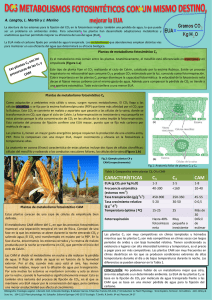
FOTORESPIRACION La fotorrespiración es un proceso que se produce en las plantas por el cual éstas utilizan oxigeno (O2) y producen dióxido de carbono (CO2) . Como dicho proceso sucede en presencia de la luz y el balance es semejante al de la respiración se denomina fotorrespiración. Pero a diferencia de la respiración, que es un proceso en el que se produce energía, la fotorrespiración no produce energía sino que la consume. Las plantas realizan fotosíntesis con el objeto de almacenar la energía solar en compuestos orgánicos altamente energéticos. En ese proceso de fotosíntesis las plantas toman dióxido de carbono del aire y liberan oxígeno. La fotorrespiración es, pues, un sistema contrario a la fotosíntesis y negativo para las plantas. La fotorrespiración se incrementa conforme aumenta la temperatura ambiente, lo cual sucede especialmente en días claros y soleados. A mayor temperatura, más tasa de fotorrespiración, llegando a igualar en ocasiones la tasa de fotosíntesis. En esos momentos el ritmo de crecimiento de las plantas se detiene. La causa de este proceso de fotorrespiración es la acción de una enzima que poseen las plantas. Esta enzima, denominada rubisco (ribulosa-1-5-bifosfato carboxilasa/oxigenasa) se comporta como fijadora de carbono en la fotosíntesis, pero a determinada temperatura empieza a comportarse como oxigenasa, es decir, capturadora de oxígeno. Las plantas que logran minimizar la fotorrespiración tienen una ventaja adaptativa sobre las demás y pueden colonizar medios áridos, secos y soleados. A las plantas que evitan la fotorrespiración se les denomina plantas C4 porque desarrollan un proceso en el que intervienen compuestos de cuatro átomos de carbono. PLANTAS C3 Es una planta «normal» —que no tiene adaptaciones fotosintéticas para reducir la fotorrespiración— se llama plantas C3. El primer paso del ciclo de Calvin es la fijación de dióxido de carbono mediante la rubisco, y las plantas que utilizan solo este mecanismo «estándar» de fijación de carbono se llaman plantas C3, por el compuesto de tres carbonos (3-PGA) que produce la reacción. Casi 85% de las especies de plantas del planeta son C3, como arroz, trigo, soya y todos los árboles. Plantas C4 En las plantas C4, las reacciones dependientes de la luz y el ciclo de Calvin están separadas físicamente: las reacciones dependientes de la luz se producen en las células del mesófilo (tejido esponjoso en el centro de la hoja) y el ciclo de Calvin ocurre en células especiales alrededor de las venas de la hoja. Estas células se llaman células del haz vascular. Para ver cómo ayuda esta división, veamos un ejemplo de la fotosíntesis C4. Primero, el CO2 atmosférico se fija en las células del mesófilo para formar un ácido orgánico simple de 444 carbonos (oxaloacetato). Este paso se lleva a cabo mediante una enzima no rubisco, PEP carboxilasa, que no tiende a unirse al O2. Después, el oxaloacetato se convierte en una molécula similar, malato, que puede transportarse hacia las células del haz vascular. Dentro de estas, el malato se descompone y libera una molécula de CO2. Luego, la rubisco fija el CO2 y lo convierte en azúcares a través del ciclo de Calvin, exactamente como en la fotosíntesis C3 Plantas C4 Este proceso tiene su precio energético: se debe gastar ATP para que la molécula de tres carbonos “ferry” regrese a la célula del haz vascular y quede lista para recoger otra molécula de CO2 atmosférico. Sin embargo, dado que las células del mesófilo constantemente bombean CO2 hacia las células del haz vascular vecinas en forma de malato, siempre hay una alta concentración de CO2 en comparación con O2 alrededor de la rubisco. Esta estrategia reduce al mínimo la fotorrespiración. La vía C4 se utiliza en cerca del 3% de todas las plantas vasculares; algunos ejemplos son el garranchuelo, caña de azúcar y maíz. Las plantas C4 son comunes en hábitats cálidos, pero son menos abundantes en zonas más frescas. En condiciones cálidas, los beneficios de una menor fotorrespiración probablemente superan el costo en ATP de pasar CO» de la célula del mesófilo a las células del haz vascular. Plantas CAM Algunas plantas adaptadas a ambientes secos, como las cactáceas y piñas, utilizan la vía del metabolismo ácido de las crasuláceas (CAM) para reducir al mínimo la fotorrespiración. Este nombre proviene de la familia de las plantas crasuláceas en las cuales los científicos descubrieron por primera vez esta vía. En vez de separar las reacciones dependientes de la luz y el uso de CO2 en el ciclo de Calvin en el espacio, las plantas CAM separan estos procesos en el tiempo. Por la noche, abren sus estomas para que el CO2 se difunda en las hojas. Este CO2 se fija en el oxaloacetato mediante la PEP carboxilasa (el mismo paso que usan las plantas C4, que luego se convierte en malato o un ácido orgánico de otro tipo. PLANTAS CAM El ácido orgánico se almacena dentro de vacuolas hasta el día siguiente. Durante el día, las plantas CAM no abren sus estomas, pero todavía pueden llevar a cabo la fotosíntesis. Eso se debe a que los ácidos orgánicos se transportan fuera de las vacuolas y se descomponen para liberar CO2, que entra en el ciclo de Calvin. Esta liberación controlada mantiene una alta concentración de CO2 alrededor de la rubisco. La vía CAM necesita ATP en varios pasos (no se muestran), así que, al igual que la fotosíntesis C4, no es un «regalo» energético. Sin embargo, las especies de plantas que usan la fotosíntesis CAM no solo evitan la fotorrespiración, sino que también usan el agua de forma muy eficiente. Sus estomas solo se abren por la noche, cuando la humedad tiende a subir y la temperatura a bajar, y ambos factores reducen la pérdida de agua de las hojas. Las plantas CAM suelen predominar en zonas muy cálidas y secas, como los desiertos. COMPARACION ENTRE C3 C4 Y CAM tIpo Separación de la fijación inicial de CO2 y el ciclo de Calvin Estomas abiertos Mejor adaptados a C3 No hay separación Día Ambientes frescos y húmedos C4 Entre el mesófilo y las células del haz vascular (en el espacio) Día Ambientes cálidos y soleados AM Entre el día y la noche (en tiempo) Noche Ambientes muy cálidos y secos Una de las clasificaciones más utilizadas a nivel académico entre las plantas angiospermas, o plantas con flor, es la que sirve para diferenciar entre monocotiledóneas y dicotiledóneas Qué es un cotiledón Los cotiledones no son otra cosa que esa primera hoja primitiva que las plantas producen en su forma de embrión. Es la que será su primera herramienta con la que recolectar luz solar y poder hacer la fotosíntesis, por eso esta pequeña hoja llamada cotiledón tan importante. Plantas monocotiledóneas Las plantas monocotiledóneas se caracterizan, como su nombre indica, por tener un único cotiledón. Pero esta dista mucho de ser su única diferencia con las dicotiledóneas. Las monocotiledóneas tienen un crecimiento muy diferente al de sus parientes de dos cotiledones. Este se debe, principalmente, a que estas no tienen cámbium, sin el que la producción de madera no es posible y, por tanto, no desarrollan tronco como tal, sino un tallo principal y más grueso. Por ejemplo, si cortas el “tronco” de una palmera, no verás anillos anuales de crecimiento como ocurre en los arbustos y árboles. Otra diferencia importante se da en las raíces de la planta. En las monocotiledóneas las raíces son adventicias, que significa que todas ellas nacen de la propia base del tallo. Esto hace que su sistema radicular sea mucho menos extenso y de un alcance menor, por lo que profundiza poco y, por tanto, tampoco pueden desarrollar un gran número de ramas. La última de las características básicas de las monocotiledóneas es que todas sus hojas muestran nervaduras visibles, que se extienden a lo largo de las mismas en paralela Plantas Dicotiledoneas Las dicotiledóneas son un grupo de plantas pertenecientes a las angiospermas, caracterizadas por la presencia de dos hojas primordiales o cotiledones en el “cuerpo” del embrión que está dentro de sus semillas. Las semillas de las plantas dicotiledóneas poseen un embrión con dos hojas embrionarias, primordiales o cotiledonares, generalmente carnosas y ricas en sustancias de reserva que nutren al embrión durante las primeras etapas de su desarrollo y durante el proceso inicial de la germinación. El embrión de una dicotiledónea está organizado anatómicamente de tal forma que se pueden distinguir: – Un vástago embrionario o plúmula, que posteriormente se convertirá el en vástago de la planta adulta – Una raíz embrionaria o radícula, a partir de la cual se desarrollará la raíz principal – Dos cotiledones u hojas embrionarias, que representan las primeras hojas de la plántula una vez germinada la semilla, y – Un hipocótilo, que es la porción que se encuentra entre la plúmula y la radícula Plantas monocotiledóneas: ejemplos Tulipan Rosales Lilium Alstroemeria Jengibre Vainilla Plantas Dicotiledóneas: ejemplos Laurel Canela Cilantro Comino Anis Cerezas Membrillos Higos ESTRUCTURAS EN VEGETALES Ramos (formaciones de menos de un año): Ramo de madera Es un brote lignificado de 70-100 cm y un diámetro superior a 1 cm, que solo posee yemas de madera. Son predominantes en nuevas plantaciones y con la edad su número se reduce. Chupón Es otro tipo de ramo vegetativo (de madera) que puede salir de cualquier parte del árbol, normalmente de ramas gruesas, por lo que tiene un gran vigor. Son fáciles de identificar ya que crecen siempre en sentido vertical, se hacen muy largos y la distancia entre los nudos es superior a la habitual. Salvo excepciones, se deben eliminar durante la poda en verde puesto que “chupan” alimento de la planta y no producen. Ramos (formaciones de menos de un año): Dardo Formación vegetativa de 7 cm de longitud como máximo, con una sola yema terminal de madera. Por su diminuto tamaño no es necesario podar, salvo en variedades muy productivas. Además, suelen estar situados en ramas gruesas, de manera que si evolucionan a lamburdas dan fruta de buena calidad. Ramos (formaciones de menos de un año): Sierpe Similar al chupón aunque procede directamente de la raiz. Deben ser eliminados, especialmente si se emplean herbicidas. Anticipado Son brotes que aparecen antes de tiempo a partir de una yema de madera, es decir, se forman y evolucionan en el mismo periodo vegetativo. En algunos casos suelen conservarse ya que podrán dar yemas de flor en la siguiente brotación. Ramos (formaciones de menos de un año): Ramo Mixto Ramo algo menos vigoroso que el de madera, con yemas laterales tanto de madera como de flor, siendo la terminal siempre de madera. Tiene un diámetro superior a 1 cm y es la formación característica de frutales de hueso. Ramo de Mayo Ramo más corto y delgado que el mixto (8-20 cm). La yema terminal es de madera, y las laterales de flor. Crece de golpe. Tamaño similar a lamburda o ramillete de mayo. Formación típica de frutales de hueso. Da fruta de menor tamaño, de manera que solo deben conservarse algunas. Ramos (formaciones de menos de un año): Brindilla Ramo vegetativo de aspecto débil, menor de 50 cm y diámetro inferior a 1 cm. Típica de frutales de pepita. En frutales de hueso es común en ciruelo . Cuando la yema terminal es fructífera se denomina brindilla coronada. Normalmente no nos interesa esta formación (tan solo si tenemos escasez de yemas) puesto que no dan la mejor producción. Ramas (formaciones de más de un año): Lamburda Formación de 5-10 cm. Es un dardo transformado, más largo, con la yema terminal de flor. (Foto izquierda) Bolsa Engrosamiento por acumulación de sustancias. Son formaciones que año tras año dan fruta. Es típica de peral, y algo menos en manzano. (Foto derecha)