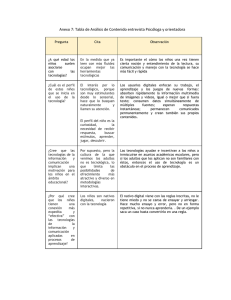
MODELOS CAUSALES CLÁSICOS Fernando Alvarez La Psicología
Anuncio

1 MODELOS CAUSALES CLÁSICOS Fernando Alvarez La Psicología decimonónica era la ciencia de la mente, y su método de análisis la introspección. La quiebra de esta tradición se gestó en varios frentes en la segunda mitad del siglo XIX. Por una parte, en el ámbito de los naturalistas en Gran Bretaña en la década de 1870, el enfoque de Douglas Spalding (1872, 1873) y Charles Darwin (1872) en la observación y experimentación con el comportamiento abriría el camino a la Etología. Por otra parte, el interés hacia el comportamiento del filósofo funcionalista John Dewey (1927-1928) y del fisiólogo Iván P. Pávlov (1927) animarían el nacimiento del Conductismo (Watson, 1913). Se revisarán a continuación los modelos teóricos producidos por ambas tradiciones científicas a lo largo del siglo XX, como explicaciones causales de la conducta. Comportamiento apetitivo y consumatorio W. Craig (1876-1954), norteamericano, discípulo del reputado ornitólogo C. O. Whitman (1842-1910), propone en 1918 el primer modelo teórico sobre la estructura del comportamiento. Según Craig, el comportamiento que él consideraba instintivo no era una mera cadena de reflejos, sino que se desarrollaba en ciclos, los que se compondrían de cuatro fases. La primera de ellas, que Craig denominaría “Comportamiento Apetitivo”, consistiría en un estado de agitación, que se mantendría mientras ciertos estímulos clave estuvieran ausentes. Este comportamiento se manifestaría en la búsqueda de dichos estímulos, así como en mayor tensión muscular. Además, los actos más 2 simples que compondrían esta fase serían flexibles, adaptándose mediante el aprendizaje a las circunstancias variables que el animal encontrara en su búsqueda. La segunda fase (el “Comportamiento Consumatorio” o de satisfacción) se desencadenaría ante la presencia del estímulo clave (los que serían después conocidos como “Estímulos-signo”). Este comportamiento sería innato y de estructura fija. Independientemente de Craig, J. von Uexküll (1909) considera esos estímulos clave, desencadenadores de actos concretos, en su propio esquema causal de la conducta. Por su parte, O. Heinroth (1910) describe la fase consumatoria del modelo de Craig como “arteigene Triebhandlungen” (o actos instintivos característicos de especie). Posteriormente, en la Etología clásica, este comportamiento sería conocido como “Pautas de Acción Fijas”. La tercera fase sería de saciedad y aversión al estímulo, y la cuarta fase sería de descanso e independencia respecto a él. El modelo de Craig, con las aportaciones de J. von Uexküll (1909) y O. Heinroth (1910), fue aceptado por K. Lorenz y N. Tinbergen, e incluido en sus propios esquemas causales del comportamiento. No obstante, el modelo presenta varias dificultades. Así, no es siempre fácil determinar qué componente del comportamiento sería el apetitivo, ya que con frecuencia no se observa ninguna conducta exploratoria (por ejemplo, en la caza al acecho), mientras que en la alarma o en la agresión hay dudas de su presencia, o no se da en absoluto. En lo que respecta a la fase consumatoria del esquema, la aplicación al comportamiento sexual (el más frecuentemente utilizado como ejemplo) también presenta problemas: la conducta de fertilización de los huevos en peces no siempre reduce la motivación sexual. Umwelt Para Jacob von Uexküll (1864-1944) los animales son sujetos cuya función esencial es la de percibir y actuar. En su obra Umwelt und Innenwelt der Tiere (1909), 3 von Uexküll huye de la falacia de un mundo único en el que se encasillarían todos los seres vivos, e introduce el concepto de Umwelt para denotar el mundo subjetivo de cada animal, y que incluye tanto su universo de percepción como de ejecución. Según esta idea, al nivel de percepción, cada animal se desenvolvería en un mundo propio compuesto de ciertos y escasos “estímulos-clave” ambientales, los necesarios para el desarrollo de sus necesidades vitales, que envuelven a cada individuo en una burbuja particular (Uexküll, 1934), y determinan su comportamiento mediante el aparato efector disponible. Para von Uexküll la relación entre el animal, con sus órganos y necesidades especiales y su universo propio, subjetivo y significativo (el Umwelt), constituyen el “sistema funcional” (Funktionskreis) o sistema interactivo. Así pues, el umwelt, propio mundo subjetivo del individuo, sería diferente de su habitat (externo y objetivo, tal como lo describiría un ecólogo) y de su nicho (en el sentido de la función ecológica de la especie en el ecosistema) (Emmeche, 2001) El mecanismo interno encargado de filtrar los estímulos (el “Esquema Desencadenante Innato”) se comparaba a una cerradura que cierra o abre el paso (mediante los “estímulos-clave”) a la realización del comportamiento innato. En su esquema del Umwelt, von Uexküll se aparta en todo momento de una visión mecanicista de la percepción y la acción, y, en consonancia con su visión vitalista, ve al animal en todo momento no como una máquina, sino más bien como el ingeniero que opera la máquina en el centro de la burbuja de su Umwelt. El Umwelt no llegó a establecerse como concepto operativo en los estudios etológicos, aunque tuvo gran influencia en la primera Etología, especialmente sobre las ideas desarrolladas por Konrad Lorenz, siendo después olvidado por largo tiempo. Sin embargo, con el desarrollo de la Biosemiótica, Zoosemiótica y Medicina Psicosomática, el concepto de Umwelt ha cobrado de nuevo interés, como mundo subjetivo en el que se desenvuelve cada ser vivo, en relación con su equipo específico de procesado de la información, y sus códigos y sistema de señales (T. von Uexküll et al., 1993). 4 La idea de imagen de búsqueda, también propuesta por von Uexküll, deriva directamente del concepto de Umwelt, y se refiere a la más fácil percepción de ciertos objetos del ambiente. Esta percepción selectiva tendría muy evidente valor adaptativo, principalmente en relación con la búsqueda de alimento, y podría variar de acuerdo al último objeto encontrado (Uexküll, 1934; Ruiter, 1952; Fig. 1). Figura 1.- La imagen de búsqueda de la ya desaparecida jarra de barro sobre la mesa impide momentáneamente ver la botella de cristal en su lugar (según ejemplo de Uexküll, 1934). No es siempre fácil saber si los animales han adquirido una imagen de búsqueda, y así, podemos observar a un animal comenzando a comer cierto alimento críptico al que antes no prestaba atención (y adquiriendo la costumbre de comerlo), pero el cambio en el hábito alimenticio puede no deberse a la adquisición de la nueva imagen de búsqueda, sino a otros procesos de aprendizaje que pueden estar ocurriendo a mismo tiempo. Hay ejemplos, no obstante, de la validez del concepto en la mayor eficacia en la detección de alimento artificialmente críptico, una vez que se detecta, por pollos de gallina (Gallus gallus domesticus), y en la respuesta por la corneja (Corvus corone) al polimorfismo artificial de presas experimentales (McFarland, 1987). 5 Modelos de aprendizaje Mediante los procesos de aprendizaje y por efecto de la experiencia se producen cambios adaptativos en el comportamiento. Se han logrado clasificar las formas de aprendizaje y se han elaborado modelos del mismo merced a los estudios de fisiólogos, etólogos y, sobre todo, psicólogos. Consideraremos aquí las formas de aprendizaje no asociativo, del asociativo y del aprendizaje complejo. Aprendizaje no asociativo A veces, como consecuencia de responder repetidamente a un determinado estímulo, un organismo cambiará la propia respuesta, bien disminuyéndola (Habituación) o incrementándola (Sensibilización). Estas dos formas de aprendizaje no asociativo (sin implicar una nueva asociación entre diferentes estímulos y la respuesta), desencadenadas por la simple estimulación repetitiva, permiten al organismo aprender acerca de las propiedades del estimulo en cuestión. Habituación. Thorpe (1963) la define como la extinción relativamente permanente de la respuesta, como resultado de la estimulación repetida, y que no se acompaña de ningún reforzamiento. Puesto que la habituación es relativamente duradera y específica del estímulo, puede considerarse diferente de la fatiga o la adaptación sensorial. En sentido amplio pues, habituación sería una forma de aprender a no responder a estímulos sin relevancia para la vida del sujeto: la habituación en la respuesta de captura de un sapo común (Bufo bufo) ante la presentación de un objeto no comestible le ahorra tiempo y energía, que puede dedicar a otras actividades. El interés de la habituación para la supervivencia será sin duda la causa de su extendida presencia en el reino animal, desde protozoos a humanos (Thorpe, 1963; Evans, 1971; Wood, 1988). El proceso de Habituación no es tan simple como podría pensarse, de forma que si el estímulo no se presenta por un largo periodo de tiempo, la respuesta inicial 6 reaparece, y si la habituación y la siguiente recuperación de la respuesta se repite varias veces, la propia habituación se realiza más rápidamente en la sucesión de respuestas. Sensibilización. La habituación suele ocurrir como respuesta a estímulos relativamente insignificantes, mientras que ante estimulación más potente puede darse el efecto diametralmente opuesto de Sensibilización (sin descartar que puedan producirse los dos efectos simultáneamente). En esta última modalidad de aprendizaje no asociativo se da un incremento más o menos permanente en la respuesta o una reducción en la latencia o en el umbral de reacción como consecuencia de la estimulación repetitiva del mismo estímulo (McConnell, 1966; Evans, 1971). Un pulpo (Octopus vulgaris) se retirará a su refugio o saldrá de él para atacar la pieza de alimento que se le ofrece en el otro extremo del acuario. La probabilidad de que salga del refugio y ataque un estímulo neutro (por ejemplo, un disco blanco de plástico) aumentará si recientemente se le ha ofrecido alimento, y disminuirá si recientemente se le han aplicado descargas eléctricas. No hay ningún premio o castigo implicados, pues el ofrecimiento de alimento o la descarga eléctrica no se aplican en relación con el comportamiento que el pulpo haya realizado (Hochner et al., 2006). La Sensibilización representa un acomodo a las condiciones ambientales, y debe tener importante repercusión en la supervivencia. Así, si se da cierta regularidad en el ambiente, debe compensarle a un animal salir de su refugio en busca de alimento si recientemente lo ha obtenido (pues es probable que lo siga encontrando), o evitará hacerlo si recientemente se ha hallado en gran peligro. Esta forma de aprendizaje parece estar muy extendida, sobre todo en invertebrados, y puede considerarse precursora del aprendizaje asociativo. 7 Aprendizaje asociativo En las formas de aprendizaje asociativo o condicionamiento, la exposición a los estímulos permite al organismo aprender acerca de las relaciones entre ellos y con las respuestas. Aunque los primeros experimentos al respecto datan de principios del siglo XX (Fig. 2), el modelo tiene precedentes en varios filósofos asociacionistas, quienes suponían que las ideas complejas se construyen por asociación de ideas más simples (Hartley, 1749; Mill, 1829). Figura 2.- Laberinto en T usado por Yerkes (1912) para el estudio del aprendizaje en Lombriz de Tierra (Lumbricus terrestris). Condicionamiento Clásico. En el condicionamiento más simple dos estímulos se presentan uno tras otro en un corto intervalo de tiempo. Uno de dichos estímulos es efectivo por sí mismo desencadenando automáticamente una respuesta concreta, sin necesidad de aprendizaje alguno, es el Estímulo Incondicionado (EI). El otro estímulo (el Estímulo Condicionado, EC) es neutro, es decir, no desencadena por sí mismo la respuesta, y puede ser de tipo discreto o contextual. Como resultado de la asociación en el tiempo del EC precediendo al EI, el EC adquiere parte de las propiedades desencadenantes del EI, pudiendo ejecutarse la respuesta en ausencia de éste, y con tan sólo la presentación del EC. El ejemplo clásico es el perro de Pávlov oyendo el sonido de una campana (EC) antes de la presentación del alimento (EI), que tiene por 8 sí el efecto de producción de saliva en los perros. Como resultado, después de varias pruebas (de la asociación entre el EI y el EC), el perro producirá saliva con tan sólo escuchar el sonido de la campana. Según Pávlov (1927), las respuestas al estímulo incondicionado podrían llegar a ser desencadenadas por otros estímulos (los condicionados) por mera asociación. Su Ley de los Reflejos Condicionados sería aplicable a los actos reflejos, y no a lo que habitualmente se conoce como respuestas voluntarias. Condicionamiento Operante. En el condicionamiento más complejo, o Condicionamiento Instrumental son las consecuencias de la conducta, en el curso de varias pruebas, las que afectan al comportamiento que las precede. Es decir, una cierta conducta se seguirá de adición o sustracción de su refuerzo o castigo, o puede seguirse de ninguna consecuencia. Así pues, la respuesta (que, en este caso, sería un acto independiente y “voluntario” del animal) precedería en el tiempo a las consecuencias, y se vería afectada por un efecto retroactivo de aquéllas. Según el modelo conocido como Ley del Efecto (Thorndike, 1901), si un comportamiento va acompañado o seguido de satisfacción (accionar el mecanismo de apertura, que permita a un gato salir de la jaula), cuando la situación surja de nuevo, el animal tenderá a repetir aquel comportamiento. Y, al contrario, si el comportamiento va acompañado de insatisfacción, el animal tenderá a evitar dicho comportamiento. En la modificación más objetivista (manos mentalista) del modelo realizada por Skinner (Ley de Refuerzo, 1938), cuando al comportamiento realizado por un animal le sigue la presentación de un refuerzo, aumentará la probabilidad de que ese comportamiento se repita. Aprendizaje Complejo Impronta Filial. Para Lorenz (1935) la Impronta es el proceso por el que los recién nacidos establecen una rápida, irreversible e intensa relación con sujetos sociales (generalmente los padres) o sus sustitutos (incluso un objeto sin ningún parecido a la 9 especie a la que los sujetos pertenecen). El efecto se da principalmente en especies de aves y mamíferos de desarrollo precoz, y tiene lugar durante un “periodo crítico” restringido en el estadio juvenil temprano. El fenómeno fue descrito por primera vez por Douglas Spalding en 1873 sobre pollitos de gallina, llegando a la conclusión que la respuesta de seguir por parte de los pollitos era instintiva y no aprendida, mientras que la capacidad de reconocer a la figura materna sí era aprendida. Además, Spalding constata la existencia de un periodo crítico. El efecto fue redescubierto por Oskar Heinroth (1911) en pollos de ansar común (Anser anser), los que recibían la impronta (einzuprägen: imprimir, estampar) de la presencia humana, asociándose a los humanos como lo harían sujetos normales hacia sus progenitores, sin mostrar ninguna inclinación por seguir a coespecíficos adultos. En base a observaciones previas de O. Heinroth y de él mismo sobre pollos de patos y gansos, Lorenz (1935) define más exactamente el proceso, que denominó Prägung, o Impronta, especificando sus características. Para Lorenz la Impronta es el resultado de un instinto, y el “periodo crítico” estaría muy restringido en un corto intervalo temporal. Estudios posteriores han demostrado que el efecto puede producirse tanto por estímulos visuales como sonoros, no estando tan claramente delimitado en el tiempo como en principio se pensaba, pudiendo también ser afectado por las circunstancias que rodean al animal. Además, cualquier objeto no tiene el mismo efecto produciendo la Impronta, de forma de que pollitos de ánade real (Anas platyrhynchos) se improntan más fácilmente a objetos verdes o amarillos, y los pollitos de gallina los prefieren azules o anaranjados. No obstante, se reconoce el proceso como muy importante en el desarrollo del comportamiento en especies precoces, y quizá también en animales altriciales, incluida la especie humana. 10 Impronta Sexual. Además de afectar a la asociación paterno-filial, la experiencia temprana tendrá efectos sobre las preferencias sexuales adultas. Mediante la Impronta Sexual, cuando adultas, muchas aves mostrarán preferencias sexuales hacia individuos del sexo contrario con características similares a las de aquel que los atendió en la fase juvenil. El fenómeno fue descrito por Lorenz (1935). El efecto de la Impronta Sexual es menos marcado que el de la Respuesta de Seguir, pudiendo restringirse a uno de los dos géneros (al macho es especies dimórficas de anátidas). Además, es más fácil que se produzca hacia la propia especie, siendo menos efectiva hacia especies emparentadas, y aun menos hacia las filogenéticamente alejadas. En mamíferos no suele darse la Impronta Sexual, aunque el cuidado y alimentación en la fase juvenil suele producir tendencias sociales hacia la especie cuidadora. Aprendizaje Observacional. Cuando un animal aprende de las acciones de otro sujeto, el cual ha de resolver un problema, podemos hablar de Aprendizaje Observacional o Social. Por este medio puede aprenderse algo tan simple como la localización del alimento, o tan complejo como una secuencia de actos que permita al sujeto que aprende obtener un premio. De esta forma, mediante la observación de aquellos individuos que iniciaran el nuevo comportamiento, debe haberse realizado la transmisión de nuevas costumbres alimenticias en el petirrojo (Erithacus rubecula), el carbonero (Parus major), el herrerillo (Parus caeruleus), y el macaco de Japón (Macaca fuscata) (Kawai, 1963; Lefebvre, 1995), y parecen estar dotados para esta modalidad de aprendizaje muy distintas especies, desde invertebrados a primates (Fiorito y Scotto, 1992; Byrne, 1995; Fritz y Kotrschal, 1999; Schaik et al., 1999). Aunque el aprendizaje por observación parecería precisar de procesos más complejos que los necesarios para el condicionamiento asociativo, el sujeto observador podría más bien estar asociando los estímulos procedentes del comportamiento del sujeto observado con las consecuencias de ese comportamiento, 11 y los resultados de experimentos de laboratorio así lo sugieren. De esta forma, en un solo ensayo, un macaco rhesus (Macaca mulatta) puede aprender la solución a un problema de discriminación planteado a otro mono, en que, de dos objetos presentados, tan sólo uno de ellos va asociado a un refuerzo (por ejemplo, alimento). En este caso, el mono observador no estará imitando al observado, sino aprendiendo de la consecuencia (apropiada o equivocada) del comportamiento de éste. Aprendizaje Latente. En esta forma de aprendizaje existe un lapso de tiempo entre el momento en que el efecto tiene lugar y aquél en que el que se manifiesta en el sujeto. Este sería el caso para las dos últimas formas descritas de aprendizaje, así como para el canto en el macho de muchos paseriformes, cuya expresión completa ocurre cuando el individuo es adulto, aunque la estimulación por el canto de su progenitor tuvo lugar en los primeros meses de vida (Thorpe, 1958). En un ejemplo de Aprendizaje Latente, a una rata saciada se le ofrece la posibilidad de explorar un laberinto, con alimento en la meta. Al alcanzar el fin del laberinto, la rata no comerá el alimento; pero, sometida más tarde, y ya hambrienta, al mismo problema, elegirá más correctamente la ruta adecuada del laberinto (Blodgett, 1929). Cognición. Mediante procesos mentales no directamente observables ciertos animales serían capaces de abstraer, de considerar por separado ciertas cualidades de los objetos a partir de los datos sensoriales, que serían elaborados, almacenados, y, en su momento, recuperados para el uso. Por ejemplo, el chimpancé (Pan troglodytes), cuando se le enseña a formar frases, puede usar símbolos abstractos para colores, sin que esos símbolos (que pueden ser figuras de plástico), tengan el color que el sujeto desee expresar (Premack y Premack, 1983). Parecen darse distintos procesos cognitivos en diferentes especies animales, respondiendo a adaptaciones a diferentes nichos ecológicos y a la propia capacidad mental. Así, aunque la paloma (Columba livia) es capaz de utilizar conceptos de tipo absoluto (como distinguir a una persona sola o en grupo, vestida o desnuda) y de tipo relacional (como discriminar igual a/diferente de) (Wasserman et al., 1995; Cook et al., 12 1997), esta última capacidad es más propia de los primates (Macaco Rhesus, Macaca mulata, y chimpancés) y cetáceos (al menos el delfín mular, Tursiops truncatus) (Köhler, 1921; Wright et al., 1983; Thompson et al., 1997; Herman, 2002). La Etología Cognitiva se ha basado directamente en los logros de la Psicología y utiliza su metodología, aunque complementada con la observación de los animales en su medio natural, sirviéndose además del enfoque ecológico y del sociobiológico. Áreas de interés en este campo son la Atención, Categorización, Conciencia, Emoción, Herramientas, Lenguaje y Memoria (Fig. 3). Figura 3.- El chimpancé se reconoce a sí mismo ante un espejo. El mayor rechazo a la Etología Cognitiva procede de los seguidores del conductismo radical, ya que en gran parte tampoco aceptan el análisis cognitivo del comportamiento humano. Por lo demás, las mayores controversias han surgido en lo referente al uso del lenguaje y la capacidad de conciencia propia. 13 Acción de los estímulos La acción de los estímulos se ha entendido, en su acepción clásica, encuadrada en las hipótesis de Mecanismo Desencadenante Innato, Suma Heterogénea de Estímulos, Estímulos Gestalt o Configuracionales y Estímulos Supernormales. Mecanismo Desencadenante Innato De acuerdo con este modelo, la especificidad con que un determinado estimulo-signo desencadena una pauta de conducta fija y concreta es el resultado de la acción de mecanismos nerviosos que detectarían la presencia de un estímulo determinado y activarían la respuesta apropiada. A este mecanismo se le denominó “Mecanismo Desencadenante Innato” (angeborener Auslösemechanismus, Lorenz, 1937,1950), y es claramente derivado del “Esquema Desencadenante Innato” de von Uexküll. Ejemplos frecuentemente aducidos en apoyo del modelo han sido el ataque por el pez espinoso (Gasterosteus aculeatus) a modelos con vientre rojo (Tinbergen, 1951), por petirrojos a plumas rojas (Lack, 1943) o por el pechiazul hacia plumas azules (Peiponen, 1960), la evitación del precipicio visual en sujetos no experimentados de pollitos, gatos, cabritos y niños (Gibson y Walk, 1960), y el picoteo preferente por pollitos de gallina a fotografías de esferas iluminadas en su parte superior, es decir, de apariencia tridimensional (Dawkins, 1968). Quizá el ejemplo que mejor ilustra la idea de “Estímulos-signo” es el propuesto por von Uexküll (1909) sobre el comportamiento de la garrapata (Ixodes sp.). Este parásito no responde a la visión del hospedador para adherirse a él, sino que, posado sobre una rama, se deja caer sobre el mamífero que pase bajo él al sentirse estimulado por los efluvios de ácido butírico producidos por el hospedador. Una vez 14 sobre él, la garrapata busca una zona de piel desnuda de pelaje, y sólo se desencadenará en la respuesta de horadar la piel y bombear sangre si sus receptores sensoriales son estimulados por una temperatura cercana a los 37ºC. Así pues, sólo ciertos estímulos desencadenan respuestas concretas. Suma Heterogénea de Estímulos Cuando varios estímulos-signo equivalentes (en cuanto desencadenantes de la misma conducta, aunque con distinta efectividad) simultanean su acción (por ejemplo, formando parte del mismo objeto o animal), sus efectos se suman. Este fenómeno, descrito por primera vez por Alfred Seitz, discípulo de Lorenz, en 1940, como ley de “Suma Heterogénea de Estímulos” (“Reizsummenregel”), propone la adición de efectos de los distintos estímulos, de forma que cada elemento añade su propia porción estimulante. Así, el vuelo de cortejo del macho de la mariposa Hypparchia semele es desencadenado de manera óptima por la combinación de estímulos de tipo de movimiento (aleteo), grado de pigmentación (oscura) y distancia al sujeto (cercanía), de forma que al eliminar alguno de estos estímulos se reduce la intensidad de la respuesta de cortejo (Tinbergen et al., 1942). La más clara verificación cuantitativa de la ley la realizaron Baerends y Kruijt (1973) mediante la observación de la respuesta de recuperación de modelos de huevo colocados en el borde del nido por la Gaviota Argéntea (Larus argentatus). Combinando estímulos de forma y color en los modelos experimentales de huevos, se comprobó que cada estímulo añade su propia contribución, que es independiente de la contribución de los otros estímulos. Estímulos-Gestalt o Configuracionales Este tipo de estímulos no cumple la ley de suma heterogénea, sino que todos sus componentes actúan conjuntamente. En consonancia con la visión de Psicología Gestalt de que un objeto estimulador es más que la suma de sus partes, a este tipo de 15 estímulos se les suele denominar también estímulos-Gestalt. Así pues, la estructura del objeto estimulador sería de relación configuracional. El fenómeno se pone de manifiesto en experimentos relativos a la orientación del comportamiento hacia la zona de la cabeza del animal portador del estímulo en el ataque defensivo del avetoro (Botaurus stellaris) y en la petición de alimento por pollos de mirlo (Turdus merula) y zorzal común (Turdus philomelos). Cuando el avetoro se ve acorralado dirige sus rápidos picotazos hacia la cabeza de su enemigo. Mas cuando, experimentalmente, se le presenta un modelo artificial de enemigo sin “cabeza”, no se desencadena la reacción defensiva, y cuando se le añade aquélla (incluso en la forma de un rudimentario modelo de cartón sobre el “cuerpo”), el ave lanza sus ataques a esa zona añadida (Portielje, 1926). En el caso de la petición de alimento por pollos de mirlo y zorzal, la orientación de los picos abiertos se realiza hacia la cabeza del progenitor que llega al nido dispuesto a cebar. La experimentación demostró que los pollos reconocen la cabeza mediante relaciones espaciales entre las partes del cuerpo del progenitor: para el pollo el tamaño absoluto de la cabeza es irrelevante, y lo que importa es la relación de tamaños entre la “cabeza” (la parte saliente superior) y el “cuerpo” (la parte más voluminosa en la parte inferior del estímulo) (Fig. 4; Tinbergen y Kuenen, 1939). Figura 4.- Como ejemplo de la acción de “Estímulos Configuracionales”, presentados dos círculos de tamaño fijo en la parte superior de otro círculo de tamaño variable, los pollos de mirlo y zorzal común elegirán dirigir su petición de alimento hacia uno u otro de los círculos superiores, dependiendo de la relación espacial (de superficie) que tengan con el círculo inferior (Tinbergen y Kuenen, 1939) 16 Estímulos supernormales Con frecuencia, algunas de las características de las señales pueden, al aumentar su intensidad, incrementar el efecto desencadenante sobre las pautas fijas correspondientes. De esta manera, exagerando artificialmente la característica desencadenadora (y produciendo así un “Estímulo Supernormal”), puede éste estímulo ser más efectivo desencadenando la respuesta que el estímulo natural (Fig. 5). Figura 5.- Elección de modelo artificial de huevo de tamaño supernormal por la gaviota argéntea (Según Baerends, 1962). Este fenómeno se puso de manifiesto en la preferencia de varias especies de aves por recuperar o incubar huevos anormalmente grandes (Lorenz y Tinbergen, 1938, en el ganso común; Tinbergen, 1951, en el ostrero Haematopus ostralegus). Este efecto debe haber intervenido en la evolución del parasitismo reproductivo por 17 parte de cuco (Cuculus canorus) y el críalo (Clamator glandarius) produciendo huevos mayores y de color más contrastado que los de sus hospedadores habituales, lo que desencadena una mayor aceptación por éstos (Alvarez et al., 1976; Alvarez, 1999, 2000, 2004). En la misma categoría debería también encuadrarse la preferencia de pollos de gaviota argéntea por picotear (desencadenando así la ceba) puntos coloreados en el pico del progenitor, o modelos de pico anormalmente finos y de color intensamente contrastado (Tinbergen, 1949; Tinbergen y Perdeck, 1950). El efecto superóptimo fue también demostrado por Magnus (1958) en la estimulación del cortejo desencadenado por la alternancia de color anaranjado y oscuro que se da durante el vuelo de la hembra de la mariposa nacarada (Argynnis paphia). Al proceder experimentalmente, Magnus demostró que, situados los dos colores anternantes en un cilindro en rotación, el efecto de desencadenar el cortejo es mayor a mayor velocidad de rotación del cilindro, superándose incluso la situación natural, y sólo deja de actuar la alternancia de colores cuando el aparato sensorial del macho es incapaz de distinguir separadamente los colores, al fusionarse cuando el cilindro gira a muy alta velocidad. Los “Estímulos Supernormales“ no precisan de aprendizaje previo para tener efecto y, en general, su elección (al menos dentro de los rangos de variación normales del estímulo) es ventajosa para el sujeto en cuestión, en cuanto le produce beneficios de supervivencia o reproducción. Así, cuando el macho del pez espinoso corteja a hembras gordas con preferencia a las delgadas, está eligiendo a aquéllas con más huevos en su vientre. Vistos conjuntamente los modelos clásicos de acción de los estímulos, con la excepción del modelo de “Mecanismo Desencadenante Innato”, todos los demás mantienen su vigencia. Aunque el modelo de “Mecanismo Desencadenante Innato” es muy sugerente, y ha animado la investigación por un tiempo, hoy se ha desechado, ya que no se han hallado procesos nerviosos que funcionen en la forma que lo haría el susodicho mecanismo. 18 Energía de Acción Específica Según este modelo, la motivación (que aumentaría con el paso del tiempo) es específica para cada tipo de comportamiento (como la alimentación, lucha, comportamiento sexual, etc.). La fuente de esa energía motivacional sería la “Energía de Acción Específica”. Según el modelo de “Energía de Acción Específica”, las “Pautas Fijas de Conducta” se asociarían durante la ejecución en grupos a un más alto nivel, siendo cada uno de esos grupos activado en el sistema nervioso central por un factor interno concreto (el “Mecanismo Desencadenante Innato”), el que entraría en acción mediante la energía acumulada de adscripción específica a ese factor y por el efecto desencadenante específico de los “Estímulos-signo”. Las analogías que más influyeron en la difusión de este concepto fueron el Modelo Psicohidráulico de Lorenz (1937, 1950) y el Modelo Jerárquico de Tinbergen (1951), así como la idea de las Actividades de Desplazamiento. Modelo Psicohidráulico de Lorenz Según el primero de estos modelos, que Lorenz (1950) veía más como una cruda aproximación a la realidad, tras la realización de una cierta agrupación de “Pautas Fijas de Conducta”, y de la descarga de la energía a su servicio (la “Energía de Acción Específica”), con el paso del tiempo ésta se iría acumulando de nuevo como lo haría el gas bombeado en un recipiente (1937) o el agua en una cisterna (1950) (Fig. 6). Como resultado de la presión del agua acumulada sobre una válvula de salida (cuya apertura, o ejecución de las “Pautas de Acción Fijas”, estaría controlada por el “Mecanismo Desencadenante Innato”), y del peso que actuara abriendo esa válvula (los “Estímulos-signo” específicos que desencadenan la respuesta concreta), la válvula finalmente cedería, liberando el agua hacia otro depósito (la agrupación de actos) con 19 orificios a diferentes niveles (las “Pautas de Acción Fijas” o elementos del grupo de actos). Figura 6.- “Energía de Acción Específica”. A) Modelo Psicohidráulico de Lorenz (1950) y B) Modelo Jerárquico de Tinbergen (1951). En A se libera la acumulación de “Energía de Acción Específica” (mediante la realización de un acto o grupo determinado de actos) por efecto de la presión de la energía acumulada y del “Estímulo-signo” específico para cada acto o grupo de actos. En B se representa la estructura jerarquizada del “centro instintivo” reproductivo del pez espinoso. Dicho “centro reproductivo” lo constituirían varios centros de nivel de complejidad inferior (se representa sólo el “centro territorial”), compuestos a su vez por centros de menor nivel, hasta alcanzar el nivel de los actos concretos, por ejemplo, el de morder al rival. Cada bloqueo, impedimento al flujo descendente de la “Energía de Acción Específica”, se eliminarían mediante la acción de los “Estímulos-signo” y “Mecanismo Desencadenante Innato” correspondientes, liberándose la realización de los actos. 20 La “Energía de Acción Específica” podría acumularse a tan alto nivel que, de no aparecer el “Estímulo-signo” correspondiente, la “Pauta de Acción Fija” se ejecutaría sin más. Se trataría de una “Actividad en Vacío” (“Leerlaufreaktion”; Lorenz, 1939). Lorenz no introdujo en su modelo ningún proceso de retroalimentación que permitiera, mediante el aprendizaje, ajustar el comportamiento futuro, pues para él las “Pautas de Acción Fijas” no cambiarían a lo largo de la vida del animal. En todo caso, aunque el modelo fue en parte aceptado durante el periodo de la Etología clásica, hoy ha caído completamente en desuso, al no hallarse ninguna estructura cerebral que sirva para el acúmulo de la “Energía de Acción Específica”, ni haberse hallado evidencia fisiológica de la existencia del “Mecanismo Desencadenante Innato”. Modelo Jerárquico de Tinbergen El Modelo Jerárquico de Tinbergen (1951) propone la existencia de centros instintivos organizados según complejidad jerarquizada. Por ejemplo, en el tan estudiado pez espinoso existiría, entre otros, un centro del instinto reproductivo, compuesto a su vez de varios instintos de nivel de complejidad inferior. Cada uno de los centros subordinados, por ejemplo, el centro de la territorialidad, se compondría a su vez de centros instintivos más simples: el centro de la nidificación, el agresivo, el de cortejo, etc. (Fig. 6). Según este modelo, existirían bloqueos, impedimentos al flujo descendente de la “Energía de Acción Específica” entre los centros. Mediante la acción de cada “Estímulo-signo” y “Mecanismo Desencadenante Innato” correspondientes, se eliminaría el bloqueo, y, fluyendo la energía, liberaría la realización de los actos. El modelo de Tinbergen presenta la misma debilidad que el de Lorenz, al basarse también en los conceptos de “Energía de Acción Específica” y “Mecanismo Desencadenante Innato”. 21 Actividades de desplazamiento Este comportamiento fue descrito por primera vez, independientemente, por Tinbergen y A. Kortlandt en 1940. La característica principal de esta conducta es su aparente irrelevancia respecto al contexto en que se da, ocurriendo con frecuencia en situaciones de conflicto entre realizar al mismo tiempo dos comportamientos incompatibles. Además, los propios actos del desplazamiento suelen ser actividades que se repiten mucho en la rutina ordinaria del animal (tales como alimentarse, o rascarse). Así, en medio del cortejo, el macho del pez espinoso puede repentinamente nadar hacia su nido y hacer los movimientos de ventilación hacia unos huevos inexistentes, pues aun no han sido puestos por la hembra, sin que este comportamiento influya en ninguna forma sobre la hembra cortejada. Además del contexto sexual, el agresivo es un origen frecuente de conflicto, y de las consiguientes actividades de desplazamiento. En el caso del espinoso, la ventilación-desplazamiento suele darse ante el conflicto entre cortejar a la hembra y atacarla como intrusa en el territorio, o, en una verdadera disputa territorial, puede hacer movimientos de excavar el nido (Tinbergen y van Iersel 1947). También, durante una disputa territorial o pelea, los escribanos nivales (Plectrophenax nivalis), alondras cornudas (Eremophila alpestris) o los gallos (Gallus g. domesticus) implicados muestran conflicto entre atacar o apartarse del contrario, mostrando entonces con frecuencia la actividad de desplazamiento de picotear el suelo, como si se alimentaran (Howard 1929, Pickwell 1931, Lorenz 1935, Tinbergen 1939). Las actividades de desplazamiento pueden ser elementos del repertorio ya mencionado de la atención o construcción del nido, o la falsa alimentación, así como conducta sexual (Tinbergen 1952), incubación (Kortlandt 1940), rascarse las plumas o pelaje (Portielje 1939, Tinbergen 1940), o incluso dormir (Makkink 1936). No obstante, estas actividades pueden darse fuera de situaciones de conflicto, por ejemplo, cuando se impide alcanzar el resultado del comportamiento que el animal 22 esté realizando. Por ejemplo, si, en el intento de obtener alimento, se impide el acceso del animal a ese alimento, o bien se retrasa su obtención, o si el alimento no tiene el gusto habitual. El oportunismo evolutivo ha integrado algunas de estas actividades al catálogo de señales comunicativas, fijando su estructura mediante un proceso de ritualización y haciéndolas menos variables, con lo que se disminuye su ambigüedad y se incrementa su eficacia en comunicación. Según Tinbergen (1952), las actividades de desplazamiento serían salidas por las que se expresarían fuertes motivaciones cuya respuesta habría sido de alguna forma impedida. Por otra parte, y al no hallarse evidencia para aquella idea, Andrew (1956) y van Iersel y Bol (1958) sugieren que los sistemas de comportamiento habitualmente ejercen inhibición sobre conductas de otros sistemas (de otra forma no podría ejecutarse ningún comportamiento), y al entrar en conflicto varios sistemas, la inhibición se anularía temporalmente, apareciendo la conducta que de otra forma no se realizaría. La mera existencia de las actividades de desplazamiento reduce la fiabilidad de modelos motivacionales basados en los conceptos de “Energía de Acción Específica” y “Mecanismo Desencadenante Innato”, pues esos modelos no pueden explicar como una agrupación de factores causales pueden producir un comportamiento “normalmente ” dependiente de otra agrupación (Hinde, 1856). Modelos de relaciones temporales Los modelos más refinados de la Etología clásica en la búsqueda de factores causales comunes para los distintos elementos de la conducta se han basado en las relaciones temporales entre actos. Dos aproximaciones al problema han proporcionado una visión global de la conducta: las secuencias temporales de actos y los modelos factoriales. 23 Secuencias temporales de actos En base principalmente a la observación del comportamiento de anátidas, Oskar Heinroth (1871-1945) propuso la existencia de secuencias de actos de comportamiento, fijas, características de cada especie y heredadas sin intervención de procesos de aprendizaje (Heinroth, 1911). Las primeras aproximaciones al problema recurrieron al simple cómputo de transiciones entre actos (Fig. 7). Este método ya proporciona un esquema de la estructura temporal del comportamiento de cortejo de los peces Lebistes reticulatus (Baerends et al. 1955), Pygosteus pungitius (Morris, 1958) y Tilapia mosambica (Neil, 1964), aunque su utilidad es limitada. Figura 7.- Secuencia de actos del cortejo del macho territorial del pez Tilapia mossambica hacia hembras a más y menos 15 cm del nido (Según Neil, 1964). Una aproximación probabilística a las transiciones proporciona una visión más objetiva, tal como se obtuvo para la agresión y el cortejo del págalo grande (Stercorarius skua, Andersson 1974) y del babuino gelada Theropithecus gelada 24 (Alvarez y Cónsul, 1978). Por otra parte, el modelo de cadenas de Markov ha resultado exitoso en el análisis de la conducta de dípteros, peces, aves y primates (Nelson, 1964; Altmann, 1965; Lemon y Chatfield, 1971; Dawkins y Dawkins, 1976; Dobson y Lemon, 1979). La técnica de cadenas de Markov debe usarse con precaución, debido sobre todo a que la probabilidad de los diferentes actos tiende a variar en el tiempo. Así pues, la obtención de secuencias puede ser una útil herramienta de descripción, aunque no convendría ir más allá tratando de obtener conclusiones sobre factores causales a partir de esa información. Además, muchos de los estudios arriba citados se realizaron sobre sujetos en situación social, asumiéndose que el comportamiento de los otros animales apenas influía sobre la secuencia de conductas. Sin embargo, el análisis de secuencias cruzadas ha demostrado el efecto comunicativo del comportamiento de unos animales sobre el de otros (Altmann, 1965; Hazlett y Bossert, 1965; Alvarez y Cónsul, 1978). Modelos factoriales Otra aproximación a las relaciones temporales es la obtención de agrupaciones de actos mediante análisis de agrupaciones (clusters), que presenta la ventaja de precisar escasas presunciones sobre la estructura del comportamiento (Morgan et al., 1976; Ghett 1978). El método más sofisticado hacia la comprensión de la estructura temporal del comportamiento es quizá el análisis factorial (Hazlett, 1977; Frey y Pimentel, 1978). Este método, que permite detectar un pequeño número de factores causales, de los que dependerían los actos (y no de ellos entre sí, como asumen las cadenas de Markov), y que explicarían la mayor parte de las correlaciones observadas entre los actos (Blalock, 1961; Slater, 1973), ha sido aplicado con éxito a especies diferentes de peces, anfibios, ardeidos, cérvidos y primates (Wiepkema, 1961; Baerends y van der Cincel, 1962; Baerends et al., 1970; van Hooff, 1970; Balthazart, 1973; Halliday, 1976; Alvarez et al., 1975; Alvarez y Cónsul, 1978; López de Ipiña et al., 1983). 25 El estudio pionero de Wiepkema (1961) sobre el análisis factorial de las relaciones temporales entre doce actos del cortejo del macho del pez ródeo (Rhodeus amarus) demostró la existencia de tres factores independientes (representables, por tanto, en un modelo tridimensional, Fig. 8), que responden al 80-90% de la varianza total: el grupo de actos agonísticos (compuesto por las tendencias a agredir y a huir), el grupo sexual, y el grupo no reproductivo. El análisis factorial del comportamiento total del babuino gelada (Alvarez y Cónsul, 1978) demostró que los 22 actos más frecuentes se organizan temporalmente según cuatro factores, que absorben al 65% de la varianza total: I. Autoestimulación, II. Limpieza social del pelaje versus Relación social intensa, III. Sexual masculino, y IV. Ataque versus Presentación sexual. Figura 8.- Modelo vectorial del comportamiento de cortejo del macho del pez ródeo (Rhodeus amarus), obtenido mediante análisis factorial de los coeficientes de correlación de las transiciones temporales entre los actos (1 al 12), representados como vectores respecto a los tres factores comunes obtenidos (I, II y III), los que se representan en forma ortogonal (según Wiepkema, 1961). La elección de la técnica de cadenas de Markov o de análisis factorial dependerá sobre todo de la importancia que a priori se atribuya a los potenciales “efectos secuenciales” o a los “efectos motivacionales”. Para los primeros se asume un estado motivacional constante, lo que es poco realista, mientras que para los 26 segundos la asunción es de un cambio más lento en los factores motivacionales que en los propios actos. Por otra parte, aunque el análisis factorial pretende explicar el comportamiento mediante unos pocas variables fundamentales, no es claro lo que esas variables verdaderamente significan, de modo que mientras para Wiepkema (1961) esos factores representarían ”tendencias” motivacionales, para Baerends (1970) serían tan sólo “áreas más densas de la red causal”, admitiéndose por tanto la dependencia del comportamiento tanto de factores externos como internos. Resumen Se revisan los siguientes modelos causales sobre comportamiento animal producidos a lo largo el siglo XX en los campos de la Etología y la Psicología Comparada. 1. Comportamiento apetitivo y consumatorio. 2. Umwelt. 3. Modelos de aprendizaje Aprendizaje no asociativo (Habituación, Sensibilización). Aprendizaje asociativo (Condicionamiento Clásico, Condicionamiento Operante). Aprendizaje complejo (Impronta Filial, Impronta Sexual, Aprendizaje Observacional, Aprendizaje Latente, Cognición). 4. Acción de los estímulos Mecanismo Desencadenante Innato Suma Heterogénea de Estímulos Estímulos-Gestalt o Configuracionales Estímulos Supernormales 5. Energía de Acción Específica Modelo Psicohidráulico de Lorenz 27 Modelo Jerárquico de Tinbergen Actividades de Desplazamiento 6. Modelos de relaciones temporales Secuencias de actos Modelos factoriales Referencias Altmann, S. A. (1965): «Sociobiology of rhesus monkeys II: Stochastics of social communication». J. Theor. Biol., 8: 490-522. Alvarez, F. (1999): «Attractive non-mimetic stimuli in Cuckoo Cuculus canorus eggs». Ibis, 141: 142-144. Alvarez, F. (2000): «Response to Common Cuckoo Cuculus canorus model egg size by a parasitized population of Rufous Bush Chat Cercotrichas galactotes». Ibis, 142: 683-686. Alvarez, F. (2004): «The conspicuous gape of the nestling Common Cuco Cuculus canorus as a supernormal stimulus for Rufous Bush Chat Cercotrichas galactotes hosts». Ardea, 92: 63-68. Alvarez, F., Arias de Reyna, L., y Segura, M. (1976): «Experimental brood parasitism of the magpie (Pica pica) ». Anim. Behav., 24: 907-916. Alvarez, F., Braza, F., y Norzagaray, A. (1975): «Etograma cuantificado del gamo (Dama dama) en libertad». Doñana, Acta Vert., 2: 93-142. Alvarez, F., y Cónsul, C. (1978): «The structure of social behaviour in Theropithecus gelada». Primates, 19: 45-59. Andersson, M. (1974): «Temporal graphical analysis of behaviour sequences». Behaviour, 51: 38-48. Andrew, R. J. (1956): «Normal and irrelevant toilet behaviour in Emberiza, Spp.». Brit. J. Anim. Behaviour, 4: 85-96. Baerends, G. P. (1962): «La reconnaissance de l’oeuf par le goeland argenté». Bull. 28 Soc. Scientifique Bretagne, 37: 193-208. Baerends, G. P. (1970): «A model of the functional organization of incubation behaviour». Behaviour Suppl., 17: 263-312. Baerends, G. P., Brouwer, R., y Waterbolk, H. Tj. (1955): «Ethological studies on Lebistes reticulatus (Peters): I. An analysis of the male courtship pattern». Behaviour, 8: 249-334. Baerends, G. P., y van der Cingel, N. A. (1962): «On the phylogenetic origin of the snap display in the coomon heron (Ardea cinerea L.)». Symp. Zool. Soc. Lond., 8: 7-24. Baerends, G. P., Drent, R. H., Glas, P., y Groenewold, H. (1970): «An ethological analysis of incubation behaviour in the herring gull». Behaviour Suppl., 17: 135235. Baerends, G. P., y Kruijt, J. P. (1973): «Stimulus selection». En: Hinde, R. A., y Stevenson-Hinde, J. (Eds.), Constraints on learning (pp. 23-49). Londres, Academic Press. Balthazart, J. (1973): «Analyse factorielle du comportement agonistique chez Tilapia macrochir (Boulenger 1912)». Behaviour, 46: 37-72. Blalock, H. M. (1961): Causal inferences in Nonexperimental Research. Chapel Hill, University of North Carolina Press. Blodgett, H. C. (1929): «The effect of the introduction of reward upon the maze performance of rats». Univ. of California Publ. Psychology, 4: 113-134. Byrne, R. W. (1995): The Thinking Ape: Evolutionary Origins of Intelligence. Oxford, Oxford Univ. Press. Cook, R. G., Katz, J. S., y Cavoto, B. R. (1997): «Pigeon same-different concept learning with multiple stimulus classes». J. Experimental Psychology: Animal Behavior Processes, 23: 417-433. Craig, W. (1918): «Appetites and Aversions as Constituents of Instincts». Biol. Bull., 34: 91-107. 29 Darwin, C. (1872): The Expression of Emotions in Man and Animals. Londres, Murray. Darwin, C. (1877): «A biographical sketch of an infant ». Mind, 2: 285-294. Darwin, C. (1877): «A biographical sketch of an infant». Mind, 2: 285-294. Dawkins, R. (1968): «The ontogeny of a pecking preference in domestic chicks». Z. Tierpsychol., 25:170-186. Dawkins, R., y Dawkins, M. (1976): «Hierarchical organization and postural facilitation: rules for grooming in flies». Anim. Behav., 24: 739-755. Dewey, J. (1927-1928): «Body and Mind». En: The Collected Works of John Dewey: Later Works, Vol. 3, 1927-1928, Essays (pp. 25-40). Chicago, SIU Press. Dobson, C. W., y Lemon, R. E. (1979): «Markov sequences in songs of American thrushes». Behaviour, 68: 86-104. Emmeche, C. (2001): «Does a robot have an Umwelt? Reflections on the qualitative biosemiotics of Jakob von Uexküll». Semiotica, 134: 653-693. Evans, S. M. (1971): «Behavior in Polychaetes». Quarterly Review of Biology, 46: 379405. Fiorito, G., y Scotto, P. (1992): «Observational learning in Octopus vulgaris». Science, 256: 545-547. Frey, D. F., y Pimentel, R. A. (1978): «Principal Component Analysis and Factor Analysis». En: Colgan, P. W. (Ed.), Quantitative Ethology (pp. 219-245). Nueva York, Wiley. Fritz, J., y Kotrschal, K. (1999): «Social learning in common ravens». Anim. Behav., 57: 785-793. Ghett, V. J. de (1978): «Hierarchical Cluster Analysis». En: Colgan, P. W. (Ed.), Quantitative Ethology (pp. 115-144). Nueva York, Wiley. Gibson, E.J., y Walk, R. D. (1960): «The visual cliff». Scient. Amer., 202: 64-71. Halliday, T. R. (1976): «The libidinous newt: an analysis of variations in the sexual behaviour of the male smooth newt, Triturus vulgaris». Anim. Behav., 24: 398414. 30 Hartley, D. (1749). Observations on Man, his Frame, his Duty, and his Expectations. Nueva Cork, Delmar, 1976. Hazlett, B. A. (1977): Quantitative Methods in the Study of Animal Behavior. Loncres, Academic Press. Hazlett, B. A., y Bossert, W. H. (1965): «A statistical analysis of the aggressive communications systems of some hermit crabs». Anim. Behav., 13: 357-373. Heinroth, 0. (1910): Beiträge zur Biologie, namentlich Ethologie und Psychologie der Anatiden. Verh. 5 int. orn. Kongr. Berlin: 589-702. Heinroth, O., y Heinroth, M. (1924-1928): Die vogel Mitteleuropas. I-IV. Bermühler Verlag, Berlín. Herman, L. M. (2002): «Exploring the cognitive world of the bottlenosed dolphin». En: Bekoff, M., Allen, C. y Burghardt, G., (Eds.), The cognitive animal: Empirical and theoretical perspectives on animal cognition (pp. 275-283). Cambridge, MA, MIT Press, . Hinde, R. A. (19566): «Ethological Models and the Concept of “Drive”». British J. Philosophy of science, 6: 321-331. Hochner, B., Shomrat, T., y Fiorito, G. (2006): «The Octopus: A Model for a Comparative Analysis of the Evolution of Learning and Memory Mechanisms». Biol. Bull., 210: 308-317. Howard, H. E. (1929): An introduction to the study of bird behaviour. Cambridge, Cambridge University Press. Iersel, J. van., y Bol, A. C. (1958): «Preening of two tern species. A study on displacement activities». Behaviour, 13: 4-89. Kawai, M. (1963): «Newly-acquired Pre-Cultural Behavior of the Natural Troop of Japanese Monkeys of Koshima Islet». Primates, 21: 141-160. Kortlandt, A. (1940): «Eine Ubersicht der angeborenen Verhaltensweisen des 31 mittleleuropäischen Kormorans (Phalacrocorax carbosinensis), ihre Funktion, ontogenetische Entwicklung und phylogenetische Herkunft ». Arch. Néerl. Zool., 4: 401-442. Köhler, W. (1921): Intelligenzenprüfungen am Menschenaffen. Berlín, Springer. Kuhn, T. S. (1962): The Structure of Scientific Revolutions. Chicago, University of Chicago Press. [Trad. Castellana: La estructura de las revoluciones científicas. México, Fondo de Cultura Económica, 1971.] Lack, D. (1943): The Life of the Robin. Cambridge, Cambridge Univ. Press. Lefebvre, L. (1995): The opening of milk bottles by birds: evidence for accelerating learning rates, but against the wave-of advance model of cultural transmission. Behavioural Processes, 34: 43-54. Lemon, R. E., y Chatfield, C. (1971): «Organization of song in cardinals». Anim. Behav., 19: 1-17. López de Ipiña, S., Braza, F., y Alvarez, F. (1983): «Análisis factorial de las expresiones faciales del babuino sagrado (Papio hamadryas)». Doñana, Acta Vert., 10: 169-175. Lorenz, K. (1935): «Der Kumpan in der Umwelt des Vogels». J. f. Ornithol., 83: 137213, 289-413. Lorenz, K. (1937): «Über die Bildung des Instinktbegriffes». Naturwiss., 25: 289-300, 307-318, 324-331. Lorenz, K. (1939): «Vergleichende Verhaltensforschung». Zoologische Anzeiger. Supplement, 12: 69-102. Lorenz, K. (1950): «The comparative method in studying innate behaviour patterns». Symp. Soc. Exp. Biol., 4: 221-268. Lorenz, K., y Tinbergen, N. (1938): «Taxis und Instinkthandlung in der Einrollbewegung Der Graugans». Z. Tierpsychol. 2: 1-29. Magnus, D. (1858): «Experimentelle Untersuchungen zur Bionomie und Ethologie des 32 Kaisermantels Argynnis paphia L. (Lep. Nymph.). I. Uber optische Auslöser von Angliegereaktionen unf ihre Bedeutung für das Sichfinden der Geschechter». Z. f. Tierpsychol., 15: 397-426. Makkink, G. F. (1936): An attempt at an ethogram of the European Avocet (Recurvirostra avosetta L.) with ethological and psychological remarks. Ardea, 25: 1-62. McConnell, J. V. (1966): «Comparative Physiology: Learning in Invertebrates». Ann. Rev. Physiol., 28: 107-136. McFarland, D. (1987): The Oxford Companion to Animal Behaviour. Oxford, Oxford University Press. Mill, J. (1829): Analysis of the Phenomena of the Human Mind. Londres, Thoemmes Continuum, 2001. Morgan, B. J. T., Simpson, M. J. A., Hanby, J. P., y Hall-Graggs, J. (1976): «Visualizing interaction and sequential data in animal behaviour: theory and application of cluster-analysis methods». Behaviour, 56: 1-43. Morris, D. (1958): «The reproductive behaviour of the ten-spined stickleback (Pygosteus pungitius L.)». Behaviour, Suppl., 6: 1-154. Neil, E. H. (1964): «An analysis of color changes and social behavior of Tilapia mossambica». Univ. California Publ. Zool., 75: 1-58. Nelson, K. (1964): «The temporal pattern of courtship behaviour in the glandulocaudine fishes (Ostariophysi, Characidae)». Behaviour, 24: 90-146. Pávlov, I. P. (1927): Conditioned Reflexes: An Investigation of the Physiological Activity of the Cerebral Cortex. Oxford, Oxford University Press. Peiponen, V. A. (I960): «Verhaltensstudien am Blaukehlchen». Ornis Fenn., 37: 69-83. Pickwell, G. B.(1931): «The prairie horned lark». Trans. Acad. Sci. St. Louis, 27: 1-153. Portielje, A. F.J. (1926): «Zur Ethologie bzw. Psychologie von Botaurus stellaris». Ardea, 15: 1-15. Portielje, A.F.J. (1939): «Triebleden bzw. Intelligente Aeusserungen beim Orang-Utan 33 (Pongo pygmaeus Hoppius) ». Bijdr. Dierk., 27: 61-114. Premack, D., y Premack, A. J. (1983): The Mind of an Ape. Norton, Nueva York. [Trad, castellana: La mente del simio. Madrid, Debate 1984.] Ruiter, L. de (1952): Some experiments on the camouflage of stick caterpillars. Behaviour, 4: 222-232. Schaik, C. P. van, Deaner, R. O., y Merrill, M. Y. (1999): «The conditions for tool use in Primates: implication for the evolution of material culture». J. Human Evol., 36: 719-741. Seitz, A. (1940): «Die Paarbildung bei einigen Cichliden. I. Die Paarbildung bei Astatotilapia strigigena (Pfeffer)». Z. f. Tierpsychol., 4: 40-84. Skinner, B. F. (1938): The behavior of organisms. Nueva York, AppletonCentury. Slater, P. J. B., (1973): «Describing sequences of behavior». En: Bateson, P. P. G., y Klopfer, P. H., (Eds.), Perspectives in Ethology (pp. 131-153). Londres, Plenum Press. Spalding, D.A. (1872): «On Instinct». Nature, 6: 485-486. Spalding, D.A. (1873): «Instinct, with Original observations on Young Animals». Macmillan’s Magazine, 27: 282-293. Thompson, R. K. R., Oden, D. L., y Boysen, S. T. (1997): «Language-naïve chimpanzees (Pan troglodytes) judge relations between relations in a conceptual matching-to-sample task». J. Experimantal Psychology: Animal Behavior Processes, 23: 31-43. Thorndike, E. L. (1901): «Animal intelligence: an experimental study of the associative processes in animals». Psychological Review Monograph Supplement, 2: 1-109. Thorpe, W. H. (1958): «The learning of song patterns by birds, with special reference to the chaffinch, Fringilla coelebs». Ibis, 100: 535-570. Thorpe, W. H. (1963): Learning and Instinct in animals. Cambridge, MA, Harvard 34 University Press. Tinbergen, N. (1939): «On the analysis of social organization among vertebrates, with special reference to birds». Amer. Midl. Nat., 21:210-234. Tinbergen, N. (1940): «Die Uebersprungbewegung». Z. f. Tierpsychol., 4: 1-40. Tinbergen, N. (1949): «Die functie van de rode vlek op de snavel van de zilvermeeuw». Bijdragen tot de Dierkunde, 28: 453-465. Tinbergen, N. (1951): The study of instinct. Oxford, Oxford University Press. [El Estudio del Instinto. México, Siglo XXI, 1969.] Tinbergen, N. (1952): «”Derived” activities; their causation, biological significance, origin, and emancipation during evolution». Quar. Rev. Biol., 27: 1-32. Tinbergen, N., y Iersel, J. van (1947): «”Displacement reactions” in the three-spined stickleback». Behaviour, 1: 56-63. Tinbergen, N., y Kuenen, J. (1939): «Über die auslösenden und die richtunggebenden Reizsituationen der Sperrbewegung von jungen Drosseln (Turdus m. merula und T. e. ericetorum Turton) ». Z. f. Tierpsychol., 3: 37-60. [Trad, castellana: «Las situaciones estímulo desencadenadoras y directoras de la respuesta de abrir el pico en las crías del mirlo común y del zorzal común». En N. Tinbergen, Estudios de Etología 2 (pp. 19-55). Alianza, Madrid, 1979.] Tinbergen, N., Meeuse, B.J.D., Boerema, L.K., y Varossieau, W.W. (1942): «Die Blaz des Samtfalters, Eumenis [= Satyrus] semele (L.)». Z. f. Tierpsychol., 5:182226. Tinbergen, N., y Perdeck, A.C. (1950): «On the stimulus situation releasing the begging response in the newly hatched herring gull chick (Larus argentatus argentatus Pont)». Behaviour, 3: 1-39. Uexküll, J. von (1909): Umwelt und Innenwelt der Tiere. Berlin, Springer. Uexküll, J. von (1934): Streifzüge durch die Umwelten von Tieren und Menschen. Berlín, Springer. Uexküll, T. von, Geigges, W., y Hermann, J. M. (1993): «Endosemiosis». Semiotica, 35 96: 5-51. Wasserman, E. A., Hugart, J. A., y Kirkpatrick-Steger, K. (1995): «Pigeons show samedifferent conceptualisation after training with complex visual stimuli». J. Experimental Psychology: Animal Behavior Processes, 21: 248-252. Watson, J. B. (1913): «Psychology as the behaviorist views it». Psychol. Review, 20: 158-177. Wiepkema, P. R. (1961): «An Ethological Analysis of the Reproductive Behaviour of the Bitterling (Rhodeus amarus Bloch)». Arch. Neerl. Zool., 14: 103-199. Wood, D. C (1988): «Habituation in Stentor: A Response-Dependent Process». J. Neuroscience, 9: 2248-2253 Wright, A. A., Santiago, H. C., Urcuioli, P. J., y Sands, S. F. (1983): «Monkey and pigeon acquisition of same/different concept using pictorial stimuli». En: Commons, P. W., Herrnstein, R. J., y Wagner, A. R., (Eds.), Quantitative Analyses of Behavior (pp. 295-317). Cambridge, MA, Ballinger. Yerkes, R. M. (1912): «The intelligence of earthworms». J.Animal Behavior, 2: 332252.