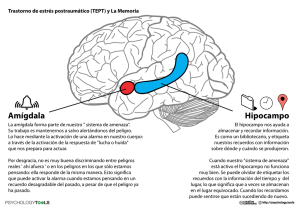
La amígdala y su relación con el autismo, los trastornos conductuales y otros trastornos del neurodesarrollo
Anuncio

CONFERENCIA MEMORIAL DR. MUÑOZ YUNTA La amígdala y su relación con el autismo, los trastornos conductuales y otros trastornos del neurodesarrollo Víctor L. Ruggieri Resumen. La amígdala está relacionada con el reconocimiento del significado afectivo del estímulo, la memoria a largo plazo, la orientación del estímulo social y la percepción de orientación de la mirada. Desempeña un papel fundamental en el reconocimiento de caras, en especial la de temor, y permite la comprensión de diversos estados emocionales, los cuales facilitarán una adecuada cognición social. Disfunciones de la amígdala se han relacionado con diversos trastornos del neurodesarrollo y con alteraciones neurocognitivas y conductuales en entidades neurogenéticas específicas. Múltiples estudios focalizados en el complejo amigdalino han permitido comprender muchos aspectos fisiopatológicos y formular nuevas hipótesis en relación con su génesis. Dado que los trastornos o entidades en que se ha evocado el papel de la amígdala son cada vez más extensos, este artículo remite a aquéllos que han despertado mayor interés en los últimos años, dividiéndolos en dos grupos: trastornos del desarrollo y conductuales (autismo, trastornos de ansiedad, trastorno bipolar, alexitimia y anorexia nerviosa), y entidades neurogenéticas específicas (síndromes del cromosoma X frágil, Rett, Prader-Willi y Williams), en las cuales se han comprobado alteraciones estructurales o disfunciones que pueden relacionarse con la sintomatología neurocognitiva y conductual de éstas. Es importante recordar que la amígdala es una estructura altamente conectada que conforma verdaderas redes funcionales, se ha asociado a diversos trastornos cuya explicación es variada e incluye diversos fenómenos fisiopatológicos, por lo que no debe simplificarse de una forma reduccionista su papel, sino también jerarquizar disfunciones de otras áreas que interactúan con ella. Servicio de Neurología. Hospital de Pediatría Prof. Dr. Juan P. Garrahan. Buenos Aires, Argentina. Palabras clave. Alexitimia. Amígdala. Anorexia nerviosa. Autismo. Síndrome de Prader-Willi. Síndrome de Rett. Síndrome de Williams. Síndrome X frágil. Cómo citar este artículo: Ruggieri VL. La amígdala y su relación con el autismo, los trastornos conductuales y otros trastornos del neurodesarrollo. Rev Neurol 2014; 58 (Supl 1): S137-48. Introducción La amígdala ocupa un lugar preponderante en el circuito de la empatía, es fundamental en el aprendizaje emocional y su regulación [1,2], y desempeña también un papel importante en el desarrollo de la teoría de la mente [3]. Desde el punto de vista filogenético, es de las estructuras más antiguas del cerebro, y su presencia se reconoce en animales 250 millones de años antes de la aparición de los mamíferos; incluso un homólogo suyo se encuentra en los anfibios [4]. Las transformaciones sustanciales de la estructura del complejo amigdalino, en la medida que se avanza desde los insectívoros a los primates, son la disminución de las regiones olfatorias (en especial las afectadas por el órgano vomeronasal y la detección de feromonas) y un aumento en el tamaño de los núcleos profundos, como el basal y el lateral [5]. Por otra parte, observamos un mayor tamaño del núcleo lateral en el cerebro humano comparado con los primates no humanos [6], lo que se relaciona con el gran desarrollo de su neocórtex y sus conexiones. www.neurologia.com Rev Neurol 2014; 58 (Supl 1): S137-S148 El complejo amigdalino interactúa con múltiples sistemas funcionales del cerebro, y puede considerarse como parte de una estructura integrada a un todo [7], conformada por una serie de núcleos (grupo basolateral, núcleo centromedial y núcleo cortical; el grupo basolateral se puede dividir, a su vez, en núcleo lateral, basal y núcleos basales accesorios) [8]. Muchos de esos componentes internos tienen importantes conexiones intrínsecas [9]. La amígdala está vinculada a la corteza orbital frontal, el cuerpo estriado y el tálamo, todas las estructuras implicadas en el procesamiento emocional. Correspondencia: Dr. Víctor L. Ruggieri. Servicio de Neurología. Hospital de Pediatría Prof. Dr. Juan P. Garrahan. Combate de los Pozos, 1881. CP 1245 Buenos Aires, Argentina. E-mail: victorruggieri@gmail.com Declaración de intereses: El autor manifiesta la inexistencia de conflictos de interés en relación con este artículo Aceptado tras revisión externa: 10.01.14. © 2014 Revista de Neurología Trastornos del neurodesarrollo y conductuales relacionados con la amígdala Autismo El autismo es considerado un síndrome conductual, de base biológica, caracterizado por déficits en la socialización, la comunicación y el lenguaje, asociado a intereses restringidos, conductas estereotipadas y trastornos sensoriales [1]. S137 V.L. Ruggieri El déficit en la cognición social es el eje sobre el que giran los aspectos relacionados con la génesis de los trastornos del espectro autista (TEA). Diversos autores han comunicado en personas con TEA déficit en el reconocimiento facial [10], en la detección de expresión de emoción, en especial de temor [11], en la mirada egocéntrica y pobre lectura de los ojos [12], todos estos hallazgos relacionados con el funcionamiento amigdalino. La afectación en la percepción de caras y emociones generaría la imposibilidad de inferir segundas intenciones, comprender miradas, deseos y conductas de los otros, lo que afecta gravemente la conducta social y, consecuentemente, la empatía [1]. El tamaño de la amígdala ha sido objeto de múltiples trabajos y observaciones en las personas con autismo, y puede variar de acuerdo con la edad de las personas estudiadas, el sexo y la gravedad de los síntomas. Diversos autores [13-15] encontraron en niños un aumento del tamaño de la amígdala aproximadamente de un 15% en comparación con controles. No obstante, otros autores, estudiando a adolescentes o adultos jóvenes, no encontraron diferencias en el tamaño o incluso detectaron menor volumen, lo que se relacionó con mayor mirada de evitación y trastornos conductuales más graves [16,17]. Complementariamente, la amígdala continúa su crecimiento a través de los años en varones con desarrollo típico, lo que parece no ocurrir en el autismo [14]. En consecuencia, parece haber un sobrecrecimiento de la amígdala en la infancia que no continuaría en la adolescencia, como ocurre en adolescentes con desarrollo típico. Estos hallazgos parecen tener correlato con la hipótesis del sobrecrecimiento cerebral temprano observado en niños con TEA [18]. El agrandamiento de la amígdala observado en niños con TEA de 36 a 56 meses se relacionó con mayor afectación social y de la comunicación, y peor evolución a los 6 años [15,19]. Schumann et al [14] estudiaron, a través de resonancia magnética (RM), el tamaño de la amígdala en niños de 2,5 años con riesgo de padecer autismo comparados con niños con desarrollo típico, y los reevaluaron a los 4 años. Encontraron que los niños con autismo tenían sobrecrecimiento amigdalino, y que éste se desarrollaba antes de los 3 años, momento en que los síntomas comenzaban a hacerse evidentes. Desde el punto de vista neuropatológico, Schumann et al [14] encontraron, en 10 personas con TEA (de 10 a 40 años), menor número de neuronas en la totalidad de la amígdala y en el núcleo lateral comparados con controles. S138 Estudios con RM funcional (RMf ) en autistas, en comparación con controles normales, pusieron en evidencia la falta de activación de la amígdala derecha en respuesta al estímulo social, en relación con la percepción de la mirada de la cara, en especial de los ojos [20], y la falta de activación del giro fusiforme, menor activación de la amígdala y giro occipital, procesamiento de imágenes en áreas diferentes, y menor tamaño de la amígdala en autistas en relación con controles [21]. Complementariamente, Monk et al [22], estudiando los circuitos neurales del procesamiento emocional de caras (alegría, tristeza y enojo) en 12 personas con TEA con RMf, observaron algo peculiar: las personas con TEA tenían mayor activación de la amígdala que las personas control que no padecían autismo. No obstante, un análisis más completo permitió observar que en los TEA había una activación más intensa entre la amígdala y la corteza prefrontal ventromedial (redes afectadas en la regulación emocional), y una conectividad más débil entre la amígdala y el lóbulo temporal (en la vía relacionada con la identificación de expresiones faciales, en especial en la región más anterior del lóbulo temporal, que está íntimamente relacionada con el procesamiento de expresiones faciales emocionales). Estos hallazgos permiten comprender las dificultades en la regulación emocional y el procesamiento de caras en los TEA, y confirman la afectación de la amígdala, aunque como parte de un circuito más complejo y complementario. Empatía, autismo y su relación con la amígdala La empatía, definida como la atribución de los estados mentales de uno mismo y de otros, es lo que permite dar sentido o entender acciones de otras personas y reaccionar adecuadamente a los estados mentales de los otros [1]. En el autismo, la empatización puede estar afectada en diversos grados. Incluso las personas con síndrome de Asperger pueden comprender emociones simples, pero no complejas. La empatía es un componente esencial para la experiencia emocional y la interacción social, denota una respuesta afectiva a estados mentales directamente percibidos o imaginados, o estados de sentimientos inferidos por otra persona [23]. Ocurre cuando la observación o imaginación de estados afectivos en otros induce estados compartidos en el observador [1] y permite ‘ponernos en la piel del otro’. A través de ella podemos comprender los estados emocionales, lo que nos permite no sólo comu- www.neurologia.com Rev Neurol 2014; 58 (Supl 1): S137-S148 Conferencia Memorial Dr. Muñoz Yunta nicar e interactuar con otros en aspectos afectivos y placenteros, sino también predecir acciones, intenciones y sentimientos de otros. Esta motivación genera una verdadera conducta prosocial [23]. Los niños con autismo tienen baja reactividad en la mayoría de las áreas de los circuitos de la empatía [24], que varía de acuerdo con las pruebas realizadas y el paradigma utilizado. Se han detectado disfunciones o procesamiento atípico en personas con TEA en regiones relacionadas con el procesamiento de la mirada, como el área posterior del sulco temporal superior y la amígdala, ante el procesamiento de caras y emociones [21,25]. La relación de la afectación de la empatía en la génesis de los TEA y la detección de disfunciones en los circuitos que la incluyen nos permiten comprender la importancia de la amígdala y todos sus circuitos en su génesis. Trastorno de ansiedad/fobia social La ansiedad puede definirse como un estado emocional aversivo que se desencadena por señales intensas en ausencia de un detonante específico o externo (a diferencia del miedo, que es una reacción fisiológica ante una amenaza real o percibida). Los trastornos de ansiedad son muy comunes en la infancia, en especial los trastornos de ansiedad por separación, cuya prevalencia se estima en el 4% de niños y adolescentes. Su inicio puede ser antes de los 18 años, y se considera inicio temprano antes de los 6 años. Los síntomas centrales son ansiedad excesiva e inapropiada para el nivel del desarrollo del sujeto concerniente a la separación respecto de su hogar, o miedo a perder un familiar o las personas con quienes está vinculado. Diversos trabajos encontraron un mayor tamaño de la amígdala derecha en niños con trastornos de ansiedad generalizada y en adolescentes con conductas de inhibición temprana en la infancia [26, 27]. No obstante, no todos los trabajos coinciden con estos hallazgos; de hecho, Milham et al [28], estudiando a niños con ansiedad de separación, fobia social o trastornos de ansiedad generalizada, comunicaron una significativa disminución del volumen de la amígdala izquierda en comparación con controles normales. Esta discrepancia orienta a pensar que los trastornos de ansiedad son un grupo heterogéneo y que muchos de los hallazgos habrá que correlacionarlos con el trastorno de ansiedad específico. Desde el punto de vista funcional, Thomas et al [29] comunicaron un mayor refuerzo de la actividad amigdalina en respuesta a caras de temor en www.neurologia.com Rev Neurol 2014; 58 (Supl 1): S137-S148 niños con ansiedad comparados con sujetos normales [29]. McClure et al [30] encontraron en adolescentes ansiosos una mayor activación de la amígdala derecha frente a caras de temor en comparación con caras felices, pero sólo cuando la evaluación estaba dirigida a valorar el rango o nivel de temor [30] (preguntando, por ejemplo: ¿cuán temeroso estás tú?), pero no durante la evaluación de otros estados (por ejemplo: ¿cuán larga es la nariz?). Incluso la hiperactivación de la amígdala durante la representación interna de miedo se comparó entre adolescentes ansiosos y deprimidos [31]. Guyer et al [32], observando adolescentes controles y ansiosos, detectaron que estos últimos son más propensos a percibir personas desconocidas y es menos probable que quieran interactuar con ellas, y muestran hiperactivación de la amígdala durante la visualización de estímulos aparentemente ino­ fensivos, como pares sonrientes [32]. La amígdala también se ha encontrado hiperactivada frente a caras emocionalmente neutrales no sólo en personas con trastornos de ansiedad, sino en adultos con inhibición social, quienes tenían altos niveles de inhibición social en la infancia [33]. Estos hallazgos permiten inferir que la amígdala es anormal tanto a nivel estructural como funcional en niños y adolescentes ansiosos, y posiblemente en niños que no desarrollan ansiedad, pero sí conductas de temperamento inhibido. Por otra parte, los trastornos funcionales observados son similares a los notificados en los niños con TEA, lo que permitiría inferir que estas disfunciones observadas estarían relacionadas con trastornos de ansiedad, los cuales tienen alta comorbilidad en los TEA [34]. Trastorno del estado de ánimo/trastorno bipolar Los síntomas de depresión en la infancia pueden variar de acuerdo con el desarrollo evolutivo del niño y de sus características cognitivas. Podríamos decir que el sello del trastorno bipolar es la desregulación del humor, que puede manifestarse con episodios de manía con alta ansiedad y energía, y momentos depresivos o hipomanía, caracterizados por sentimientos de desesperanza y aislamiento, los cuales interfieren en la vida diaria [35]. Por otra parte, la manía y la depresión pueden coocurrir, fluctuar e incluso alternar rápidamente y resultar en cuadros de paranoia. Los episodios de manía se pueden manifestar por irritabilidad y conductas explosivas asociadas a pobre funcionamiento psicosocial, lo que afecta gra- S139 V.L. Ruggieri vemente al niño y su familia. Muchas veces pueden expresarse con formas menos graves, como sobreactividad, disminución del sueño, hablar mucho, aumento de la actividad social, pobre juicio, etc. Se ha notificado, en niños con trastorno bipolar, impedimento en el reconocimiento de emociones de expresiones faciales y en el funcionamiento social, lo que puede vincularse a una disfunción de la amígdala [36]. Desde el punto de vista estructural, se ha encontrado con RM un menor tamaño de la amígdala en comparación con personas con desarrollo típico, en un porcentaje del 10-16% [37]. Incluso se ha descrito una disminución del volumen de la amígdala en adolescentes con trastorno bipolar (lo que no ocurre en adolescentes con desarrollo típico) [38]. No obstante, estos hallazgos deben tomarse con cautela porque podrían estar influidos por la evolución de la enfermedad, comorbilidades, alto estrés asociado a estos trastornos o efectos neurotóxicos. Alexitimia El término ‘alexitimia’ se refiere al impedimento en la habilidad de identificar y describir los propios sentimientos y emociones [39]. Este trastorno afecta, sin duda, el desarrollo de la empatía, con la consiguiente afectación en la cognición social [1]. Es difícil definir un área específica relacionada con este trastorno. Debemos tener en cuenta que, si bien ciertas funciones cognitivas están localizadas en áreas específicas del cerebro, en general se acepta que la complejidad de los procesos del cerebro es posible gracias a que diversas regiones son multifuncionales. De hecho, es lo que nos permite entender activaciones o desactivaciones de una misma área en respuesta a diferentes estímulos, por lo que deben tenerse en cuenta el contexto, el diseño y el paradigma de las pruebas. Justamente en la alexitimia deben considerarse diversos paradigmas para poder identificar su respuesta y disfunción específica en relación con ella. En un interesante trabajo, Moriguchi y Komaki [39] analizaron, en personas con alexitimia, las respuestas de diversas áreas a diversos paradigmas con RMf, a los que dividieron en cuatro categorías: estímulo emocional externo, imaginación y fantasía, estímulo sensoriomotor y somatosensitivo, y estímulo que contiene contexto social. Sólo se analizará el relacionado con la respuesta al estímulo emocional externo, dado que en éste se encuentra involucrada la amígdala. Partiendo de la hipótesis de que las personas con alexitimia tienen déficits en el procesamiento emo- S140 cional, se realizaron trabajos analizando las respuestas neurales a diferentes estímulos emocionales externos, como mostrar expresiones de caras o situaciones emocionales. Se hicieron diversos estudios utilizando tomografía por emisión de positrones o RMf en los que se evaluaba el grado de alexitimia a través de la escala de alexitimia de Toronto (TAS-20) [40], basada en medir dificultades para identificar sentimientos, y describir sentimientos y pensamientos orientados externamente. Cuando los estudios se focalizaron en la observación de estados afectivos faciales, como el temor, se detectó baja activación de la amígdala y, a mayor grado de alexitimia ante expresión de tristeza, se encontró menor actividad de la amígdala, la ínsula, las áreas temporal superior y occipitoparietal, y el hipocampo [39]. Si bien existen múltiples estudios y teorías fisiopatológicas, sólo he puntualizado algunos que permiten demostrar que en la alexitimia se observa disminución de la activación de la amígdala y otras áreas implicadas en el procesamiento afectivo del estímulo del mundo externo. Por otra parte, la actividad neural de la amígdala y la corteza cingulada anterior y posterior se correlacionaba negativamente con los niveles de alexitimia, incluso en personas con anorexia nerviosa [41]. En conclusión, se han demostrado en la alexitimia bajos niveles del procesamiento afectivo en respuesta a disparadores de estímulos emocionales externos. Esto coincide con la impresión de los clínicos de que las personas con alexitimia se muestran emocionalmente aburridas y no afectuosas [39]. Anorexia nerviosa La anorexia nerviosa es un trastorno mental alimentario grave, con una prevalencia aproximada del 0,6%, con claro predominio en mujeres (80%), de comienzo general promedio a los 18 años (rango: 10-30 años), con una mortalidad del 5-18% por década, y posiblemente relacionada con caquexia o suicidio [42]. Se caracteriza por el rechazo a ingerir alimentos, el control del peso y la imagen corporal de forma obsesiva, y el temor a aumentar de peso, asociado a trastornos comórbidos, como ansiedad y depresión. En general, son personas que se mantienen físicamente activas, hacen ejercicio y rechazan hablar de su problema o recibir ayuda. Si bien la etiología aún no está aclarada, es probable que factores genéticos, neurobiológicos, psicológicos y socioculturales confluyan para desencadenar este trastorno. www.neurologia.com Rev Neurol 2014; 58 (Supl 1): S137-S148 Conferencia Memorial Dr. Muñoz Yunta En la anorexia nerviosa se reconocen diversos mecanismos fisiopatológicos, los cuales analizaremos muy sucintamente, sin intentar sobresimplificar su explicación. Algunos síntomas específicos de la anorexia nerviosa y sus comorbilidades están directamente relacionados con la amígdala, el sistema límbico y conexiones. Entre los más significativos están: – Preocupación por el peso y el aspecto corporal, donde la amígdala, la corteza frontal y la somatosensitiva se relacionan con la observación del tamaño del cuerpo. – Baja autoestima, vergüenza y disgusto, que, junto con la ínsula, están funcionalmente relacionados con la experiencia corporal, sentimientos y pensamientos. – Ansiedad relacionada con la amígdala y el hipocampo en la detección de peligro. – Impedimento en la empatía, ligando la experiencia corporal con pensamientos y sentimientos. Diversos factores hormonales han demostrado su importancia en la génesis de la anorexia nerviosa. La influencia del factor liberador de la corticotropina, la hipercortisolemia, los opioides, la grelina, la leptina y la oxitocina se han relacionado con diversas áreas del cerebro vinculadas con la conducta e intensión alimentaria, el sentido de saciedad y el temor [43,44]. Entre las áreas involucradas en la conducta alimentaria relacionadas con la influencia hormonal se encuentran el hipotálamo, el hipocampo, el eje hipotálamo-hipofisario, la amígdala y el complejo amigdalino, junto con sus conexiones y la ínsula. Se ha propuesto un mecanismo psiconeuroquímico en relación con los opioides [45], que, al comenzar la dieta, genera una sensación de bienestar y adaptación al apetito y disminución del gasto de energía, lo que lleva a un paso final de adicción a la dieta, adaptándose a padecer hambre [43]. Justamente, diversos tipos de opioides se relacionan con el comportamiento mediado con la recompensa y con la autoestimulación [43]. Es claro que la activación del eje hipotálamo-hipofisario adrenal y la hipercortisolemia desempeñan, probablemente, un papel importante en la anorexia nerviosa, y que hay una superposición de las áreas involucradas en la respuesta al estrés y las vías de motivación por los alimentos (por ejemplo, amígdala, hipotálamo, hipocampo e ínsula). La oxitocina ha demostrado también que desempeña un papel importante en la génesis de la anorexia nerviosa [44]. Existen proyecciones centrales de las neuronas de oxitocina a regiones a tra- www.neurologia.com Rev Neurol 2014; 58 (Supl 1): S137-S148 vés del cerebro y receptores involucrados con la oxitocina que se relacionan con la motivación alimentaria, incluyendo el hipotálamo, la amígdala, la corteza prefrontal y la ínsula. Los receptores de oxitocina son muy ricos en la amígdala y el hipotálamo (áreas integradoras de señales del apetito e importantes en el aprendizaje de la saciedad y del valor de su sentido) [44]. Estudios funcionales en pacientes con anorexia nerviosa y controles pusieron en evidencia, usando paradigmas relacionados con la alimentación, hipoactivación en el hipotálamo, amígdala, hipocampo e ínsula, áreas relacionadas con la motivación alimentaria, en mujeres con anorexia nerviosa activa en comparación con mujeres que se encuentran en recuperación de anorexia nerviosa y mujeres sanas [46]. Lawson et al [44] sugieren que la oxitocina tiene funciones en la regulación del apetito y su disfunción puede contribuir, junto con otros factores, a los síntomas de los trastornos alimentarios. Desde el punto de vista neuropsicológico y conductual, si bien muchas observaciones son controvertidas, es claro que muchos aspectos relacionados con la cognición social en la anorexia nerviosa se encuentran involucrados en las disfunciones de la amígdala. Diversos autores han comunicado afectación en la comprensión de pruebas de teoría de la mente y reconocimiento de emociones faciales (lectura de la mente en las pruebas de observación de los ojos) [42,47]. Hambrook et al [48] estudiaron perfiles psicométricos de empatización y sistematización y realizaron pruebas de cociente de espectro autista, y sólo detectaron que las mujeres con anorexia nerviosa tenían puntuaciones más elevadas de dicho cociente que controles normales, aunque sin cumplir con criterios de autismo. Gillberg et al [49] encontraron déficits en el cociente de empatía en mujeres con anorexia nerviosa e incluso notificaron convergencia entre la anorexia nerviosa y características de la alexitimia. Las alteraciones estructurales y funcionales detectadas en estudios realizados con RMf permiten inferir que la amígdala desempeña un papel importante en la neurobiología de la anorexia nerviosa y la cognición social [42]. Mediante diversos paradigmas, y sin involucrar necesariamente la teoría de la mente, se han detectado alteraciones en el funcionamiento emocional a través de pruebas del cociente de empatía y del test de alexitimia de Toronto [42]. Por otra parte, Gallagher y Frith [50] proponen que la amígdala no está relacionada con actividades S141 V.L. Ruggieri mentalistas per se, lo que permitiría comprender la falta de afectación de la teoría de la mente. No obstante, es importante tener en cuenta las características de la población estudiada en personas con anorexia nerviosa, ya que los trastornos de la teoría de la mente se encontraron en pacientes con muchos años de evolución, mientras que los estudios con pacientes de corta evolución no mostraron déficit en teoría de la mente [42]. Esto puede ser esencial para el abordaje temprano y el apoyo emocional en personas con anorexia nerviosa. Entidades neurogenéticas específicas Dado que no es el objetivo de este trabajo profundizar sobre las características específicas de cada una de las entidades que abordaré, simplemente haré una breve introducción de cada una de ellas, puntualizando sus aspectos clínicos y genéticos (muchos de los cuales serán los relacionados con disfunciones o alteraciones del complejo amigdalino y sus conexiones). Síndrome X frágil El síndrome X frágil (SXF) es la causa hereditaria ligada al cromosoma X reconocida más frecuente de discapacidad intelectual, TEA y trastornos del aprendizaje, y afecta a 1 de cada 4.000 varones y a 1 de cada 6.000 mujeres [51]. Los varones presentan un fenotipo orientador, aunque no específico, con alto peso al nacer, macrocefalia, cara alargada, mentón prominente, frente alta, orejas grandes, laxitud ligamentosa y macroorquidismo de aparición en la pubertad. La mayoría presenta retraso mental, que guarda una relación directa con el tamaño de la ampliación de la isla CGG [51]. Desde el punto de vista conductual, la hiperactividad, obsesiones y conductas autistas, caracteri­ zadas por rechazo al contacto visual, estereotipias, perseveraciones, obsesiones, aleteos, conductas de evitación, impulsividad, autoagresiones, trastorno en la comunicación verbal y no verbal, son los hallazgos más significativos. El autismo se ha descrito incluso en niños que portan la permutación. Desde el punto de vista genético, este síndrome se produce por una expansión inestable del trinucleótido repetido CGG, que se ubica en la región 5’ del gen FRM1, cuyo locus está ubicado en el extremo distal del brazo largo del cromosoma Xq28. La mutación completa presenta, usualmente, hipermetilación de la isla CpG, lo que resulta en el S142 silenciamiento completo del gen, el cual se expresa usualmente en los varones, que tienen el fenotipo clásico y más grave [51]. Desde el punto de vista fisiopatológico, esto genera la ausencia de la proteína correspondiente FRMP1, que regula el desarrollo citoplasmático neuronal, es mayor en la región postsináptica glutaminérgica [51], y está implicada en el desarrollo y la plasticidad neuronal. Justamente, la ausencia de dicha proteína sería la responsable de los defectos en la sinaptogénesis y la estructuración sináptica posnatal observadas en estos pacientes. Resumiendo, las personas con SXF tienen generalmente, desde el punto de vista clínico, discapacidad intelectual, dificultades sociales, rechazo al contacto visual, impulsividad, ansiedad e hiperexcitación. En el sistema nervioso central, los cambios volumétricos estructurales más consistentes en niños y jóvenes comparados con controles sanos incluyen el agrandamiento del caudado y la disminución de la amígdala y el vermis cerebeloso posterior [52]. La disminución del tamaño de la amígdala se describió también en un trabajo comparativo de personas con SXF (con o sin autismo), personas con autismo no sindrómico y controles normales [53]. Los niños con SXF tenían menor volumen de la amígdala, independientemente de que cumplieran criterios de autismo, comparados con aquéllos con autismo no SXF o controles. No obstante, no todos los trabajos son concluyentes con estos hallazgos. Watson et al [54] no encontraron diferencias volumétricas en el tamaño de la amígdala en 51 jóvenes con una media de 35 meses. Desde el punto de vista funcional, en estudios con RMf en niños y jóvenes se encontraron diferencias en la activación de la amígdala. Tomando como paradigma el rechazo al contacto visual, Watson et al [54] detectaron que, cuando los niños con SXF miraban a la cara con dirección a los ojos, mostraban mayor activación de la amígdala que los controles, e incluso dicha hiperactivación persistía aún ante la observación de sucesivas caras. Esto podría explicarse por una clara situación de ansiedad a la mirada a los ojos y permitiría entender la tendencia de estos niños a evitarla. Si bien son muchos los interrogantes del papel de la función de la amígdala en el SXF, es claro que la disminución del volumen y las alteraciones funcionales podrían indicar su importancia en el comportamiento ocular y los trastornos de ansiedad en él. Incluso Hessl et al [55] encontraron disfunciones y alteraciones estructurales en la amígdala en individuos con premutaciones. www.neurologia.com Rev Neurol 2014; 58 (Supl 1): S137-S148 Conferencia Memorial Dr. Muñoz Yunta Síndrome de Rett El síndrome de Rett es, probablemente, el mejor ejemplo de conductas autistas biológicamente determinadas, en el que las niñas afectadas cumplen el mismo patrón conductual y evolutivo sin importar los aspectos sociales, ambientales o familiares. Afecta casi exclusivamente a mujeres, aunque hay raros casos de varones afectados, y su frecuencia es de 1 por 10.000 niñas [56]. Clínicamente, los primeros síntomas pueden ma­ nifestarse entre los 6 meses y los 3 años, siendo los 18 meses la edad habitual de inicio. Entre los síntomas más persistentes se encuentran detención y deterioro psicomotor progresivo, excesiva ansiedad, déficits conductuales sociales, detención del crecimiento cefálico y conductas estereotipadas manuales (movimientos de lavado de manos, palmoteo, golpeteo de manos en la boca), que son altamente orientadores, junto con la pérdida del uso propositivo de las manos, bruxismo, episodios de aspiración de aire compulsivo con pausa respiratoria, convulsiones (75% de los casos), escoliosis y espasticidad [57]. Episodios de ansiedad, pánico, gritos y terror o períodos de depresión se comunicaron en el 70% de los casos [57]. Genéticamente, se trata de una entidad producida por un déficit de la proteína Mecp2 (metil CpG binding protein 2), la cual se une a las zonas metiladas del ADN [56]. Es un gen regulador de genes y, por mecanismos epigenéticos, mantiene inactivas otras proteínas (gen represor de genes) con elevada presencia en el cerebro y facilita el normal funcionamiento de otros genes, lo que permite el desarrollo de los tejidos embrionarios y la plasticidad cerebral [58]. Se ha comunicado un gran número de mutaciones diferentes en este gen, lo que podría explicar la variabilidad de expresión clínica [56,58]. Un hallazgo interesante en ratones knockout deficientes de MECP2 en áreas específicas de la corteza cerebral (amígdala, estriado e hipocampo) [59] fue que manifestaron síntomas conductuales similares al síndrome de Rett (trastornos de coordinación, mayor ansiedad, menor intensión social y déficit en condicionamiento del miedo) [60]. Teniendo en cuenta la región basolateral de la amígdala en la percepción y modulación de emociones (incluyendo temor, estrés y ansiedad), Adachi et al [61] examinaron si la pérdida de MECP2 en la región basolateral de la amígdala era suficiente para generar un fenotipo de síndrome de Rett. Desarrollaron un ratón en el que se silenciaba la actividad de MECP2 en esa zona (a través de un vec- www.neurologia.com Rev Neurol 2014; 58 (Supl 1): S137-S148 tor viral). Esto produjo en los ratones déficits similares al síndrome de Rett en cuanto a la ansiedad y la conducta de la memoria de aprendizaje. Estos autores inyectaron luego un complejo de histona deacetilasa, que es habitualmente reprimido por el MECP2, a ratones sanos, y observaron los mismos síntomas que en los que no tenían MECP2. Esto permite comprender que la ausencia de inhibición de la histona deacetilasa produce conductas simi­ lares a las que son generadas por la ausencia del MECP2, y que el MECP2 es un represor transcripcional de éste, que se expresa en la amígdala al no ser reprimido por el MECP2, generando entonces déficits en la amígdala que producen el fenotipo de ansiedad y afectación cognitiva social en el síndrome de Rett [61]. Es interesante destacar que éstos no producen los trastornos motores observados en el síndrome de Rett, simplemente permiten comprender aspectos conductuales de éste y la importancia de la amígdala en estos aspectos. Síndrome de Prader-Willi El síndrome de Prader-Willi es una entidad de presentación esporádica, cuya alteración genética se caracteriza por una deleción en el brazo largo del cromosoma 15 (15q11-13) de origen paterno o disomía uniparental materna. Se han sido identificado múltiples genes como responsables del fenotipo de este síndrome [62]: SNRPN, GABRB3, NECDIN, PAR1, PAR5, PAR7 y MAGEL2. Estos niños presentan grave hipotonía en el período neonatal, hiporreflexia, dificultades alimentarias en los primeros meses, manos y pies pequeños, hipogonadismo y afectación cognitiva [57,62]. Las dificultades alimentarias van cediendo a lo largo del primer año y son reemplazadas por una incontrolable hiperfagia. La hiperfagia, junto con la obesidad mórbida, domina posteriormente el fe­ notipo. Presentan retraso mental leve a moderado, con déficits en aritmética, escritura, memoria visual y auditiva inmediata, y afectación en la atención auditiva. Desde el punto de vista conductual, el elemento más distintivo es su actitud alimentaria. Muestran a partir del primer año de vida un apetito insaciable, que rápidamente los lleva a la obesidad mórbida si el niño no recibe tratamiento nutricional y conductual adecuado. Intentan obtener alimentos incluso a escondidas o robándolos. Ante la postura de límites respecto de la alimentación, presentan ansiedad, berrinches e incluso auto y heteroagresión. También S143 V.L. Ruggieri se han comunicado trastornos de sueño, conductas autistas, depresión y tendencia al suicidio [62]. Justamente, el fenotipo conductual alimentario en este síndrome es objeto de constante análisis pa­ ra poder comprender sus bases fisiopatológicas e incluso las de la obesidad. Los hallazgos neuropatológicos no han sido suficientemente consistentes para explicar la hiperfagia, y se ha descrito aumento del tamaño de los ventrículos laterales [63,64], anormalidades en el núcleo dentado del caudado [64] y atrofia cortical frontal [63], entre otros hallazgos. Los estudios funcionales a través de RMf han demostrado algunos hallazgos interesantes. Las áreas implicadas en el apetito y la motivación alimentaria son las regiones límbicas y paralímbicas del cerebro, incluyendo la corteza orbitofrontal, la prefrontal media, la ínsula y la amígdala [65]. En las personas obesas, la corteza prefrontal desarrolla mayor actividad durante la saciedad [66], lo que indicaría un reclutamiento de dicha área con el objeto de suprimir las áreas de motivación del alimento (límbica y paralímbica). Holsen et al [66], en un interesante trabajo con personas obesas que se compararon con personas con síndrome de Prader-Willi, utilizando RMf y un paradigma de observación de imágenes de comida, de animales o figuras borrosas, cuatro horas antes de comer y posprandial (con 500 kcal), demostraron que las personas obesas tenían mayor activación ante las imágenes de comida antes de la alimentación que después de ésta en la amígdala, la corteza media prefrontal y el opérculo frontal, mientras que las personas con síndrome de Prader-Willi mostraron mayor activación frente a imágenes de alimentos después de la alimentación que antes de ésta en la corteza occipitofrontal, la ínsula, el hipocampo, la corteza prefrontal, el giro parahipocampal y la amígdala. Esto explicaría un distinto mecanismo en la hiperfagia entre obesos y síndrome de PraderWilli. Después de comer, las personas con síndrome de Prader-Willi muestran hiperfunción del sistema límbico y paralímbico, regiones que conducen a conducta alimentaria y motivación alimentaria (por ejemplo, amígdala), e hipoactivación de las áreas corticales que suprimen la intensión alimentaria y el autocontrol de la decisión de alimentación [67]. La amígdala aumenta su activación en estado de apetito y en respuesta a la visión de comidas calóricas, lo que la relaciona con la respuesta motivacional alimentaria y define el comer mientras tiene apetito. Incluso existe una conexión entre la amígdala y la respuesta alimentaria diferente de acuerdo S144 con el estímulo alimentario en el contexto de diversos niveles calóricos [68]. En resumen, la amígdala exhibe una gran activación después de la alimentación, lo que llevaría a la falta de sentido de saciedad o a continuar con intensión alimentaria. Estos hallazgos permiten mantener la hipótesis de que la conducta alimentaria en el síndrome de Prader-Willi se produce por la hiperactivación de los circuitos subcorticales de recompensa (la amígdala) relacionados con la falta de saciedad postalimentaria y la hipoactivación de las áreas corticales inhibitorias del apetito tras la alimentación (corteza orbitofrontal) [67]. Síndrome de Williams El síndrome de Williams es, probablemente, uno de los síndromes con fenotipo cognitivo y conductual más homogéneo y claramente definido. Se produce por una microdeleción en el brazo largo del cromosoma 7 (7q11-23) [69]. Desde el punto de vista clínico, los niños con este síndrome se caracterizan por tener facies peculiar de ‘duendecillo’ y presentan retraso en el desarrollo psicomotor asociado a cardiopatía, en general estenosis pulmonar [69]. En general, presentan retraso mental, aunque un 5% alcanza un rendimiento limítrofe [57]. Tienen claros trastornos en la motricidad gruesa y fina y afectación en los aspectos visuoperceptuales. Por otra parte, demuestran ser habilidosos en tareas de teoría de la mente. Su lenguaje expresivo es mejor que el comprensivo, y se caracteriza por frases gramaticalmente correctas, fluidas e incluso complejas, pero superficiales, estereotipadas o fuera de contexto. Conductualmente se caracterizan por ser amistosos y sociables con los adultos, pero su relación con los pares es pobre. Justamente esa sociabilidad y el lenguaje hacen sobrestimar sus capacidades. Debe tenerse en cuenta que la mayoría son incapaces de llevar una vida independiente [57]. La desinhibición social, la ansiedad, la distractibilidad y la tendencia a ser extremadamente amigables sin temor a los desconocidos juegan justamente en contra de su independencia e inserción laboral [70]. La mayoría de las personas con síndrome de Williams tiene, al menos, un trastorno de ansiedad [71]. Desde el punto de vista neuropatológico, Galaburda y Bellugi [72], analizando el cerebro de una persona fallecida con síndrome de Williams, detectaron disminución del tamaño de la amígdala, en especial del área dorsal del núcleo lateral. www.neurologia.com Rev Neurol 2014; 58 (Supl 1): S137-S148 Conferencia Memorial Dr. Muñoz Yunta Desde el punto de vista estructural, Järvinen-Pasley et al [73] comunicaron disminución del volumen del cerebro en áreas parietales y occipitales, mientras que estudios de volumetría [74] describieron que el tamaño de la amígdala estaba desproporcionadamente aumentado en 43 personas con síndrome de Williams en comparación con controles. Estudios funcionales evidenciaron disfunciones en la amígdala en personas con síndrome de Williams. Meyer-Lindenberg et al [75] evaluaron la respuesta de la amígdala a estímulos de amenazas sociales y no sociales (por ejemplo, caras o escenas) en adultos con síndrome de Williams y controles. Las personas con síndrome de Williams mostraban una menor activación de la amígdala ante caras, pero aumentaban la activación ante escenas de amenaza. Incluso en un estudio subsecuente detectaron hiperactivación de la amígdala a escenas de amenaza no social [76]. La hipoactivación de la amígdala a escenas de temor social también se demostró y se detectó una menor activación ante caras de temor [77]. Esto se relacionó, en un trabajo posterior de Haas et al [78], con la tendencia de estas personas a acercarse a extraños, justamente asociada a menor activación da la amígdala izquierda. Las imágenes funcionales sugieren que los individuos con síndrome de Williams muestran un aumento de la activación de la amígdala ante amenazas de estímulos no sociales y disminución de su respuesta ante amenaza social, hallazgos opuestos a los observados en la amígdala en personas sanas. Por otra parte, diversos autores han detectado que existe una reducción en la conectividad funcional de los centros de procesamiento de la cara en el cerebro entre el área fusiforme facial con la amígdala y corteza prefrontal en adultos con síndrome de Williams [79]. Nuevos estudios en niños serán fundamentales para poder comprender cómo todos estos hallazgos influyen en el desarrollo y el fenotipo conductual en el síndrome de Williams. Conclusiones Se ha analizado la importancia de la amígdala, el complejo amigdalino y sus conexiones en la génesis de diversos trastornos del neurodesarrollo y en su relación con los trastornos conductuales en diversas entidades neurogenéticas. Es importante jerarquizar el papel de la amígdala en fenómenos como la ansiedad y temores anormales, los cuales pueden ser el problema central o comórbido de diversos trastornos del desarrollo. www.neurologia.com Rev Neurol 2014; 58 (Supl 1): S137-S148 La identificación de fenómenos fisiopatológicos relacionados con la amígdala en diversos trastornos o entidades específicas permitirá probablemente, en un futuro, desarrollar terapias para esas disfunciones y de esa manera mejorarlas o incluso curarlas. Complementariamente, una manera interesante para comprender la importancia de las disfunciones del complejo amigdalino sería estudiar las enfermedades en las cuales éste se encuentra directamente afectado, entre otras, por ejemplo, la enfermedad de Urbach-Wiethe (proteinosis lipoídica), entidad autonómica recesiva [80] con calcificaciones amigdalinas progresivas asociadas a déficits en el juicio de expresiones faciales, asociación figuraolor, recuerdos de figuras negativas y positivas, e incluso ataques de pánico [80,81]; el síndrome de Capgras [82], producido por una desconexión traumática entre la amígdala y la corteza inferotemporal que genera desconocimiento de las personas conocidas, aunque reconociéndolas como iguales, pero pensando que son farsantes, lo que podría explicarse debido a la falta de la carga afectiva y emocional generada por esta desconexión a pesar de reconocerlas en todos los otros aspectos; la esquizofrenia, considerada un trastorno del desarrollo en el cual se han demostrado alteraciones funcionales y estructurales de la amígdala, aunque la forma infantil podría tener bases etiológicas diferentes a las del adolescente [79,83]; o las epilepsias del lóbulo temporal, asociadas a déficits en el reconocimiento de expresiones faciales, en especial el miedo, lo que podría relacionarse con los trastornos de conducta asociados a ellas [84]. Por otra parte, es importante analizar aspectos relacionados con la plasticidad del sistema nervioso central, dado que un abordaje precoz puede evitar mayor deterioro o incluso mejorar los síntomas. Seguramente el reconocimiento de genes específicos que tienen relación con el desarrollo del sistema amigdalino permitirá comprender sus funciones e interacciones, y así permitir un abordaje a la medida de cada una de estas disfunciones. Es probable que no haya una sola explicación y tampoco un abordaje específico sólo para la amígdala, pero el hecho de plantearse este desafío implicará en el futuro respuestas para la comprensión y tratamiento de muchos de estos trastornos. Bibliografía 1. 2. Ruggieri VL. Empatía, cognición social y trastorno del espectro autista. Rev Neurol 2013; 56 (Supl 1): S13-21. Morris JS, Friston KJ, Buchel C. A neuromodulatory role for the human amygdala in processing emotional facial expressions. Brain 1998; 121: 47-57. S145 V.L. Ruggieri 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. 16. 17. 18. 19. 20. 21. 22. 23. S146 Shaw P, Lawrence EJ, Radbourne C, Bramham J, Polkey CE, David AS. The impact of early and late damage to the human amygdala on ‘theory of mind’ reasoning. Brain 2004; 127: 1535-48. Laberge F, Mühlenbrock-Lenter S, Grunwald W, Roth G. Evolution of the amygdala: new insights from studies in amphibians. Brain Behav Evol 2006; 4: 177-87. Stephan H, Frahm HD, Baron G. Comparison of brain structure volumes in insectivora and primates. VII. Amygdaloid components. J Hirnforsch 1987; 28: 571-84. Barger N, Stefanacci L, Semendeferi K. A comparative volumetric analysis of the amygdaloid complex and basolateral division in the human and ape brain. Am J Phys Anthropol 2007; 134: 392-403. Swanson LW, Petrovich GD. What is the amygdala? Trends Neurosci 1998; 21: 323-31. Amunts K, Kedo O, Kindler M, Pieperhoff P, Mohlberg H, Shah N, et al. Cytoarchitectonic mapping of the human amygdala, hippocampal region and entorhinal cortex: intersubject variability and probability maps. Anat Embryol (Berl) 2005; 210: 343-45. Pitkänen A, Kelly JL, Amaral DG. Projections from the lateral, basal, and accessory basal nuclei of the amygdala to the entorhinal cortex in the macaque monkey. Hippocampus 2002; 12: 186-205. Dawson G, Webb S, McPartland J. Understanding the nature of face processing impairment in autism: Insights from behavioral and electrophysiological studies. Dev Neuropsychol 2005; 27: 403-24. Howard MA, Cowell PE, Bowcher J, Broks P, Mayes A, Farrant A, et al. Convergent neuroanatomical and behavioural evidence of an amygdala hypothesis of autism. Neuroreport 2000; 11: 2931-35. Calder AJ, Lawrence AD, Young AW. Neuropsychology of fear and loathing. Nat Rev Neurosci 2001; 2: 352-63. Mosconi MW, Cody-Hazlett H, Poe MD, Gerig G, GimpelSmith R, Piven J. Longitudinal study of amygdala volume and joint attention in 2- to 4-year-old children with autism. Arch Gen Psychiatry 2009; 66: 509-16. Schumann CM, Hamstra J, Goodlin-Jones BL, Lotspeich LJ, Kwon H, Buonocore MH, et al. The amygdala is enlarged in children but not adolescents with autism; the hippocampus is enlarged at all ages. J Neurosci 2004; 14: 6392-401. Sparks BF, Friedman SD, Shaw DW, Aylward EH, Echelard D, Artru AA, et al. Brain structural abnormalities in young children with autism spectrum disorder. Neurology 2002; 59: 184-92. Haznedar MM, Buchsbaum MS, Wei TC, Hof PR, Cartwright C, Bienstock CA, et al. Limbic circuitry in patients with autism spectrum disorders studied with positron emission tomography and magnetic resonance imaging. Am J Psychiatry 2000; 157: 1994-2001. Nacewicz BM, Dalton KM, Johnstone T, Long MT, McAuliff EM, Oakes TR, et al. Amygdala volume and nonverbal social impairment in adolescent and adult males with autism. Arch Gen Psychiatry 2006; 63: 1417-28. Courchesne E, Pierce K, Schumann CM, Redcay E, Buckwalter JA, Kennedy DP, et al. Mapping early brain development in autism. Neuron 2007; 25: 399-413. Munson J, Dawson G, Abbott R, Faja S, Webb SJ, Friedman SD, et al. Amygdalar volume and behavioral development in autism. Arch Gen Psychiatry 2006; 63: 686-93. Baron-Cohen S, Ring HA, Wheelwright S, Bullmore ET, Williams SE. Social intelligence in the normal and autistic brain: an fMRI. Eur J Neurosci 1999; 11: 1891-8. Pierce, Muller RA, Ambrose J, Allen G, Courchesne E. Face processing occurs outside the fusiforme ‘face area’ in autism: evidence from functional MRI. Brain 2001; 124: 2059-73. Monk CS, Weng SJ, Wiggins JL, Kurapati N, Louro HM, Carrasco M, et al. Neural circuitry of emotional face processing in autism spectrum disorders J Psychiatry Neurosci 2010; 35: 105-14. Batson CD. These things called empathy. In Decety J, Ivkes W, eds. The social neuroscience of Empathy. Cambridge: MIT Press; 2009. 24. Di Martino A, Ross K, Uddin LQ, Sklar AB, Castellanos FX, Milham MP. Functional brain correlates of social and non­social processes in autism spectrum disorders: an activation likelihood estimation meta-analysis. Biol Psychiatry 2009; 65: 63-74. 25. Wang AT, Lee SS, Sigman M, Dapretto M. Neural basis of irony comprehension in children with autism: the role of prosody and context. Brain 2006; 129: 932-43. 26. De Bellis MD, Casey BJ, Dahl RE, Birmaher B, Williamson DE, Thomas KM. A pilot study of amygdala volumes in pediatric generalized anxiety disorder. Biol Psychiatry 2000; 48: 51-7. 27. Hill SY, Tessner K, Wang S, Carter H, McDermott M. Temperament at 5 years of age predicts amygdala and orbitofrontal volume in the right hemisphere in adolescence. Psychiatry Res 2010; 182: 14-21. 28. Milham MP, Nugent AC, Drevets WC, Dickstein DP, Leibenluft E, Ernst M, et al. Selective reduction in amygdala volume in pediatric anxiety disorders: a voxel-based morphometry investigation. Biol Psychiatry 2005; 57: 961-6. 29. Thomas KM, Drevets WC, Whalen PJ, Eccard CH, Dahl RE, Ryan ND, et al. Amygdala response to facial expressions in children and adults. Biol Psychiatry 2001; 49: 309-16. 30. McClure EB, Monk CS, Nelson EE, Parrish JM, Adler A, Blair RJ, et al. Abnormal attention modulation of fear circuit function in pediatric generalized anxiety disorder. Arch Gen Psychiatry 2007; 64: 97-106. 31. Beesdo K, Lau JY, Guyer AE, McClure-Tone EB, Monk CS, Nelson EE, et al. Common and distinct amygdala-function perturbations in expressed vs anxious adolescents. Arch Gen Psychiatry 2009; 66: 275-85. 32. Guyer AE, Lau JY, McClure-Tone EB, Parrish J, Shiffrin ND, Reynolds RC, et al. Amygdala and ventrolateral prefrontal cortex function during anticipated peer evaluation in pediatric social anxiety. Arch Gen Psychiatry 2008; 65: 1303-12. 33. Schwartz CE, Wright CI, Shin LM, Kagan J, Rauch SL. Inhibited and uninhibited infants ‘grown up’: adult amygdalar response to novelty. Science 2003; 300: 1952-3. 34. Schumann CM, Amaral DG, eds. The human amygdala in autism. New York: Guilford Press; 2009. 35. American Psychiatric Association. Diagnostic and statistical manual of mental disorders. 5 ed. Arlington, VA: APA; 2013. 36. McClure EB, Treland JE, Snow J, Schmajuk M, Dickstein DP, Towbin KE, et al. Deficits in social cognition and response flexibility in pediatric bipolar disorder. Am J Psychiatry 2005; 162: 1644-51. 37. Garrett A, Chang K. The role of the amygdala in bipolar disorder development. Dev Psychopathol 2008; 20: 1285-96. 38. Blumberg HP, Donegan NH, Sanislow CA, Collins S, Lacadie C, Skudlarski P, et al. Preliminary evidence for medication effects on functional abnormalities in the amygdala and anterior cingulate in bipolar disorder. Psychopharmacology (Berl) 2005; 183: 308-13. 39. Moriguchi Y, Komaki G. Neuroimaging studies of alexithymia: physical, affective, and social perspectives. Biopsychosoc Med 2013; 7: 1-12. 40. Taylor Gb, Bagby RM, Parker JD. The 20-item Toronto Alexitimia Scale IV. Reliability and factorial validity in different languages and cultures. J Psychosom Res 2003; 55: 277-83. 41. Miyake Y, Okamoto Y, Onoda K, Shirao N, Okamoto Y, Yamawaki S. Brain activation during the perception of stressful word stimuli concerning interpersonal relationships in anorexia nervosa patients with high degrees of alexithymia in an fMRI paradigm. Psychiatry Res 2012; 20: 113-9. 42. Adenzato M, Todisco P, Ardito RB. Social cognition in anorexia nervosa: evidence of preserved theory of mind and impaired emotional functioning. PLoS One 2012; 7: e44414. 43. Hasan TF, Hasan H. Anorexia nervosa: a unified neurological perspective Int J Sci 2012; 8: 679-703. 44. Lawson EA, Holsen LM, Santin M, Meenaghan E, Eddy KT, Becker AE, et al. Oxytocin secretion is associated with severity of disordered eating psychopathology and insular cortex hypoactivation in anorexia nervosa. J Clin Endocrinol Metab 2012; 97: 1898-908. www.neurologia.com Rev Neurol 2014; 58 (Supl 1): S137-S148 Conferencia Memorial Dr. Muñoz Yunta 45. Yeomans MR, Gray RW. Opioid peptides and the control of human ingestive behaviour. Neurosci Biobehav Rev 2002; 26: 713-28. 46. Holsen LM, Lawson EA, Blum J, Ko E, Makris N, Fazeli PK, et al. Food motivation circuitry hypoactivation related to hedonic and nonhedonic aspects of hunger and satiety in women with active anorexia nervosa and weight-restored women with anorexia nervosa. J Psychiatry Neurosci 2012; 37: 322-32. 47. Tchanturia K, Happé F, Goldey J, Tresure J, Bara Carril N. ‘Theory of mind’ in anorexia nervosa. Eur Eat Disord Rev 2004; 12: 361-6. 48. Hambrook D, Tchanturia K, Schmidt U, Russell T, Treasure J. Empathy, systemizing, and autistic traits in anorexia nervosa: a pilot study. Br J Clin Psychol 2008; 47: 335-9. 49. Gillberg IC, Råstam M, Gillberg C. Anorexia nervosa 6 years after onset: Part I. Personality disorders. Compr Psychiatry 1995; 36: 61-9. 50. Gallagher HL, Frith CD. Functional imaging of ‘theory of mind’. Trends Cogn Sci 2003; 7: 77-83. 51. Arberas C, Fernández M, Altamirano E, Tello A. Síndrome de X frágil. Síndrome de retardo mental ligado al X con macroorquidismo. Síndrome de Martin Bell. Rev Hosp Niños 2002; 19: 35-41. 52. Gothelf D, Furfaro JA, Hoeft F, Eckert MA, Hall SS, O’Hara R, et al. Neuroanatomy of fragile X syndrome is associated with aberrant behavior and the fragile X mental retardation protein (FMRP). Ann Neurol 2008; 63: 40-51. 53. Hazlett HC, Poe MD, Lightbody AA, Gerig G, Macfall JR, Ross AK, et al. Teasing apart the heterogeneity of autism: same behavior, different brains in toddlers with fragile X syndrome and autism. Neurodev Disord 2009; 1: 81-90. 54. Watson C, Hoeft F, Garrett AS, Hall SS, Reiss AL. Aberrant brain activation during gaze processing in boys with fragile X syndrome. Arch Gen Psychiatry 2008; 65: 1315-23. 55. Hessl D, Rivera S, Koldewyn K, Cordeiro L, Adams J, Tassone F, et al. Amygdala dysfunction in men with the fragile X premutation. Brain 2007; 130: 404-16. 56. Ruggieri V, Arberas C. Trastornos generalizados del desarrollo. Aspectos clínicos y genéticos. Medicina (Bs Aires) 2007; 67: 569-85. 57. Ruggieri V, Arberas C. Fenotipos conductuales: patrones neuropsicológicos biológicamente determinados. Rev Neurol 2003; 37: 239-53. 58. Arberas C, Ruggieri V. Autismo y epigenética. Un modelo de explicación para entender la génesis de los trastornos del espectro autista. Medicina (Bs Aires) 2013; 73 (Supl 1): S20-9. 59. Chen RZ, Akbarian S, Tudor M, Jaenisch R. Deficiency of methyl-CpG binding protein-2 in CNS neurons results in a Rett-like phenotype in mice. Nat Genet 2001; 3: 327-31. 60. Gemelli T, Berton O, Nelson ED, Perrotti LI, Jaenisch R, Monteggia LM. Postnatal loss of methyl-CpG binding protein 2 in the forebrain is sufficient to mediate behavioral aspects of Rett syndrome in mice. Biol Psychiatry 2006; 595: 468-76. 61. Adachi M, Autry AE, Monteggia LM. MeCP2-mediated transcription repression in the basolateral amygdala may underlie heightened anxiety in a mouse model of Rett syndrome. J Neurosci 2009; 13: 4218-27. 62. Arberas C, Guillamondegui M, Fernández M, Tello A. Síndrome de Prader-Willi. Rev Hosp Niños 2005; 215: 311-20. 63. Hashimoto T, Mori K, Yoneda Y, Yamaue T, Miyazaki M, Harada M, et al. Proton magnetic resonance spectroscopy of the brain in patients with Prader-Willi syndrome. Pediatr Neurol 1998; 18: 30-5. 64. Hayashi M, Itoh M, Kabasawa Y, Hayashi H, Satoh J, Morimatsu Y, et al. A neuropathological study of a case of the Prader-Willi syndrome with an interstitial deletion of the proximal long arm of chromosome 15. Brain Dev 1992; 14: 58-62. 65. Holsen LM, Zarcone JR, Thompson TI, Brooks WM, Anderson MF, Ahluwalia JS, et al. Neural mechanisms underlying food www.neurologia.com Rev Neurol 2014; 58 (Supl 1): S137-S148 66. 67. 68. 69. 70. 71. 72. 73. 74. 75. 76. 77. 78. 79. 80. 81. 82. 83. 84. motivation in children and adolescents. Neuroimage 2005; 27: 669-76. Holsen LM, Zarcone JR, Brooks WM, Butler MG, Thompson TI, Ahluwalia JS, et al. Neural mechanisms underlying hyperphagia in Prader-Willi syndrome. Obesity (Silver Spring) 2006;14: 1028-37. Holsen LM, Savage CR, Martin LE, Bruce AS, Lepping RJ, Ko E, et al. Importance of reward and prefrontal circuitry in hunger and satiety: Prader-Willi syndrome vs simple obesity. Int J Obes (Lond) 2012; 36: 638-47. Killgore WD, Young AD, Femia LA, Bogorodzki P, Rogowska J, Yurgelun-Todd DA. Cortical and limbic activation during viewing of high- versus low-calorie foods. Neuroimage 2003; 19: 1381-94. Arberas C, Fernández M, Lapunzina P, De Eilzalde M, Tello A. Síndrome de Williams (síndrome de Williams-Beuren o hipercalcemia idiopática infantil). Rev Hosp Niños 1999; 183: 180-6. Einfeld SL, Tonge BJ, Florio T. Behavioral and emotional disturbance in individuals with Williams syndrome. Am J Ment Retard 1997; 102: 45-53. Woodruff-Borden J, Kistler DJ, Henderson DR, Crawford NA, Mervis CB, et al. Longitudinal course of anxiety in children and adolescents with Williams syndrome. Am J Med Genet C Semin Med Genet 2010; 15: 277-90. Galaburda AM, Bellugi UV. Multi-level analysis of cortical neuroanatomy in Williams syndrome. J Cogn Neurosci 2000; 12 (Suppl 1): S74-8. Järvinen-Pasley A, Bellugi U, Reilly J, Mills DL, Galaburda A, Reiss AL, et al. Defining the social phenotype in Williams syndrome: a model for linking gene, the brain, and behavior. Dev Psychopathol 2008; 20: 1-35. Reiss AL, Eckert MA, Rose FE, Karchemskiy A, Kesler S, Chang M, et al An experiment of nature: brain anatomy parallels cognition and behavior in Williams syndrome. J Neurosci 2004; 24: 5009-15. Meyer-Lindenberg A, Hariri AR, Muñoz KE, Mervis CB, Mattay VS, Morris CA, et al. Neural correlates of genetically abnormal social cognition in Williams syndrome. Nat Neurosci 2005; 8: 991-3. Meyer-Lindenberg A, Hariri AR, Mervis CB, Mattay VS, Morris CA. Abnormalities in neural processing of emotional stimuli in Williams syndrome vary according to social vs. non-social content. Neuroimage 2010; 50: 340-6. Haas BW, Mills D, Yam A, Hoeft F, Bellugi U, Reiss A. Genetic influences on sociability: heightened amygdala reactivity and event-related responses to positive social stimuli in Williams syndrome. J Neurosci 2009; 28: 1132-9. Haas BW, Hoeft F, Searcy YM, Mills D, Bellugi U, Reiss A. Individual differences in social behavior predict amygdala response to fearful facial expressions in Williams syndrome. Neuropsychologia 2010; 48: 1283-8. Schumann CM, Bauman MD, Amaral DG. Abnormal structure or function of the amygdala is a common component of neurodevelopmental disorders. Neuropsychologia 2011; 49: 745-59. Siebert M, Markowitsch HJ, Bartel P. Amygdala, affect and cognition: evidence from 10 patients with Urbach-Wiethe disease. Brain 2003; 126: 2627-37. Wiest G, Lehner-Baumgartner E, Baumgartner C. Panic attacks in an individual with bilateral selective lesions of the amygdala. Arch Neurol 2006; 63: 1798-801. Ramachandran VS. Consciousness and body image: lessons from phantom limbs, Capgras syndrome and pain asymbolia. Philos Trans Royal Soc B Biol Sci 1998; 353: 1851-9. Arango C, Kahn R. Progressive brain changes in schizophrenia. Schizophr Bull 2008; 34: 310-1. Golouboff N, Fiori N, Delalande O, Fohlen M, Dellatolas G, Jambaqué I. Impaired facial expression recognition in children with temporal lobe epilepsy: impact of early seizure onset on fear recognition. Neuropsychologia 2008; 46: 1415-28. S147 V.L. Ruggieri The amygdala and its relation to autism, behavioural disorders and other neurodevelopmental disorders Summary. The amygdala is related with the recognition of the emotional meaning of stimuli, long-term memory, the orientation of social stimuli and the perception of gaze orientation. It plays a fundamental role in the recognition of faces, especially those expressing fear, and makes it possible to comprehend different emotional states, which will facilitate an appropriate social cognition. Dysfunctions of the amygdala have been associated to a number of different neurodevelopmental disorders as well as neurocognitive and behavioural disorders in specific neurogenetic entities. A number of studies focused on the amygdalic complex have allowed researchers to understand many pathophysiological aspects and to formulate new hypotheses regarding their origins. Given that the disorders or conditions in which the role of the amygdala has been evoked are becoming increasingly more extensive, this article refers the reader to those that have aroused the most interest in recent years. Thus, they can be divided into two groups: developmental and behavioural disorders (autism, anxiety disorders, bipolar disorder, alexithymia and anorexia nervosa) and specific neurogenetic entities (fragile X, Rett, Prader-Willi and Williams syndromes), in which structural or dysfunctional alterations have been observed that may be related with their neurocognitive and behavioural symptoms. It is important to remember that the amygdala is a highly connected structure that forms truly functional networks and has been associated to different disorders with varied explanations and includes several different pathophysiological phenomena. Its role must not, therefore, be simplified in a reductionistic manner, but also placed upon a hierarchy of dysfunctions in other areas that interact with it. Key words. Alexithymia. Amygdala. Anorexia nervosa. Autism. Fragile X syndrome. Prader-Willi syndrome. Rett syndrome. Williams syndrome. S148 www.neurologia.com Rev Neurol 2014; 58 (Supl 1): S137-S148