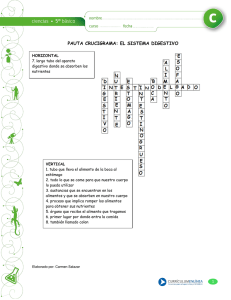
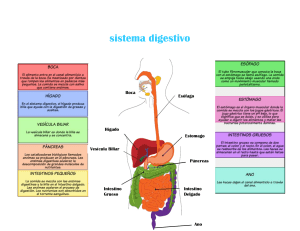
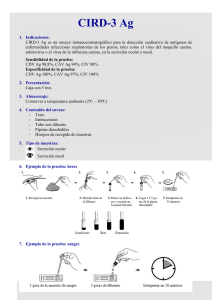
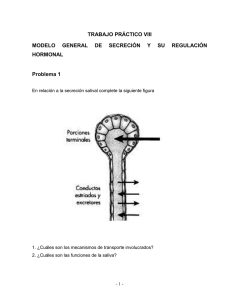
")
GUÍA BÁSICA FISIOLOGÍA DEL APARATO DIGESTIVO PARTE 1: MOTILIDAD El tubo digestivo es un conducto que trabaja de manera integrada con sus glándulas anexas. La luz representa el medio externo y siempre está llena de líquido. Las paredes del tubo digestivo pueden tener músculo liso (involuntario) y músculo estriado (voluntario). Músculo estriado: en primera porción del esófago (1/3 superior), faringe y esfínter anal externo Músculo liso: en el resto del tubo digestivo. Se dispone en capas que son: Circular interna: sincicio eléctrico (gran cantidad de uniones nexus), pocos receptores para Ach (acetilcolina), muchos receptores inhibitorios Longitudinal externa: de características opuestas Oblicua interna: sólo presente en el estómago, debajo de la submucosa En la capa submucosa se encuentra el plexo submucoso de Meissner, que regula las funciones de secreción y absorción. Mientras que el plexo mientérico de Auerbach, que está entre las dos capas musculares, es el centro integrador de todas las actividades. A este último le llegan aferencias vagales (colinérgicas) y del SNA, mediante proyecciones noradrenérgicas. Ambos forman parte del SNE (sistema nervioso entérico) Actividad eléctrica de la célula muscular lisa Una onda lenta es una variación eléctrica de la célula que no constituye un potencial de acción, tiene un componente rápido (fase 3) y uno lento (fase 4). La sucesión coordinada en tiempo y espacio de ondas lentas determinada por las células de Cajal (fibras especializadas auto excitables) constituyen el REB (ritmo eléctrico básico). Las células intersticiales de Cajal son similares a las fibras musculares pero se tiñen histológicamente como células nerviosas. Se encuentran sobre la capa circular interna o en el plexo mientérico de Auerbach. Las células de Cajal se agrupan formando estructuras denominadas marcapasos (osciladores), como es el caso del marcapasos gástrico, con un ritmo de 3 ciclos por minuto (cpm). El REB nunca desaparece en los sectores del tubo digestivo donde exista (es decir en estómago distal, todo el intestino delgado y todo el colon). Sin embargo, el REB puede desaparecer transitoriamente en colon bajo dos circunstancias determinadas: en sueño profundo (fase 4 o delta del sueño) y en ayuno. Motilidad Se define como un conjunto de fenómenos eléctricos y mecánicos que se producen en el tubo digestivo. Las células musculares pueden contraerse o relajarse de manera particular: CONTRACCIÓN TÓNICA: es una contracción constante, como sucede en el fundus gástri co y en los esfínteres CONTRACCIÓN FÁSICA: es una contracción periódica, si es coordinada hablamos de contracción propulsiva, si no es coordinada hablamos de mezcla. RELAJACIÓN ACTIVA: es la relajación directa RELAJACIÓN INACTIVA: es la relajación indirecta Unidad Motora Formada por el SNE, la célula intersticial de Cajal y la célula muscular lisa. Las fibras musculares (100 micrones) son la unidad eléctrica, mientras que los fascículos musculares (1200 micrones) son la unidad contráctil. Modelo general de motilidad (reflejo peristáltico) Estímulo: el bolo alimenticio (a nivel de la luz) sobre mecanorreceptores parietales. Vía Aferente Centro integrador: SNE, principalmente plexo mientérico de Auerbach Vía Eferente Efecto: peristalsis (motilidad de mezcla o motilidad propulsiva) *MOTILIDAD PROPULSIVA: contracción y relajación coordinada en tiempo y espacio *MOTILIDAD DE MEZCLA: contracción y relajación incoordinada y fásica Motilidad Propulsiva Por detrás del Bolo Alimenticio: Contracción de la capa circular interna (por Ach), estrechando la luz Relajación de la capa longitudinal externa (por SST, VIP, CCK que tienen como vía final común la producción de NO que actúa como relajante muscular) Por delante del Bolo Alimenticio: Relajación de la capa circular interna, aumentando la luz Contracción de la capa longitudinal externa, mediando la propulsión anterógrada Modelo de ayuno Se activa recién cuando se ha absorbido el 90% del contenido intestinal. Interviene el CMM (complejo motor migrante). Este es un conjunto de fases motoras que se va expandiendo por un gran sector del tubo digestivo: desde el marcapasos gástrico hasta la válvula ileocecal. El marcapasos gástrico está a 3 cm del cardias, sobre la cara posterior y tendiendo hacia la curvatura mayor, en la porción media del cuerpo gástrico. El marcapasos gástrico tiene influencia hasta la región antro-pilórica, dominando por su mayor frecuencia a otros marcapasos del sector, los cuales ciclan con una frecuencia de hasta 1 cpm. El marcapasos duodenal, por su parte, tiene una frecuencia de 12-14 cpm, la cual es decreciente en sentido distal: 9 cpm. En yeyuno e ileon 4 cpm. En el colon existe una innumerable cantidad de marcapasos, separados por distancias muy pequeñas, pero con una muy baja capacidad de acopl e entre las diferentes mesetas de frecuencia. Una meseta o platau es un sector determinado del tubo digestivo dominado por un marcapasos (oscilador) en particular. La propagación de las señales eléctricas (ya sean ondas lentas o espi gas) se produce gracias al acople eléctrico entre las mesetas. El CMM parte del marcapasos gástrico, pero se extiende hasta la válvula ileo cecal, pasando por todas las mesetas dominadas por el marcapasos correspondiente a cada sector del tubo digesti vo por donde pasó (entre el estómago y la válvula íleo cecal) La duración del CMM es de entre 70 y 110 minutos (media de 90 minutos). El CMM puede dividirse en fases: FASE I: fase de quiescencia. Se observa sólo el REB (sólo actividad eléctrica), ocupa entre el 40-60% del CMM. El REB del estómago, que funciona como marcapasos hasta la región antro-pilórica, cicla a 3-4 ciclos por minuto (ondas lentas). Los demás marcapasos: duodeno (12-14 cpm), yeyuno alto (10 cpm), yeyuno medio (9 cpm), yeyuno bajo (8 cpm), íleon proximal (7 cpm), íleon distal (6 cpm). FASE II: fase polifásica. Se caracteriza por presencia de REB y espigas que aumentan en frecuencia e intensidad a lo largo del tiempo, pero que nunca llegan a una re l aci ón onda lenta - espiga de 1:1. Ocupa entre un 20 y 30 % del total de la duración del CMM. FASE III: frente de actividad. Sólo espigas (actividad motora), relación onda lenta – e spi ga de 1:1. Las contracciones son intensas y con la máxima frecuencia, según el marcapasos de cada sector del tubo digestivo con CMM, por ejemplo: 3 cpm en el estómago, 12-14 cpm en el duodeno y así sucesivamente. FASE IV: fase polifásica inversa. Va disminuyendo paulatinamente la frecuencia y la intensidad. FUNCIONES DEL CMM: Sobre todo en la fase III. La principal función es la de evitar el sobre crecimiento bacteriano a nivel del estómago y del intestino delgado (el colon tiene flora bacteriana). Algunas ondas del marcapasos del estómago no llegan a porcione s más distales porque se agotan en el camino, por lo que va aumentando el tamaño de las colonias bacterianas a nivel distal (intestino grueso o colon). Movilizan grandes volúmenes de fluidos, por grandes distancias. Producen el barrido de alimentos y sustancias no digeribles. Modelo de ingesta: Interrumpe al modelo del ayuno (CMM) y es característico de la deglución. Se divide en 2 etapas: MASTICACIÓN DE ALIMENTOS: procesado mecánico y químico mediante la saliva (con enzimas como la amilasa y la lipasa) DEGLUCIÓN DEL BOLO ALIMENTICIO: desde la boca hasta el estómago. Tragar sólo implica pasar el alimento desde la boca hasta la faringe. En la porción inicial de la faringe, e l bol o distiende las paredes faríngeas y producen un reflejo que permite expandir l a cavidad para facilitar el paso del alimento y a su vez propulsarlo. La deglución, que es posible gracias al modelo de ingesta, es el proceso que lleva el alimento desde la boca hasta el estómago. Comprende varias etapas: oral, faríngea (mediante el refl ejo peristáltico) y esofágica (mediante las ondas esofágicas primarias o secundarias). 1) FASE ORAL: Es automática, voluntaria y consciente. Produce el procesamiento del alimento a través de la masticación, el contacto con las enzimas, y la imbibición e n sal i va (que facilita los anteriores fenómenos), formando el denominado bolo alimenticio. La masticación es un ejemplo de motilidad de mezcla, mientras que la motilidad de propulsión permite el pasaje del bolo a la orofaringe, donde al contactar con los me cano receptores (corpúsculos de Pommerenke) disparan el reflejo peristáltico (característico de la siguiente fase). 2) FASE FARINGEA: (reflejo peristáltico) Los mecano-receptores de las paredes son los corpúsculos de Pommerenke, sensibles a la distensión. Se estimulan y descargan por la vía aferente (mediante los pares craneales V, VII, IX y X) hasta el centro integrador e n e l SNC (núcleos sensitivos del NTS), el que opera deglución, vómito y defecación, solame nte. EL control se ejerce desde la formación reticular bulbar, sobre el piso de 4to ventrículo, cercano al centro de la defecación y del vómito. La vía eferente parte de los núcleo s motores de los pares IX y X (en este caso del núcleo dorsal del Vago), que tienen como inervación: boca, lengua, glándulas salivales, estómago. El efector siempre es el múscul o (estriado o liso) y los efectos se dan en forma simultánea. Este reflejo peristáltico es simultáneo a la activación de mecanismos de seguridad y precede a la onda esofágica (que es característica de la próxima fase) 3) FASE ESOFÁGICA: (ondas esofágicas) La onda esofágica se acopla en tiempo y espacio al reflejo peristáltico (perteneciente a la fase faríngea de la deglución) para hacer progre sar el bolo alimenticio en sentido distal. Las ondas esofágicas pueden ser primarias (precedidas de la deglución) efectivas o inefectivas, y secundarias (ondas de vaci amie nto para superar ondas primarias inefectivas). Las ondas terciarias son de mezcla (no de propulsión como las anteriores) y se consideran patológicas, excepto en neonatos y ancianos. Zonas de alta presión (ZAP): ESFINTER ESOFÁFICO SUPERIOR (EES): constituido por el músculo cricofaringeo. Es un esfínter anatómico, una ZAP, que se mantiene entre 40 y 80 mmHg por contracción tóni ca mediada por los pares craneales IX y X (en un 70%) y por su propia fuerza elástica (en un 30%). ESFINTER ESOFÁGICO INFERIOR (EEI): es un esfínter fisiológico, no anatómico. Conformado por 3-4 cm de músculo liso esofágico. Se mantiene entre 10-50 mmHg mediante: el propio músculo por un fenómeno miogénico (70%) ya que el umbral eléctrico es muy cercano al mecánico. Luego hay fuerzas externas como los pilares del diafragma (en un 20%) y las fuerzas elásticas (en un 10%). El EEI nunca se relaja en un 100%, solo podría llegar hasta un 95%. FUNDUS GÁSTRICO: mantiene una presión constante de 12 mmHg, por contracción tónica. Tiene función de reservorio de alimentos. Estadíos del modelo de ingesta: 1) Los mecanismos de seguridad se ponen en juego para evitar que los alimentos entren por una vía que no sea la digestiva. Entre estos mecanismos se cuentan: el cierre de las coanas para evitar la invasión a la nariz, hay una elevación del paladar blando para evitar la retrodifusión (el sistema de seguridad eleva la bóveda palatina y se cierra la glotis para evitar el reflujo del bolo alimenticio). Existe además, una apnea transitoria de aproximadamente 2 segundos por circuitos laterales en bulbo (donde se encuentran cercanos el centro respiratorio y el de la deglución). Todos estos fenómenos se producen de manera simultánea, por estímulo sobre los corpúsculos de Pommerenke, señal que se procesa a nivel del SNC. 2) Propulsión faríngea (reflejo peristáltico) 3) Relajación del EES, es pasiva. La inhibición se realiza a nivel de los núcleos de los pares IX y X. La relajación es de un 100% y permite que el bolo atraviese el esfínter. 4) Propulsión del esófago donde una onda esofágica pri maria efectiva permite que el bolo llegue hasta el sector superior del EEI. De resultar inefectiva no llegaría a ese nivel. 5) Relajación del EEI que es activa. Se produce sobre el músculo liso mediado por NO (vía final común de todos los relajantes musculares). Así el bolo está apto para llegar al fundus gástrico 6) Relajación receptiva refleja (RRR) del fundus gástrico. La presiónd e 12 mmHg se mantiene, ya que se relaja la fibra, pero manteniendo el volumen. Se da por control bulbar, y es el último evento del programa de la deglución. Se produce en respuesta de l a distensión del esófago distal ante el pasaje del bolo alimenticio, preparando las condiciones apropiadas para que el fundus pueda recibirlo. Todos estos eventos se dan simultáneamente desde el momento en que se “dispara” el programa de la deglución. Llenado gástrico: El llenado gástrico es consecuencia de la deglución y se produce en los 2/3 proximales donde se despliega la capa muscular oblicua externa (fundus motor con función de reservorio). La relajación adaptativa refleja (RAR) no forma parte del reflejo de la deglución y consiste en la rel ajaci ón de l fundus con la finalidad de poder recibir aún más carga. El centro integrador se encuentra en los plexos intramurales. Permite adaptar la longitud de la fibra muscular manteniendo constante la presión de 12 mmHg del fundus a pesar del incremento del volumen. Ocurre que al finalizar el proceso de la deglución, la RRR se agota en pocos minutos, con lo cual el fundus busca su tamaño anterior (recordar que estaba tónicamente contraído) y el contenido aumentado (por la entrada de bolo dentro de la luz) hace que se estimulen mecano-receptores de la pared del fundus, promoviendo la adaptación a los nuevos contenidos, mediante la RAR. Además, la RAR, es una respuesta al pasaje del bolo alimenticio al antro gástrico, estímulo para mecano -receptores parietales, integrando la señal en el SNE. La consecuencia de este estímulo es la relajaci ón de l as paredes musculares a este nivel. El cuerpo y el antro gástrico tienen motilidad propulsiva con función de mezcla por la existencia de una tercera capa muscular: la oblicua externa. Permite que el bolo llegue al píloro (porción distal de estómago). Este tipo de motilidad produce un flujo turbulento que transforma la fase sólida del bolo en una fase semilíquida, hasta lograr llegar a la misma, realiza pulsión y retropulsión, tendiendo a formar bolos de menos de 2 mm de diámetro, tamaño adecuado para poder atravesar el píloro. En el píloro solo pasan los bolos de menos de 2 mm. Si tiene un mayor tamaño hay retropulsión hasta que alcance el tamaño adecuado (menos de 2 mm). El antro se encarga, entonces, de que las partículas estén en condiciones físicas de atravesar el píloro, pero no es por sí solo el responsable de la evacuación gástrica. Evacuación Gástrica: El vaciamiento depende de dos variables: 1) del tamaño de la partícula y 2) del acoplamiento eléctrico o coordinación antro-píloro-duodenal dado por el SNE a través de circuitos locales. El vaciamiento se encuentra condicionado por el duodeno: el pasaje a través del mismo dispara el denominado efecto enterogastrona (de naturaleza neurohormonal, mediante SST y CCK – somatostatina y colecistokinina, respectivamente-), que frena, en mayor o menor medida el vaciamiento gástrico al provocar la incoordinación o desacople eléctrico a nivel del SNE. Tanto el exceso de osmolaridad, de volumen, o de carga lipídica, retrasan el vaciamiento gástri co (entonces estos son estímulos del efecto enterogastrona, o sea, se sensa en el duodeno que características tiene el contenido que ha abandonado el estómago y ha atravesado el píloro, de acuerdo a esas características se libera CCK y SST, quienes pasan a circulación y cuando llegan al estómago, inducen la incoordinación antro-píloro-duodenal para que se frene el vaciamiento gástrico y el duodeno tenga tiempo de cumplir sus funciones con la carga previamente re ci bi da – motilidad, secreción pancreática y biliar, absorción, etc-) El vaciamiento es diferente para sólidos y para líquidos: EVACUACIÓN GÁSTRICA DE LÍQUIDOS: En caso de que el líquido sea isoosmótico es inmediatamente evacuado por diferencia de presión entre el fundus y el duodeno, sin interrumpir el CMM. Si fuera hiperosmótico, se evacúa en forma directa pero interrumpiendo el CMM, con una mayor latencia. La velocidad de vaciamiento es proporcional al volumen, exponencialmente. Los líquidos se vacían directamente, gracias a la diferencia de presión entre el fundus y el duodeno, sin mediar la peristalsi s, l a cual e s fundamental para los sólidos, que son procesados por el antro hasta alcanzar el di áme tro adecuado para pasar a través del píloro. EVACUACIÓN GÁSTRICA DE SÓLIDOS: Nunca es una evacuación directa, el tamaño de las partículas es sensado en el píloro, además de analizar sus propiedades en cuanto a que sean partículas digeribles o no. La evacuación es lineal o semi -logarítmica, no es directamente proporcional a la velocidad de vaciamiento con el volumen. El estómago distingue entre sustancias digeribles y no digeribles, luego verifica el tamaño mediante e l píloro. Una vez que el alimento llega al duodeno, se dispara el reflejo peristáltico y el efecto enterogastrona (de acuerdo a las características del quimo) Motilidad específica: 1) BOCA: motilidad típica de mezcla, procesado enzimático mediante la saliva, da lugar a la formación del bolo alimenticio. Las enzimas más importantes son la lipasa lingual (formada por las glándulas de Von Ebner) que actúa sobre lípidos de cadena corta, y la enzima ptialina (o alfa-amilasa) que actúa sobre hidratos de carbono (de estas dos, la última es de mayor relevancia, ya que existe una lipasa pancreática, ergo la amilasa salival cobra más importancia con respecto a la lipasa salival). 2) FARINGE: motilidad propulsiva. El bolo tarda 10 segundos en pasar de la faringe al esófago proximal, momento en el que actúan las enzimas salivales, salvo la lipasa lingual que se activa dentro del medio ácido que provee el estómago. 3) ESÓFAGO: ondas esofágicas. Tiene un potencial de membrana de -90 mV, no tiene onda lenta (ni REB). ondas esofágicas pueden ser: PRIMARIAS (ondas propulsivas precedidas por una deglución -10 seg previos- es procesada en el SNC; puede ser efectiva o inefectiva) o SECUNDARIAS (también son propulsivas pero no son precedidas por una deglución, son ondas de vaciamiento esofágico, procesadas por el SNC y modulada por el SNE). Hay ondas TERCIARIAS (que suelen ser patológicas, el esófago adquiere motilidad de mezcla. Sol o e s fisiológica en recién nacidos y ancianos, por falta de maduración o por deterioro de los plexos nerviosos). 4) FUNDUS: tónicamente contraído (presión de 12 mmHg), su función es de reservorio. No se observa REB; el potencial de membrana se mantiene entre -25 y -35 mV. Se producen algunas variaciones en RRR y RAR, pero su actividad eléctrica es estable. 5) CUERPO Y ANTRO GÁSTRICO: en ayuno hay CMM, motilidad propulsiva con función de mezcla durante la ingesta. 6) DUODENO Y YEYUNOILEON: en ayuno hay CMM, motilidad propulsiva y de mezcla. 7) COLON: hay REB, ya que hay células de cajal. El derecho cicla a 4-11 cpm, con mezcla y absorción electrolítica, hay flora fermentativa. El transverso cicla entre 11-30 cpm, con peristalsis y contracciones en masa. El izquierdo cicla entre 15-36 cpm, con motilidad de mezcla, hay flora putrefactiva. Durante el sueño el colon tiene una actividad cero, durante el periodo de ayuno-postingesta el colon produce un mayor acople eléctrico para provocar defecación, aumentando la peristalsis. 8) COLON SIGMOIDE: función de reservorio de materia fecal. Cuando pasa a nivel del re cto, su distensión se vuelve conciente. El esfínter anal externo está formado por músculo estriado, y por lo tanto, es controlado por el SNC. 9) VESÍCULA BILIAR Y ESFINTER DE ODDI: en ayuno se observan oscilaciones en el CMM, en ingesta hay un control neurohumoral. Para algunos autores serían parte del CMM (en este caso CMS, complejo motor secretor), al igual que el EEI. ESOFAGOMANOMETRÍA : Mediante cambios en las presiones se puede estudiar la motilidad esofágica y del fundus estomacal. El esofagomanómetro utiliza un tubo de nitrógeno líquido conectado a una bomba de perfusión con agua destilada o solución fisiológica, con perfusión continua. Hay conectado a la bomba un sistema de tubos de polietileno con muy baja compliance al ser poco fl e xibl es (por l o que mantienen la misma presión de perfusión), que finaliza en un catéter, es decir un tubo obliterado en su extremo, por donde sale el líquido por diferencia de presiones. Si la presión intraesofágica fuera mayor que la del tubo, habría un reflujo del líquido perfundido, lo que es captado por una placa A/D conectada a un transductor, que registra una deflexión positiva cuando la presión intraesofágica resulta ser mayor y el líquido retorna. La esofagomanometría puede ser pasiva (en reposo) o activa (durante la deglución). La esofagomanometría pasiva utiliza un catéter que llega hasta el fundus gástrico, donde se observa una ondulación positiva en el registro (hay 12 mm de Hg de presión), pero se ven fluctuaciones que corresponden a las fases de contracción y relajación (por aumento de la presión en inspiración y disminución de la misma en espiración). Luego se retira el catéter algunos centímetros y en determinado sector la presión es de 50 mmHg, al llegar al EEI donde tambi é n se observan fluctuaciones por la misma razón. Se sigue retirando el catéter y se determina que el EEI se encuentra a 40 cm de la arcada de ntari a superior que es utilizada como referencia, teniendo en cuenta que el tubo digestivo está calibrado. En la porción torácica del esfínter se encuentra el punto de inversión respiratoria (PIR): ahora la inspiración provoca una deflexión negativa (hacia abajo, presión negativa) y la espiración una deflexión positiva (hacia arriba, presión positiva). El PIR se encuentra a 38 cm de la arcada dentaria superior. Se continua retirando el catéter hasta que la presión cae hasta 0 mm de Hg. Se encuentra en el tercio inferior del esófago, donde la espiración produce una deflexión positiva y la inspiración una deflexión negativa. Luego se registra un aumento de la presión a nivel del EES, a una di stanci a de 20 cm. Finalmente se registra una presión de 0 mm de Hg a nivel de la faringe, ampliamente comunicada con el exterior. Mediante la esofagomanometría activa puede estudiarse la motilidad del esófago en un se nti do dinámico. Se procede al registro de presiones mientras el paciente deglute, en este caso, agua. Se utilizan normalmente entre 5 y 7 catéteres intercomunicados y separados por una di stanci a de 5 cm uno del otro. Alteraciones en la motilidad esofágica Las alteraciones en la motilidad esofágica se traducen en un síntoma muy frecuente en clínica denominado disfagia. Esta alteración puede ser clasificada como orgánica (como en e l caso de l a esclerodermia y del cáncer de esófago) o inorgánica (también denominada funcional). En el caso de las alteraciones funcionales se trata de una sensación de atascamiento por algún trastorno sensorial, mientras que no existe una correlación entre el lugar percibido y el l ugar re al posible de atascamiento. Generalmente se realizan pruebas para localizar el sitio de la l e sión o l a mas ocupante mediante RX, sin arrojar resultados positivos. Este tipo de alteraciones funci onale s suele relacionarse más con el plano psico-somático que con el orgánico. Dentro de las alteraciones orgánicas pueden destacarse el espasmo esofágico difuso (EED), el nut cracker, la seudoacalasia y la acalasia. El EED se caracteriza por una motilidad de mezcla por superposición de ondas a nivel del cuerpo del esófago, mientras que el EEI no presenta alteraciones. El denominado “nut cracker” (rompe nueces), se caracteriza por ondas de alta intensidad y larga duración que provocan sensación dolorosa, mientras que el EEI no presenta alteración alguna. La seudoacalasia suele darse como consecuencia de un tumor del techo gástrico y por l a fal ta de relajación del EEI y por bloqueo de neuronas inhibitorias por la producción de sustancias de origen tumoral. La acalasia propiamente dicha se caracteriza por aperistalsis en el cuerpo esofágico y la no relajación del EEI. Radiológicamente se observa la característica imagen de mega esófago. En la acalasia congénita existe una falla en la migración de neuronas inhibitorias que a través de la producción de NO (óxido nítrico) controlan el EEI. Clearence ácido del esófago El clearence ácido del esófago es el barrido de la carga ácida proveniente del fundus gástrico. Define la capacidad que tiene el esófago para deshacerse de los protones (H+) que, generando un muy bajo pH, lesionarían gravemente la mucosa esofágica, la que no cuenta con barreras defensivas como la pared estomacal o la duodenal, por lo que debe recurrir a este mecanismo. Este es uno de los mecanismos de defensa indispensables contra el reflujo gastro -esofágico responsable de los síntomas como la pirosis (vulgarmente denominada acidez). Es de tener en cuenta que, si bien la mayoría de los reflujos son ácidos (reflujos gastro esofágicos), en un 20% estos reflujos son alcalinos. El último es el caso del reflujo entero -gástrico, que se produce generalmente ante una falla del cierre del píloro, con pasaje del contenido intestinal: HCO3-, sale s biliares, lisofosfolípidos (productos de degradación de los TAG), y enzimas, todos con capacidad de lesionar el epitelio esofágico (donde se vierten desde el estómago), con conse cue nci as aún más graves que en el caso del reflujo ácido. No menos de 1/3 de la población occidental ha sufrido alguna vez fallas en el clearence esofágico, provocando el cuadro característico de pirosis, con ardo retro esternal. Este cuadro, de resultar persistente, podría desencadenar en graves lesiones en la mucosa esofágica que, incluso, pue den terminar en cáncer de esófago. La causa más frecuente de falla del clearence esofágico es la relajación fuera de tiempo e inadecuada del EEI (en el 80% de los casos), además de otros problemas de motilidad. El clearence ácido del esófago es un mecanismo que cuenta con dos componentes: 1) HCO3-: bicarbonato proveniente de la saliva 2) ONDA ESOFÁGICA PRIMARIA: que moviliza el bicarbonato al tercio inferior del esófago, donde es necesario neutralizar la carga ácida proveniente del fundus gástrico por algún reflujo. El esófago posee un pH levemente alcalino (7,2-7,4) y posee un epitelio pavimentoso (plano estratificado) no queratinizado, el que podría dañarse por la carga ácida del HCl (ácido clorhídrico) de la luz estomacal. En condiciones fisiológicas, durante la deglución, el EEI se relaja, mientras que la presión del fundus gástrico se mantiene (12 mm de Hg), por lo cual el contenido ácido puede acceder por diferencia de presiones al tercio distal del esófago (con 0 mm de Hg de presión), el cual no ti e ne desarrollada una capa mucosa adecuada para defenderse de la misma, por lo cual necesita del mecanismo de clearence. La secreción salival provee de bicarbonato (con concentración de 0.16 M y en 0,7 ml por deglución) al tercio inferior del esófago gracias a una onda esofágica primaria efectiva (motilidad). El clearence se activa cuando el pH esofágico cae a nivel del pH estomacal (0,8-1,2) y se manti e ne hasta que el pH sube hasta 4, nivel que el esófago puede resistir sin suf rir daños mayores. Para alcanzar este nivel, se requieren, habitualmente entre 6 y 12 degluciones. Es de destacar que durante el sueño profundo (sueño delta) no hay deglución de saliva (ya que la motilidad e s nul a), con lo cual si hubo reflujo previo no podría hacerse efectivo el mecanismo de clearence . Por otra parte hay algunos alimentos como el café, el chocolate y la menta que aumentan el riesgo de reflujo gastro esofágico. Valoración del clearence ácido del esófago Puede realizarse mediante pruebas de clearence exógeno y pruebas de clearence endógeno. Clearence exógeno: se realiza mediante la instilación de 15 ml de HCL en concentración de 0,1 M a través de una sonda naso gástrica que llega hasta el tercio inferior del esófago. Se realizan degluciones cada 30 segundos y si el pH esofágico (registrado por un medi dor de pH) alcanza el valor de 4 UI (Unidades Internacionales) entre 6 y 12 degluciones, el clearence se considera normal. Clearence endógeno: se realiza mediante una sonda naso gástrica que se mantiene e ntre 4 y 24 hs, registrando número, duración e intensidad de los reflujos. Mediante este método pueden registrarse el reflujo más prolongado y el momento en el se produjo. PARTE 2: SECRECIÓN DE AGUA Y ELECTROLITOS, SALIVAL, GÁSTRICA Y PANCREÁTICA Otra de las funciones del tubo digestivo es la secreción de agua y electrolitos, que se cumple sobre la base del modelo general de secreción, el que (con características diferenciales) se repite en todos los tipos celulares que desarrollan esta función. Primero es necesario tener en cuenta las concentraciones de los iones más relevante s, tanto de l liquido intracelular (LIC) como del extracelular (LEC): ION Concentración en LIC Concentración en LEC Na+ 10-12 meq/l 135-145 meq/l K+ 135-145 meq/l 3,5-5 meq/l Cl+ 5-7 meq/l 110 meq/l Modelo general de secreción (MGS) Todas las células epiteliales que secretan agua y electrolitos utilizan mecanismos básicos comune s y mecanismos específicos (de los cuales depende la diferencia en los productos finales: protones o bicarbonato). Según el predominio de un mecanismo específico sobre el otro, se define el ti po de secreción de la célula. Teniendo en cuenta la polaridad de las células epiteliales, se di fere ncia un sector luminal (en contacto con la luz del tubo digestivo) y un sector basolateral (en contacto con la circulación y con las células vecinas). El modelo general de secreción cuenta con cinco componentes básicos que se repiten en todos los tipos celulares con función secretora y, por otro lado, herramientas específicas que determinan l a funcionalidad de las diferentes células (según tengan secreción básica o ácida). La dinámica del MGS depende del estímulo neurohormonal (como el de la secretina, por ejemplo), el que interactúa con receptores basolaterales que activan el mecanismo de secreción a travé s de la señalización por segundos mensajeros (como el AMPc y el IP3). La transducción de señal es tiene como objetivo final la fusión de túbulo-vesículas (TV) conteniendo las herramientas de se cre ción (preformadas) a la membrana luminal. Componentes básicos del MGS 1) Cotransporte 2Cl/Na+/K+: basolateral, la fuerza de arrastre osmótica el Na+ promue ve l a energía necesaria para este mecanismo (este transportador es el mismo que se encuentra en el asa ascendente gruesa de henle, que se bloquea por diuréticos del asa como la furosemida) 2) Canal de Cl: luminal 3) Canal de k+: luminal o basolateral, es voltaje dependiente. 4) Bomba Na+/K+ ATP asa: exclusivamente basolateral. Es el único mecanismo activo. 5) Vía paracelular: también denominada Shunt Path, se evidencia mejor en epitelios abiertos. Herramientas específicas del MGS 1) Contra transporte (antiporte) Cl/HCO3-: basolateral, absorción de cloro y secreción de bicarbonato. Sirve para alcalinizar el medio. 2) Antiporte Na+/H+: basolateral, absorbe sodio y secreta protones. Sirve para producir una secreción hipotónica (aunque isoosmótica) y para neutralizar el bicarbonato (formando ácido carbónico -H2CO3-) 3) Bomba de H+/K+: luminal, secreta protones y absorbe potasio Secreción salival Las glándulas salivales se dividen, a grandes rasgos, en mayores y menores. Las mayores tienen una secreción serosa (mayor concentración proteica) como las parótidas; una secreción mucosa (mayor concentración de agua) como las sublinguales; y una secreción mixta como las submaxilares. Las menores, por otra parte, producen un menor porcentaje de la carga salival pero se encargan de la secreción constitutiva (constante), a diferencia de la secreción de las glándulas mayores que necesitan de estímulos para desencadenarse. Las glándulas de Von Ebner, ubicadas a nivel de l a V lingual, producen la enzima lipasa lingual. En ambos casos, la unidad funcional de la glándula es el salivón, que se encuentra formado por un adenómero (con una porción acinar y otra ductular), los plexos nerviosos que lo inervan, y la microcirculación que lo irriga. Por su parte, la porción acinar del adenómero, se ocupa de la secreción proteica, mientras que su porción ductular se ocupa de la secreción hidro-electrolítica. El acino es el encargado de la producción de saliva primaria, que es isotónica, y con contenido predominantemente proteico. La misma se transforma en su trayecto dentro del sistema ductular, por el antiporte Na+/H+. La relativa acidez consecuente, activa luego al antiporte Cl -/HCO3-, para ahora, hacer a la saliva levemente alcalina, mediante un mecanismo de contracorriente. Finalmente, la saliva definitiva es hipotónica, alcalina e incolora. El volumen de secreción promedio es de entre 1000 ml y 1500 ml diarios. Es para tener en cuenta que todas las secreciones del tubo digestivo son de entre 1000-1500 ml, excepto la secreción pancreática y estomacal que son de entre 1500-2000 ml por día. La saliva cuenta con una gran concentración de Ca2+ y fosfato, mientras que el principal ani on e s el HCO3- (en mayores concentraciones que en el plasma). Entre los componentes orgáni cos más importantes se encuentran la proteína R (importante en la absorción de Vit B 12), tiacinatos (antibacterianos), IgA secretora (dimérica) y la enzima lipasa lingual (producida por las gl ándul as de Von Ebner). Secreción pancreática (exócrina) Esta vez la unidad funcional se denomina pancreón, el que está conformado por u n ade nóme ro, los plexos nerviosos y la microcirculación. El producto de secreción pancreática es incolor, isotónico y alcalino. El 85% está conformado por agua, el anión principal es el HCO3-, mientras que el catión pri nci pal es el Ca2+ (Na+ y K+ se encuentran en concentraciones similares al plasma). La precipitación de Ca2+ puede determinar la formación de cálculos, que normalmente se e vi tan por la acción de litostatinas (o PSP). Dentro de los componentes orgánicos se encuentran enzimas (en un 95%), las que se acti van por medio de la tripsina, a su vez activada mediante una enteroquinasa producida por la membrana luminal de los enterocitos, y por un mecanismo autocatalítico. El factor inhibidor de Katzal evita l a conversión de la proenzima tripsinógeno en tripsina dentro del conducto pancreático, evitando su acción lítica sobre la misma célula pancreática. Secreción estomacal La secreción de HCL (ácido clorhídrico), importante en la digestión de proteínas y en procesos como la absorción de Vit B12, corre por cuenta de las células parietales de la región ácido secretora del estómago (más que nada a nivel del cuerpo gástrico). Las células parietales, ade más de secretar HCL, secretan el denominado factor intrínseco de Castle. Por otra parte, las áreas mucosecretoras se ubican a nivel subcardial y prepilórica. Allí e xiste una gran densidad de células principales (productoras de pepsinógeno) y células mucosas. Ademas hay una alta concentración de células del sistema APUD (de producción neurohormonal) y glándulas tubulares ramificadas. Producción ácido clorhídirco La célula parietal tiene un estado de reposo y un estado activo. En repos o conserva una forma piramidal y en su citoplasma se evidencia gran concentración de túbulo vesículas (TV). Por su parte, luego de recibir el estímulo adecuado la célula se activa, produciendo la fusión de las TV. Los estímulos pueden ser: Gastrina, Histamina o Acetil colina (Ach). GASTRINA: producida por las células G antrales y duodeno-yeyunales. Estimulan al receptor basolateral denominado CCK-B, que tiene al PIP 2 y al Ca2+ como segundos mensajeros, hete aquí que se encuentra acoplado a proteína Gq, que estimula a una PLC (fosfolipasa C) que degrada el PIP2 (fosfatidil inositol difosfato) en IP3 (inositol trifosfato) que aumenta la conductancia al Ca2+ y al DAG (diacilgricerol) que estimula a la PKC (protein kinasa C) HISTAMINA: producida por mastocitos, enterocormafin like cells y por el sistema enzimático histidina decarboxilasa. Estimulan al receptor basolateral H2, que se encuentra acoplado a proteína Gs, con lo cual si activación produce un aumento de la concentraci ón de AMPc (segundo mensajero). Ach: desde los plexos nerviosos del SNE, actuando sobre receptores basolaterales M3, acoplados a proteína Gq, con lo cual tienen como segundos mensajeros al PIP2 y Ca2+ Los segundos mensajeros (AMPc y PIP2-Ca2+) promueven la fusión de las TV, con la instaura ci ón de los mecanismos necesarios para la secreción del HCL sobre la membrana luminal de la célula parietal. Luego de la fusión de las TV se activan inmediatamente los canales luminales para el K+, que tiende a salir de la célula y es vital para activar luminalmente la bomba H+/K+. Esta última hace que el K++ regrese al LIC y que se secrete H+ (derivado del H2O). En tercer lugar se produce la apertura de canales para el Cl-, que secretan este ion a la luz tubular. Por otra parte, dentro de la célula parietal se acumulan K+ y HCO3- (producto de la degradación del ácido carbónico por la enzima anhidrasa carbónica, la cual puede catalizar la reacción inversa). La membrana basolateral utiliza un antipote Cl-/HCO3- que ingresa cloro y expulsa bicarbonato. Además, el antiporte Na+/K+, que expulsa de la misma célula al H+, es un mecanismo para neutralizar la acidosis provocada por el anterior mecanismo. Ahora el mismo Na+ que ingresó sal e de la célula por la bomba Na+/K+ ATP asa basolateral. La bomba H+/K+ luminal puede ser inhibida por antiácidos muy recientes como el omeprazol, que se administra cada 24 hs, teniendo en cuenta que ese lapso coincide con el tiempo de renovaci ón de la bomba en la membrana luminal de la célula parietal. Otros antiácidos inhiben al rece ptor H2 (de histamina) como es el caso de la ranitidina. De todas maneras es mucho más efectiva la inhibición de la producción de ácido clorhídrico por parte del omeprazol (inhibidor de la bomba de protones) ya que la inhibición es de manera irreversible, por lo tanto el efecto es mucho más potente. Regulación de la secreción de ácido clorhídrico En el período interdigestivo, existe una secreción basal de HCL, que confiere a la luz estomacal un pH de entre 0,8 y 1,8, que se produce por el estímulo basal neurohormonal que parte del SNE, que mantiene activadas a las células parietales. Se produce un aumento progresivo de la secreción, que no es tónica, relacionado con el CMM, que en este caso, actúa como CMS (complejo motor secretor). Durante el período interdigestivo se suceden tres fases: cefálica, gástrica y finalmente intestinal: FASE CEFÁLICA: es controlada por reflejos condicionados en base a los diferentes estímulos (vista, olfato, audición), es mediada por el nervio vago (par X) que estimula a l as células parietales mediante fibras descendentes del SNE. FASE GÁSTRICA: basalmente el pH es de entre 0,8 y 1,8, como antes se mencionó. Las proteínas de los alimentos deglutidos, actúan a nivel gástrico y hacen que el pH se ele ve a 5 (dependiendo de la cantidad de aminoácidos básicos), estimulando luminalmente la secreción de HCL por estímulo sobre la célula G (que libera gastrina) e inhibiendo a la célula D antral (que ahora deja de liberar SST, que antes se encontraba inhibiendo la secreción de HCL). Otros estímulos son, además de los aminoácidos aromáticos de las proteínas de la dieta, la distensión mecánica del antro y el estímulo del Ca2+ de la dieta. FASE INTESTINAL: esta es la fase inhibitoria por excelencia de la secreción de HCL. Los estímulos luminales en el intestino proximal son: el bajo PH del quimo que viene del estómago (es decir, el bolo alimenticio parcialmente digerido y embebido en HCL) y las proteínas, hidratos de carbono y lípidos que forman parte del mismo. Estos estímulos luminales activan a las células I (que liberan CCK) y a las células S (que liberan se cre tina), que descargan a la circulación sus respectivas neurohormonas. Las mismas (CCK y secretina), producen el denominado efecto entero-gastrona, sobre las células D fúndi cas gástricas, las cuales son cerradas (sólo son estimuladas basolateralmente a través de neurohormonas, ya que no contactan con la luz), y liberan SST. Esta última frena la secreciónd e HCL al inhibir a las células G, que se ven impedidas en liberar gastri na (para estimular a las células parietales), y además provoca un retardo del vaciamiento gástri co por incoordinación antro-píloro-duodenal (por un desacople eléctrico), proporcional al volumen de carga y al grado de composición lipídica del bolo. Tipos celulares y sus productos: MASTOCITOS: histamina ELC (enterocromafin like cells): histamina PARIETALES: HCl y factor intrínseco D: somatostatina (SST) G: gastrina I: colecistokinina (CCK) S: secretina Respuesta potenciada: se presenta en la fase cefálica, y se produce entre las células G, P y ECL y las terminaciones nerviosas del SNE (a través de neurohormona bombesina-simil). Cada tipo celular cuenta con receptores para los productos que no fabrica, de manera tal que se produce una estimulación múltiple, dando lugar a una respuesta potenciada. Test de histamina El objetivo del test de histamina es determinar la masa funcional de células parietales, determinando la capacidad máxima de secreción de HCL, y también la capacidad de secreción basal. Se supone que el número de células parietales (masa funcional) es proporcional al peso corporal magro, mayor en el hombre que en la mujer. Para la prueba, en primer lugar se realiza un vaciamiento gástrico que no debe superar los 200ml y no presentar restos de alimentos. La sonda se mantiene durante una hora para, entonces, volver a retirar el contenido gástrico. Es entonces cuando se mida la BAO (muestra basal de HCL)que normalmente es de 0-4 meq/hora. Se espera una hora más para administrar entonces un inhibidor de receptores H1 (para e vi tar un shock anafiláctico) y así entonces se puede administrar histamina que sólo estimulará a los receptores H2 (que no se encuentran inhibidos), y que están involucrados en la secreción de HCL. La histamina puede administrarse por perfusión constante o en una única administración (en el caso de optar por el test de histamina máxima). Se suelen administrar 0,1mg de clorhidrato de histamina por kg de peso magro, como medida standard. A 0,4 mg se logra activar el total de las células parietales y entonces se determina la MAO (máximo débito ácido) que suele ser de entre 10 y 24 meq/hora. En el test de perfusión constante hay un aumento progresivo de la secreción ácida en relación a l a variable concentración de receptores para histamina (sensibilidad particular para cada una de cada una de las células), hasta que en un punto, generalmente al administrar 0,4mg de hi stami na se logran activar todos los receptores H2 y la curva se estabiliza. Respuesta normal y casos patológicos del Test de Histamina Grado NORMAL I II III MAO 10-24 2 0,50 0 BAO 0-4 25 5 0,5 Secreción NORMAL AUMENTADA DISMINUIDA DISMINUIDA Patología probable NINGUNA Síndrome ulceroso Atrofia parcial de las células parietales Atrofia total de las células parietales con anemia IV 48 68 AUMENTADA perniciosa Gastrinoma (tumor secretor Barrera defensiva mucosa La barrera mucosa es una compleja estructura de defensa contra el pH ácido (cuyo origen es el HCL), antígenos, sales biliares, y enzimas líticas que pudieran lesionar el epitelio mucoso de l tubo digestivo a nivel estomacal y del intestino proximal (duodeno-yeyuno). Esta barrera se di vi de e n tres sectores: pre-epitelial, epitelial y post-epitelial 1. PRE-EPITELIAL: formada, desde el sector más externo al más interno, por la capa de agua no removible, la secreción alcalina, la capa de moco y finalmente el factor surfactante: CAPA DE AGUA NO REMOVIBLE: es la primera en contacto con la luz. SECRECIÓN ALCALINA: proviene de la secreción epitelial de bicarbonato, el que queda retenido en la capa de agua, con la finalidad de neutralizar los protones que podrían retrodifundir desde la luz. Un 70% del bicarbonato se secreta por antiporte con cloro (que ingresa al enterocito), un 20% sale por vía paracelular, mientras que el 10% restante se secreta por canales luminales electrogénicos. En el último caso, el HCO3- proviene del simporte (co-transporte) HCO3/Na+ de la submucosa (mecanismo estimulado por las prostaglandinas) CAPA DE MOCO: el moco está formado por una porción central (de naturaleza proteica) envuelta por hidratos de carbono, con el fin de hidratarlo y protegerlo contra enzimas proteolíticas. Existen dos tipos de moco a este nivel, dependiendo del sector donde se produce. El más superficial (producido por células más externas de la glándula), es el moco no sulfatado, que es más diluído por carecer de gran cantidad de pue nte s disulfuro, pero muy rico en NANA (ácido N acetil neramínico, PAS +) con l a capacidad de neutralizar H+ mediante sus cargas negativas libres. Por el contrario, el moco sulfatado, es más profundo (producido por células profundas del cuello glandular), y líquido, con mayor cantidad de pue nte s disulfuro, pobre en NANA, es producido en grandes cantidades y a gran velocidad generando una corriente hacia la luz (barriendo elementos que podrían llegar al fondo de la glándula gástrica, y favoreciendo la sal i da de los H+ secretados por las células parietales). La capa de moco es mayor e n dudodeno, donde también es importante la producción de HCO3-, mientras que en el estómago no tienen gran relevancia, como sí la tiene l a capa de surfactante; como diferencia entre la barrera mucosa gástrica y l a duodenal hay que tener en cuenta que una mayor secreción de bicarbonato a nivel gástrico interferiría en gran medida con la secreción de H+. que es característica de este sector. La capa de moco actuaría como un soporte para la secreción alcalina, al concentrarse a este nivel, se genera un gradiente tal que del lado luminal de la capa, el pH es ácido, mientras que en el lado epitelial el pH es neutro. FACTOR SURFACTANTE: forma una capa de glicoesfingolipidos (molécul as anfipáticas formadas por un ácido graso, y un esfingol con colas hidrófobas y un hidrato de carbono como cabeza hidrofílica), constituyendo una monocapa: las cabezas hidrofílicas interactúan con la membrana, mientras que las colas hidrófobicas se orientan hacia la luz impidiendo la formaci ón de HCL, ya que evitan la hidratación de los H+. En la membrana de la célula parietal, este factor especialmente, resulta indispensable. 2. EPITELIAL: barrera formada por las mismas células epiteliales. El H+ tiende a ingresar al medio interno por vía paracelular o por vía trans-celular. En el primer caso, la célula podría entrar en acidosis y destruirse de no contar con un mecanismo como el antiporte Na+/H+. Entonces el H+ que había retrodifundido es eliminado del interior celular y la célula puede conservar su equilibrio ácido-base. Los mecanismos defensivos a nivel epitelial i ncluyen: uniones GAP intercelulares, renovación celular por mitosis (a partir de las células basal e s de Paneth) cada 72 hs. También puede haber restitución celular rápida en 24 hs con reemplazo por células vecinas que migran en dirección de un tapón mucoso en el sitio lesionado, además participan de este proceso el cortisol y el EGF-1 (factor de crecimi e nto epidérmico). Este último mecanismo sólo es posible si la membrana basal y la microcirculación están indemnes. 3. POST-EPITELIAL: formada por los capilares y los vasos de la submucosa (microcirculación), que aportan glucosa y O2 para poder mantener todo el resto del sistema, HCO3- para neutralizar los H+ retrodifundidos hasta este nivel y barrerlos (house keeping). Este e s e l sector más importante de toda la barrera, teniendo en cuenta que su falla haría fracasar todo el sistema por falta de O2, nutrientes y buffer bicarbonato. El HCO3- puede prove ni r de la circulación sistémica, pero también puede tener origen local: en glándulas oxínti cas (en estómago, productoras de HCL), la secreción de H+ por la célula parietal hace que se acumule bicarbonato dentro de la célula, que va a ser liberado a la circulación por el antiporte Cl-/HCO3- (basolateral). Este mismo difunde vía circulatoria hasta la parte alta de la misma glándula, donde es recaptado por células del epitelio superficial, el cual luego lo libera a la luz, contribuyendo a la carga de HCO3-, a nivel de la capa de moco; mediante este mecanismo se define la “marea alcalina”. PARTE 3: REGULACIÓN DE LA FUNCIÓN DIGESTIVA Homeostasis La homeostasis es el equilibrio del medio interno, mantenido de manera natural mediante respuestas adaptativas que promueven la conservación de ciertas variables consideradas normales (por ejemplo, la temperatura normal). Diversos mecanismos sensoriales de retroalimentación y control actúan para mantener este estado constante. Algunas de las funciones o variables controladas por mecanismos homeostáticos son la secreción glandular, el equilibrio hidroelectrolítico, la temperatura corporal, la presión arterial y la hematopoyesis. Cada vez que actúe un factor del medio externo alterando el estado de equilibrio de l organi smo (la homeostasis), será necesario un cambio de conducta (un cambio de programa) ante la nueva situación, que tenderá a mantener estable el equilibrio del medio interno con respe cto al me di o externo. A su vez, pueden distinguirse dos tipos de conductas (programas) que tienden a mantener la homeostasis: CONDUCTAS REACTIVAS: consiste en una reacción del medio interno frente a determinado estímulo del medio externo, mediante un mecanismo de retroalimentación (feedback), por lo que se denominan respuestas a lazo cerrado. Este cambio de conducta dura tanto como dure el estímulo estresante. CONDUCTAS PREDICTIVAS: son programas determinados (como el sueño o la conducta de preservación de la especie en los animales) que se ejecutan con o sin estímulo externo (por ejemplo, el sueño carece de estímulo bien definido). Estas conductas (programas) pueden encontrarse reforzados por conductas reactivas, las que producen las adaptaciones necesarias para alcanzar el objetivo predeterminado (en este caso). Las conductas predictivas se basan en cronorritmos o ritmos biológicos (circadianos, ultradianos, circanuales), por lo que se las denomina respuestas a lazo abierto (al care ce r de un mecanismo de retroalimentación), y se caracteriza por ponerse en marcha y no detenerse hasta haberse completado el programa, independientemente de la presenci a o no de un estímulo. Sistemas y programas de trabajo Un sistema es un conjunto de entidades organizadas (subsistemas) que se encuentran regidos por leyes o reglas comunes, que tienden a cumplir con un objetivo que sería imposible de lograr individualmente. Los distintos sistemas son los que ejecutan los diferentes programas de trabajo. Por otra parte, un programa es un conjunto de funciones organizadas, con el fin de cumplir un objetivo determinado (por ejemplo, la secreción de HCl). Dentro del concepto de sistema, se desprende el concepto de sistema integrativo, un caso especial que es necesario tener presente al tratar la fisiología del aparato digestivo. Un sistema integrativo es un sistema que se caracteriza por: Recibir la información proveniente de receptores periféricos (como los corpúsculos de Pommerenke, que son mecanorreceptores), a través de una rama aferente. Analizar la información en un Centro Integrador (que en general es el SNE, salvo en los casos de las funciones controladas por el SNC: deglución, vómito, defecación) y generar una respuesta (ejecutando un programa predeterminado). Transmitir la información desde el Centro Integrador hasta los efectores (por e jempl o, e l músculo liso del tubo digestivo), mediante una rama eferente. Ejecutar la acción propiamente dicha, por parte de los efectores (por ejemplo, la contracción del músculo liso para producir la peristalsis). Niveles de organización del control neural Primero es necesario diferenciar dos conceptos: 1. NIVEL DE PROCESAMIENTO: funcionalmente constituye un arco reflejo, es decir que es capaz de recibir información, procesarla, elaborar una respuesta y transmitirla a los efectores. Los programas se procesan en sistemas integrativos. 2. NIVEL DE MODULACIÓN: la respuesta que elabora sólo puede modular (acelerar, retrasar o inhibir) la respuesta del nivel anterior Una vez esclarecido estos dos conceptos diferentes, se detalla cada nivel de organización: NIVEL I: comprende las actividades del plexo mientérico y del plexo submucoso, ambos partes de l SNE. El submucoso procesa programas relacionados con la secreción y la absorción; el mienté rico (de mayor jerarquía), procesa programas relacionados con la motilidad y además modula al submucoso. Se encuentran en estrecha relación con los efectores. Este nivel procesa la mayoría de los programas del tubo digestivo como: CMM, ritmo secretorio, reflejo secretor intestinal. NIVEL II: comprende básicamente los ganglios PRE y PARAVERTEBRALES de la cadena simpática. Recibe señales aferentes de receptores periféricos y elabora una respuesta que es transmi tida al NIVEL I que procesa la función o programa (conjunto de funciones) y ejecuta una respuesta: Ejemplo de ellos son los reflejos gastro-colónicos, duodeno-pancreáticos, recto-colónicos, y anocolónicos. Este nivel actúa como una estación intermedia para la información que llega desde la periferia para luego enviar señales eferentes al NIVEL I. NIVEL III: comprende desde la médula espinal hasta la corteza cerebral. El área más importante e s la región bulbo-protuberancial donde se encuentran los centros sensitivos y motores de los nervios craneales que inervan al tubo digestivo: V, VII, IX, Y X. Es el nivel de modulación por excelencia, por ejemplo, la fase cefálica de la secreción de HCl: en este nivel no se inicia ni se genera sino que se modula. Además este nivel procesa tres programas: deglución, vómito y defecación. Péptidos reguladores GASTRINA: Es un péptido derivado de la familia de la CCK, pero con efectos (en su mayoría) antagónicos. Producida por las células G antrales y duodenales. Las isoformas más importantes son la 68 (big-big gastrina), la 34 (big gastrina)y la 17 (Little gastrina). Los segundos mensajeros que utiliza son: PIP2 y Ca2+ (así que su receptor debe se r de 7 TMS acoplado a proteína G q) Estímulos: Pueden ser de dos tipos A) BASOLATERALES: el péptido bombesina-símil (del SNE) y la histamina, estimulan su secreción, por otro lado, la SST (de las células D) ejerce efecto inhibitorio B) LUMINALES: el aumento del pH (hasta 5), los aminoácidos aromáticos, y el Ca2+ estimulan su secreción. La disminución del pH (hasta 1-2) la inhibe. Efectos: A) Estimulación de las células parietales sobre receptores CCK-B. B) Efecto trófico sobre la mucosa gástrica (acción que comparte con la CCK). C) Modulación de la contracción del EEI. D) Leve efecto ecbólico (liberación enzimática) a nivel del páncreas; leve efecto increti na y estímulo de la liberación de polipéptido pancreático (PP) COLECISTOKININA (CCK): Las isoformas más importantes son la 33 (en tubo digesti vo) y 8 (a nivel neural), con acción similar pero diferente vida media (mayor para la isoforma 8 neural). La CCK es producida, en tubo digestivo, por las células I duodeno-yeyunal es. Los segundos mensajeros que utiliza son: PIP2 y Ca2+ Estímulos: lípidos, proteínas, principalmente hidrolizados, actuando a nivel luminal. Efectos: son diferentes de acuerdo al sitio de acción A) SOBRE EL PÁNCREAS: ejerce un efecto ecbólico al estimular al acino para la secre ci ón proteica, mientras que la Ach (por un mecanismo neural) lo favorece. Además tiene un efecto trófico y es el estímulo principal para la liberación de PP. B) SOBRE LA VÍA BILIAR: ejerce un efecto colagogo (estimula la contracción), el que consiste en la contracción de la pared vesicular e indirectamente (por un circuito en e l que intervienen encefalinas) relajación del esfínter de Oddi. C) SOBRE EL ESTÓMAGO: ejerce un efecto entero-gastrona, el que media la inhibición de la secreción de HCl y el retraso del vaciamiento gástrico. Además tiene un efecto trófico sobre la mucosa gástrica y estimula la liberación de bicarbonato. D) SOBRE EL INTESTINO DELGADO: inhibe la absorción de agua y electrolitos, además de favorecerla liberación de bicarbonato, al igual que en el estómago. E) SOBRE EL COLON: estimula indirectamente su motilidad al actuar sobre el SNE, que al liberar Ach contrae el músculo liso a este nivel. SECRETINA: No se registran isoformas, pertenece a la familia del VIP (péptido intestinal vasoactivo). Se libera en forma endócrina. El segundo mensajero que utiliza es el AMPc Estímulos: descenso del pH a nivel del yeyuno íleon, además de las sales biliares, si e mpre a nivel luminal. Efectos: son diferentes de acuerdo al sitio de acción A) SOBRE EL DÚCTULO PANCREÁTICO: estimula la secreción de agua y electrolitos, favoreciendo por vía neural el efecto colerético del VIP. B) SOBRE DÚCTULO BILIAR: tiene un efecto colerético similar. C) A NIVEL GÁSTRICO: es parte del efecto entero-gastrona junto con la CCK. PEPTIDO INSULINO-TROPICO GLUCOSA DEPENDIENTE (GIP): Antiguamente denominado péptido inhibidor gástrico. Es producido por la célula K duodeno-yeyunal, que es una célula cerrada (sólo recibe estímulo basolateral, sin contacto con la luz). Estímulos: a nivel basolateral, al ser superado en un 20% el valor basal de glucosa (que normalmente es de 70-110 mg%). Efectos: son diferentes de acuerdo al sitio de acción A) SOBRE EL PÁNCREAS ENDÓCRINO: favorece la liberación de insulina por las células beta del páncreas mediante un efecto incretina. B) A NIVEL GÁSTRICO: inhibe todas las funciones gástricas. Clasificación de los péptidos reguladores según familia Familia dela Gastrina: Gastrina y CCK Familia de la Secretina: Secretina, VIP, GIP, Entero-glucagon Reflejo secretor intestinal (RSI) El reflejo secretor intestinal es parte de lo que se denomina un sistema integrativo. Un sistema e s un conjunto de entes que interactúan de manera sinérgica y coordinada en pos de una finalidad común, funcionando bajo los mismos principios. En especial, los sistemas integrativos se caracterizan por estar formados por un polo aferente en relación con el estímulo, un centro integrador y un polo eferente en relación con el efector. En el caso del RSI, el receptor es una célula endócrina, el centro integrador es el SNE, el efector e s el enterocito y el efecto final es la secreción de agua y electrolitos. Por su parte, tanto la vía aferente como la eferente, están representadas por vías peptidérgicas o por vías colinérgicas. 1) AFERENCIA: el estímulo es de origen mecánico (por la distensión provocada por el bolo alimenticio). Los receptores para el estímulo no forman parte de los denominados “receptores clásicos”, ya que en este caso se trata de células endócrinas como las NT (productoras de neurotensina), las enterocromafines (productoras de serotonina o 5-HT), las células S, D y G. Los plexos nerviosos y los plexos capilares (microcirculación), llegan hasta el sector basolateral de estas células, con lo cual una parte de las neurohormonas (según corresponda para cada tipo celular) pasa a la circulación y la otra parte estimula a los plexos nerviosos. Estas células endócrinas (en este caso con función de receptores), liberan, luego de ser estimuladas, sus productos (5-HT, neurotensina, secretina, etc.), l os que actúan sobre los terminales de neuronas internunciales del plexo mientérico, las que a su vez liberan neurotransmisores: sustancia P, 5-HT, y Ach, los que actúan sobre las neuronas integradas en las estaciones ganglionares del SNE (el centro integrador). El resto de los productos de liberación, como ya se comentó, pasa directamente a la circulación. 2) CENTRO INTEGRADOR: la señal se integra a nivel del plexo mientérico (parte del SNE), a nivel de las estaciones ganglionares correspondientes. A este nivel se produce la estimulación mediante los neurotransmisores sustancia P, 5-HT y Ach. Por otro lado, puede haber inhibición, esta vez mediante SST y encefalinas, que provienen de otros tipos celulares. 3) EFERENCIA: desde el plexo mientérico se libera VIP (segundo mensajero: AMPc), y Ach (segundo mensajero: PIP2-Ca2+), que estimulan la liberación hidro-electrolítica por parte del enterocito. A su vez, este último puede ser inhibido en su función secretora medi ante adrenalina (origen en SNA), actuando a través de receptores alfa-2 (disminuyen los niveles de AMPc al estar acoplados a proteína G i). Todo esto representa la vía neuronal. Simultáneamente por vías endócrinas (a través de la circulación) llegan al enterocito las neurohormonas liberadas originalmente a la circulación por las células endócrinas que actuaban como receptores, como es el caso de la neurotensina (NT), serotonina (5-HT), y secretina. Se considera que la vía neural es más rápida, aunque más fugaz que la endócrina. Sensorialidad del tubo digestivo Antiguamente se pensaba que las vísceras eran insensibles a los estímulos dolorosos y que carecían de terminales nerviosos. Hoy se sabe que las vísceras huecas poseen terminaciones nerviosas y que estímulos adecuados pueden inducir dolor. También se sabe que en el lóbulo de la ínsula se configuran tanto el homúnculo sensorial como el homúnculo motor del aparato digestivo. El principal estímulo doloroso para una víscera hueca es su distensión, contracción, o estiramiento. Además, pueden provocar dolor la distensión de la cápsula de los órganos sólidos como el hígado, la inflamación del peritoneo (su hoja parietal que está inervada, a diferencia de la visceral que no está inervada) adyacente a una víscera y la isquemia (falta de perfusión sanguínea) de algún sector del tubo digestivo por oclusión vascular. El mecanismo de dolor visceral no está totalmente claro, sin embargo se conocen algunos he chos de gran utilidad para la clínica, sobretodo. El dolor visceral tiene marcadas diferencias con respecto del dolor somático (como el dolor que se origina en el músculo estriado o en la pi e l , por ejemplo). Por empezar, la sensibilidad somática tiene una correlación 1 a 1 entre dermatoma (sector definido inervado) y metámera (sector medular hacia donde va la inervación sensorial). Mientras que en la sensibilidad visceral un solo dermatoma puede encontrarse relacionado con hasta cinco metámeras, que a su vez se encuentran relacionadas con cinco sendos dermatomas somáticos. Además el dolor somático es bien puntual (el paciente puede indicarlo con un de do), mientras que el dolor visceral es difuso (el paciente lo refiere indicando con toda una mano). Esto último es la base del dolor referido, es decir que un estímulo doloroso en un sector de una víscera hueca (dermatoma visceral) se transmite divergentemente hacia cinco metámeras medulares, conectadas a su vez con cinco sectores de la piel o del músculo estriado superficial (dermatomas somáticos) dando la sensación consciente de que el dolor se originó en estos últimos. Entonces, de acuerdo con su mecanismo, se diferencian dos tipos de dolor abdominal: el dolor visceral verdadero y el dolor referido. DOLOR VISCERAL VERDADERO: es iniciado por un estímulo que actúa en terminales nerviosos de la víscera hueca, generalmente por distensión o contracción de la mi sma. El estímulo es conducido a la médula mediante fibras aferentes viscerales, sin la participación de fibras somáticas (cerebro espinales). Este dolor es difuso y no se asoci a a defensa muscular (contracción al mínimo tacto de la pared abdominal) ni a hiperalgesia cutánea. DOLOR REFERIDO: de acuerdo a las características antedichas, puede ser víscero-sensitivo o víscero-cutáneo. EL dolor referido víscero-sensitivo es producido en especial por la inflamación de la víscera. Es conducido por fibras aferentes viscerales en relación con fibras somáticas (cerebro espinales) como se describió antes. Por esta razón, el dolor es referido a las zonas laterales del abdomen y está asociado con la defensa muscular y la hiperalgesia cutánea. Por su parte el dolor referido víscero-cutáneo se produce cuando hay inflamación del tejido subperitoneal, peritoneal y mesentérico. Es un dolor bien localizado (puntual) y se acompaña de rigidez permanente de la pared abdominal en el sitio exacto de ubicación de la víscera inflamada. PARTE 4: FISIOLOGÍA HEPATO BILIAR El hígado es un órgano constituido por células con gran capacidad proliferativa, que trabaja normalmente al 20% de su capacidad máxima. Como glándula exócrina, su principal producto de secreción es la bilis: líquido isotónico, alcalino y pigmentado. La composición biliar es: AGUA: en un 90% ELECTROLITOS: los principales cationes son Cu2+ y Fe3+, mientras que el anión principal es el HCO3-. El resto de los iones se encuentran en concentraciones similares a las del plasma. PROTEINAS: la principal es la albúmina. Además hay un contenido importante de IgA secretora (dimérica). ELEMENTOS LIPOSOLUBLES: colesterol (en una concentración de 6mM), fosfolípidos (en una concentración de 20mM, del que el 98% corresponde a lecitina, también denominada fosfatidilcolina), sales biliares y bilirrubina (presente en su forma conjugada). Sales biliares Su producción hepática diaria representa entre el 2-6% del total y es de entre 0,2 – 0,6 gramos. El pool de ácidos biliares en el organismo es de 2-4 g (en concentración de 5mM). Su síntesis es a partir del colesterol (esta es una de las formas de eliminación del mismo). La absorción de sales biliares, que además sufren circuito entero hepático, es de un 80-90% a nivel del íleon terminal. La enzima clave en la producción de sales biliares es la 7-alfa-hidroxilasa (o 7-alfamonooxigenasa), enzima regulable asociada al citocromo p450 que tiene como producto al áci do biliar primario (ácido cólico y quenodesoxicólico), los mismos pasan a sales biliares al conjugarse con taurina o glicina, además de Na (OH) o hidróxido de sodio. ÁCIDO CÓLICO QUENODESOXICÓLICO (con 3 grupos –OH) ÁCIDO (con 2 grupos –OH) Glicina O Taurina Na (OH) SAL BILIAR SAL BILIAR Posibles sales biliares a partir del ácido cólico: taurocolato de sodio, glicolato de sodio. Posibles sales biliares a partir del ácido quenodesoxicólico: tauroquenodesoxicolato de sodio, glicoquenodesoxicolato de sodio. Las sales biliares, producidas en el hepatocito, se vierten al canalículo biliar, mediante un carri e r ATP-dependiente y por el carrier MOAT, a una presión de 30 mm de Hg, generándose una diferencia de presiones (delta P) de 10 mm de Hg, favorable para el eflujo. El canalículo biliar se encuentra formado por las paredes celulares de los hepatocitos. Ya en el canalículo biliar la bilis se isotoniza. Se absorben ácidos biliares en forma activa y se produce la colangio cetosis (absorción de glucosa), que tiende a evitar la proliferación de poblaciones bacterianas que realizan glucólisis. De no mediarse este mecanismo de colangio cetosis (o en caso de falla del CMM), habría sobrecrecimiento bacteriano de colonias que hacen glucólisis anaeróbica (dejando como residuo al ácido láctico), provocando la acidificación del medio, que puede tener como consecuencia la desconjugación de las sales biliares. Es decir, la baja concentración de glucosa mantiene la esterilidad de la vía biliar. Además hay absorción de H+ y secreción de HCO3- (por estímulo de la secretina) que alcaliniza la bilis. Finalmente, las sales biliares se desconjugan en intestino delgado cuando el pH del medio desciende por la llegada del quimo ácido proveniente del estómago. Vesícula biliar Su función es concentrar y almacenar la bilis (una función NO indispensable para la vida). Ade más se encarga de acidificar la bilis al absorber HCO3-, mediante el antiporte (contratransporte) Na+/H+, es decir que se produce absorción de HCO3- por protonización previa (estímulo para este mecanismo). En caso de colecistectomía (resección quirúrgica de la vesícula biliar), la bilis se almacena en intestino proximal y se libera frente a estímulos determinados. Metabolismo de las sales biliares En el íleon terminal, un 80% de las sales biliares (producidas por el hepatocito y almacenadas por la vesícula biliar), se reabsorben como tal, mientras que otro 10% se reabsorbe como ácidos biliares. A este nivel, la absorción se realiza mediante un cotransporte con Na+ (simporte) denominado IBAT. La absorción de los ácidos biliares es favorecida por la protonización (disminuyendo las cargas libres, pasando a ser más liposolubles) que sufren antes de contactar con la membrana del enterocito. Este microclima ácido que circunda la membrana del enterocito se logra mediante una bomba H+/K+, que secreta H+, que a su vez se conjuga con los ácidos biliare s. El restante 10% (el que quedaba, recordar que del 100% el 80% se reabsorbía como sal biliar y el 10% como ácido biliar, así que quedaba un 10% remanente sin describir), pasa al colon, donde l as bacterias producen deshidroxilación de los ácidos biliares primarios, que pasan a ser secundarios. El pool de ácidos biliares es de 2-4 gramos (en una concentración de 5mM) y se movili za e ntre 612 veces por día, mediante el circuito enterohepático. Funciones de las sales biliares 1) Solubilizan el colesterol y otros lípidos, permitiendo la excreción del primero (colesterol) 2) Facilitan la absorción intestinal de los lípidos 3) Regulan negativamente su propia síntesis 4) Facilitan la absorción de vitaminas liposolubles (K, E, D, A) La circulación del líquido biliar (que contiene, entre otros, a las sales biliares), se produce gracias a “bombas químicas”, constituidas por las membranas del hepatocito y de los enterocitos a nivel del íleon terminal, y “bombas mecánicas”, constituidas por la motilidad de la vesícula biliar y la motilidad intestinal. Estas bombas median tanto la absorción como la secreción de la bilis. Dentro de la bilis se destacan tres fracciones: 1) Fracción ácido biliar dependiente: origen en el hepatocito, proviene de la recaptación biliar por el mecanismo IBAT, y posterior liberación pasiva. 2) Fracción ácido biliar no dependiente: origen en el hepatocito, junto con la fracci ón áci do biliar dependiente conforman el 80% de la bilis. Consiste en la secreción de HCO3-. 3) Fracción ductular: origen en conductos biliares menores, representa el 20% de la bilis. Consiste en la liberación de HCO3-. Metabolismo de la bilirrubina El 75% de la bilirrubina es producto de la metabolización del grupo hemo de la Hemoglobina (Hb), mientras que el resto proviene de la degradación del grupo hemo de la míoglobina y de los citocromos. La vida media de un eritrocito es de 120 días: cuando los lípidos de su membrana se oxidan cada vez más y la van haciendo cada vez menos flexible, ya no pueden atravesar las re de s e spl éni cas, donde van a quedan atrapados para luego ser degradados por los macrófagos (esplénicos), liberando Hb, la que se degrada a su vez en globina y grupo hemo. El hemo mediante una enzima hemooxigenasa, pasa a ser biliverdina, la que por una enzima reductasa pasa a ser bilirrubina (indirecta, aún no conjugada en el hígado, liposoluble). Esta bilirrubina no conjugada (BNC) es poco soluble y resulta tóxica, debida a su falta de solubilidad en el plasma, debe ser transportada por una proteína, la albúmina. La BNC, cuando se une a la albúmina, no puede filtrar a través de la barrera de filtración glomerular (no se filtra a nivel renal), esto se debe a que el glomérulo renal es imperme abl e a l as moléculas de gran tamaño y peso, como lo es la albúmina. Además, hay restricción del filtrado glomerular dependiente de cargas: la membrana basal glomerular (con alta densidad de proteoglicanos con cargas negativas libres) tiene afinidad por moléculas catiónicas, y por el contrario, repele moléculas aniónicas, como es la albúmina. Por otro lado, la escasa cantidad de albúmina (en este caso tiene unida BNC) que podría llegar a filtrarse, se reabsorbe inmediatamente. La bilirrubina no conjugada (unida a la albúmina, o bilirrubina indirecta), es captada por receptores específicos en el polo capilar del hepatocito. Los receptores se encuentran acoplados a un transportador denominado OATP (organic anions transport protein), transportador de aniones orgánicos que gasta un mol de ATP por mol de BNC ingresada al hepatocito por el polo capilar o captador. Ya dentro del hepatocito, la BNC se une a las ligandinas Z e Y, con la finalidad de evitar que la BNC retrodifunda: estas ligandinas separan la BNC de la albúmina al presentar mayor afinidad que esta última por la BNC. Así una vez que se unen las ligandinas a la BNC suceden dos cosas: por un l ado la albúmina vuelve a la circulación y por el otro la BNC es transportada hasta el REL del denominado polo conjugador, donde la enzima UDP-GLUCURONIL TRANSFERASA (UDP-G) conjuga dos veces a la bilirrubina: en el 80% de los casos con ácido glucurónico y el restante con sulfatos y fosfatos. Ahora la bilirrubina no conjugada (BNC) pasa a denominarse bilirrubina conjugada (BC) o directa. La BC es soluble, predomina en bilis, sí puede filtrar a nivel renal y en exceso produce coluria (color oscuro y de aspecto espumoso de la orina). Sin embargo es conveniente no confundir e ste col or patológico del color normal (ámbar) de la orina, que es consecuencia de la presencia de urocromos (producto del catabolismo de aminoácidos cíclicos como la tirosina y la fenil-alanina). La BC es liberada activamente mediante una bomba que gasta un mol de ATP por cada mol de BC liberada, y se encuentra en el polo canalicular o secretor del hepatocito (que da hacia la vía biliar). Existe una diferencia de presiones (∆p) de 10 mm de Hg entre el hepatocito y el canalícul o bi l i ar, favorable para la salida de la BC hacia la vía biliar. Mediante la misma, la BC pasa a la segunda porción del duodeno mediante el conducto colédoco, cruza todo el intestino delgado y finalmente llega al colon derecho. En colon derecho, las bacterias fermentativas, mediante enzimas beta-glucuronidasas, forman un núcleo de bilirrubina libre (al desconjugarla). Esta bilirrubina libre, también por acción bacteriana, pasa a estercobilinógeno (ECB). Un 20 % del ECB (del 20% que se reabsorbe en el íleon distal, pasando a la circulación), se filtra a nivel renal, para luego liberarse en la orina, donde se denomina urobilinógeno. Durante la micción, en contacto con el O2 del aire, el urobi l i nógeno se oxida a urobilina, que NO da el color característico de la orina (ese lo daban los UROCROMOS! VER MÁS ARRIBA). El restante 80% del ECB (siempre del 20% que se reabsorbe en íleon distal), realiza circulación enterohepática, para reconjugarse a nivel hepático. El restante 80% del núcleo de bilirrubina libre (que no se reabsorbió en íleon distal, y que era producto de la acción bacteriana sobre el colon derecho) continúa como ECB (estercobili nógeno) para excretarse en materia fecal, donde pasa a ser estercobilina por el contacto con el O2 del ai re ambiental (por oxidación), dando el color amarronado característico de la materia fecal (MF). En resumen, la BC continua las siguientes vías: BC (100%) 80% Se excreta en MF como estercobilina 20% se reabsorbe en íleon distal, pasando a la circulación 80% realiza circulación enterohepática (reconjugación) 20% se filtra en el riñón y se transforma en urobilina en la orina Ictericia La ictericia es un signo clínico (objetivo) que se caracteriza por la coloración amarillenta de la pi e l , mucosas y escleróticas. La hipervitaminosis A (dando un cuadro denominado pseudoictericia) provoca una coloración similar, pero no en escleróticas, lo cual sirve para el diagnóstico diferenciación una ictericia. La ictericia suele producirse cuando los niveles de bilirrubina supe ran los 2 mg%. Los parámetros clínicos que pueden variar de manera importante según el paciente, son: BILIRRUBINEMIA NORMAL: concentración de bilirrubina de 0,8-1,2 mg%. Normalmente el 80% corresponde a BNC y el restante 20%a la BC. HIPERBILIRRUBINEMIA SUBCLÍNICA: que no provoca ictericia; concentración de bilirrubina de 1,2-2 mg%. HIPERBILIRRUBINEMIA CLÍNICA: se observa ictericia; concentración de bilirrubina mayor a 2 mg%. Las ictericias pueden clasificarse, según su origen, en prehépatica, hepática y posthepática. La mayoría de las noxas habituales (hepatitis viral, por ej.) afectan, en mayor o menor medida, a cada uno de los tres polos hepáticos (captador, conjugador, excretor), pero el excretor suele resultar frecuentemente afectado en primer lugar, por razones aún no esclarecidas. ICTERICIA PRE-HEPÁTICA: se encuentra afectada la zona comprendida entre la vía sanguínea y el polo capilar (o captador) del hepatocito. CAUSAS POSIBLES: hemólisis acelerada (como en la eritroblastosis fetal), ane mia hemolítica o estasis sanguínea. En todos los casos, el hígado está sano. SIGNOS: gran aumento de la BNC (unida a la albúmina) en la sangre, con aumento de la BC en la bilis (pero menor, teniendo en cuenta al polo conjugador del hepatocito como factor limitante). En sangre también hay un leve aume nto de l a BC, en este caso por conjugación extrahepática (a nivel del bazo, por ejemplo). L a orina es normal (aumenta la urobilina, que NO produce coluria), pero las heces son hipercólicas (más oscuras) por aumento de la estercobilina. ICTERICIA HEPÁTICA: se encuentran afectados los tres polos (captador, conjugador y excretor), es decir, todo el hepatocito, por lo que también puede denominarse ictericia mixta. Este tipo de ictericias es la más frecuente y se caracteriza por el aumento tanto e n los niveles de BNC como de BC, ya que resulta afectado todo el hepatocito (en sus tres polos), aunque hay que tener en cuenta que no todos los hepatocitos del hígado están comprometidos, sino un porcentaje determinado. Al estar lesionado el hepatocito (hígado enfermo), se registran niveles aumentados de la enzima fosfatasa alcalina (FAL) , y sobre todo de las transaminasas: ALAT y ASAT (o GOT y GPT), fundamentales para el diagnóstico de hepatitis. La hepatitis (aguda o crónica) puede ser de origen viral (la mayoría) o por causas tóxicas, entre otras. En este tipo de ictericia, los signos pueden variar, teniendo en cuenta el grado de compromiso del hígado: si la lesión es relativamente leve, puede haber hipocolia y no haber coluria. Generalmente, el cuadro comienza con elevación de la BC, teniendo en cuenta que el polo excretor es el primero en ser afectado. CAUSAS POSIBLES: hepatitis, cirrosis. Las causas congénitas se deben al défici t de la enzima glucuronil-transferasa. Además es muy común la ictericia neonatal (en el 65% de los nacimientos, sobre todo en prematuros), por falta de maduraci ón de l hígado. SIGNOS: cada uno de los polos es afectado de diferente manera. 1. POLO CAPILAR: aumentan tanto BC como BNC (esta en mayor medida). En plasma se manifiesta solamente como aumento de la BNC, mi e ntras que el nivel de BC se mantiene estable. También puede haber hipocolia y orina normal. 2. POLO CONJUGADOR: aumenta la BNC (que se va acumulando), mientras que la BC puede ser normal. 3. POLO EXCRETOR: es el primero en ser afectado y se considera el polo más débil. El nivel de BNC es normal, pero la BC aumenta (se acumula al no poder ser excretada). Hay acolia y coluria (por el filtrado renal de la BC). ICTERICIA POST-HEPÁTICA: se encuentra afectada la zona comprendida entre el pol o excretor (polo biliar) hasta la segunda porción del duodeno, donde termina el colédoco (que vierte el líquido biliar) y el conducto pancreático o conducto de Wirsung (que vi e rte el contenido del jugo pancreático, incluyendo las enzimas pancreáticas). Ambos conductos desembocan en la ampolla de Vater (segunda porción del duodeno: los dos juntos – e n e l 75% de los casos-, o por separado). CAUSAS POSIBLES: litiasis biliar (cálculos en la vía biliar), o cáncer de cabeza de páncreas que comprima al colédoco. Ambas situaciones provocan un variable trastorno del flujo biliar denominado colestasis. En todos los casos, el hígado se encuentra sano. SIGNOS: la BC no puede pasar al duodeno ya que la vía biliar se encuentra bloqueada, por lo que retrodifunde a la sangre (aumentando la concentración plasmática de BC). Llega un momento en el que el hepatocito ya no puede conjugar bilirrubina, al “invertirse los polos” excretor y conjugador: la BC retrodifunde al plasma para luego filtrar a nivel renal, provocando coluria, además del daño al hepatocito mismo. La coluria es la coloración oscura de la orina, que se caracteriza por una capa de espuma superficial amarillenta, producto de la tensión superficial ejercida por una capa de espuma superficial amarillenta, producto de la tensión ejercida por las sales biliares a ese nivel. También hay leve aumento de l a BNC (acumulación al no poder ser metabolizada en hígado por falla del polo conjugador del hepatocito). La BC nunca llega al colon y en materia fecal, consecuentemente, se observa acolia (falta del color normal en la MF). PARTE 5: DIGESTIÓN Y ABSORCIÓN DE NUTRIENTES HIDRATOS DE CARBONO La dieta aporta normalmente un 55% de glúcidos: básicamente lactosa, sacarosa y almidón. El almidón puede ser intracelular (como en el caso de las harinas refinadas) o extracelular (como e s el caso de la cubierta celulósica de las legumbres, con uniones beta 1-4 glicosídicas, las que no pueden ser degradadas por ninguna enzima humana). Estos glúcidos sufren un proceso de degradación y posterior absorción. Digestión de hidratos de carbono En la misma actúan diferentes enzimas, como la amilasa salival, la amilasa pancreática y las disacaridasas de membrana de la chapa estriada del intestino del gado. AMILASA SALIVAL: o ptialina, producida en las glándulas parótidas. Degrada las uniones alfa 1-4, generando oligosacáridos y dextrinas límite. Las últimas son residuos de oligosacáridos que no pudieron ser degradados por impedimento estérico (la disp osi ción molecular de la proteína no permite el contacto con el sitio activo de la enzima a este nivel, que queda como remanente). AMILASA PANCREÁTICA: actúa sobre el almidón, al igual que la anterior, sobre uniones alfa 1-4, generando disacáridos como la maltosa (con uniones alfa 1-4), la isomaltosa (con uniones alfa 1-6) y trisacáridos como la maltotriosa (con uniones alfa 1-4). DISACARIDASAS DE MEMBRANA: se encuentran formando parte de la membrana plasmática del enterocito (intestino delgado). Estas enzimas son: lactasa, sacarasa y maltasa (sobre uniones alfa 1-4), e isomaltasa (sobre uniones alfa 1-6), dando como productos de degradación monosacáridos (glucosa, galactosa y fructosa). Estas enzimas actúan en la digestión a nivel de la membrana y se encuentran asociadas al cotransportador Na+-Glu (denominado SGLT) por vía transcelular. Esta disposición tan cercana de la enzima con el transportador determina la importancia de las enzi mas de l a chapa estriada, ya que evitan que, debido a la acción enzimática, se produzca un exceso osmolar (al aumentar el número de partículas osmóticamente activas) a nivel luminal, mediante la rápida entrada de monosacáridos a través del transportador. Absorción de hidratos de carbono Se realiza a nivel del duodeno y yeyuno proximal, a través del cotransportador Na+-Glu (SGLT) asociado a las disacaridasas de membrana, y por vía paracelular. Cuando la osmolaridad es muy alta, como en los sectores altos del tubo digestivo (duodeno y yeyuno proximal) durante el período post ingesta, en este caso se produce difusión simple por vía paracelular. En todos los casos, la membrana plasmática es el factor limitante para la absorción de hidratos de carbono (debido a su carácter hidrofílico y la presencia de cargas negativas libres). La glucosa que entra por vía transcelular sale luego por canales GLUT basolaterales hacia la circulación. En el caso de los hidratos de carbono extracelulares (celulosa, con uniones beta 1-4), no son digeridas por la ausencia en el humano de una beta 1-4 glicosidasa, y llegan a nivel del colon donde las bacterias (fermentativas), los degradan a ácidos volátiles (denominados aniones orgánicos), que al ser osmóticamente activos, confieren el volumen y la consistencia a l a mate ri a fecal. Si se producen aniones orgánicos en exceso (por una ingesta importante de hidratos de carbono o por insuficiencia pancreática que determina la falta de enzimas sacarolíticas) puede producirse diarrea osmótica y acidorrea. PROTEINAS El proceso digestivo de las proteínas comienza con el proceso mecánico de trituración a nivel de l a boca, prosigue en el estómago (donde el pH bajo del HCl desnaturaliza a las proteínas) y final i za a nivel del intestino delgado (donde son finalmente degradadas por enzimas pancreáticas e intestinales, para luego ser absorbidas. Degradación de proteínas A NIVEL DEL ESTOMAGO: Las estructuras proteicas son desnaturalizadas por el bajo pH, por lo que sólo se conserva su estructura aminoacídica lineal. Además, las células principales producen pepsinógeno (pro-enzima) que se activa por el bajo pH atacando l a estructura lineal de las proteínas antes desnaturalizadas. A NIVEL DEL INTESTINO: (que además recibe enzimas pancreáticas) se produce n e nzimas que luego son liberadas hacia la propia luz. Estas enzimas son aminopeptidasas (que actúan sobre el amino terminal de la estructura lineal) y dipeptidasas (que separan dipéptidos en aminoácidos libres). A NIVEL PANCREÁTICO: se produce la pro enzima tripisinógeno, la que se activa a tri psi na mediante la enzima enteroquinasa (ubicada en la membrana de la chapa estriada), también por un mecanismo autocatalítico (utilizando Ca2+ como cofactor). La tripsina tiene una alta especificidad por aminoácidos aromáticos, los que resultan ser el e stímulo más importante para la liberación de las enzimas pancreáticas. La tripsina, a su vez, acti va a todas las demás proenzimas liberadas a la luz intestinal por el páncreas, como procarboxipeptidasa (que se activa a carboxipeptidasa, una exopeptidasa que actúa sobre aminoácidos del extremo COOH), el quimiotripsinógeno (que pasa a tripsinógeno) y la proelastasa (que pasa a elastasa). Absorción de proteínas La absorción más frecuente de los aminoácidos remanentes es la dependencia de Na+, por transporte activo secundario, mediante el Modelo de Crane. Por otra parte, existe un mecani smo de absorción independiente de Na+, a través de canales para aminoácidos. En caso de no poder degradarse satisfactoriamente, por insuficiencia funcional pancreática (por ej.), las proteínas tienden a pudrirse por acción de las bacterias putrefactivas del colon izquie rdo. El exceso de animas en la MF (el VN es de 2 g por día) se denomina azotorrea, mientras que la aparición de fibras de carne no digeridas, se denomina cratorea. LÍPIDOS: digestión y absorción La dieta aporta dentro de la fracción lipídica (variable en cada individua) un 90% de TAG, mientras que el restante 10% corresponde al colesterol y otros lípidos. Los mismos sufren, en el tubo digestivo, tres procesos importantes: solubilización, degradación y absorción. 1) SOLUBILIZACIÓN: la misma permite un aumento en la interfase enzima-sustrato, y se logra gracias a un proceso denominado micelación. Los lípidos llegan al duodeno como una micela exógena (o gástrica), conteniendo la masa lipídica de la dieta, con gran hidrofobicidad. En la micela exógena, los lípidos se ubican de manera característica: los más hidrofóbicos (como el colesterol esterificado) en el centro, mientras que los más hidrofílicos (como los ácidos grasos) se ubican en la periferia. Por otro lado, la micela endógena (o biliar), conteniendo las sales biliares, llegan a la segunda porción del duodeno para allí unirse a la micela exógena para formar una micela mixta, mucho más soluble y posibilitando una mejor interacción con las enzimas lipolíticas, que van a degradar a los lípidos. 2) DEGRADACIÓN: la misma ocurre luego del proceso de micelación (solubilización de los lípidos), y en ella intervienen diferentes enzimas como: LIPASA LINGUAL: es fabricada por las glándulas de Von Ebner (a nivel de la V lingual), que tienen un rango óptimo para su acción a un pH igual al presente en la luz estomacal, donde son activas. LIPASA PANCREÁTICA: las cargas negativas de la micela (por las sales biliares) evitan su contacto, por lo que necesita de la co-enzima colipasa (producto de la conversión de procolipasa por efecto de la tripsina y del bajo pH estomacal). La enzima lipasa es una alfa-lipasa, por lo que no puede remover el ácido graso esterificado con el hidroxilo en la posición Beta (segundo carbono de la mol é cul a de glicerol). Es por esto que debe actuar a este nivel una enzima isomerasa, que traslada el ácido graso desde la posición beta hasta una posición alfa (primer carbono, desde un extremo o desde el opuesto). FOSFOLIPASAS: son enzimas que degradan fosfolípidos, dando lugar a un ácido graso libre y a un lisofosfolípido. COLESTEROL ESTER HIDROLASA: transforma el colesterol esterificado en colesterol libre, liberando un ácido graso. 3) ABSORCIÓN: los ácidos grasos libres y los MAG (monoacilglicéridos) pueden ser absorbidos gracias a la previa solubilización por parte de las sales biliares a nivel l umi nal . Además, la bomba H+/K+ genera protonización del medio: los H+ cercanos a la membrana del enterocito anulan las cargas negativas libres y facilitan la difusión simple a través de l a membrana plasmática. El factor limitante para la absorción de los lípidos es la capa de agua no removida. Las sales biliares “rodean” al lípido (solubilizándolo), y lo hacen atravesar esa capa. Ya en la vecinidad con la membrana plasmática los lípidos atraviesan fácilmente la misma por difusión (dada su liposolubilidad) mientras que las sales biliares (de naturaleza anfipática, es decir, formadas por sectores hidrofóbicos e hidrofílicos en la misma molécula) no pueden hacer lo mismo, y sólo pueden ingresar en cotransporte con Na+ (mecanismo IBAT). Dentro de la célula, un 30% de los ácidos grasos libres resintetizan TAG (gastando 2 mol e s de ATP), mientras que el restante 70% pasa a circulación linfática. Por otra parte, los ácidos grasos de cadena corta y media, pasan, debido a su menor tamaño, directamente a los capilares sanguíneos. La obstrucción de la ampolla de Vater (que recibe al conducto pancréatico y al col é doco) por un lito (cálculo originado en la vesícula biliar) puede dar lugar a una insuficiencia pancreática funcional por la no llegada de las enzimas lipolíticas a la luz del duodeno. Esto se traduce en esteatorrea, es decir, un exceso en la grasa de la MF (el valor normal de grasa en MF es el 5% del total ingerido). Cuando hay esteatorrea, las grasas tiende a “enranciarse” (al oxidarse), a nivel del colon, por acción bacteriana, dando un olor característico de la esteatorrea. CONCEPTO DE MICELA Una micela es una estructura formada como consecuencia de un fenómeno físico-químico espontáneo que se produce por la interacción de un grupo determinado de moléculas anfipáticas que contacta con una fase líquida, manteniendo su estabilidad. La micela hace que la interfas e enzima-sustrato aumente, partiendo siempre de un mismo volumen, y tiene como función de favorecer el accionar de las enzimas lipolíticas, al disminuir la tensión superficial generado por l as moléculas lipídicas (hidrofóbicas). Este proceso se ve favoreci do por el aumento de la temperatura y de la motilidad. La concentración micelar crítica (CMC) es la mínima concentración de sales biliares para que la absorción de lípidos no descienda. El valor normal de la CMC es de 1-2 mM. En caso contrario puede generarse un fenómeno de esteatorrea (aumento de lípidos en la MF). Se pueden describir distintos tipos de micelas: MICELA EXÓGENA: también denominada micela gástrica, se origina a partir de los l ípi dos provenientes de la dieta. Las moléculas se van ordenando según su hidrofobicidad (las más hidrofóbicas se ubican más hacia el centro y las menos hidrofóbicas a nivel de la superficie). Desde el centro de la micela exógena hacia la periferia, se disponen: colesterol esterificado, TAG, fosfolípidos, DAG, MAG, ácidos grasos libres (AGL) y finalmente las lipoproteínas. MICELA ENDÓGENA: también denominada micela biliar, se forma en la vesícula biliar al lograr una CMC, circulando por la vía biliar. Las sales biliares son las responsables de la micelación definitiva, al unirse y solubilizar a la micela exógena a nivel del duodeno, formando la denominada micela mixta. Desde el centro de la micela endógena hacia la periferia, se disponen: colesterol no esterificado, fosfolípidos, y sales biliares. Ahora se genera una interfase adecuada para la acción de las enzimas lipolíticas, responsables de l a degradación lipídica. PARTE 6: MANEJO HIDROELECTROLÍTICO DEL TUBO DIGESTIVO La eficiencia, en cuanto al manejo de agua y electrolitos, depende de la relación entre la carga que llega a determinado sector del tubo digestivo y la carga que sale del mismo, en definitiva depende del porcentaje absorbido, independientemente del volumen absoluto. Para el manejo de agua, un sector como el colon se considera muy eficiente, teniendo en cuenta que, a pe sar de que pose e una absorción diaria neta menor que el duodeno (por ejemplo), es capaz de absorber un mayor porcentaje de esa carga que le llega. En cuanto al manejo de electrolitos, se considera que un sector del tubo digestivo es más eficiente si puede transportar electrolitos aún en contra de gradiente químico, sin importar el volumen neto. En este sentido, entonces, el colon es el sector más eficiente para la absorción de a gua y electrolitos. Se considera que el colon tiene una eficiencia del 90%, mientras que el duodeno yeyuno tienen un 40% y el íleon un 50%. El TCD (Túbulo contorneado distal), y el túbulo col e ctor, son los sectores más eficientes del sistema renal y además, al igual que el colon, son epitelios cerrados. El ángulo de Treitz (ángulo duodeno-yeyunal) es el límite en el cual el quimo (hipertónico), se isotoniza. Es necesario considerar que en epitelios abiertos, como en los sectore s al tos de l tubo digestivo (el punto de corte es el ángulo de Treitz), las concentraciones de Na+ y K+ se manti e ne n en equilibrio con el plasma, debido a que estos iones se distribuyen fácilmente por difusión a ambos lados del enterocito. Por otro lado, ya a nivel del íleon hay absorción de Cl- con secreción de HCO3- (por el me cani smo de antiporte Cl-/HCO3-). Se secreta HCO3- hasta que el Cl- alcanza una concentración intrace l ular de 50-60 meq/l, mientras que el HCO3- aumenta en la misma proporción pero en la luz. En el colon hay una alta secreción de HCO3- y de K, mientras que se sigue absorbiendo Cl-. Diferencia entre tonicidad y osmolaridad Los términos tonicidad (híper, hipo) y osmolaridad (híper, hipo) se utilizan con mucha frecuencia y muchas veces como sinónimos, lo cual es un error. En realidad la tonicidad se refiere a las cargas eléctricas (partículas eléctricamente activas) que se distribuyen a ambos lados de una membrana o con respe cto del plasma. Por ejemplo: en la MF hay hipertonicidad para el K+ e hipotonicidad para el Na+ y el Cl -, ya que estos dos últimos se encuentran menos concentrados respecto del plasma. Como éstos son iones y están cargados eléctricamente, se cuantifican en equivalentes (eq) o en miliequivalentes (meq). Por otro lado, la osmolaridad (Osm) se refiere a las partículas osmóticamente activas (con capacidad para arrastrar agua) que se distribuyen a ambos lados de una membrana o con respecto del plasma. Por ejemplo, en el período postprandial, la osmolaridad de la luz del duodeno es mucho mayor que la osmolaridad plasmática (que es de aproximadamente 300 mOsm). Como éstas son partículas osmóticamente activas, se cuantifican en osmoles o en miliosmoles. Como puede imaginarse, no todas las partículas osmóticamente activas son partículas cargadas eléctricamente (iones), ejemplo de esto, es la glucosa, que carece de cargas. Por tanto, no siempre tonicidad es sinónimo de osmolaridad. La tonicidad de la luz del tubo digestivo se equilibra con la del plasma antes del ángulo de Treitz, a partir del cual los contenidos luminales son isotónicos, mientras se trate de intestino delgado. Entonces en los sectores altos del tubo digestivo, (intestino delgado), la concentración luminal de cada uno de los iones es la misma que la del plasma. Como se verá, en el colon habrá ciertas modificaciones en cuanto a la tonicidad. Diferencias entre los tipos de epitelios 1) BARRERAS EPITELIALES: están conformadas por los enterocitos. Estas células se encuentran polarizadas: poseen una membrana luminal o mucosa (de cara a la luz del tubo digestivo) y una membrana basolateral o serosa (la que contacta con la circulación y con las células vecinas). Las uniones intercelulares determinan que el epitel io sea abierto o cerrado, en función de que estas uniones estrechas seas más o menos permeabl es. Estas uniones estrechas son diferenciaciones de membrana que acumulan en su superficie cargas negativas, por lo cual tienen gran afinidad por los cationes, sobre todo si son i one s monovalentes como el Na+ o el K+. La diferencia de potencial transepitelial se genera a partir de la separación que l a barre ra epitelial (formada por los enterocitos) establece entre la luz del tubo digestivo y el intersticio. 2) EPITELIO ABIERTO: o epitelio LEACKY, es un epitelio cuyas uniones estrechas intercelulares son muy permeables, y permiten la libre difusión de agua y electrolitos mediante la llamada vía paracelular (shunt path). Hay que tener en cuenta que el epitelio digestivo, forma una suerte de membrana o barrera que separa el medio luminal del tubo dige sti vo (medio externo) del medio interno (la circulación, en este caso). Este ti po de e pi tel io e s característico de los sectores altos del tubo digestivo: duodeno, yeyuno e íle on. CARACTERISTICAS: Uniones estrechas permeables Alta conductancia de membrana Baja resistencia de membrana Diferencia de potencial transepitelial de 0 mV No es capaz de mantener gradientes químicos (porque si disipan fácilmente) Maneja altos volúmenes (hasta 9000 ml) Presente en duodeno, yeyuno, vesícula biliar y riñón (en el TCP = túbulo contorneado proximal) 3) EPITELIO CERRADO: o epitelio TIGHT, se caracteriza por presentar uniones estrechas poco permeables para los iones; el transporte característico es el transcelular, y en algunos casos activo. En el caso del manejo del agua, estas uniones estrechas (poco permeables para los iones) no representan ningún obstáculo para su pasaje. Este tipo de e pi te l ios se encuentra en los sectores bajos del tubo digestivo, como el colon. CARACTERÍSTICAS: Uniones estrechas poco permeables para los iones, pero no para el agua que difunde libremente Baja conductancia de membrana Alta resistencia de membrana Diferencia de potencial transepitelial de -20mV Es capaz de mantener gradientes químicos Es capaz de generar gradiente osmótico Maneja volúmenes relativamente bajos (hasta 1500 ml) Presente en íleon terminal, colon, y riñón (túbulo contorneado distal y túbulo colector). ENTRADA DE AGUA AL DUODENO (EN 24HS) Desde la dieta: 2000 ml Secreción salival: 1000 ml Secreción estomacal: 2000 ml Secreción pancreática: 2000 ml Secreción biliar: 1000 ml Secreción intestinal: 1000 ml TOTAL de agua que arriba al duodeno = 9000 ml En el duodeno se absorben 4000 ml, mientras que 5000 ml pasan al íleon. En el íleon se absorben 3500 ml, mientras que 1500 ml pasan al colon. En el colon se absorben 1400 ml (es decir, el 90% de lo que le llega), con lo cual se define como el sector más eficiente del tubo digestivo, en cuanto a la capacidad de absorber agua en contra de gradiente y en tal proporción. No se debe confundir el concepto de eficiencia con el de eficacia: en tal caso el duodeno sería el sector más eficaz, ya que absorbe un volumen de agua mucho mayor (4000 ml), al del colon (1400 ml), pero no es el más eficiente, ya que el duodeno absorbe e l 40% de lo que le llega. Pasan a formar parte de la materia fecal 100 ml de agua, provenientes del colon, esto en 24 hs. Materia Fecal La materia fecal es el producto de desecho del tubo digestivo y tiene una composición orgáni ca e inorgánica (electrolítica) determinada. Los aniones orgánicos provienen de la fermentación bacteriana (en el colon derecho) de hi dratos de carbono que no han sido absorbidos en el intestino delgado y son un componente muy importante de la materia fecal. Estos hidratos de carbono que no pueden ser absorbi dos, pue de n ser la celulosa, que no puede ser degradada por la falta de una enzima Beta 1-4 glicosidasa, o bie n almidón, que en exceso puede fermentar y generar gran cantidad de anión orgánico. La materia fecal es pobre en Na+ y Cl- y es abundante en K+ y anión orgánico. Los aniones orgánicos son ácidos grasos de cadena corta, muy volátiles, como ser el ácido fórmico, el acé ti co, el láctico, el propiónico y el butírico. Todos estos son productos de la fermentación de hidratos de carbono por parte de la flora bacteriana fermentativa del colon derecho. Corresponden al 70% de los aniones totales (orgánicos e inorgánico) en materia fecal, y su elevado poder osmóti co e s e l factor determinante de la cantidad de agua en materia fecal (por arrastre osmótico), defin iendo el volumen, la consistencia y el peso de la misma. Por otra parte el Cl - y el HCO3- son los aniones inorgánicos más importantes. El exceso de aniones orgánicos es causa de meteorismo, es decir, la producción de gases por la exagerada ingesta de hidratos de carbono, y además pueden determinar la producción de diarre a osmótica (por arrastre de agua). La concentración habitual de aniones orgánicos en MF sue l e se r de 220 meq/l. Las características de la MF son: PESO (en 24hs): 120-200 g (el 80% corresponde al agua). VOLUMEN (en 24 hs): 100-200 ml pH: aproximadamente 7 CONCENTRACIÓN IÓNICA: abundante en K+ y anión orgánico; pobre en Na+ y Cl OSMOLARIDAD: la osmolaridad real es siempre de 300 mOsm/l. Mientras que la osmolaridad Teórica Calculada suele ser de 250-280 mOsm/l ¿Por qué la osmolaridad real de la materia fecal se mantiene siempre en 300 mOsm/l? Porque e l colon es incapaz de producir contenidos luminales hipo o hiperosmolares, es decir, que de habe r un aumento en la osmolaridad luminal, éste siempre será compensado con un aumento en la secreción de agua hacia la luz, con lo cual el volumen neto de agua en la materia fecal termina siendo aumentado y la osmolaridad es mantenida en 300 mOsm/l. Diarreas Se considera diarrea cuando se superan los 200g diarios de MF, además se observan otros he chos como la disminución de la consistencia (heces más líquidas), el aumento del volumen y del número de deposiciones diarias. El ritmo intestinal varía mucho para cada individuo, y depende de hábitos alimentarios y socioculturales, sin embargo se suele tomar como parámetro normal desde 3 deposiciones diarias hasta 3 deposiciones semanales; recién por debajo de este parámetro se habla de constipación. Otro parámetro importante a considerar es la emergencia evacuatoria nocturna: si el paciente refiere levantarse de noche a causa de emergencia evacuatoria siempre se considera un rasgo patológico de origen orgánico, teniendo en cuenta que la motilidad colónica sue l e e star abol ida durante el sueño profundo. En este caso se considera diarrea orgánica (por una neoplasia endócrina por ejemplo), para diferenciarla de una diarrea funcional (de naturaleza psicosomática o de naturaleza motora, que jamás podría producirse con el individuo durmiendo). Veamos diferentes clasificaciones de las diarreas: 1) SEGÚN LA CAUSA: a) Diarrea Fermentativa: por alta ingesta de hidratos de carbono, que produce saturación de los mecanismos de absorción del intestino delgado y, consecuentemente, mayor sustrato para las bacterias fermentativas del colon derecho. También puede ser consecuencia de un síndrome de malabsorción por déficit de enzimas sacarolíticas o por déficit en la funcionalidad del epitelio absorti vo intestinal, con lo cual los hidratos de carbono no absorbidos llegan a nivel de l a luz de l colon y resultan osmóticamente activos, además de ser sustrato para las enzimas bacterianas fermentativas, con mayor producción de anión orgánico y aún mayor arrastre osmótico. b) Diarrea Secretoria: por exacerbación del MGS (Modelo General de Secre ción). El mejor ejemplo de una noxa que produzca este cuadro es el vibrio chole rae (bacte ria productora del cólera), que aumenta la activación del MGS por estímulo sostenido de l AMPc actuando sobre receptores para el VIP. c) Diarrea osmótica: mucha pérdida de agua por uso de laxantes alcalinos, por ejempl o, sales de magnesio. 2) SEGÚN LA OSMOLARIDAD: a) Diarrea Fermentativa: menor o igual a 250 mOsm/l b) Diarrea Secretoria: mayor a 250 mOsm/l c) Diarrea Osmótica: menor o igual a 250 mOsm/l 3) SEGÚN EL pH: a) Diarrea Fermentativa: menor o igual a 5,5 (por aumento del anión orgânico –HCO3-) b) Diarrea Secretoria: 5,5 - 7 c) Diarrea Osmótica: mayor a 7 (por laxantes, por aumento de partículas alcalinas) La diarrea secretoria también puede ser de tipo inflamatoria o infecciosa. En el caso de la bacteria del cólera, ésta ingresa por vía oral (en alimentos o en agua contaminada) y tiene e fe cto mediante su toxina, responsable de la patogenia. La toxina tiene dos subunidades: la B5 se une a la membrana del enterocito, mientras que la región A2 actúa como “muñón funcional”. Esta úl ti ma se divide a su vez en dos regiones: la región A1 que es la que ADP-ribosila a la subunidad alfa de la proteína Gs, con pérdida irreversible de la ATPasa (que pasa ATP a ADP, inactivando el mecanismo), con lo cual la proteína Gs estimula indefinidamente a la enzima adenilato ciclasa (que produce altas cantidades de AMPc), que sobreexcita a todo el MGS. La otra región, A2, es accesoria. Manejo electrolítico del tubo digestivo 1) DIFUSIÓN SIMPLE: es el pasaje de partículas desde uno hacia otro lado de una membrana, cuya fuerza impulsora para dicho pasaje es la presión hidrostática dada por la diferencia de concentración de solutos a ambos lados de la membrana; el pasaje de partículas es desde el compartimiento más concentrado al menor concentrado, para “equilibrar” las concentraciones en ambos compartimientos. Este es un mecanismo pasivo, ya que no hay gasto directo de ATP. 2) DIFUSIÓN FACILITADA: es el pasaje de partículas a través de transportadores específicos y saturables denominados carriers (proteínas transmembrana) que tienen un movi miento de flip-flop o flipping que les permite captar a la molécula específica del lado luminal, rotar dentro de la membrana y finalmente liberar la molécula al medio intracelular. En un momento se llega a un flujo máximo en el que el sistema se satura y que es independiente de la diferencia de concentración. Por otro lado, si no existiera gradiente transmembrana no existiría pasaje alguno, por más que todos los transportadores estén libres. 3) DIFUSIÓN POR CANAL: el pasaje de las moléculas es pasivo y no existe gran especificidad: es decir, si el interior de la proteína canal (transmembrana) está “tapizado” de cargas negativas, entonces este canal tendrá afinidad por iones de carga positiva (cationes). En este caso, además del gradiente químico es necesario un gradiente eléctri co para que e l mecanismo funcione. Los canales, a su vez, pueden ser voltaje dependientes o ligando dependientes. 4) COTRANSPORTE: también denominado symport, es un transporte activo secundario, es decir que utiliza como fuerza impulsora para el pasaje al gradiente de disipación de l Na+, creado por el antiporte Na+/K+ (que gasta ATP directamente). Este mecanismo se uti li za para el pasaje de moléculas hidrosolubles como glucosa, aminoácidos y algunas vitaminas (como la C y las del complejo B). En este transporte, la utilización del ATP como fuente de energía es indirecta. 5) CONTRA-TRANSPORTE: también denominado antiport es el transporte de un elemento en dirección opuesta al elemento que provee la energía disipadora (suele ser el Na+). Ejemplo de este mecanismo es el contra-transporte Na+/H+. También se considera un transporte activo secundario. 6) TRANSPORTE ACTIVO (primario): utiliza directamente energía proveniente del ATP para transportar partículas en contra de gradiente electroquímico. Teóricamente, como todo transportador, es un mecanismo saturable. La bomba Na+/K+ ATPasa basolateral es el mejor ejemplo. 7) INTERNALIZACIÓN MEDIADA POR RECEPTOR: es el pasaje de partículas con especifici dad por una proteína de membrana que es internalizada por la misma para liberar la partícul a al interior celular (donde se pierde la afinidad del ligando con el receptor). El pasaje es proporcional a la concentración de las partículas (en principio), siendo el factor l i mitante para este mecanismo, el número de receptores. Mecanismos específicos de transporte SODIO 1) SECRECIÓN DE Na+: mediante la vía paracelular por el MGS 2) ABSORCIÓN DE Na+: A) Contratransporte con nutrientes: como aminoácidos, vitaminas hidrosolubles, glucosa (en este caso, denominado Modelo de Crane). Este último se ve más que nada a ni ve l del yeyuno y no se observa en el colon. B) Difusión simple: por la vía paracelular (vía selectiva para cationes, por las cargas negativas de la superficie de las uniones estrechas). El Mg2+ de algunos antiácidos neutraliza las cargas negativas, evitando el pasaje de H+ (acción antiácida), pero también evita el pasaje de otros cationes monovalentes como lo es el Na+. C) Solvent drag: es el pasaje por la vía paracelular cuando se produce la absorción de glucosa (el gradiente osmótico de la misma en este caso arrastra al Na+). Se observa sobre todo en yeyuno. D) Antiporte Na+/H+: se pone en marca cuando se secreta HCO3- (por el antiporte Cl/HCO3-) como mecanismo compensatorio. Se ve en yeyuno y colon. E) Cotransporte Na+ - Cl-: se observa en el colon. F) Canales electrogénicos: son de ubicación luminal, regulados basolateralmente por hormonas como la aldosterona y la ANG II (en el colon izquierdo). El colon es el sitio con mayor capacidad de conservación del Na+. G) Cotransporte Na+ - AO: a nivel del colon derecho, el sector donde la flora fermentativa produce anión orgánico a partir de hidratos de carbono no digeridos. CLORO 1) SECRECIÓN: por canales electrogénicos de ubicación luminal y presentes en todo e l tubo digestivo, formando parte del MGS. 2) ABSORCIÓN: A) Difusión simple: por vía paracelular, sólo al atenuar las cargas negativas de la superficie de las uniones estrechas, por ejemplo tras utilizar sales de magnesio. Se distribuyen en todo el tubo digestivo formando parte del MGS. B) Cotransporte Na+ - Cl: a nivel del colon C) Antiporte HCO3-Cl-: su activación induce al antiporte Na+/H+ al disminuir la concentración de bicarbonato intracelular. Se ve a nivel del íleon y en duodeno. POTASIO 1) SECRECIÓN: A) Canales electrogénicos: de ubicación luminal, presentes en todo el tubo digestivo, formando parte del MGS. B) Canales electrogénicos del colon izquierdo: de ubicación luminal, activados basolateralmente por la unión de la aldosterona a su receptor específico. 2) ABSORCIÓN: A) Difusión simple: mediante la vía paracelular, es parte del MGS, a nivel del intestino delgado. B) Bomba H+/K+: tiene la capacidad de secretar protones hasta lograr un gradie nte de hasta 2000 unidades de diferencia con respecto al medio interno. Se encuentra e n l a célula parietal y en el colon. Este mecanismo se inhibe por la droga omeprazol (IBP = inhibidor de la bomba de protones). BICARBONATO 1) SECRECIÓN: A) Antiporte Cl-/HCO3-: se observa en íleon y colon. B) Canales electrogénicos: de ubicación luminal, se observan en duodeno y colon. El duodeno es el sitio de mayor secreción de HCO3- (como parte de la barrera defensiva, similar a la presente en el estómago). C) Difusión simple: a través de la vía paracelular del MGS, en todo el tubo digestivo. 2) ABSORCIÓN: A) Antiporte Na+/H+: con secreción de H+ y aumento de la concentración intracelular de HCO3-, y este puede pasar a la circulación utilizando el antiporte cloro bicarbonato de la membrana del polo basal del enterocito. Se observa sobre todo en yeyuno. En realidad no hay una absorción desde la luz, pero la salida del H+ produce aumento de l HCO3- intracelular, teniendo en cuenta que ambos son el producto de la hidrólisis de l ácido carbónico. B) Bomba H+/K+: con secreción de H+, en este caso tampoco hay realmente absorción de HCO3- desde la luz, pero la salida de H+ (producto del ácido carbónico, mediada por la anhidrasa carbónica), hace que aumente la concentración intracelular de bicarbonato sucediendo lo que en el ítem anterior se explicó.