 ). La liberación de E-cadherina provoca la translocación de β-catenina, que está involucrada en el fenotipo tolerogénico de las CD ( 22 ). La señalización Wnt canónica, que está mediada por β-catenina, es un regulador principal del comportamiento de los queratinocitos en la homeostasis y la enfermedad de la piel ( 23 ). Por lo tanto, sería interesante investigar el comportamiento de LC en modelos de ratón de activación de βcatenina cutánea aberrante con el fin de evaluar la participación de las CL en la alteración del comportamiento de los queratinocitos. En un siguiente paso, las LC necesitan cruzar la membrana basal para migrar a la dermis y transitar al nódulo linfático. Por lo tanto, localmente secretan metaloproteinasas de matriz que degradan el colágeno, lo que permite reordenamientos dérmicos y la motilidad transdérmica de LC ( 24 ). Una vez en la dermis, las LC pueden ingresar a los vasos linfáticos para viajar a los ganglios linfáticos regionales. La resembración de LC desde la médula ósea hacia la epidermis ocurre a través de rutas estrictamente mediadas a lo largo del folículo piloso. Los queratinocitos presentes en la región por encima del bulbo capilar secretan un ligando de quimioquina (motivo C-C) (CCL) -2 y CCL-20 ( 25 ). Un factor de crecimiento crucial para la regulación adecuada de la biología de LC ha demostrado ser el factor de crecimiento transformante (TGF) -β y sus factores de transcripción asociados PU.1, Id2 y RUNX3. Los ratones deficientes para una de estas proteínas carecen de LC completamente desarrolladas ( 26 - 28 ). TGF-β también impulsa la diferenciación de células precursoras humanas a células con características de LC in vitro ( 29 , 30 ). La señalización de TGF-β puede ser inducida tanto por queratinocitos como por LC. Sin embargo, el hallazgo de que los ratones con una deleción condicional de TGF-β en LC exhiben números de LC reducidos es indicativo de un ciclo autocrino crucial para TGF-β en la biología de LC ( 31 ). PU.1 juega un papel fundamental en muchos aspectos de la hematopoyesis temprana y la diferenciación de células mieloides y regula la expresión del gen LC esencial Runx3 en respuesta a TGF-β ( 32 ). Curiosamente, LC en estado estable y condiciones inflamatorias exhiben requisitos diferenciales para el factor de transcripción inducido por TGF-β ID2, lo que apoya el hallazgo de que las LC inflamatorias surgen de diferentes precursores que sus contrapartes de estado estable ( 32 , 33 ).La molécula del adaptador endosomal tardío p14 (LAMTOR2) también es indispensable para la homeostasis de LC y esto parece deberse a cambios en la sensibilidad del TGF-β ( 34 , 35 ). Curiosamente, las LC deficientes en p14 muestran una sensibilidad incrementada a estímulos apoptóticos y proliferación defectuosa, en parte debido a la regulación a la baja del receptor II de TGF-β. Otro miembro de la familia de TGF-β que se demostró importante para los primeros pasos de la diferenciación de LC es la proteína morfogenética ósea (BMP) 7. Los ratones que carecen de BMP7 tienen números de LC significativamente reducidos, y los LC parecen menos dendríticos que en los compañeros de camada de control ( 36 ). Estos autores demostraron que el desarrollo de LC humana podría ser impulsado por BMP7 en el caso de señalización redundante de TGF-β, como es el caso en la epidermis prenatal y en la capa epidérmica basal ( 36 ). En conclusión, el número de LC en la epidermis en condiciones homeostáticas se mantiene mediante la autorrenovación que repone la constante migración de bajo nivel desde la epidermis a los ganglios linfáticos que drenan. El desarrollo, la diferenciación y la proliferación de LC están estrechamente controlados por una red de factores de transcripción, citoquinas y factores de crecimiento, que están muy influenciados por el microambiente tisular. LC EN INFLAMACIÓN CUTÁNEA Y CICATRIZACIÓN DE HERIDAS Las células de Langerhans son, debido a su ubicación dentro de los epitelios estratificados, parte de la primera línea de defensa frente a los patógenos presentes en el medio ambiente. Las LC no activadas migran constantemente a los ganglios linfáticos para presentar autoantígeno y establecer tolerancia inmune en condiciones homeostáticas ( 37 , 38 ). Durante el encuentro patogénico, los receptores de reconocimiento de patógenos codificados en la línea germinal reconocen patrones moleculares en los microbios, lo que resulta en la secreción de citoquinas proinflamatorias y la posterior movilización de las LC a los ganglios linfáticos ( 39 ). La importancia funcional de las LC en la infección y su interacción con PRR es ampliamente revisada por West et al. en este tema de investigación ( 5 ). La inflamación cutánea, la infección o la lesión provocan una migración extensa de LC desde la epidermis, generando un nicho vacío que se repobla por una onda inicial de LC de vida corta que surge de los monocitos Gr-1 hi circulantes ( 33 , 40 ). Seré et al. informaron que estas LC derivadas de monocitos que se infiltran en la piel durante la inflamación son efímeras e independientes de Id2, pero su reclutamiento en la piel depende de CCR-2 y -6 ( 33 ). Estas LC de vida corta son reemplazadas gradualmente por una segunda ola de LC expresando Id2 que permanecen a largo plazo en la epidermis y expresan altos niveles de Langerin, CD24 y EpCAM, lo que los distingue de las LC de vida corta ( 33 ). Estas observaciones indican la importancia de mecanismos distintos para el desarrollo de LC en condiciones homeostáticas frente a inflamatorias. Los LC están impresos por la epidermis, ya que requieren interleucina (IL) -34 derivada de queratinocitos para su desarrollo, homeostasis y regeneración ( 41 ). Las respuestas de curación de heridas en la piel implican reordenamientos celulares múltiples para asegurar la restauración rápida de la función de barrera de permeabilidad epidérmica. Las respuestas cutáneas regenerativas se pueden subdividir en múltiples fases, a saber, la etapa inflamatoria, la formación de nuevo tejido y la fase de remodelación tisular que se caracterizan cada una por respuestas moleculares y celulares específicas ( 42 ). Los queratinocitos que se estresan durante las heridas regulan rápidamente al alza los ligandos del grupo natural killer 2D (NKG2D), lo que provoca la migración de poblaciones de LC fuera de la epidermis, seguido de la aparición de células T αβ en la epidermis ( 43 ). Se han observado niveles de expresión mejorados del ligando murino Rae-1ε de NKG2D en la piel herida, y los ratones que carecen de NKG2D presentan cicatrización cutánea tardía ( 44 ), corroborando la importancia de la diafonía entre queratinocitos y LC durante procesos regenerativos en la piel. Esta interacción intercelular ya es aparente en respuesta a una lesión menor, como el desdoblamiento repetitivo de la epidermis ( 45 ). En estas condiciones inflamatorias, se ha demostrado que las CL penetran en las uniones estrechas que unen los queratinocitos, permitiendo la endocitosis de los antígenos ( 46 ). El diálogo cruzado LC-queratinocitos se establece en parte mediante la producción de citocinas proinflamatorias, como IL-1, factor estimulante de colonias de granulocitos macrófagos (GM-CSF) y factor de necrosis tumoral (TNF), que estimulan la migración de LC ( 47 ). Hasta ahora, no se sabe si esas citocinas pueden alterar directamente el comportamiento de LC in vivo o si esto ocurre indirectamente a través de la activación de queratinocitos.La repoblación de la epidermis por LC ocurre en las etapas finales de la curación de la herida, es decir, la etapa de remodelación del tejido ( 48 ). Todavía no está claro si la repoblación de LC después de la herida ocurre a través de una infiltración sustancial de LC derivadas de monocitos oa través de una onda inicial de una pequeña población de células LC infiltrantes que proliferan in situ . Los datos in vitro que usan células endoteliales linfáticas dérmicas primarias humanas muestran que la estimulación con TNF da como resultado la sobreexpresión de moléculas clave de adhesión superficial ( 49 ). En la epidermis humana no ulcerada de pacientes diabéticos, que han retrasado notablemente las respuestas reparadoras cutáneas, se observó un aumento de las LC en relación con los pacientes normoglucémicos. Un mayor número de LC presentes en las úlceras del pie diabético se asociaron con un mejor resultado de curación, lo que indica un papel beneficioso para las LC humanas en la inflamación reparadora cutánea ( 50 ). En contraste, la observación de que la reparación de lesiones de la piel ocurre normalmente en ratones que carecen del factor de transcripción PU.1, que es crucial para el desarrollo de LC ( 32 ), argumentaría un papel redundante de las LC en las respuestas regenerativas cutáneas ( 51 ). Claramente, el papel de las LC en la mediación de las diferentes etapas de la regeneración de heridas todavía está bajo debate, y queda por investigar si dirigir esta población de células sería una vía terapéutica interesante para tratar la inflamación aguda o crónica inducida por la herida. LC EN REACCIONES ALÉRGICAS DE LA PIEL Las células de Langerhans se han estudiado ampliamente en reacciones alérgicas de la piel. Sin embargo, el papel de las LC en la hipersensibilidad de contacto y la sensibilización alérgica sigue siendo controvertido. Contacto Hipersensibilidad La dermatitis de contacto alérgica ocurre en individuos que desarrollan una hipersensibilidad de tipo retardado tipo IV contra alérgenos de contacto, en su mayoría pequeñas moléculas orgánicas con reactividad química (los denominados sensibilizadores químicos como metales, antibióticos tópicos o conservantes). Estos sensibilizadores químicos penetran en las capas de piel cornificadas debido a su pequeño tamaño y se unen covalentemente a las proteínas epidérmicas para formar los llamados "neoantígenos" que pueden ser reconocidos por APCs ( 52 ). Este proceso de haptización puede evitarse experimentalmente mediante la aplicación de haptenos a la piel murina. Las respuestas tisulares posteriores imitan las que surgen durante la dermatitis de contacto alérgica en humanos. Las LC son una de las primeras APCs en encontrar estos haptenos, pero su papel exacto en CHS sigue sin estar claro debido a los resultados contradictorios obtenidos de diferentes modelos de ratón, que se revisan extensamente en otra parte ( 53 ). Los informes iniciales demostraron que el agotamiento de las LC no podía derogar por completo las respuestas de CHS a los haptenos ( 54 , 55 ). Esto enfatizó la redundancia de los diferentes subconjuntos de DC en la epidermis y la dermis, ya que los dípteros también pueden activar CD dérmicas, ya que son lo suficientemente pequeños como para penetrar en la dermis. Los estudios que intentaron desentrañar el papel de las LC en CHS utilizaron diferentes modelos de ratón para eliminar las LC funcionales, lo que dio lugar a hallazgos contradictorios. La ablación genética de las LC se puede lograr inyectando toxina diftérica en ratones que expresan el receptor de toxina diftérica (DTR) humano o simio impulsado por el promotor CD207 (Langerin). Esta toxina bloquea la traducción de proteínas e induce la muerte celular solo en células que expresan transgénicamente DTR, ya que el ortólogo de ratón de este receptor es significativamente menos sensible a la toxina ( 56 ). En la piel de los ratones Langerin-DTR, todas las LC y Langerin + cDC1 pueden agotarse con una única inyección de DT. Al usar este sistema transgénico, dos informes mostraron que las LC eran en gran medida prescindibles para la inducción de CHS ( 54 , 55 ). Sin embargo, la noción de que Langerin + cDC1s dérmicos también se agotan en estos ratones requiere una reconsideración de los hallazgos previos. Para investigar específicamente la contribución de las LC en CHS, las respuestas de CHS pueden evaluarse 1 semana después de la inyección de DT, ya que Langerin +cDC1s se reabastecen de precursores circulantes en 7 días, mientras que las LC permanecen ausentes de la epidermis durante al menos 2 semanas. El cebado de células T para haptenos aplicados tópicamente disminuyó una vez que se reconstituyeron Langerin + cDC1s, mientras que las LC todavía estaban agotadas, lo que indica un papel para las CL en la sensibilización a CHS ( 57 ). Sin embargo, existe una redundancia funcional entre LC y Langerin + cDC1s, dependiendo de la dosis de hapteno ( 58 ). Un método alternativo para evaluar la importancia de las LC en las respuestas de CHS es hacer uso de ratones humanos (h) Langerin-DTR, que constitutivamente carecen de LC en presencia de Langerin +cDC1 funcionales debido a una regulación de promotor diferente del gen CD207 humano.Sorprendentemente, estos ratones mostraron una mayor sensibilización cutánea a los haptenos, lo que sugiere un papel supresor de las LC en CHS ( 59 ). Sin embargo, estas conclusiones fueron refutadas por otros informes que demuestran que las respuestas de CHS se redujeron en ratones que solo carecen de LC debido a la deleción condicional del receptor de TGFβ 1 o p14 en las células de Langerin + ( 35 , 60 ). De hecho, las LC se han implicado en la inducción de células T reguladoras y son cruciales para establecer la tolerancia a la sensibilización alérgica de contacto leve al expandir las células T reguladoras mientras se eliminan las respuestas de células T CD8 + específicas de alérgeno ( 61 - 63 ). Por consiguiente, la sobreexpresión transgénica de RANKL por los queratinocitos reduce las respuestas de CHS, y esto se mostró mediado por una capacidad potenciada de las LC para expandir las células T reguladoras CD4 +CD25 + ( 64 ). Estas funciones supresoras de las LC de estado estacionario se informaron anteriormente en la piel humana, donde se demostró que las LC activan las células T reguladoras residentes en la piel ( 10 ).Curiosamente, estos estudios demostraron que el estado de activación de las LC determinó su respuesta inmunológica, ya que las LC activadas indujeron células T efectoras y la activación limitada de las células T reguladoras tanto en ratones ( 63 ) como en humanos ( 10 ). En conclusión, al estudiar las respuestas de CHS a los haptenos, se hizo evidente que las LC pueden contribuir a la sensibilización por alérgenos de contacto en función de la dosis de hapteno y en el modelo de ratón experimental que se utiliza. Haciendo uso de un modelo de ratón transgénico que expresa CD1a, una proteína presentadora de lípidos que normalmente no está presente en ratones, se demostró que las LC que expresaban CD1a transgénicamente mostraban un CHS suprimido a dinitrofluorobenceno. Sin embargo, estos CD1a + LCs pudieron inducir respuestas Th17 a la aplicación tópica de lípidos de plantas inflamatorias ( 65 ). Esto indica que la naturaleza del hapteno puede provocar la presentación de antígenos por LC que sesga las respuestas de las células T en una dirección específica. Sin embargo, no está claro por qué en algunos pacientes se rompe la tolerancia y se produce dermatitis de contacto, y cómo las LC están involucradas en este proceso. En conclusión, es crucial tener en cuenta la dosis, la naturaleza y la ruta de aplicación del hapteno al sacar conclusiones de los modelos de ratón de CHS. Además, la pérdida de LC debido a un estímulo o lesión inflamatoria severa se reabastecerá circulando monocitos Gr-1 hi , mientras que la pérdida de LC debido a un estímulo menos severo puede resolverse mediante la proliferación local de CL ( 20 ).Esto implica nuevamente que la composición del conjunto de LC y su contribución a CHS puede variar entre modelos experimentales. Sensibilización alérgica a antígenos proteicos Al igual que los alérgenos de contacto, los alergenos proteicos de, por ejemplo, moho, polen, ácaro del polvo doméstico (HDM) y cucarachas pueden estar presentes en grandes cantidades en la superficie de la piel. Sin embargo, cuando se estudia la sensibilización alérgica, es importante señalar que los haptenos penetran fácilmente en las capas más profundas de la piel y activan las DC dérmicas, mientras que hasta ahora no está claro si los alérgenos proteicos pueden hacerlo sin una interrupción previa de la barrera. Por lo tanto, muchos modelos experimentales infligen una disrupción de la barrera epidérmica antes de la exposición al antígeno proteínico, por ejemplo, mediante extracción con cinta adhesiva o mediante el uso de irritantes como la acetona. Estas manipulaciones también activarán los queratinocitos y las células inmunes en la piel e inducirán una inflamación leve ( 66 ). En consecuencia, la perturbación de la barrera epidérmica, antes de la exposición al alergeno, influye en el resultado de diversos modelos experimentales utilizados para estudiar las respuestas alérgicas y debe tenerse en cuenta. Nakajima et al. informaron que las LC son cruciales para la sensibilización epicutánea a la ovoalbúmina (OVA) ( 67 ), mientras que recientemente demostramos que los CDC tipo 2 dérmicos y no las CL son responsables de la sensibilización de la piel a la HDM ( 68 ). Descubrimos que los ratones con depleción de LC pueden sensibilizarse eficientemente a HDM a través de la piel y que los LC cargados con HDM no pueden transferir la sensibilización a los ratones que no han recibido tratamiento previo ( 68 ). La mayor discrepancia entre estos modelos radica en el hecho de que OVA es una proteína inerte que requiere cinta previa, lo que permite la penetración y funciona como un adyuvante activando los queratinocitos y las células inmunes, mientras que el extracto HDM contiene enzimas proteolíticas activas y endotoxinas contaminantes que puede activar los queratinocitos y las células inmunes para producir citocinas proalérgicas. El hecho de que estos modelos induzcan la activación de los queratinocitos de una manera diferente podría explicar los roles divergentes de las CL durante la sensibilización. La evidencia emergente revela una importante interacción entre las células epiteliales y las CD durante la sensibilización alérgica, tanto en los pulmones como en la piel ( 69 ). Anteriormente mostramos que la señalización del receptor tipo peaje inducida por endotoxinas (TLR4) en las células epiteliales bronquiales se requiere para el cebado de Th2 a HDM ( 70 ) mediante la producción de citoquinas Th1skewing ( 71 ). Del mismo modo, los queratinocitos activados pueden producir citoquinas pro-alérgicas típicas que sesgan las CD para cebar las células Th2, como la linfopoyetina estromal tímica (TSLP), IL-33, IL-1α y GM-CSF ( 4 ). Curiosamente, TSLP estimula las LC humanas para inducir células T pro-alérgicas ( 72 ). Es de destacar que el efecto de maduración proalérgico de la TSLP se limitó a las CL epidérmicas y no se produce en las LC derivadas de CD34 + ( 73 ). Sin embargo, las LC son prescindibles para el cebado Th2 dirigido por TSLP en un modelo de ratón donde se evoca la sobreproducción de TSLP mediante la aplicación de un análogo de vitamina D3 ( 74 ). Sorprendentemente, encontramos que ni la perturbación de barrera previa a la sensibilización ni la actividad proteolítica presente en HDM se requieren para la sensibilización epicutánea eficiente a HDM ( 68 ). Aunque no encontramos un papel crucial para las LC durante la sensibilización de la piel a la HDM, demostramos que la falta de LC da como resultado un aumento en la producción de citocinas tipo 2 por las células T reestimuladas con HDM en los ganglios linfáticos ( 68 ). Nuestros hallazgos están en línea con estudios previos que muestran que las LC pueden ejercer funciones reguladoras ( 9 , 10 , 62 ) y, por lo tanto, también podrían ser capaces de suprimir Th2 cebado a HDM. De nuevo, como en CHS, estos hallazgos contradictorios indican que múltiples factores determinan si las LC contribuyen a la sensibilización epicutánea y cómo lo hacen. Por ejemplo, si los alergenos entran en contacto con la barrera de la piel alterada de individuos genéticamente predispuestos, pueden eludir la función homeostática supresora potencial de las LC y obtener acceso directo a las CD dérmicas. Además, en pacientes con un grado mínimo de inflamación en la piel, las LC pueden estar sesgadas hacia un estado proalérgico. En conclusión, el papel exacto de las LC en la sensibilización epicutánea aún no se ha dilucidado, y los hallazgos recientes apuntan a una alta redundancia funcional entre diferentes subconjuntos de DC de la piel. LC EN DERMATITIS ATÓPICA (AD) El eccema o enfermedad de Alzheimer es una enfermedad causada por Th2 que a menudo precede a las respuestas alérgicas o asmáticas en un proceso progresivo denominado marcha atópica. Aunque el papel de las LC en la sensibilización a los alérgenos proteínicos todavía está bajo debate, las LC también pueden contribuir a la fase efectora de las reacciones alérgicas en la piel, causando síntomas de AD (Figura 2 ). Sin embargo, los informes de pacientes con AD son limitados y el papel exacto de las LC en este contexto aún no se ha dilucidado. En biopsias de piel de pacientes con EA, hay más LC presentes, aunque no está claro si estos son LC residentes en la piel o derivados de monocitos ( 75 ). Estos autores muestran que TSLP causa proliferación de LC residentes en la piel, mientras que no afecta la proliferación de una onda marginal de LC derivadas de monocitos ( 75 ). Las LC humanas están en un estado activado en lesiones AD como lo demuestra la expresión mejorada de CD80 y CD86 ( 76 ), que fue corroborada por la observación de que las LC murinas también expresan marcadores de maduración incrementados en un modelo murino de AD y esto se correlaciona con severidad AD ( 77 ). La piel humana con función de barrera epidérmica defectuosa, como es el caso en pacientes con mutaciones en el gen de filagrina que codifica una proteína estructural principal en queratinocitos, predispone al desarrollo de EA ( 78 ). Curiosamente, recientemente se demostró que en biopsias de piel de pacientes que albergan mutaciones nula filagrina, LC están en un estado más activado en comparación con los pacientes control, independientemente de si estos pacientes sufrieron de AD ( 79 ). Estos autores demuestran que los productos de degradación de filagrina, que actúan como factores naturales de hidratación en la piel, reducen el fenotipo de maduración de las CD derivadas de monocitos generadas ex vivo . En conjunto, esto indica que las mutaciones de filagrina pueden afectar la función de LC (Figura 2 ], pero si esto resulta en LC hiperactivación que instiga los síntomas de AD aún no se ha explorado. Otra indicación de que las LC pueden ejercer un papel importante en la fase efectora de la EA es la observación de que las LC de las lesiones de AD muestran niveles de expresión aumentados del receptor de inmunoglobulina (Ig) E de alta afinidad (FcεRI) ( 80 , 81 ). Este receptor se une a moléculas de IgE específicas de alérgeno, promoviendo así el antígeno que se enfoca en la superficie celular y la posterior absorción de antígeno ( 82 ). De manera interesante, en los LC derivados de CD34 + humanos, la ligación de TLR2 por los productos de Staphylococcus aureus dio como resultado la regulación a la baja de la expresión de FcεRI ( 83 ). Recientemente, se postuló que el efecto de resolución de la AD de la fototerapia podría estar mediado por LC. La irradiación ultravioleta (UV) B genera fotoproductos que activan el receptor de hidrocarburo de arilo ( 84 ), que a su vez regula negativamente FcεRI en LC derivadas de CD34 + generadas ex vivo ( 85 ). Además, un informe reciente mostró que la activación de AhR por el benzopireno presente en el humo del cigarrillo indujo la movilización de LC en asociación con niveles reducidos de E-cadherina en los queratinocitos ( 86 ). Además, las LC deficientes en AhR mostraron una maduración alterada, que dio como resultado una reducción de CHS a los haptenos ( 87 ). Ambos eventos indican que el benzopireno podría activar directa o indirectamente las vías proinflamatorias en las LC que pueden empeorar la EA, pero esto aún no se ha investigado. Los niveles de expresión de AhR también se incrementan en la piel con AD ( 86 ), y recientemente se propuso que este receptor vincule la contaminación del aire con la EA ( 88 ). En conjunto, estos informes implican que la naturaleza del ligando AhR podría definir un estímulo pro o antiinflamatorio para las LC que pueden contribuir al desenlace de la enfermedad. Las células de Langerhans son importantes para aumentar las respuestas Th17 cutáneas a ciertas infecciones bacterianas. Curiosamente, se encontró que las células T específicas de HDM de pacientes con AD presentaban un fenotipo Th2, Th17 o Th2 / Th17 ( 89 ). Además, el porcentaje de células Th17 y el nivel de citocinas circulantes de IL-17 / IL-23 en sangre periférica de pacientes con EA se correlacionan con la gravedad de la enfermedad ( 90 , 91 ). Todavía no está claro cómo surge exactamente esta respuesta Th17 adicional en la EA, una enfermedad mediada por Th2. Una posible explicación es que una barrera defectuosa, inducida por defectos genéticos o arañazos, podría facilitar la disbiosis microbiana de la piel. De hecho, los pacientes con AD regularmente sufren de colonización concomitante de S. aureus , que agrava la enfermedad ( 92 ). Kobayashi et al. demostraron que los ratones que desarrollan piel eccematosa con una barrera epidérmica alterada son más propensos a desarrollar colonización por S. aureus y respuestas inmunitarias cutáneas Th17 concomitantes. Este fenotipo inflamatorio fue mediado por la infiltración de células T γδ y células Th17 y fue abrogado cuando los ratones carecían de LC ( 93 ). Del mismo modo, se demostró que las LC humanas tienen una alta capacidad para inducir la producción de IL-22 por las células T γδ, que a su vez tienen potentes efectos de limpieza sobre S. aureus ( 94 ). En conclusión, los LC pueden ejercer diferentes funciones durante la EA dependiendo de los estímulos que reciben del microambiente. Esto explica por qué, a pesar de muchos esfuerzos, los investigadores hasta ahora no han podido atribuir un papel inequívoco para los LC en AD. Figura 2 Representación esquemática de las funciones de la célula de Langerhans (LC) en la dermatitis atópica (AD).En la piel de pacientes con EA, una barrera defectuosa puede deberse a mutaciones en el gen que codifica la proteína estructural filagrina. Durante la diferenciación de los queratinocitos, el monómero de filagrina se degrada al nivel de los factores naturales de hidratación, lo que puede alterar el comportamiento de las células dendríticas. La barrera epidérmica defectuosa conduce a la inducción de LC, que puede mejorar sus tasas de proliferación y exhibir un estado de activación mejorado. Al detectar insultos ambientales, se monta una respuesta de tipo 17 adicional, y las LC migran al nódulo linfático para presentar el antígeno. LC EN PSORIASIS La psoriasis es un trastorno cutáneo inflamatorio crónico que se manifiesta por la presencia de escamas plateadas en las regiones afectadas y que se caracteriza por una mayor proliferación y una diferenciación anormal de los queratinocitos. En la piel psoriásica, hay una acumulación de células inmunitarias, de las cuales las CD y las células T son las más esenciales ( 95 ). El conocimiento actual sobre el papel de las LC en pacientes psoriásicos humanos se revisa extensamente en otro artículo de revisión en este tema de investigación ( 5 ). En ratones, una respuesta inmune similar a la psoriasis en la piel puede imitarse mediante la aplicación tópica del imiquimod ligando TLR7 y -8 (IMQ), y la dermatitis inducida en este modelo experimental depende de la señalización IL-23 / Th17 ( 96 ). Los ratones con depleción de LC muestran una disminución de la dermatitis inducida por IMQ y niveles significativamente reducidos de IL-23, lo que indica que las CL son una fuente importante de IL-23 en este modelo ( 97 ). Por el contrario, Wohn et al. informaron que las cDC, y no las LC, son responsables de la producción de IL-23 y la dermatitis posterior ( 98 ). Estos hallazgos contradictorios, ambos deducidos al usar la misma cepa de ratón Langerin-DTR, podrían deberse a la diferencia en la aplicación tópica de IMQ, es decir, en la oreja frente a la piel de la espalda. Esto podría indicar que, en función de su ubicación dentro del cuerpo, las LC pueden inducir diferentes respuestas tisulares. Los ratones que expresan ectópicamente la proteína presentadora de lípidos humanos CD1a en las LC muestran una respuesta psoriática agravada en el tratamiento IMQ, que podría reducirse mediante la administración de un anticuerpo bloqueante de CD1a. También en pacientes humanos, la respuesta inflamatoria de las citoquinas típica de la psoriasis se bloqueó mediante el tratamiento con anticuerpos anti-CD1a ( 65 ). Estas observaciones ejemplifican la importancia de la presentación del antígeno lipídico en la patogénesis de la psoriasis. Entre los mediadores inflamatorios de la psoriasis, se ha demostrado que el TNF y la IL-1β son componentes esenciales de la tormenta de citocinas que es típica en las erupciones activas de la enfermedad ( 99 ). Recientemente, se demostró que las principales CD responsables de la producción de estas citoquinas proinflamatorias son CD derivadas de monocitos, incluida una población de LC ( 100 ). Estas observaciones recientes vuelven a enfatizar el papel de las LC en el montaje del brazo Th17 de la respuesta inmune que ocurre en la piel psoriática. Como la neutralización de IL-17α es altamente efectiva en el tratamiento de la psoriasis ( 101 ), el direccionamiento terapéutico específico de las LC en el contexto de la psoriasis podría representar una vía terapéutica interesante. PATOLOGÍAS ASOCIADAS A LC La histiocitosis de células de Langerhans es un trastorno de etiología desconocida con diferentes manifestaciones clínicas y gravedad de la enfermedad, que se caracteriza por la infiltración de DC activadas en lesiones multifocales ( 102 , 103 ). Una característica clínica común de la histiocitosis LC es una erupción de la piel poco después del nacimiento, que puede resolverse espontáneamente o propagarse a otras regiones del cuerpo. Aunque se han observado gránulos de Birbeck en las células dentro de estas lesiones, el origen LC de estas histiocitosis no se ha demostrado formalmente. Las histiocitosis LC abarcan muchos subtipos con una amplia gama de manifestaciones clínicas, que se caracterizan por inflamación e hiperproliferación de leucocitos y pueden afectar a la piel, los huesos, los pulmones, la médula ósea, el sistema nervioso central y los ganglios linfáticos ( 104 ). Informes recientes han demostrado que en dos tercios de los pacientes las mutaciones activadoras en la vía RAF / MEK / ERK se producen dentro de las lesiones histiocíticas LC ( 105 , 106 ). Los tratamientos terapéuticos actuales abarcan inhibidores de MEK o BRAF o una combinación de ambos, tal como ha sido revisado recientemente por Haroche et al. ( 104 ) El modelo actual de ratón desarrollado para estudiar la histiocitosis LC, mediante la expresión transgénica de los oncogenes derivados del virus simio 40 bajo el control del promotor CD11c, no solo imita ciertas características de la enfermedad sino que también muestra características del sarcoma LC ( 107 ). Por lo tanto, este modelo animal podría no representar la mejor herramienta para estudiar la etiología de la histiocitosis LC. El sarcoma de células de Langerhans es un raro trastorno de LC neoplásico que puede formarse de nuevo o puede originarse por histiocitosis LC ( 108 ). Hasta el momento, no se han identificado modificaciones cromosómicas recurrentes en ambos trastornos, pero son necesarios más estudios para clasificar correctamente y potencialmente estratificar estas enfermedades. LC EN CÁNCER Los tumores de piel no melanoma incluyen carcinomas de células basales y carcinomas de células escamosas (SCC), que típicamente exhiben una presencia de LC muy reducida en las regiones peritumoral y tumoral ( 109 ). Como las DC son capaces de provocar respuestas inmunes antitumorales potentes, generalmente se exceptúa que desempeñan un papel crucial en la vigilancia inmune del cáncer. Sin embargo, las CD pueden presentar alteraciones funcionales en ciertos tipos de tumores y volverse menos potentes en la estimulación de células T y la producción de interferones, lo que permite escapar de la vigilancia inmune ( 110 , 111 ). Varios informes apuntan a un papel crucial para las LC en la mediación de la progresión maligna en la piel. En los SCC humanos, se observó la supresión de la funcionalidad de DC, mientras que las LC aisladas de los SCC se han mostrado como potentes estimuladores inmunes de tipo 1 in vitro ( 112 ). Curiosamente, las LC migran anormalmente desde la piel de SCC y tienen capacidades reducidas de cebado de células T dentro del ganglio linfático ( 113 ). Muchas citoquinas que se sabe que median en la migración de LC de la dermis, como TSLP, IL-1 y TNF, se han implicado en modelos de cáncer cutáneo ( 114 - 116 ). Las aberraciones en la barrera de permeabilidad epidérmica preparan la secreción de estas citocinas proinflamatorias, lo que da como resultado una susceptibilidad al cáncer alterada ( 117 ). Sin embargo, queda por establecer si en estas condiciones, la migración aumentada de LC es una causa subyacente de tumorigénesis de la piel. Dado el reciente descubrimiento de DC que expresan Langerin más allá de las LC, es plausible que muchos de los efectos de la biología tumoral atribuidos a las LC se deben en realidad a otras células Langerin + en la piel, y por lo tanto, estas observaciones deben revisarse. Se ha demostrado que las LC humanas son activadores más eficientes de las células T CD8 + vírgenes que las CD dérmicas, lo que podría tener implicaciones importantes para las respuestas antitumorales ( 118 ). En el modelo ampliamente establecido de dos etapas DMBA / TPA de carcinogénesis cutánea química, los ratones deficientes en Langerina están completamente protegidos de la papilomagenesis y la posterior formación de SCC ( 119 ). Estos autores demuestran que las células Langerin + pueden mejorar el daño en el ADN inducido por DMBA, aunque a la luz de los hallazgos recientes, queda por establecer si las LC son las principales CD de Langerin + que son capaces de metabolizar DMBA. De hecho, varios informes ahora muestran que la actividad de metabolización de DMBA de LC es prescindible para infligir mutaciones de H-Ras en queratinocitos ( 120 , 121 ). El tratamiento con radiación ionizante se usa con frecuencia como terapia tumoricida localizada en pacientes con cáncer; sin embargo, los datos recientes apuntan a la inducción de respuestas inmunitarias antitumorales adicionales mediante radioterapia, como la infiltración de células T reguladoras dentro del tumor y la activación de APC ( 122 , 123 ). Las LC son notablemente resistentes a la radiación y son muy potentes para reparar rápidamente el daño infligido al ADN, que se demostró que estaba mediado por CDKN1A (inhibidor de la cinasa dependiente de ciclina) ( 124 ). El daño repetitivo por radiación UVB predispone a la formación de melanoma y dado el asombroso éxito de las intervenciones inmunoterapéuticas en pacientes con melanoma, es fundamental para comprender la dinámica de las células inmunes dentro del microambiente del melanoma. En base a la coexpresión de S100 ( 125 ) y CD1, se ha informado una reducción de las LC en la epidermis que recubre al melanoma ( 126 ). Este número reducido de LC puede representar un evento temprano después de la fotodaña de UVB, ya que la migración de LC a los ganglios linfáticos ocurre después de la irradiación con UVB ( 19 ). Un informe reciente demuestra que los queratinocitos reducen la expresión de la integrina αvβ6 y αvβ8 después del daño de UVB, dando como resultado una regulación a la baja de la señalización de TGF-β en las LC y la posterior migración de LC de la epidermis ( 127 ). Durante la conversión maligna de tumores, las células epiteliales pueden perder su polaridad celular y propiedades adhesivas con el fin de volverse más invasivas y mesenquimales. Las LC también pueden desempeñar un papel en estas transiciones epitelio-mesenquimales (EMT) en cánceres cutáneos, ya que se sabe que muchas moléculas que intervienen en la migración de LC dentro y fuera de la epidermis desempeñan un papel en diversas etapas del proceso EMT ( 128 ) El TGF-β como factor de transcripción principal implicado en el comportamiento de LC también actúa como regulador principal de los procesos de EMT en cánceres de piel ( 129 ). BMP7 es importante para mantener las LC en la epidermis en estado de reposo. En los cánceres, BMP7 desempeña un papel similar ya que promueve la EMT invertida, es decir, la transición mesenquimal a epitelial, que actúa claramente como una señal de referencia hacia el epitelio ( 36 , 130 ). Uno de los principales eventos que caracterizan EMT es la pérdida de la expresión de E-cadherina, que como se mencionó anteriormente es un requisito previo para la migración de LC fuera de la epidermis ( 2 ). Curiosamente, como se mencionó anteriormente, la E-cadherina secretada induce la estabilización de la βcatenina, un evento que ocurre en diversos tumores cutáneos y conduce a la tolerancia en las CD ( 22 , 23 ). Queda por investigar si las LC tolerogénicas contribuyen a la tumorogénesis en tumores cutáneos activados por Wnt. El receptor tirosina quinasa Met se expresa en todas las DC y originalmente se caracterizó como un oncogen. La señalización de Met es crucial para la migración de LC de la piel a los ganglios linfáticos, ya que las LC en ratones que carecen de Met no llegan al ganglio linfático que drena la piel en condiciones inflamatorias ( 131 ). Curiosamente, la señalización Met media la actividad enzimática de MMP-2 y -9, las proteasas antes mencionadas que son cruciales para que las CL rompan la membrana basal al emigrar fuera de la epidermis ( 132 ). DISCUSIÓN Y PREGUNTAS PENDIENTES A pesar del hecho de que las LC se describieron por primera vez hace más de un siglo, todavía hay muchas preguntas pendientes con respecto a sus funciones biológicas y redundancias en la homeostasis y la patología de la piel. La identificación de subconjuntos dérmicos de Langerin + DC, independientemente de las CL epidérmicas que están en tránsito hacia los nódulos linfáticos que drenan la piel, cuestionó la validez del paradigma LC y, por lo tanto, la función biológica de las LC en diferentes contextos patológicos debe revisarse. Para comprender mejor la biología de LC, se necesitan estudios más amplios sobre biología de LC humana. Estudios altamente elegantes han indicado recientemente la complejidad de las CD humanas, revelando la existencia de múltiples precursores pre-DC que tienen propiedades funcionales distintas ( 133 ). También se necesitan modelos de mouse que permitan la orientación específica de LC, sin afectar a otros subconjuntos de DC. El uso de modelos de ratón que permitan la selección inducible de LC puede ser beneficioso sobre la deleción LC constitutiva, ya que podría ser que la ausencia de subconjuntos leucocitarios específicos durante toda la vida afecte el desarrollo normal de la piel y la homeostasis. Una mejor comprensión de las contribuciones relativas de los subtipos de DC residentes en la piel en el montaje de la plétora de respuestas tisulares que ocurren en la piel ante diversos insultos y traumatismos mejorará el desarrollo de las vacunas de DC y otras terapias dirigidas. La piel alberga trillones de microorganismos que han demostrado ser cruciales para la homeostasis y la inmunidad de los tejidos. Se demostró que los ratones que carecen de LC funcionales montan respuestas de células T normales en relación con los compañeros de camada de control después de la infección con la piel comensal Staphylococcus epidermidis ( 134 ). Sin embargo, las CL son cruciales para elevar una respuesta protectora humoral después de la aplicación tópica de la toxina A exfoliativa derivada de S. aureus ( 135 ). Estos datos apuntan al uso potencial de las terapias basadas en Langerina en la protección contra la infección, aunque la restricción microbiana por LC parece estar basada en el uso del receptor y, por lo tanto, solo se puede utilizar contra ciertos microbios ( 136 ). Queda por investigar si las LC son actores clave en el establecimiento de un microbioma saludable de la piel. También valdría la pena investigar si las LC se desarrollan normalmente en ausencia de microbios comensales, como sería el caso en animales libres de gérmenes. Está claro que la alteración de las funciones de LC podría ser una vía terapéutica interesante en diversos contextos de enfermedad. Los glicanos son determinantes cruciales en la unión de los receptores de lectina de tipo C, tales como Langerin, a sus ligandos y pueden modificarse para alterar la inmunomodulación en muchos niveles, desde el reconocimiento inmunitario de patógenos hasta la regulación de la respuesta inmune adaptativa, que podría explotarse para modular la vigilancia inmune, como se revisó en otra parte ( 137 , 138 ). En la era actual de las tecnologías de células individuales, es posible preguntarse si existen subconjuntos múltiples dentro de la población LC y cómo estos podrían afectar la respuesta inmune del tejido. Tales estudios podrían arrojar luz sobre los roles controvertidos que se han atribuido a las LC en la infección y el cáncer, y allanarían el camino para la orientación terapéutica de este tipo celular único.